

Moderate-Intensity Exercise Improves Mesenteric Arterial Function in Male UC Davis Type-2 Diabetes Mellitus (UCD-T2DM) Rats: A Shift in the Relative Importance of Endothelium-Derived Relaxing Factors (EDRF)
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Experimental Animals
2.3. Maximal Oxygen Consumption
2.4. Measurement of Metabolic Parameters in the Plasma
2.5. Blood Pressure Measurement
2.6. Measurement of Mesenteric Arterial Tension
2.6.1. Relaxation Responses to ACh
2.6.2. Relaxation Responses to Sodium Nitroprusside (SNP)
2.6.3. Contractile Responses to PE
2.6.4. Measurement of Myogenic Tone and Wall Thickness
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
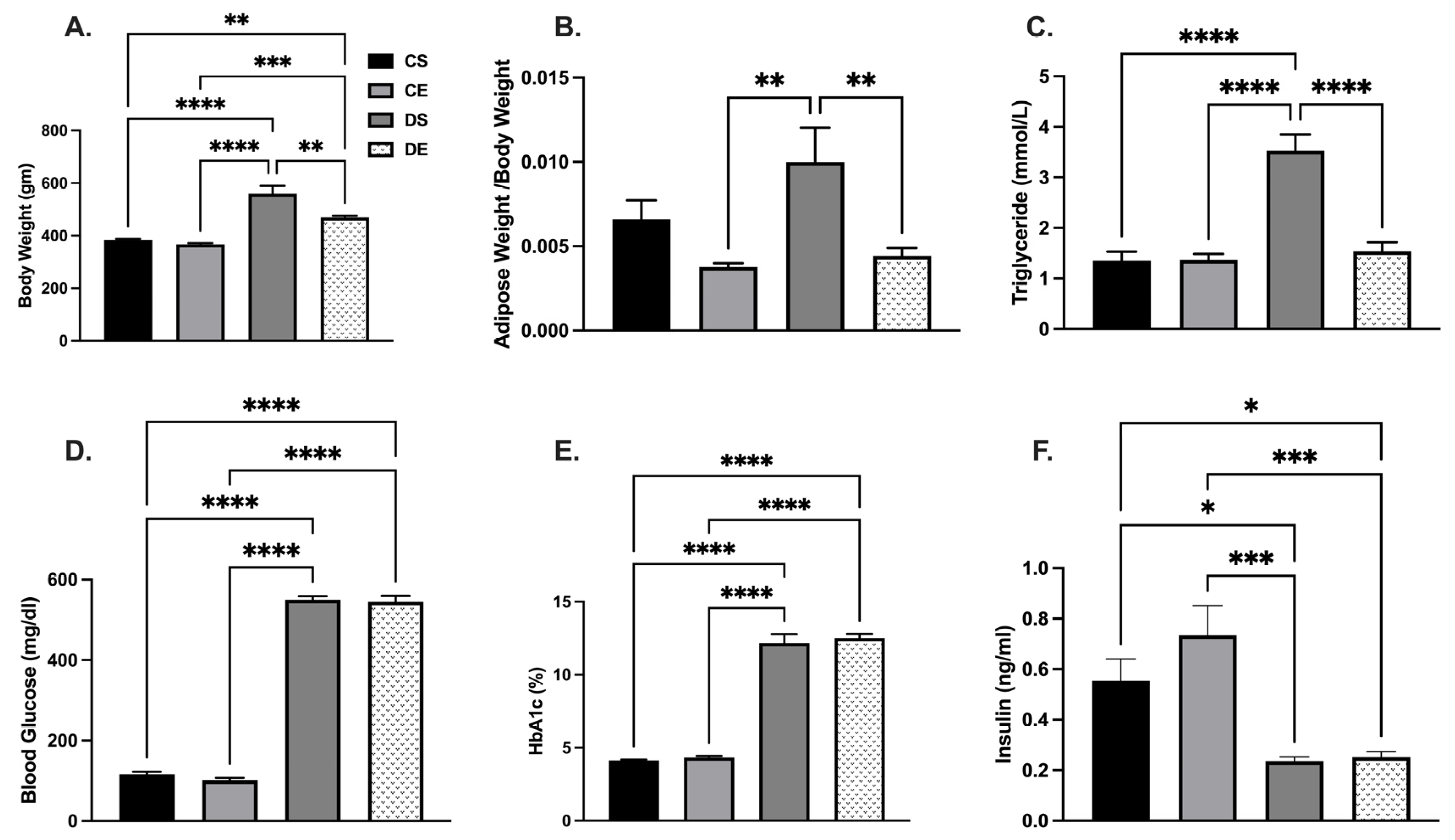
3.1. Effects of MIE on Metabolic Parameters and Blood Pressure
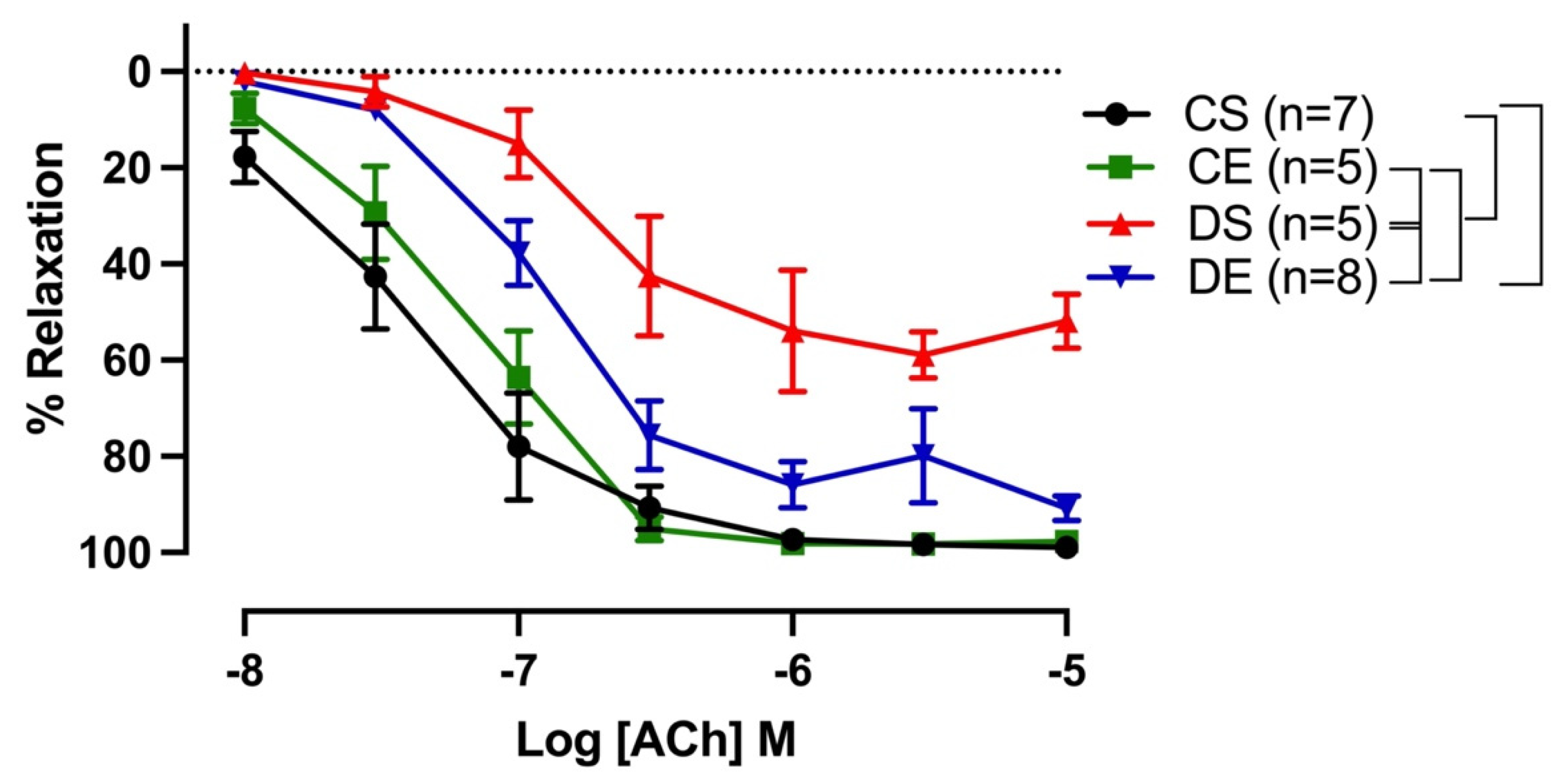
3.2. Effects of MIE on Relaxation Responses to ACh
3.3. Effects of MIE on the Relative Contribution of EDRF
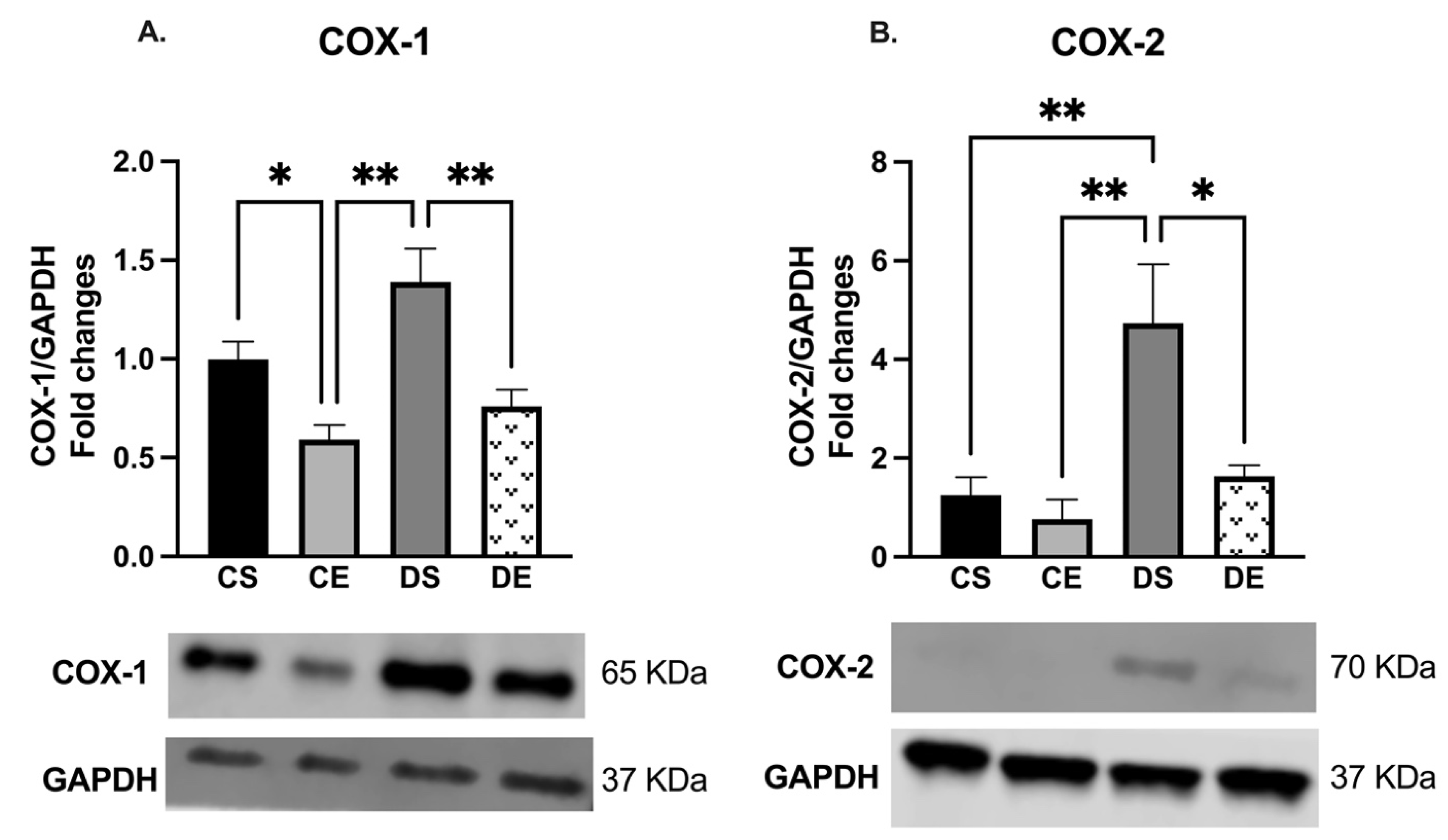
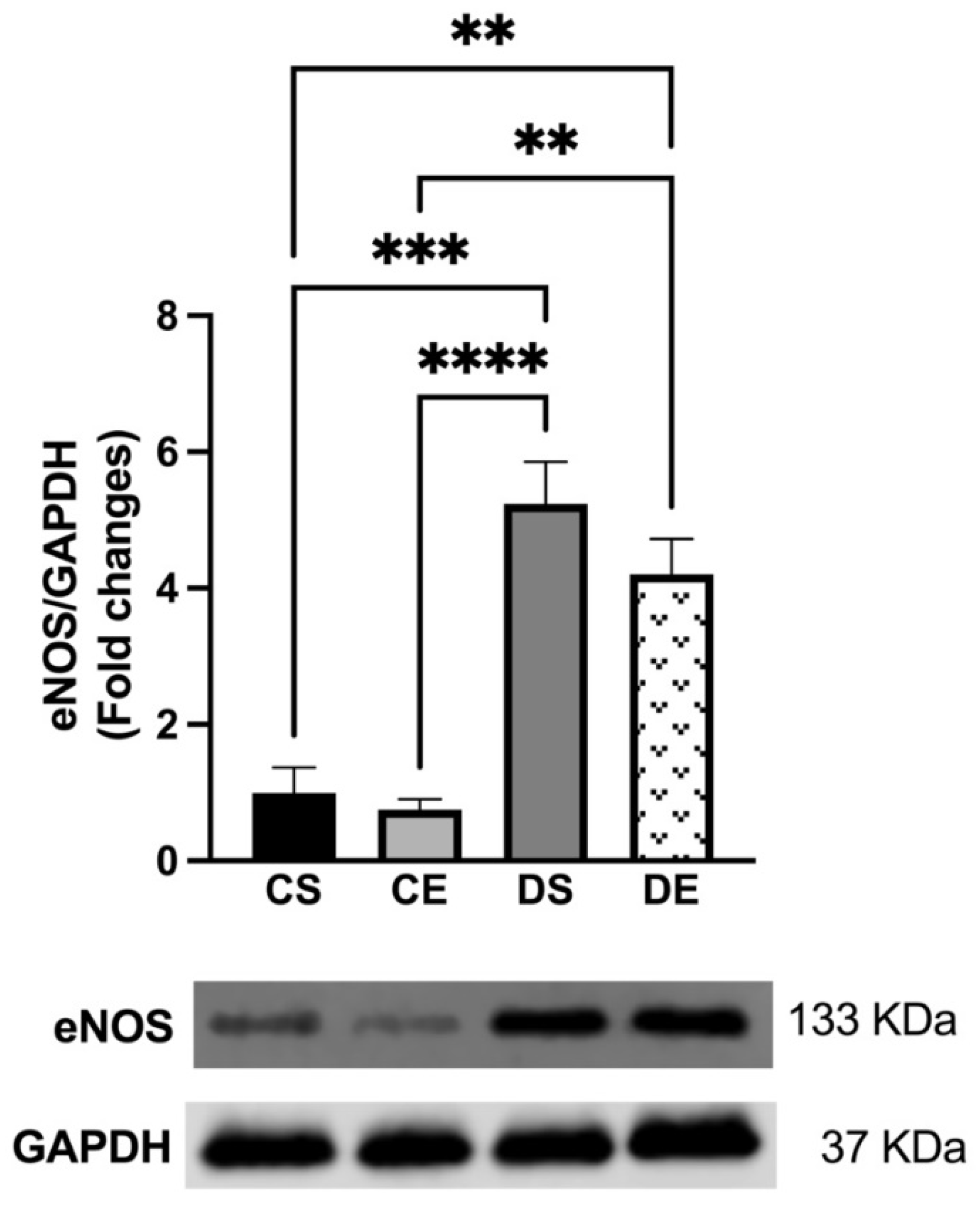
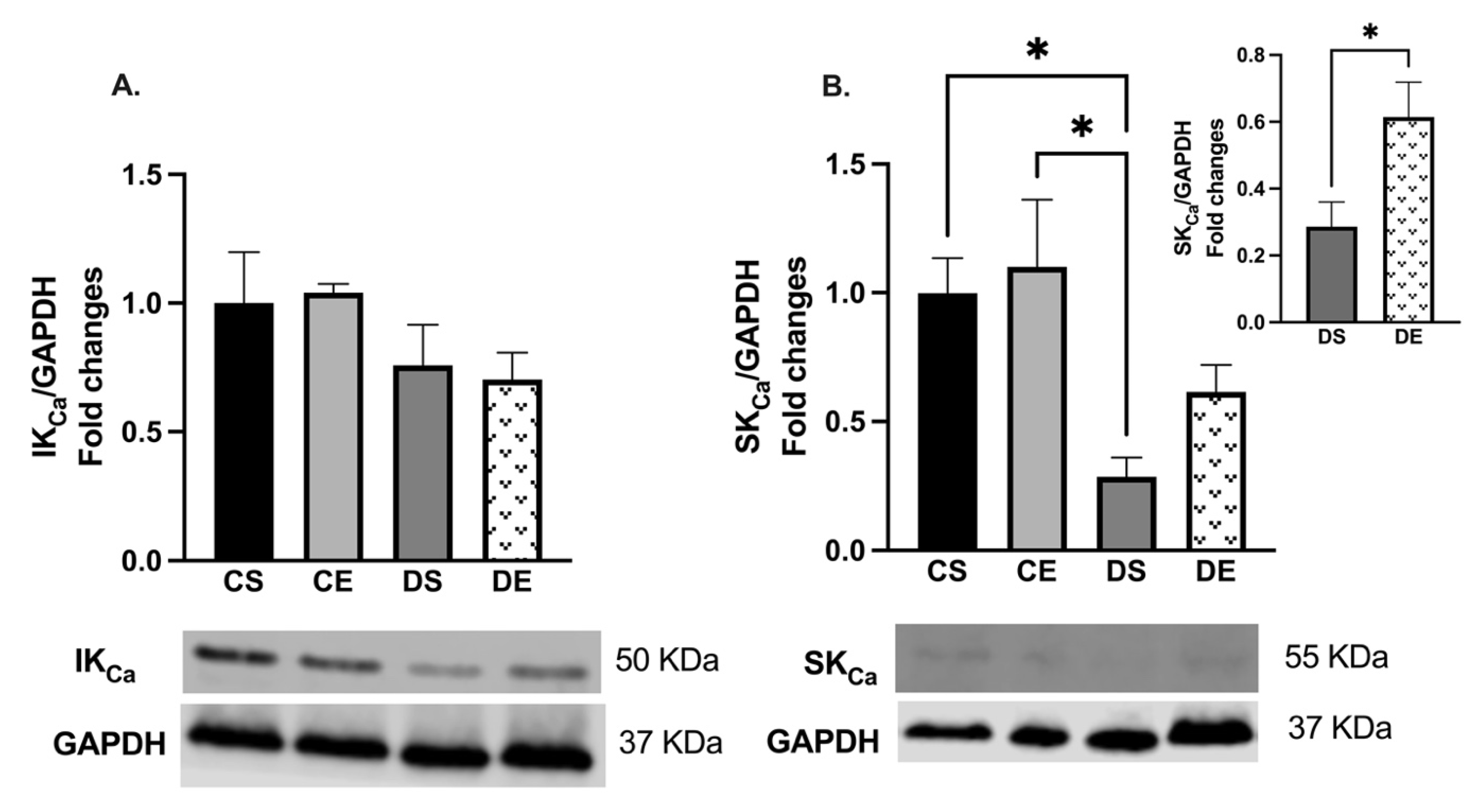
3.4. Effects of MIE on COX, eNOS, and KCa Expression
3.5. Effects of MIE on the SNP-induced Relaxation
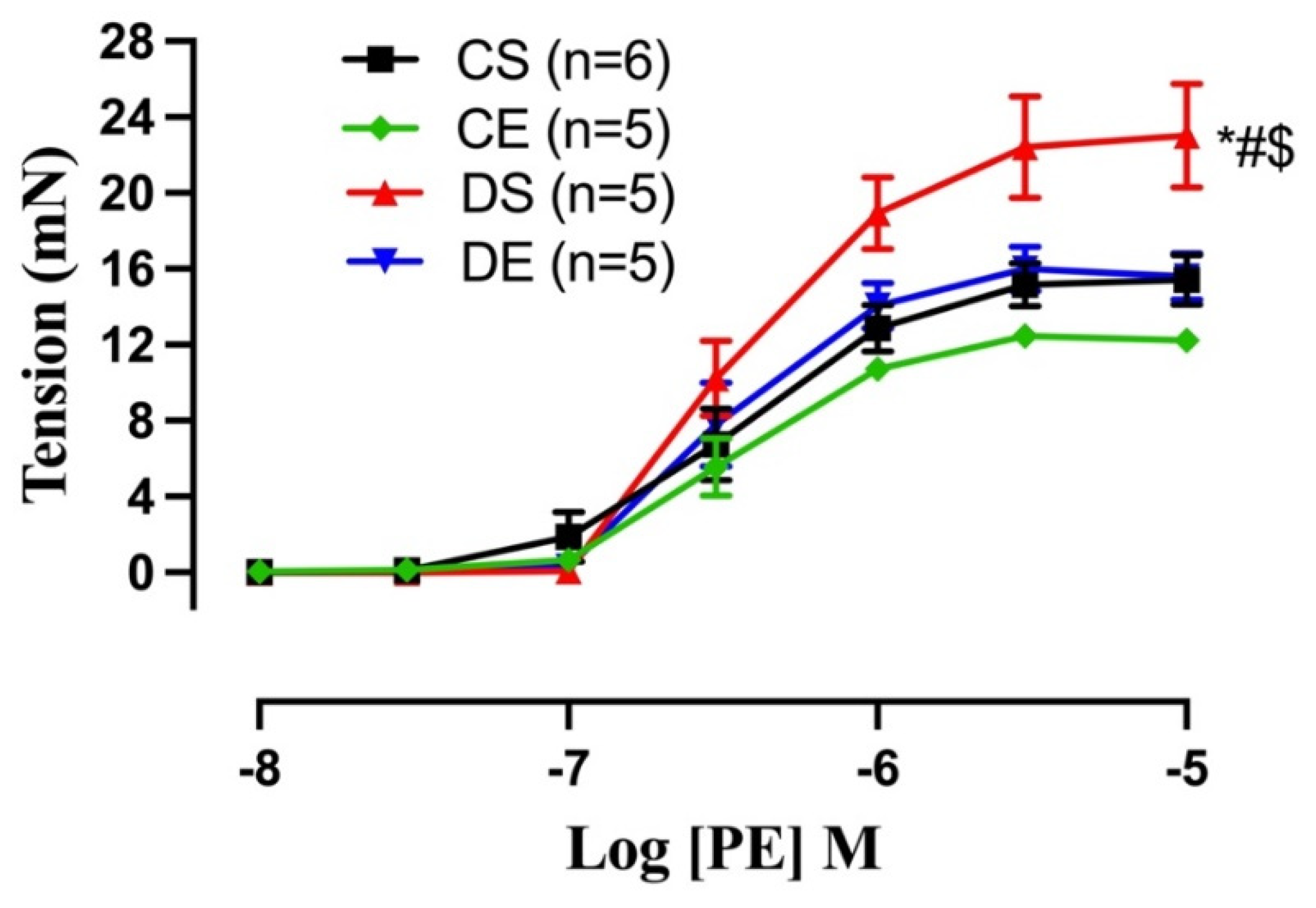
3.6. Effects of MIE on the PE-induced Contraction
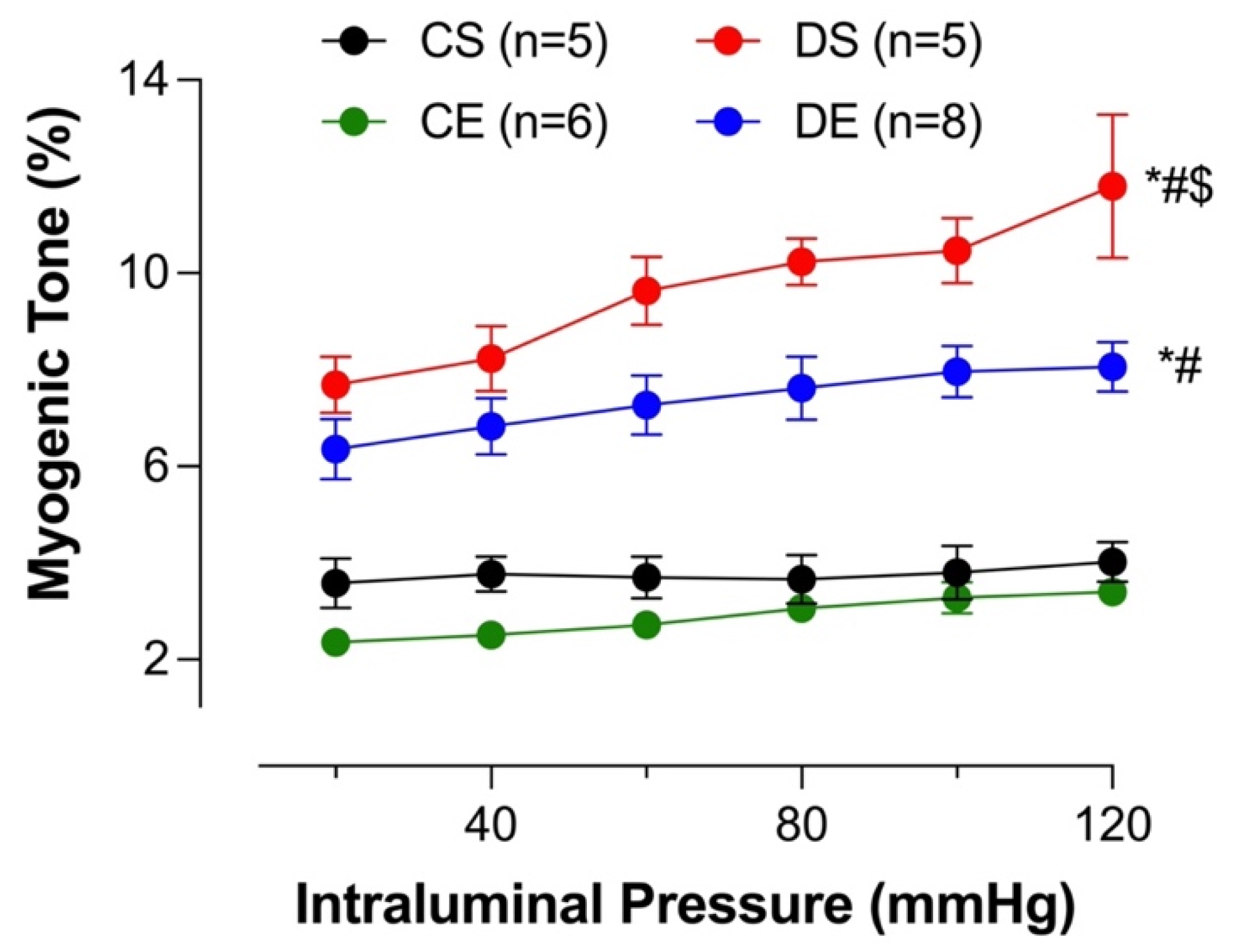
3.7. Effects of MIE on the Myogenic Tone
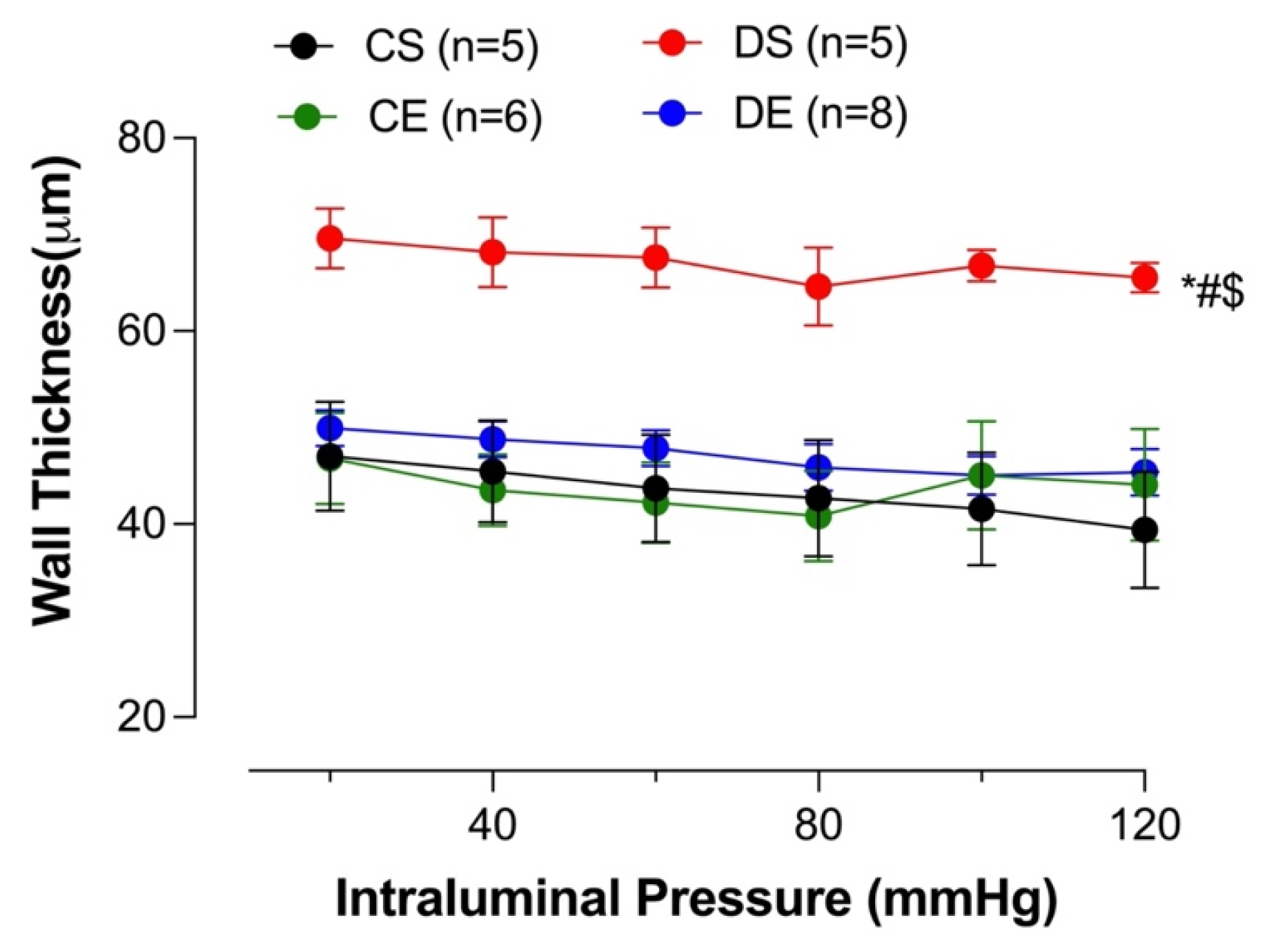
3.8. Effects of MIE on Wall Thickness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kannel, W.B.; McGee, D.L. Diabetes and Cardiovascular Disease. The Framingham Study. JAMA 1979, 241, 2035–2038. [Google Scholar] [CrossRef]
- Morrish, N.J.; Wang, S.L.; Stevens, L.K.; Fuller, J.H.; Keen, H. Mortality and Causes of Death in the WHO Multinational Study of Vascular Disease in Diabetes. Diabetologia 2001, 44 (Suppl. S2), S14–S21. [Google Scholar] [CrossRef] [Green Version]
- Asif, M. The Prevention and Control the Type-2 Diabetes by Changing Lifestyle and Dietary Pattern. J. Educ. Health Promot. 2014, 3, 1. [Google Scholar] [CrossRef]
- Zhang, R.; Thor, D.; Han, X.; Anderson, L.; Rahimian, R. Sex Differences in Mesenteric Endothelial Function of Streptozotocin-Induced Diabetic Rats: A Shift in the Relative Importance of EDRFs. Am. J. Physiol. Heart Circ. Physiol. 2012, 303, H1183–H1198. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Shaligram, S.; Zhang, R.; Anderson, L.; Rahimian, R. Sex-Specific Vascular Responses of the Rat Aorta: Effects of Moderate Term (Intermediate Stage) Streptozotocin-Induced Diabetes. Can. J. Physiol. Pharmacol. 2016, 94, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Shaligram, S.; Akther, F.; Razan, M.R.; Graham, J.L.; Roglans, N.; Alegret, M.; Parsa, A.G.; Stanhope, K.L.; Havel, P.J.; Rahimian, R. Mesenteric Arterial Dysfunction in the UC Davis Type 2 Diabetes Mellitus Rat Model Is Dependent on Pre-Diabetic versus Diabetic Status and Is Sexually Dimorphic. Eur. J. Pharmacol. 2020, 879, 173089. [Google Scholar] [CrossRef]
