
Suppressive Effects of Lactobacillus on Depression through Regulating the Gut Microbiota and Metabolites in C57BL/6J Mice Induced by Ampicillin

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lactobacillus Strains and Culture
2.2. Animal Feed
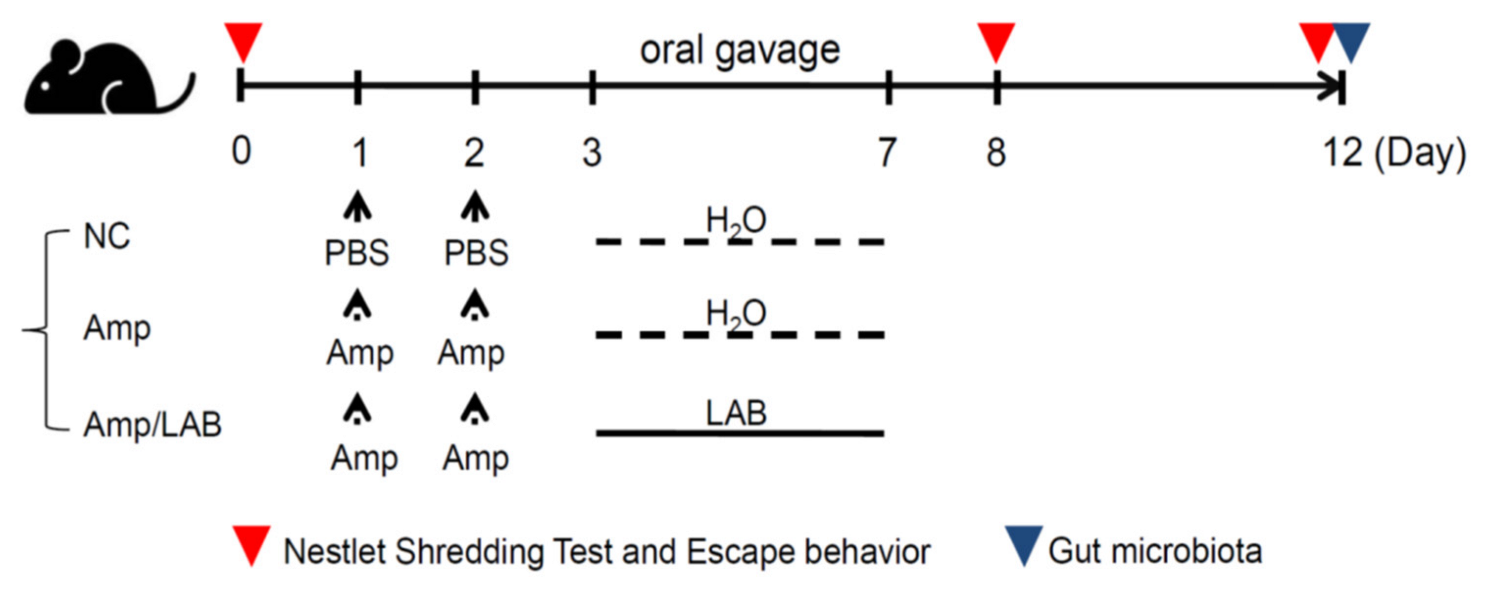
2.3. Animal Treatment-Prescreening and Identification of the Lactobacillus Strain with the Most Potential for Recovering Depressive Behavior
2.4. Behavioral Test for Depression
2.4.1. Nest-Building Test
2.4.2. Forced Swimming Test (FST)
2.5. 16S Ribosomal RNA Gene Amplicon Sequencing
2.6. Bioinformatic Analysis
2.7. Measurement of Short-Chain Fatty Acid (SCFA) Concentration in Mouse Cecal Contents
2.8. Analysis of RNA Expression of Intestinal Tight Junction Protein-Related, Inflammatory Cytokine, and Gut-Derived 5-HT Biosynthesis Genes
2.9. Statistical Analysis
3. Results
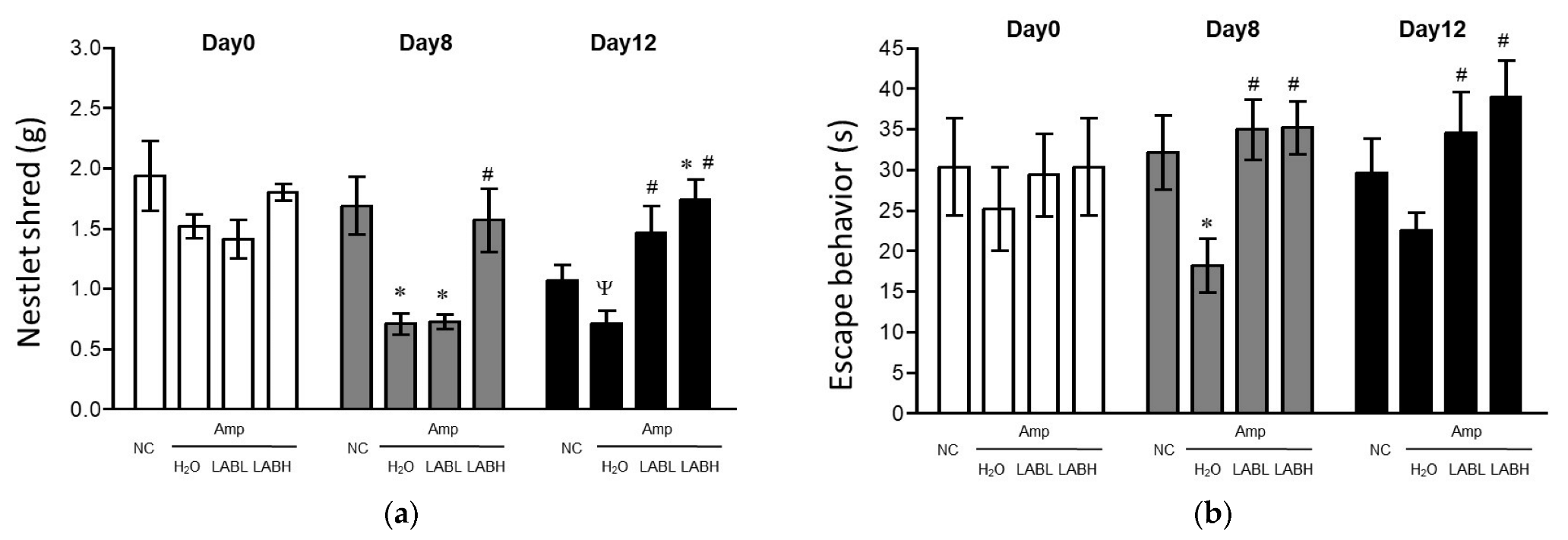
3.1. Composite Lactobacillus spp. Improves Features of Amp-Induced Depression in C57BL/6J Mice
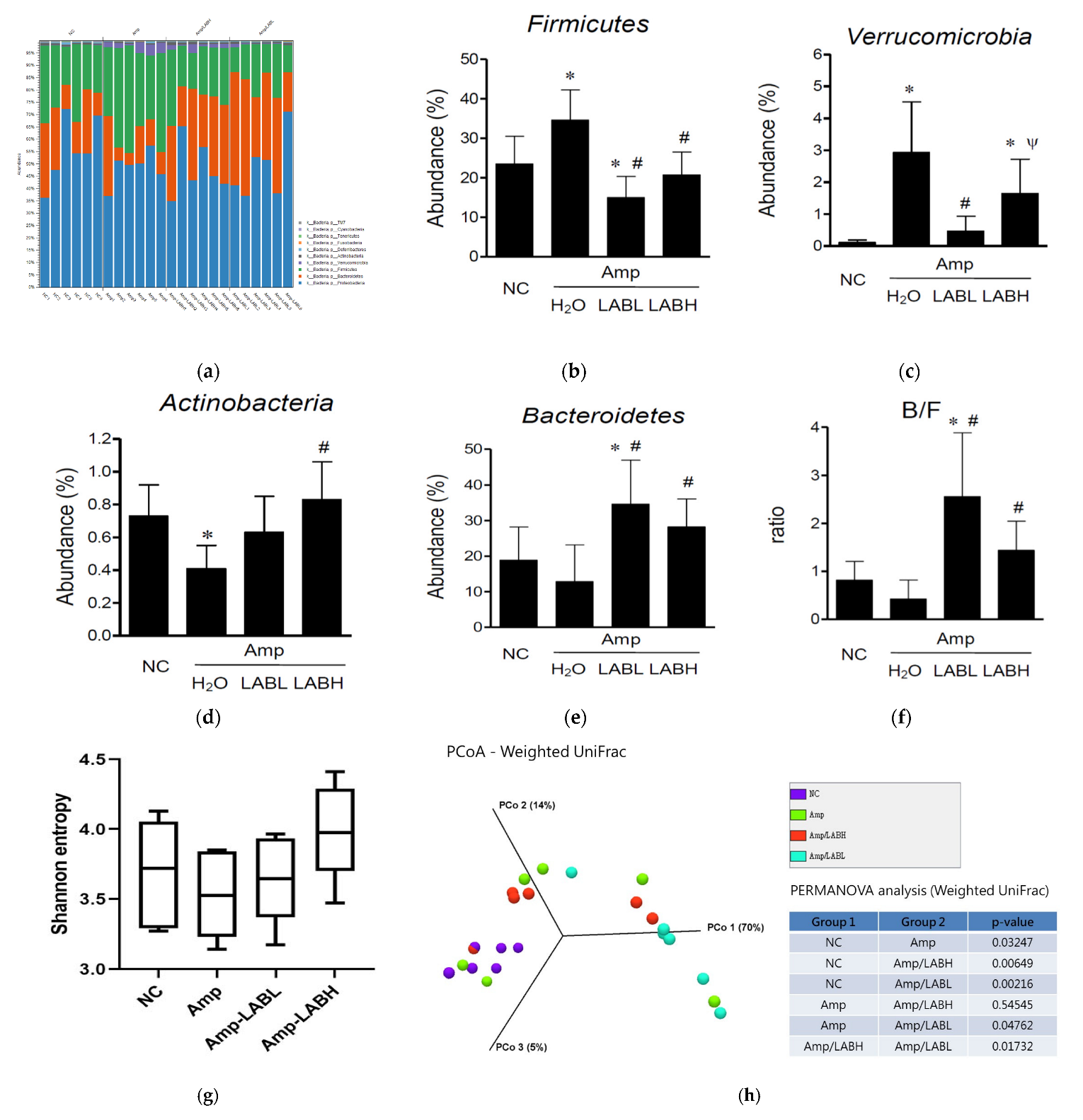