- Félétou, M.; Huang, Y.; Vanhoutte, P.M. Endothelium-Mediated Control of Vascular Tone: COX-1 and COX-2 Products. Br. J. Pharmacol. 2011, 164, 894–912. [Google Scholar] [CrossRef] [Green Version]
- Cummings, B.P.; Digitale, E.K.; Stanhope, K.L.; Graham, J.L.; Baskin, D.G.; Reed, B.J.; Sweet, I.R.; Griffen, S.C.; Havel, P.J. Development and Characterization of a Novel Rat Model of Type 2 Diabetes Mellitus: The UC Davis Type 2 Diabetes Mellitus UCD-T2DM Rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R1782–R1793. [Google Scholar] [CrossRef] [Green Version]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.M.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schürmann, A.; et al. Animal Models of Obesity and Diabetes Mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Akther, F.; Razan, M.R.; Shaligram, S.; Graham, J.L.; Stanhope, K.L.; Allen, K.N.; Vázquez-Medina, J.P.; Havel, P.J.; Rahimian, R. Potentiation of Acetylcholine-Induced Relaxation of Aorta in Male UC Davis Type 2 Diabetes Mellitus (UCD-T2DM) Rats: Sex-Specific Responses. Front. Physiol. 2021, 12, 616317. [Google Scholar] [CrossRef]
- Haram, P.M.; Kemi, O.J.; Wisloff, U. Adaptation of Endothelium to Exercise Training: Insights from Experimental Studies. Front. Biosci. 2008, 13, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Nieman, D.C.; Wentz, L.M. The Compelling Link between Physical Activity and the Body’s Defense System. J. Sport Health Sci. 2019, 8, 201–217. [Google Scholar] [CrossRef]
- Okada, S.; Hiuge, A.; Makino, H.; Nagumo, A.; Takaki, H.; Konishi, H.; Goto, Y.; Yoshimasa, Y.; Miyamoto, Y. Effect of Exercise Intervention on Endothelial Function and Incidence of Cardiovascular Disease in Patients with Type 2 Diabetes. J. Atheroscler. Thromb. 2010, 17, 828–833. [Google Scholar] [CrossRef] [Green Version]
- Wisløff, U.; Helgerud, J.; Kemi, O.J.; Ellingsen, O. Intensity-Controlled Treadmill Running in Rats: VO(2 Max) and Cardiac Hypertrophy. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H1301–H1310. [Google Scholar] [CrossRef] [Green Version]
- de Lade, C.G.; Andreazzi, A.E.; Bolotari, M.; Costa, V.M.G.; Peters, V.M.; Guerra, M. de O. Effects of Moderate Intensity Endurance Training vs. High Intensity Interval Training on Weight Gain, Cardiorespiratory Capacity, and Metabolic Profile in Postnatal Overfed Rats. Diabetol. Metab. Syndr. 2018, 10, 70. [Google Scholar] [CrossRef]
- Chung, E.; Diffee, G.M. Moderate Intensity, but Not High Intensity, Treadmill Exercise Training Alters Power Output Properties in Myocardium from Aged Rats. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 1178–1187. [Google Scholar] [CrossRef] [Green Version]
- Howarth, F.C.; Almugaddum, F.A.; Qureshi, M.A.; Ljubisavljevic, M.; Ljubisavijevic, M. Effects of Varying Intensity Exercise on Shortening and Intracellular Calcium in Ventricular Myocytes from Streptozotocin (STZ)-Induced Diabetic Rats. Mol. Cell. Biochem. 2008, 317, 161–167. [Google Scholar] [CrossRef]
- Yang, H.-L.; Hsieh, P.-L.; Hung, C.-H.; Cheng, H.-C.; Chou, W.-C.; Chu, P.-M.; Chang, Y.-C.; Tsai, K.-L. Early Moderate Intensity Aerobic Exercise Intervention Prevents Doxorubicin-Caused Cardiac Dysfunction through Inhibition of Cardiac Fibrosis and Inflammation. Cancers 2020, 12, 1102. [Google Scholar] [CrossRef]
- Tain, Y.-L.; Chang-Chien, G.-P.; Lin, S.; Hou, C.-Y.; Hsu, C.-N. Iodomethylcholine Inhibits Trimethylamine-N-Oxide Production and Averts Maternal Chronic Kidney Disease-Programmed Offspring Hypertension. Int. J. Mol. Sci. 2023, 24, 1284. [Google Scholar] [CrossRef]
- Morales-Ríos, E.I.; García-Machorro, J.; Briones-Aranda, A.; Gómez-Pliego, R.; Espinosa-Raya, J. Effect of Long-Term Intake of Nutritive and Non-Nutritive Sweeteners on Metabolic Health and Cognition in Adult Male Rats. J. Med. Food 2022, 25, 1059–1065. [Google Scholar] [CrossRef]
- Shaligram, S.; Sangüesa, G.; Akther, F.; Alegret, M.; Laguna, J.C.; Rahimian, R. Differential Effects of High Consumption of Fructose or Glucose on Mesenteric Endothelial Function in Female Rats. J. Nutr. Biochem. 2018, 57, 136. [Google Scholar] [CrossRef]
- Bridges, L.E.; Williams, C.L.; Pointer, M.A.; Awumey, E.M. Mesenteric Artery Contraction and Relaxation Studies Using Automated Wire Myography. J. Vis. Exp. 2011, 55, e3119. [Google Scholar] [CrossRef]
- Wenceslau, C.F.; McCarthy, C.G.; Earley, S.; England, S.K.; Filosa, J.A.; Goulopoulou, S.; Gutterman, D.D.; Isakson, B.E.; Kanagy, N.L.; Martinez-Lemus, L.A.; et al. Guidelines for the Measurement of Vascular Function and Structure in Isolated Arteries and Veins. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H77–H111. [Google Scholar] [CrossRef]
- Razan, M.R. Impact of 17-Beta Estradiol and Moderate-Intensity Exercise on Mesenteric Arterial Function of UC Davis Type-2 Diabetes Mellitus Rats. Ph.D. Thesis, University of the Pacific, Stockton, CA, USA, 2021. [Google Scholar]
- Razan, M.R.; Akther, F.; Islam, R.A.; Graham, J.L.; Stanhope, K.L.; Havel, P.J.; Rahimian, R. 17β-Estradiol Treatment Improves Acetylcholine-Induced Relaxation of Mesenteric Arteries in Ovariectomized UC Davis Type 2 Diabetes Mellitus Rats in Prediabetic State. Front. Physiol. 2022, 13, 900813. [Google Scholar] [CrossRef]
- Kjeldsen, S.E. Hypertension and Cardiovascular Risk: General Aspects. Pharmacol. Res. 2018, 129, 95–99. [Google Scholar] [CrossRef]
- Sunano, S.; Watanabe, H.; Tanaka, S.; Sekiguchi, F.; Shimamura, K. Endothelium-Derived Relaxing, Contracting and Hyperpolarizing Factors of Mesenteric Arteries of Hypertensive and Normotensive Rats. Br. J. Pharmacol. 1999, 126, 709–716. [Google Scholar] [CrossRef] [Green Version]
- Matchkov, V.V.; Kudryavtseva, O.; Aalkjaer, C. Intracellular Ca2+ Signalling and Phenotype of Vascular Smooth Muscle Cells. Basic Clin. Pharm. Toxicol. 2012, 110, 42–48. [Google Scholar] [CrossRef]