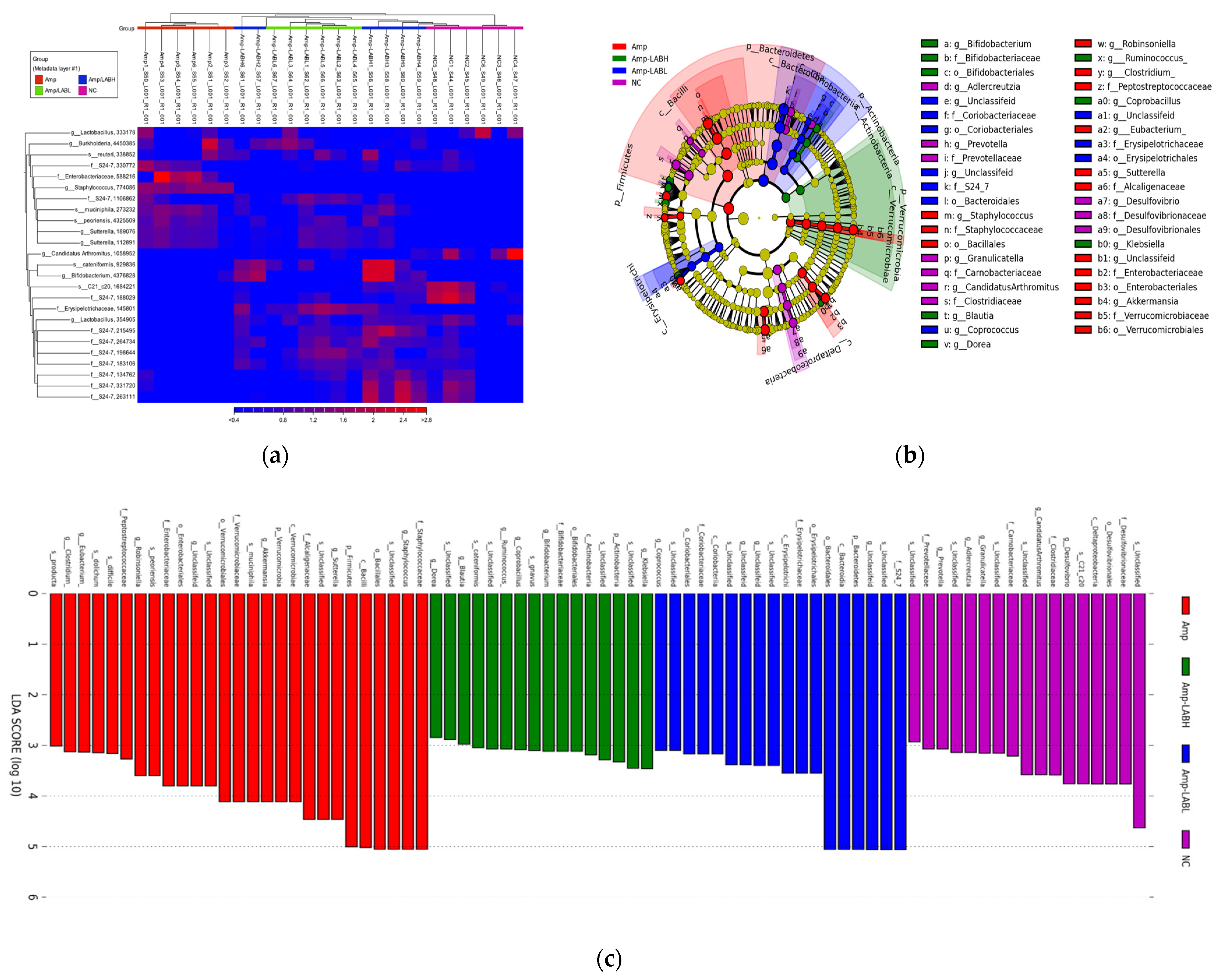
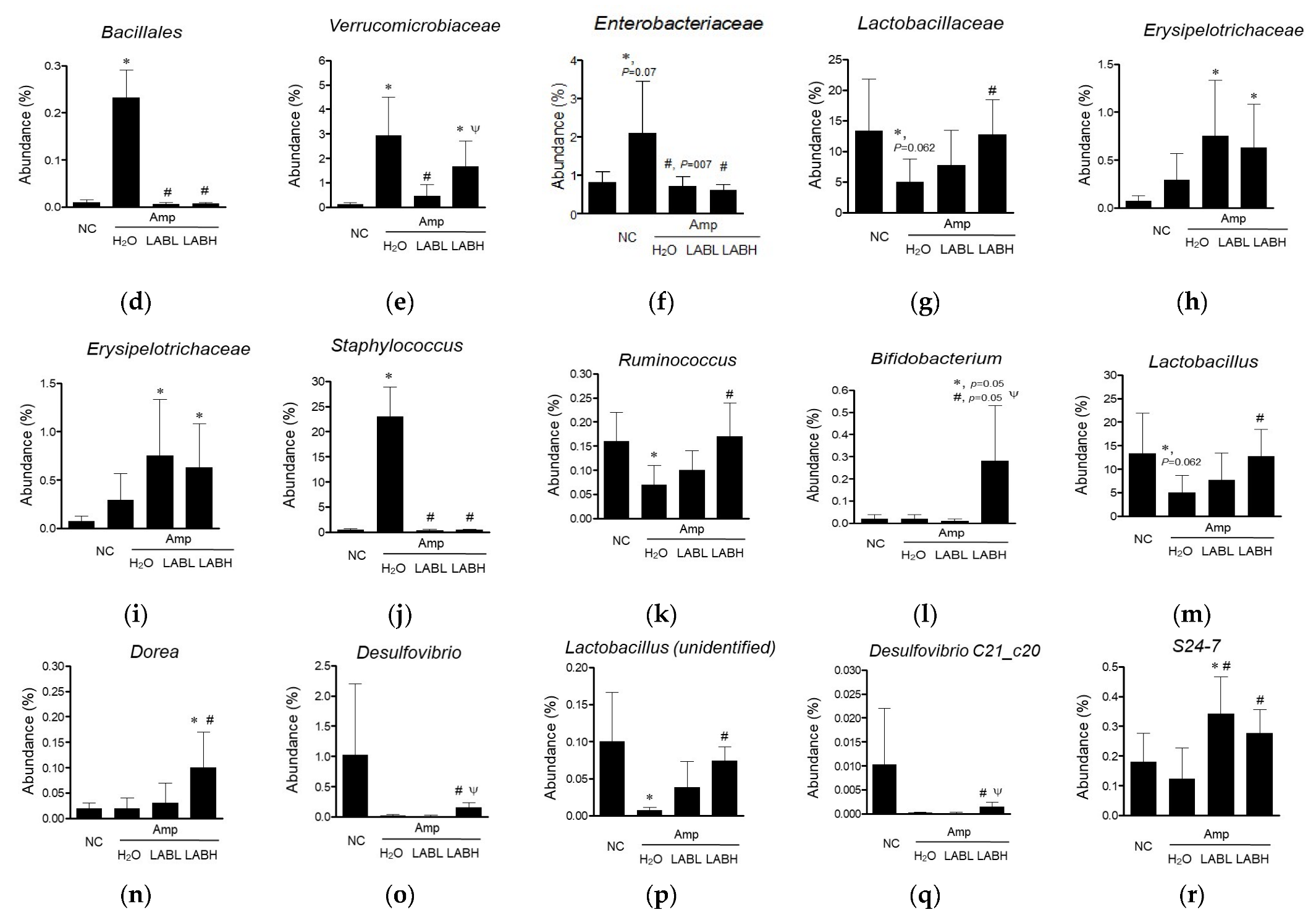
3.2. Composite Lactobacillus spp. Regulates Gut Microbiota Composition in C57BL/6J Mice Treated with Amp
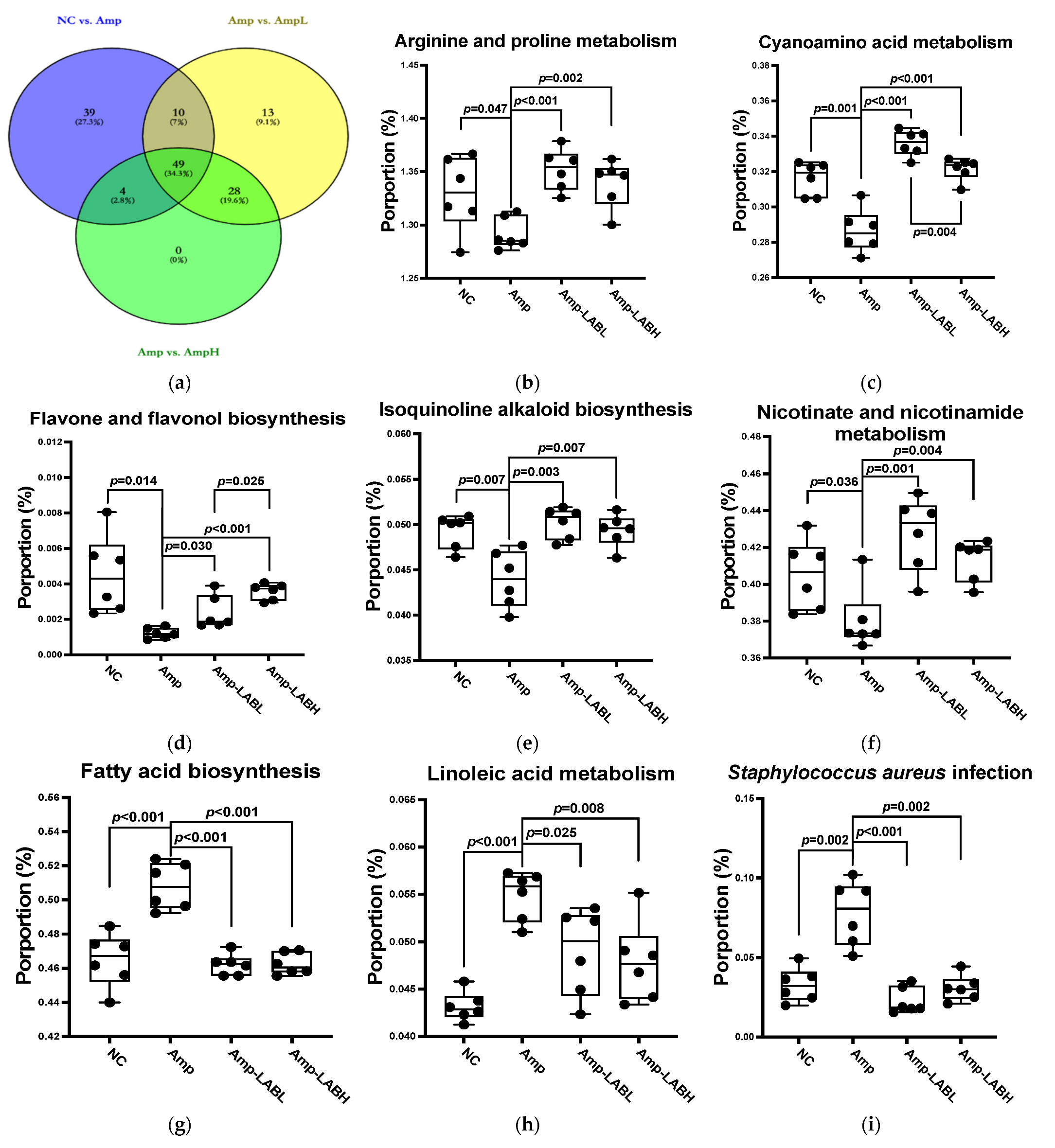
3.3. Composite Lactobacillus spp. Regulates Gut Microbe-Related Pathways in C57BL/6J Mice Treated with Amp
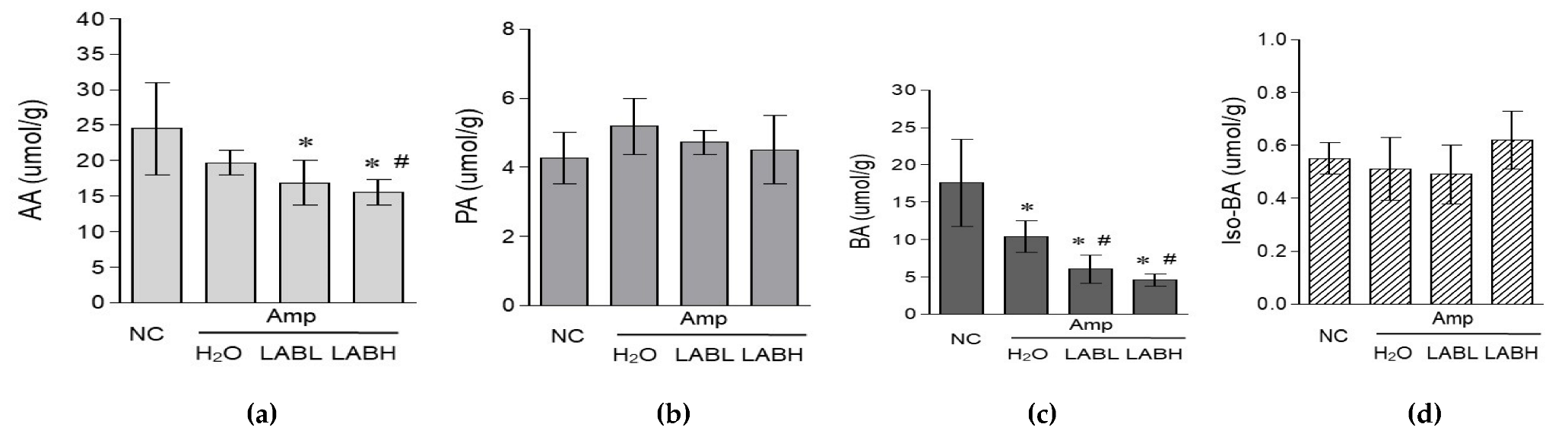
3.4. Composite Lactobacillus spp. Regulates Fecal Fatty Acid Composition in C57BL/6J Mice Treated with Amp
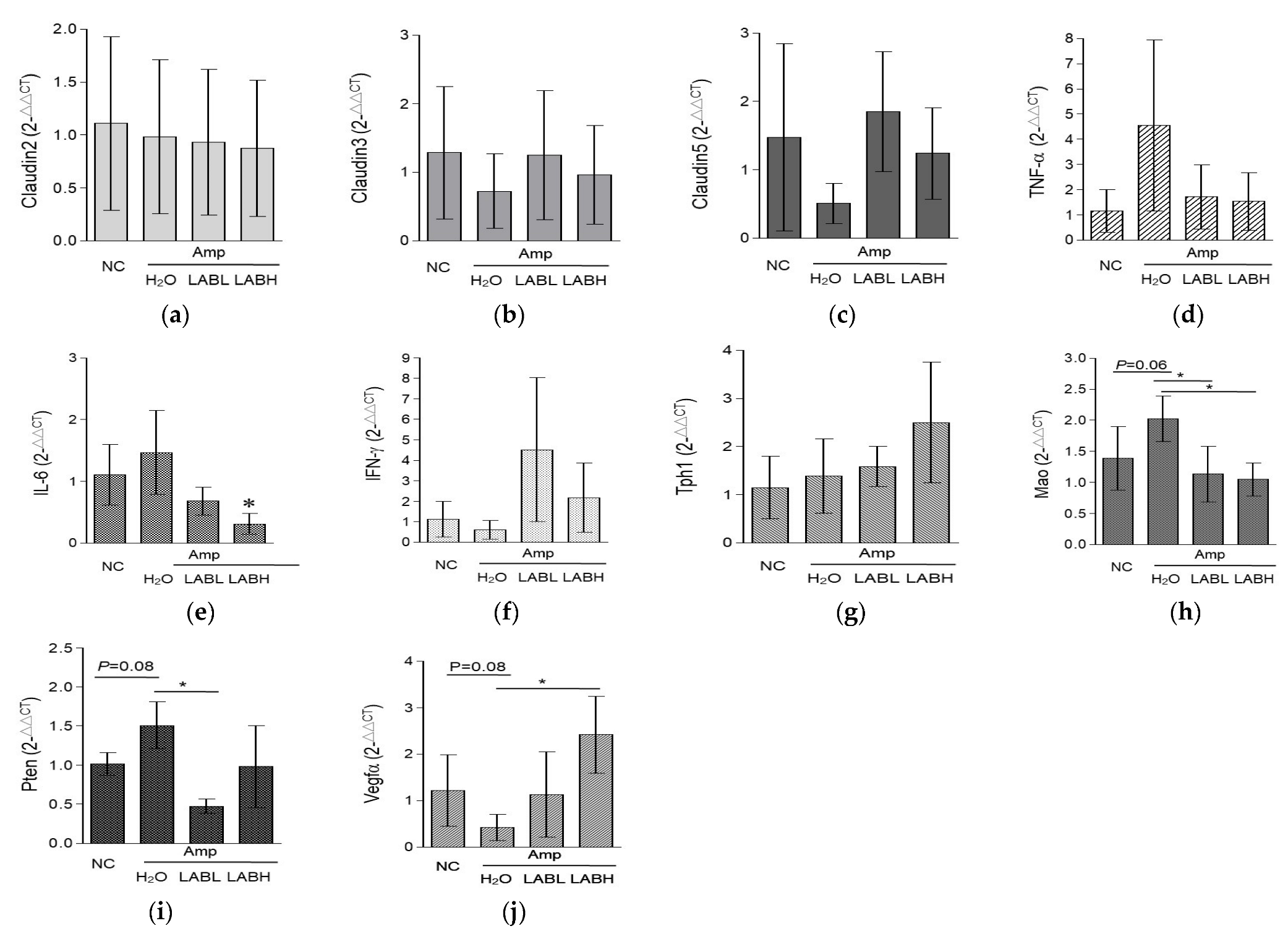
3.5. Composite Lactobacillus spp. Regulates Intestinal Tight Junction, Inflammatory Response Factors, and Gut-Derived 5-HT Biosynthesis Genes in C57BL/6J Mice Treated with Amp
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.A.; Mishra, A.K.; Tantray, J.A.; Alatawi, H.A.; Saeed, M.; Rahman, S.; Jan, A.T. Gut microbiota and cardiovascular system: An intricate balance of health and the diseases state. Life 2022, 12, 1986. [Google Scholar] [CrossRef] [PubMed]