- Sangüesa, G.; Shaligram, S.; Akther, F.; Roglans, N.; Laguna, J.C.; Rahimian, R.; Alegret, M. Type of Supplemented Simple Sugar, Not Merely Calorie Intake, Determines Adverse Effects on Metabolism and Aortic Function in Female Rats. Am. J. Physiol.—Heart Circ. Physiol. 2017, 312, H289–H304. [Google Scholar] [CrossRef] [Green Version]
- McCloskey, K.; Vuillermin, P.; Ponsonby, A.-L.; Cheung, M.; Skilton, M.R.; Burgner, D. Aortic Intima-Media Thickness Measured by Trans-Abdominal Ultrasound as an Early Life Marker of Subclinical Atherosclerosis. Acta Paediatr. 2014, 103, 124–130. [Google Scholar] [CrossRef]
- Crijns, F.R.L.; Wolffenbuttel, B.H.R.; de Mey, J.G.R.; Boudier, H.A.J.S. Mechanical Properties of Mesenteric Arteries in Diabetic Rats: Consequences of Outward Remodeling. Am. J. Physiol. Heart Circ. Physiol. 1999, 276, H1672–H1677. [Google Scholar] [CrossRef]
- Souza-Smith, F.M.; Katz, P.S.; Trask, A.J.; Stewart, J.A., Jr.; Lord, K.C.; Varner, K.J.; Vassallo, D.V.; Lucchesi, P.A. Mesenteric Resistance Arteries in Type 2 Diabetic Db/Db Mice Undergo Outward Remodeling. PLoS ONE 2011, 6, e23337. [Google Scholar] [CrossRef]
- Sanz, C.; Gautier, J.-F.; Hanaire, H. Physical Exercise for the Prevention and Treatment of Type 2 Diabetes. Diabetes Metab. 2010, 36, 346–351. [Google Scholar] [CrossRef]
- Avery, L.; Flynn, D.; van Wersch, A.; Sniehotta, F.F.; Trenell, M.I. Changing Physical Activity Behavior in Type 2 Diabetes: A Systematic Review and Meta-Analysis of Behavioral Interventions. Diabetes Care 2012, 35, 2681–2689. [Google Scholar] [CrossRef] [Green Version]
- Umpierre, D.; Ribeiro, P.A.B.; Schaan, B.D.; Ribeiro, J.P. Volume of Supervised Exercise Training Impacts Glycaemic Control in Patients with Type 2 Diabetes: A Systematic Review with Meta-Regression Analysis. Diabetologia 2013, 56, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Roberts, C.K.; Little, J.P.; Thyfault, J.P. Modification of Insulin Sensitivity and Glycemic Control by Activity and Exercise. Med. Sci. Sport. Exerc. 2013, 45, 1868–1877. [Google Scholar] [CrossRef]
- Colberg, S.R.; Sigal, R.J.; Fernhall, B.; Regensteiner, J.G.; Blissmer, B.J.; Rubin, R.R.; Chasan-Taber, L.; Albright, A.L.; Braun, B. Exercise and Type 2 Diabetes: The American College of Sports Medicine and the American Diabetes Association: Joint Position Statement. Diabetes Care 2010, 33, e147–e167. [Google Scholar] [CrossRef] [Green Version]
- American Diabetes Association Physical Activity/Exercise and Diabetes. Diabetes Care 2004, 27 (Suppl. S1), S58–S62. [CrossRef] [Green Version]
- Chen, Y.; Sloan, F.A.; Yashkin, A.P. Adherence to Diabetes Guidelines for Screening, Physical Activity and Medication and Onset of Complications and Death. J. Diabetes Complicat. 2015, 29, 1228–1233. [Google Scholar] [CrossRef]
- De Vriese, A.S.; Verbeuren, T.J.; Van de Voorde, J.; Lameire, N.H.; Vanhoutte, P.M. Endothelial Dysfunction in Diabetes. Br. J. Pharmacol. 2000, 130, 963–974. [Google Scholar] [CrossRef] [Green Version]
- Moien-Afshari, F.; Ghosh, S.; Khazaei, M.; Kieffer, T.J.; Brownsey, R.W.; Laher, I. Exercise Restores Endothelial Function Independently of Weight Loss or Hyperglycaemic Status in Db/Db Mice. Diabetologia 2008, 51, 1327–1337. [Google Scholar] [CrossRef] [Green Version]
- Takenouchi, Y.; Kobayashi, T.; Taguchi, K.; Matsumoto, T.; Kamata, K. Gender Differences in Vascular Reactivity of Aortas from Streptozotocin-Induced Diabetic Mice. Biol. Pharm. Bull. 2010, 33, 1692–1697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, S.; Cai, X.; Yin, H.; Sun, Z.; Zügel, M.; Steinacker, J.M.; Schumann, U. Exercise Training and Endothelial Function in Patients with Type 2 Diabetes: A Meta-Analysis. Cardiovasc. Diabetol. 2018, 17, 64. [Google Scholar] [CrossRef]
- Sakamoto, S.; Minami, K.; Niwa, Y.; Ohnaka, M.; Nakaya, Y.; Mizuno, A.; Kuwajima, M.; Shima, K. Effect of Exercise Training and Food Restriction on Endothelium-Dependent Relaxation in the Otsuka Long-Evans Tokushima Fatty Rat, a Model of Spontaneous NIDDM. Diabetes 1998, 47, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Y.; Lin, Y.W.; Clemont, A.; Feener, E.P.; Hein, K.D.; Igarashi, M.; Yamauchi, T.; White, M.F.; King, G.L. Characterization of Selective Resistance to Insulin Signaling in the Vasculature of Obese Zucker (Fa/Fa) Rats. J. Clin. Investig. 1999, 104, 447–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potenza, M.A.; Marasciulo, F.L.; Chieppa, D.M.; Brigiani, G.S.; Formoso, G.; Quon, M.J.; Montagnani, M. Insulin Resistance in Spontaneously Hypertensive Rats Is Associated with Endothelial Dysfunction Characterized by Imbalance between NO and ET-1 Production. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H813–H822. [Google Scholar] [CrossRef] [PubMed]
- Symons, J.D.; McMillin, S.L.; Riehle, C.; Tanner, J.; Palionyte, M.; Hillas, E.; Jones, D.; Cooksey, R.C.; Birnbaum, M.J.; McClain, D.A.; et al. Contribution of Insulin and Akt1 Signaling to Endothelial Nitric Oxide Synthase in the Regulation of Endothelial Function and Blood Pressure. Circ. Res. 2009, 104, 1085–1094. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, S.H.; Karstoft, K.; Winding, K.; Holst, J.J.; Pedersen, B.K.; Solomon, T.P.J. Effects of Acute Exercise on Pancreatic Endocrine Function in Subjects with Type 2 Diabetes. Diabetes Obes. Metab. 2015, 17, 207–210. [Google Scholar] [CrossRef]