- Paray, B.A.; Albeshr, M.F.; Jan, A.T.; Rather, I.A. Leaky Gut and autoimmunity: An intricate balance of individuals health and the diseased state. Int. J. Mol. Sci. 2020, 21, 9770. [Google Scholar] [CrossRef]
- Khan, S.; Imran, A.; Malik, A.; Chaudhary, A.A.; Rub, A.; Jan, A.T.; Syed, Z.B.; Rolfo, C. Bacterial imbalance and gut pathologies: Association and contribution of E. coli in inflammatory bowel diseases. Crit. Rev. Clin. Lab. Sci. 2018, 56, 1–17. [Google Scholar] [CrossRef]
- Lone, J.B.; Koh, W.N.; Parray, H.A.; Paek, W.K.; Lim, J.H.; Rather, I.; Jan, A.T. Gut microbiome: Microflora association with obesity and obesity related commorbidities. Microb. Pathogen. 2018, 124, 266–271. [Google Scholar] [CrossRef]
- Tap, J.; Mondot, S.; Levenez, F.; Pelletier, E.; Caron, C.; Furet, J.P.; Ugarte, E.; Muñoz-Tamayo, R.; Paslier, D.L.; Nalin, R.; et al. Towards the human intestinal microbiota phylogenetic core. Environ. Microbiol. 2009, 10, 2574–2584. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar]
- Osadchiy, V.; Martin, C.R.; Mayer, E.A. The gut-brain axis and the microbiome: Mechanisms and clinical implications. Clin. Gastroenterol. Hepatol. 2019, 17, 322–332. [Google Scholar] [CrossRef]
- Tang, W.; Zhu, H.; Feng, Y.; Guo, R.; Wan, D. The impact of gut microbiota disorders on the blood-brain barrier. Infect. Drug Resist. 2020, 13, 3351–3363. [Google Scholar] [CrossRef]
- Rutsch, A.; Kantsjö, J.B.; Ronchi, F. The gut-brain axis: How microbiota and host inflammasome influence brain physiology and pathology. Front Immunol. 2020, 11, 604179. [Google Scholar] [CrossRef]
- Tan, F.H.P.; Liu, G.; Lau, S.A.; Jaafar, M.H.; Park, Y.H.; Azzam, G.; Li, Y.; Liong, M.T. Lactobacillus probiotics improved the gut microbiota profile of a Drosophila melanogaster Alzheimer’s disease model and alleviated neurodegeneration in the eye. Benef. Microbes 2020, 11, 79–89. [Google Scholar] [CrossRef]
- Lu, C.S.; Chang, H.C.; Weng, Y.H.; Chen, C.C.; Kuo, Y.S.; Tsai, Y.C. The add-on effect of Lactobacillus plantarum PS128 in patients with Parkinson’s disease: A pilot study. Front. Nutr. 2021, 8, 650053. [Google Scholar] [CrossRef] [PubMed]
- Gu, F.; Wu, Y.; Liu, Y.; Dou, M.; Jiang, Y.; Liang, H. Lactobacillus casei improves depression-like behavior in chronic unpredictable mild stress-induced rats by the BDNF-TrkB signal pathway and the intestinal microbiota. Food Funct. 2020, 11, 6148–6157. [Google Scholar] [CrossRef]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Kiely, B.; Cryan, J.F.; Dinan, T.G. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience 2010, 170, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Bravo, J.A.; Forsythe, P.; Chew, M.V.; Escaravage, E.; Savignac, H.M.; Dinan, T.G.; Bienenstock, J.; Cryan, J.F. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 2011, 108, 16050–16055. [Google Scholar] [CrossRef]
- Institute of Health Metrics and Evaluation. Global Health Data Exchange (GHDx). 2019. Available online: http://ghdx.healthdata.org/gbd-results-tool?params=gbd-api-2019-permalink/d780dffbe8a381b25e1416884959e88b (accessed on 1 May 2021).