- Ito, D.; Cao, P.; Kakihana, T.; Sato, E.; Suda, C.; Muroya, Y.; Ogawa, Y.; Hu, G.; Ishii, T.; Ito, O.; et al. Chronic Running Exercise Alleviates Early Progression of Nephropathy with Upregulation of Nitric Oxide Synthases and Suppression of Glycation in Zucker Diabetic Rats. PLoS ONE 2015, 10, e0138037. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, L.; Xu, D. Aerobic Exercise Reduces Triglycerides by Targeting Apolipoprotein C3 in Patients with Coronary Heart Disease. Clin. Cardiol. 2019, 42, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Zicha, J.; Pechánová, O.; Cacányiová, S.; Cebová, M.; Kristek, F.; Török, J.; Simko, F.; Dobesová, Z.; Kunes, J. Hereditary Hypertriglyceridemic Rat: A Suitable Model of Cardiovascular Disease and Metabolic Syndrome? Physiol. Res. 2006, 55 (Suppl. S1), S49–S63. [Google Scholar] [CrossRef]
- Sattar, N.; Petrie, J.R.; Jaap, A.J. The Atherogenic Lipoprotein Phenotype and Vascular Endothelial Dysfunction. Atherosclerosis 1998, 138, 229–235. [Google Scholar] [CrossRef]
- Wang, L.; Gill, R.; Pedersen, T.L.; Higgins, L.J.; Newman, J.W.; Rutledge, J.C. Triglyceride-Rich Lipoprotein Lipolysis Releases Neutral and Oxidized FFAs That Induce Endothelial Cell Inflammation. J. Lipid Res. 2009, 50, 204–213. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, S.; Gotoh, N.; Hishinuma, S.; Abe, Y.; Shimizu, Y.; Katano, Y.; Ishihata, A. The Role of Hypertriglyceridemia in the Development of Atherosclerosis and Endothelial Dysfunction. Nutrients 2014, 6, 1236–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucero, D.; López, G.I.; Gorzalczany, S.; Duarte, M.; González Ballerga, E.; Sordá, J.; Schreier, L.; Zago, V. Alterations in Triglyceride Rich Lipoproteins Are Related to Endothelial Dysfunction in Metabolic Syndrome. Clin. Biochem. 2016, 49, 932–935. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, M.; Maruhashi, T.; Matsumoto, T.; Iwamoto, Y.; Iwamoto, A.; Oda, N.; Kishimoto, S.; Matsui, S.; Aibara, Y.; Hidaka, T.; et al. Relationship between Serum Triglyceride Levels and Endothelial Function in a Large Community-Based Study. Atherosclerosis 2016, 249, 70–75. [Google Scholar] [CrossRef]
- Tomioka, H.; Hattori, Y.; Fukao, M.; Sato, A.; Liu, M.; Sakuma, I.; Kitabatake, A.; Kanno, M. Relaxation in Different-Sized Rat Blood Vessels Mediated by Endothelium-Derived Hyperpolarizing Factor: Importance of Processes Mediating Precontractions. J. Vasc. Res. 1999, 36, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Zygmunt, P.M.; Ryman, T.; Högestätt, E.D. Regional Differences in Endothelium-Dependent Relaxation in the Rat: Contribution of Nitric Oxide and Nitric Oxide-Independent Mechanisms. Acta Physiol. Scand. 1995, 155, 257–266. [Google Scholar] [CrossRef]
- Feletou, M.; Vanhoutte, P.M. Endothelium-Dependent Hyperpolarization of Canine Coronary Smooth Muscle. Br. J. Pharmacol. 1988, 93, 515–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komori, K.; Lorenz, R.R.; Vanhoutte, P.M. Nitric Oxide, ACh, and Electrical and Mechanical Properties of Canine Arterial Smooth Muscle. Am. J. Physiol. 1988, 255, H207–H212. [Google Scholar] [CrossRef]
- Doughty, J.M.; Plane, F.; Langton, P.D. Charybdotoxin and Apamin Block EDHF in Rat Mesenteric Artery If Selectively Applied to the Endothelium. Am. J. Physiol. 1999, 276, H1107–H1112. [Google Scholar] [CrossRef]
- Bitar, M.S.; Wahid, S.; Mustafa, S.; Al-Saleh, E.; Dhaunsi, G.S.; Al-Mulla, F. Nitric Oxide Dynamics and Endothelial Dysfunction in Type II Model of Genetic Diabetes. Eur. J. Pharmacol. 2005, 511, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Desrois, M.; Clarke, K.; Lan, C.; Dalmasso, C.; Cole, M.; Portha, B.; Cozzone, P.J.; Bernard, M. Upregulation of eNOS and Unchanged Energy Metabolism in Increased Susceptibility of the Aging Type 2 Diabetic GK Rat Heart to Ischemic Injury. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1679–H1686. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Harrison, D.G. Endothelial Dysfunction in Cardiovascular Diseases: The Role of Oxidant Stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef] [Green Version]
- Bauersachs, J.; Popp, R.; Hecker, M.; Sauer, E.; Fleming, I.; Busse, R. Nitric Oxide Attenuates the Release of Endothelium-Derived Hyperpolarizing Factor. Circulation 1996, 94, 3341–3347. [Google Scholar] [CrossRef]
- Brandes, R.P.; Schmitz-Winnenthal, F.-H.; Félétou, M.; Gödecke, A.; Huang, P.L.; Vanhoutte, P.M.; Fleming, I.; Busse, R. An Endothelium-Derived Hyperpolarizing Factor Distinct from NO and Prostacyclin Is a Major Endothelium-Dependent Vasodilator in Resistance Vessels of Wild-Type and Endothelial NO Synthase Knockout Mice. Proc. Natl. Acad. Sci. USA 2000, 97, 9747–9752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garland, C.J.; Plane, F.; Kemp, B.K.; Cocks, T.M. Endothelium-Dependent Hyperpolarization: A Role in the Control of Vascular Tone. Trends Pharmacol. Sci. 1995, 16, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Malakul, W.; Thirawarapan, S.; Suvitayavat, W.; Woodman, O.L. Type 1 Diabetes and Hypercholesterolaemia Reveal the Contribution of Endothelium-Derived Hyperpolarizing Factor to Endothelium-Dependent Relaxation of the Rat Aorta. Clin. Exp. Pharmacol. Physiol. 2008, 35, 192–200. [Google Scholar] [CrossRef]
- Kilpatrick, E.V.; Cocks, T.M. Evidence for Differential Roles of Nitric Oxide (NO) and Hyperpolarization in Endothelium-Dependent Relaxation of Pig Isolated Coronary Artery. Br. J. Pharmacol. 1994, 112, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Oniki, H.; Fujii, K.; Kansui, Y.; Goto, K.; Iida, M. Effects of Angiotensin II Receptor Antagonist on Impaired Endothelium-Dependent and Endothelium-Independent Relaxations in Type II Diabetic Rats. J. Hypertens. 2006, 24, 331–338. [Google Scholar] [CrossRef]
- Félétou, M.; Vanhoutte, P.M. EDHF: New Therapeutic Targets? Pharmacol. Res. 2004, 49, 565–580. [Google Scholar] [CrossRef]
- Minami, A.; Ishimura, N.; Harada, N.; Sakamoto, S.; Niwa, Y.; Nakaya, Y. Exercise Training Improves Acetylcholine-Induced Endothelium-Dependent Hyperpolarization in Type 2 Diabetic Rats, Otsuka Long-Evans Tokushima Fatty Rats. Atherosclerosis 2002, 162, 85–92. [Google Scholar] [CrossRef]