- Evans-Lacko, S.; Aguilar-Gaxiola, S.; Al-Hamzawi, A.; Alonso, J.; Benjet, C.; Bruffaerts, R.; Chiu, W.T.; Florescu, S.; de Girolamo, G.; Gureje, O.; et al. Socio-economic variations in the mental health treatment gap for people with anxiety, mood, and substance use disorders: Results from the WHO World Mental Health (WMH) surveys. Psychol. Med. 2018, 48, 1560–1571. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.T.; Lai, J.B.; Du, Y.L.; Xu, Y.; Ruan, L.M.; Hu, S.H. Current understanding of gut microbiota in mood disorders: An update of human studies. Front. Genet. 2019, 10, 98. [Google Scholar] [CrossRef]
- Ma, Q.; Xing, C.; Long, W.; Wang, H.Y.; Liu, Q.; Wang, R.F. Impact of microbiota on central nervous system and neurological diseases: The gut-brain axis. J. Neuroinflamm. 2019, 16, 53. [Google Scholar] [CrossRef]
- Wallace, C.J.K.; Milev, R. The effects of probiotics on depressive symptoms in humans: A systematic review. Ann. Gen. Psychiatry 2017, 16, 14. [Google Scholar] [CrossRef]
- Yun, S.W.; Kim, J.K.; Han, M.J.; Kim, D.H. Lacticaseibacillus paracasei NK112 mitigates Escherichia coli-induced depression and cognitive impairment in mice by regulating IL-6 expression and gut microbiota. Benef. Microbes 2021, 12, 541–551. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, S.; Yang, X.; Li, W.; Si, J.; Yang, X. The antidepressant potential of lactobacillus casei in the postpartum depression rat model mediated by the microbiota-gut-brain axis. Neurosci. Lett. 2022, 774, 136474. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, S.; Zhang, X.; Zhou, H.; Wen, T.; Wang, J. Depression-like symptoms due to Dcf1 deficiency are alleviated by intestinal transplantation of Lactobacillus murine and Lactobacillus reuteri. Biochem. Biophys. Res. Commun. 2022, 593, 137–143. [Google Scholar] [CrossRef]
- Chen, X.; Meng, S.; Yu, Y.; Li, S.; Wu, L.; Zhang, Y. The role of probiotic intervention in regulating gut microbiota, short-chain fatty acids and depression-like behavior in lead-exposed rats. Int. J. Occup. Med. Environ. Health 2022, 35, 95–106. [Google Scholar] [CrossRef]
- Wakade, C.; Chong, R.; Seamon, M.; Purohit, S.; Giri, B.; Morgan, J.C. Low-dose niacin supplementation improves motor function in US veterans with Parkinson’s Disease: A single-center, randomized, placebo-controlled trial. Biomedicines 2021, 9, 1881. [Google Scholar] [CrossRef] [PubMed]
- Matarazzo, I.; Toniato, E.; Robuffo, I. Psychobiome feeding mind: Polyphenolics in depression and anxiety. Curr. Top Med. Chem. 2018, 18, 2108–2115. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.M.; Lee, H.J.; Jang, S.E.; Han, M.J.; Kim, D.H. Evidence for interplay among antibacterial-induced gut microbiota disturbance, neuro-inflammation, and anxiety in mice. Mucosal. Immunol. 2018, 11, 1386–1397. [Google Scholar] [CrossRef] [PubMed]
- Ceylani, T.; Jakubowska-Doğru, E.; Gurbanov, R.; Teker, H.T.; Gozen, A.G. The effects of repeated antibiotic administration to juvenile BALB/c mice on the microbiota status and animal behavior at the adult age. Heliyon 2018, 4, e00644. [Google Scholar] [CrossRef]
- Chen, Y.H.; Tsai, W.H.; Wu, H.Y.; Chen, C.Y.; Yeh, W.L.; Chen, Y.H.; Hsu, H.Y.; Chen, W.W.; Chen, Y.W.; Chang, W.W.; et al. Probiotic Lactobacillus spp. act against Helicobacter pylori-induced inflammation. J. Clin. Med. 2019, 8, 90. [Google Scholar] [CrossRef]
- Bergner, C.L.; Smolinsky, A.N.; Hart, P.C.; Dufour, B.D.; Egan, R.J.; Laporte, J.L.; Kalueff, A.V. Mouse models for studying depression-like states and antidepressant drugs. Methods Mol. Biol. 2010, 602, 267–282. [Google Scholar]
- Jhong, J.H.; Tsai, W.H.; Yang, L.C.; Chou, C.H.; Lee, T.Y.; Yeh, Y.T.; Huang, C.H.; Luo, Y.H. Heat-killed Lacticaseibacillus paracasei GMNL-653 exerts antiosteoporotic effects by restoring the gut microbiota dysbiosis in ovariectomized Mice. Front. Nutr. 2022, 9, 804210. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Niccolai, E.; Baldi, S.; Ricci, F.; Russo, E.; Nannini, G.; Menicatti, M.; Poli, G.; Taddei, A.; Bartolucci, G.; Calabrò, A.S. Evaluation and comparison of short chain fatty acids composition in gut diseases. World J. Gastroenterol. 2019, 25, 5543–5558. [Google Scholar] [CrossRef] [PubMed]
- Wilkowska, A.; Szałach, Ł.P.; Cubała, W.J. Gut microbiota in depression: A focus on ketamine. Front. Behav. Neurosci. 2021, 15, 693362. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol. Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.C.; Yao, W.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Hashimoto, K. Blockade of interleukin-6 receptor in the periphery promotes rapid and sustained antidepressant actions: A possible role of gut-microbiota-brain axis. Transl. Psychiatry 2017, 7, e1138. [Google Scholar] [CrossRef]
- Xie, Y.; Wu, Z.; Zhou, L.; Sun, L.; Xiao, L.; Wang, G. Swimming exercise modulates gut microbiota in CUMS-induced depressed mice. Neuropsychiatr. Dis. Treat. 2022, 18, 749–760. [Google Scholar] [CrossRef]
- Szyszkowicz, J.K.; Wong, A.; Anisman, H.; Merali, Z.; Audet, M.C. Implications of the gut microbiota in vulnerability to the social avoidance effects of chronic social defeat in male mice. Brain Behav. Immun. 2017, 66, 45–55. [Google Scholar] [CrossRef]
- Bai, S.; Bai, H.; Li, D.; Zhong, Q.; Xie, J.; Chen, J.J. Gut microbiota-related inflammation factors as a potential biomarker for diagnosing major depressive disorder. Front. Cell. Infect. Microbiol. 2022, 12, 831186. [Google Scholar] [CrossRef]
- Shen, Y.; Yang, X.; Li, G.; Gao, J.; Liang, Y. The change of gut microbiota in MDD patients under SSRIs treatment. Sci. Rep. 2021, 11, 14918. [Google Scholar] [CrossRef]
- Driscoll, T.; Gabbard, J.L.; Mao, C.; Dalay, O.; Shukla, M.; Freifeld, C.C.; Hoen, A.G.; Brownstein, J.S.; Sobral, B.W. Integration and visualization of host-pathogen data related to infectious diseases. Bioinformatics 2011, 27, 2279–2287. [Google Scholar] [CrossRef] [PubMed]
- Bublitz, D.C.; Wright, P.C.; Bodager, J.R.; Rasambainarivo, F.T.; Bliska, J.B.; Gillespie, T.R. Epidemiology of pathogenic enterobacteria in humans, livestock, and peridomestic rodents in rural Madagascar. PLoS ONE 2014, 9, e101456. [Google Scholar] [CrossRef] [PubMed]
- Slattery, C.; Cotter, P.D.; O’Toole, P.W. Analysis of health benefits conferred by Lactobacillus species from kefir. Nutrients 2019, 11, 1252. [Google Scholar] [CrossRef] [PubMed]
- Drozdov, V.N.; Astapovskiy, A.A.; Serebrova, S.Y.; Lazareva, N.B.; Shikh, E.V. Clinical efficacy of probiotic strains of the Bifidobacterium and Lactobacillus. Vopr. Pitan. 2020, 89, 107–115. [Google Scholar] [PubMed]
- Bonnechère, B.; Amin, N.; van Duijn, C. The role of gut microbiota in neuropsychiatric diseases—Creation of an atlas-based on quantified evidence. Front. Cell. Infect. Microbiol. 2022, 12, 831666. [Google Scholar] [CrossRef] [PubMed]
- Ormerod, K.L.; Wood, D.L.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic characterization of the uncultured Bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef]
- Ramalho, J.B.; Spiazzi, C.C.; Bicca, D.F.; Rodrigues, J.F.; Sehn, C.P.; da Silva, W.P.; Cibin, F.W.S. Beneficial effects of Lactococcus lactis subsp. cremoris LL95 treatment in an LPS-induced depression-like model in mice. Behav. Brain Res. 2022, 426, 113847. [Google Scholar] [CrossRef]
- Yang, C.; Ni, H.Y.; Yin, J.J.; Zhou, T.; Gu, Q.X.; Chen, T.T.; Cai, C.Y. Atorvastatin ameliorates depressive behaviors via regulation of α7nAChR expression by PI3K/Akt-BDNF pathway in mice. Biochem. Biophys. Res. Commun. 2022, 593, 57–64. [Google Scholar] [CrossRef]
- Mayneris-Perxachs, J.; Castells-Nobau, A.; Arnoriaga-Rodríguez, M.; Martin, M.; de la Vega-Correa, L.; Zapata, C.; Burokas, A.; Blasco, G.; Coll, C.; Escrichs, A.; et al. Microbiota alterations in proline metabolism impact depression. Cell Metab. 2022, 34, 681–701. [Google Scholar] [CrossRef]
- German-Ponciano, L.J.; Rosas-Sánchez, G.U.; Rivadeneyra-Domínguez, E.; Rodríguez-Landa, J.F. Advances in the preclinical study of some flavonoids as potential antidepressant agents. Scientifica 2018, 2018, 2963565. [Google Scholar] [CrossRef]
- Khan, H.; Perviz, S.; Sureda, A.; Nabavi, S.M.; Tejada, S. Current standing of plant derived flavonoids as an antidepressant. Food Chem. Toxicol. 2018, 119, 176–188. [Google Scholar] [CrossRef]
- Gasperi, V.; Sibilano, M.; Savini, I.; Catani, M.V. Niacin in the central nervous system: An update of biological aspects and clinical applications. Int. J. Mol. Sci. 2019, 20, 974. [Google Scholar] [CrossRef] [PubMed]
- Elia, I.; Broekaert, D.; Christen, S.; Boon, R.; Radaelli, E.; Orth, M.F.; Verfaillie, C.; Grünewald, T.G.P.; Fendt, S.M. Proline metabolism supports metastasis formation and could be inhibited to selectively target metastasizing cancer cells. Nat. Commun. 2017, 8, 15267. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Liu, L.; Zhang, Y.; Pu, J.; Yang, L.; Zhou, C.; Yuan, S.; Zhang, H.; Xie, P. Metabolomics identifies perturbations in amino acid metabolism in the prefrontal cortex of the learned helplessness rat model of depression. Neuroscience 2017, 343, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Corso, M.; Ziliotto, F.; Rizzini, F.M.; Teo, G.; Cargnello, G.; Bonghi, C. Sensorial, biochemical and molecular changes in Raboso Piave grape berries applying “Double Maturation Raisonnée” and late harvest techniques. Plant Sci. 2013, 208, 50–57. [Google Scholar] [CrossRef]
- Fan, J.; Zhang, K.; Jin, Y.; Li, B.; Gao, S.; Zhu, J.; Cui, R. Pharmacological effects of berberine on mood disorders. J. Cell. Mol. Med. 2019, 23, 21–28. [Google Scholar] [CrossRef]
- Liu, N.; Sun, S.; Wang, P.; Sun, Y.; Hu, Q.; Wang, X. The mechanism of secretion and metabolism of gut-derived 5-Hydroxytryptamine. Int. J. Mol. Sci. 2021, 22, 7931. [Google Scholar] [CrossRef]
- Spohn, S.N.; Mawe, G.M. Non-conventional features of peripheral serotonin signalling—The gut and beyond. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Saraf, M.K.; Piccolo, B.D.; Bowlin, A.K.; Mercer, K.E.; LeRoith, T.; Chintapalli, S.V.; Shankar, K.; Badger, T.M.; Yeruva, L. Formula diet driven microbiota shifts tryptophan metabolism from serotonin to tryptamine in neonatal porcine colon. Microbiome 2017, 5, 77. [Google Scholar] [CrossRef]
- Warner-Schmidt, J.L.; Duman, R.S. VEGF is an essential mediator of the neurogenic and behavioral actions of antidepressants. Proc. Natl. Acad. Sci. USA 2007, 104, 4647–4652. [Google Scholar] [CrossRef]
- Wang, X.Q.; Zhang, L.; Xia, Z.Y.; Chen, J.Y.; Fang, Y.; Ding, Y.Q. PTEN in prefrontal cortex is essential in regulating depression-like behaviors in mice. Transl. Psychiatry 2021, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 2019, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Skonieczna-Żydecka, K.; Grochans, E.; Maciejewska, D.; Szkup, M.; Schneider-Matyka, D.; Jurczak, A.; Łoniewski, I.; Kaczmarczyk, M.; Marlicz, W.; Czerwińska-Rogowska, M. Faecal short chain fatty acids profile is changed in polish depressive women. Nutrients 2018, 10, 1939. [Google Scholar] [CrossRef]
- Wu, M.; Tian, T.; Mao, Q.; Zou, T.; Zhou, C.J.; Xie, J.; Chen, J.J. Associations between disordered gut microbiota and changes of neurotransmitters and short-chain fatty acids in depressed mice. Transl. Psychiatry 2020, 10, 350. [Google Scholar] [CrossRef]
- Lewis, K.; Lutgendorff, F.; Phan, V.; Söderholm, J.D.; Sherman, P.M.; McKay, D.M. Enhanced translocation of bacteria across metabolically stressed epithelia is reduced by butyrate. Inflamm. Bowel. Dis. 2010, 16, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Kayama, H.; Okumura, R.; Takeda, K. Interaction between the microbiota, epithelia, and immune cells in the intestine. Annu. Rev. Immunol. 2020, 38, 23–48. [Google Scholar] [CrossRef]
- Miranda-Ribera, A.; Ennamorati, M.; Serena, G.; Cetinbas, M.; Lan, J.; Sadreyev, R.I.; Jain, N.; Fasano, A.; Fiorentino, M. Exploiting the zonulin mouse model to Eestablish the role of primary impaired gut barrier function on microbiota composition and immune profiles. Front Immunol. 2019, 10, 2233. [Google Scholar] [CrossRef]
- Zhou, J.M.; Zhang, H.J.; Wu, S.G.; Qiu, K.; Fu, Y.; Qi, G.H.; Wang, J. Supplemental xylooligosaccharide modulates intestinal mucosal barrier and cecal microbiota in laying hens fed oxidized fish oil. Front. Microbiol. 2021, 12, 635333. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | Species | Group | Rate of Recovery from Depressive Behaviors |
|---|---|---|---|
| (%, Mean ± SEM) | |||
| 1 | L. rhamnosus; GMNL-74 | A1 # | 107.4 ± 25.9 |
| 2 | L. reuteri | A2 # | 45.2 ± 27.4 |
| 3 | L. fermentum | A3 # | 77.4 ± 19.4 |
| 4 | L. plantarum; GMNL-141 | A4 # | 156.2 ± 46.7 |
| 5 | L. acidophilus; GMNL-185 | A5 # | 132.1 ± 54.0 |
| 6 | L. casei | A6 # | 69.4 ± 14.0 |
| 7 | L. fermentum | A7 # | 5.4 ± 22.1 |
| 8 | L. paracasei | A8 # | 39.2 ± 15.3 |
| 9 | L. rhamnosus | A9 # | 44.6 ± 14.0 |
| 10 | L. plantarum; GMNL-662 | A10 # | 10.8 ± 38.8 |
| 11 | L. rhamnosus + L. plantarum + L. acidophilus | A1 + A4 + A5 # | 171.4 ± 71.8 |
| 12 | L. rhamnosus + L. plantarum + L. acidophilus | A1 + A4 + A5 & | 234.3 ± 64.4 |
| Gene | Primer Sequence (5′–3′) * | Size (bp) | GenBank Accession No. |
|---|---|---|---|
| GADPH (mice) | F-GCACAGTCAAGGCCGAGAAT | 151 | JN958248.1 |
| R-GAATCCTTCTGACCCATGCC | |||
| Claudin2 | F-TCCGGGACTTCTACTCACCA | 190 | XM_021188667.2 |
| R-CTCCTAGTGGCAAGAGGCTG | |||
| Claudin3 | F-GTTTCGGCATTCATCGGCA | 180 | XM_021186977.2 |
| R-TGCCAGTAGGATAGACACCAC | |||
| Claudin5 | F-GCTCTCAGAGTCCGTTGACC | 235 | NM_013805.4 |
| R-CTGCCCTTTCAGGTTAGCAG | |||
| IL-6 | F-TCTCTCCGCAAGAGACTTCCA | 235 | XM_021191538.1 |
| R-ATACTGGTCTGTTGTGGGTGG | |||
| TNFα | F-CTAGCCAGGAGGGAGAACAG | 149 | NM_001278601.1 |
| R-GCTTTCTGTGCTCATGGTGT | |||
| IFNγ | F-TCCTTTGGACCCTCTGACTT | 130 | NM_008337.4 |
| R-GTAACAGCCAGAAACAGCCA | |||
| Tph1 | F-ACTGCGACATCAGCCGAGAA | 162 | XM_036152912.1 |
| R-CGCAGAAGTCCAGGTCAGAAATC | |||
| Mao | F-GGAGAAGCCCAGTATCACAGG | 113 | NM_173740.3 |
| R-GAACCAAGACATTAATTTTGTATTCTGAC | |||
| VEGF-α | F-GCT ACT GCC GTC CGA TTG A | 163 | NM_001025257.3 |
| R-ATG GTG ATG TTG CTC TCT GA | |||
| Pten | F-GGA AGT AAG GAC CAG AGA CAA | 287 | XM_006526769.3 |
| R-CAC CAC ACA CAG GCA ATG |
| Pathway (Significance, p < 0.05) | Amp vs. NC | Amp-LABL vs. Amp | Amp-LABH vs. Amp |
|---|---|---|---|
| Amoebiasis | I * | D | D |
| Arginine and proline metabolism | D | I | I |
| Atrazine degradation | I | D | D |
| Bacterial toxins | D | I | I |
| Biosynthesis of siderophore group nonribosomal peptides | I | D | D |
| Biosynthesis of vancomycin group antibiotics | D | I | I |
| Cellular antigens | D | I | I |
| Chromosome | D | I | I |
| Cyanoamino acid metabolism | D | I | I |
| Cytoskeleton proteins | D | I | I |
| D-Alanine metabolism | I | D | D |
| D-Arginine and D-ornithine metabolism | I | D | D |
| Energy metabolism | D | I | I |
| Fatty acid biosynthesis | I | D | D |
| Flavone and flavonol biosynthesis | D | I | I |
| Glycerolipid metabolism | I | D | D |
| Glycerophospholipid metabolism | I | D | D |
| Isoquinoline alkaloid biosynthesis | D | I | I |
| Linoleic acid metabolism | I | D | D |
| Meiosis–yeast | I | D | D |
| Membrane and intracellular structural molecules | D | I | I |
| Metabolism of cofactors and vitamins | I | D | D |
| Nicotinate and nicotinamide metabolism | D | I | I |
| Nitrogen metabolism | I | D | D |
| Other ion-coupled transporters | I | D | D |
| Penicillin and cephalosporin biosynthesis | D | I | I |
| Phosphonate and phosphinate metabolism | I | D | D |
| Phosphotransferase system (PTS) | I | D | D |
| Plant-pathogen interaction | D | I | I |
| Polyketide sugar unit biosynthesis | D | I | I |
| Pores ion channels | D | I | I |
| Primary immunodeficiency | D | I | I |
| Protein folding and associated processing | I | D | D |
| Protein processing in the endoplasmic reticulum | D | I | I |
| Proximal tubule bicarbonate reclamation | D | I | I |
| Pyruvate metabolism | I | D | D |
| Renal cell carcinoma | I | D | D |
| Riboflavin metabolism | I | D | D |
| Shigellosis | I | D | D |
| Signal transduction mechanisms | I | D | D |
| Staphylococcus aureus infection | I | D | D |
| Streptomycin biosynthesis | D | I | I |
| Sulfur metabolism | I | D | D |
| Sulfur relay system | I | D | D |
| Taurine and hypotaurine metabolism | D | I | I |
| Tetracycline biosynthesis | I | D | D |
| Ubiquitin system | I | D | D |
| alpha-Linolenic acid metabolism | I | D | D |
| beta-Lactam resistance | D | I | I |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, W.-H.; Yeh, W.-L.; Chou, C.-H.; Wu, C.-L.; Lai, C.-H.; Yeh, Y.-T.; Liao, C.-A.; Wu, C.-C. Suppressive Effects of Lactobacillus on Depression through Regulating the Gut Microbiota and Metabolites in C57BL/6J Mice Induced by Ampicillin. Biomedicines 2023, 11, 1068. https://doi.org/10.3390/biomedicines11041068
Tsai W-H, Yeh W-L, Chou C-H, Wu C-L, Lai C-H, Yeh Y-T, Liao C-A, Wu C-C. Suppressive Effects of Lactobacillus on Depression through Regulating the Gut Microbiota and Metabolites in C57BL/6J Mice Induced by Ampicillin. Biomedicines. 2023; 11(4):1068. https://doi.org/10.3390/biomedicines11041068
Chicago/Turabian StyleTsai, Wan-Hua, Wen-Ling Yeh, Chia-Hsuan Chou, Chia-Lin Wu, Chih-Ho Lai, Yao-Tsung Yeh, Chorng-An Liao, and Chih-Chung Wu. 2023. "Suppressive Effects of Lactobacillus on Depression through Regulating the Gut Microbiota and Metabolites in C57BL/6J Mice Induced by Ampicillin" Biomedicines 11, no. 4: 1068. https://doi.org/10.3390/biomedicines11041068
APA StyleTsai, W. -H., Yeh, W. -L., Chou, C. -H., Wu, C. -L., Lai, C. -H., Yeh, Y. -T., Liao, C. -A., & Wu, C. -C. (2023). Suppressive Effects of Lactobacillus on Depression through Regulating the Gut Microbiota and Metabolites in C57BL/6J Mice Induced by Ampicillin. Biomedicines, 11(4), 1068. https://doi.org/10.3390/biomedicines11041068