- Moran, T.H.; Bi, S. Hyperphagia and Obesity in OLETF Rats Lacking CCK-1 Receptors. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1211–1218. [Google Scholar] [CrossRef] [Green Version]
- Villar, I.C.; Hobbs, A.J.; Ahluwalia, A. Sex Differences in Vascular Function: Implication of Endothelium-Derived Hyperpolarizing Factor. J. Endocrinol. 2008, 197, 447–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, W.B. New Role for Epoxyeicosatrienoic Acids as Anti-Inflammatory Mediators. Trends Pharm. Sci. 2000, 21, 125–127. [Google Scholar] [CrossRef] [PubMed]
- Node, K.; Huo, Y.; Ruan, X.; Yang, B.; Spiecker, M.; Ley, K.; Zeldin, D.C.; Liao, J.K. Anti-Inflammatory Properties of Cytochrome P450 Epoxygenase-Derived Eicosanoids. Science 1999, 285, 1276–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, M.; Sánchez, A.; Pilar Martínez, M.; Benedito, S.; López-Oliva, M.-E.; García-Sacristán, A.; Hernández, M.; Prieto, D. COX-2 Is Involved in Vascular Oxidative Stress and Endothelial Dysfunction of Renal Interlobar Arteries from Obese Zucker Rats. Free Radic. Biol. Med. 2015, 84, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Avendaño, M.S.; Martínez-Revelles, S.; Aguado, A.; Simões, M.R.; González-Amor, M.; Palacios, R.; Guillem-Llobat, P.; Vassallo, D.V.; Vila, L.; García-Puig, J.; et al. Role of COX-2-derived PGE2 on Vascular Stiffness and Function in Hypertension. Br. J. Pharmacol. 2016, 173, 1541–1555. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Revelles, S.; Avendaño, M.S.; García-Redondo, A.B.; Alvarez, Y.; Aguado, A.; Pérez-Girón, J.V.; García-Redondo, L.; Esteban, V.; Redondo, J.M.; Alonso, M.J.; et al. Reciprocal Relationship between Reactive Oxygen Species and Cyclooxygenase-2 and Vascular Dysfunction in Hypertension. Antioxid. Redox Signal. 2013, 18, 51–65. [Google Scholar] [CrossRef]
- Alvarez, Y.; Pérez-Girón, J.V.; Hernanz, R.; Briones, A.M.; García-Redondo, A.; Beltrán, A.; Alonso, M.J.; Salaices, M. Losartan Reduces the Increased Participation of Cyclooxygenase-2-Derived Products in Vascular Responses of Hypertensive Rats. J. Pharmacol. Exp. Ther. 2007, 321, 381–388. [Google Scholar] [CrossRef]
- Santiago, E.; Martínez, M.P.; Climent, B.; Muñoz, M.; Briones, A.M.; Salaices, M.; García-Sacristán, A.; Rivera, L.; Prieto, D. Augmented Oxidative Stress and Preserved Vasoconstriction Induced by Hydrogen Peroxide in Coronary Arteries in Obesity: Role of COX-2. Br. J. Pharmacol. 2016, 173, 3176–3195. [Google Scholar] [CrossRef] [Green Version]
- Fleming, I.; Bauersachs, J.; Schäfer, A.; Scholz, D.; Aldershvile, J.; Busse, R. Isometric Contraction Induces the Ca2+-Independent Activation of the Endothelial Nitric Oxide Synthase. Proc. Natl. Acad. Sci. USA 1999, 96, 1123–1128. [Google Scholar] [CrossRef] [Green Version]
- Dora, K.A.; Hinton, J.M.; Walker, S.D.; Garland, C.J. An Indirect Influence of Phenylephrine on the Release of Endothelium-Derived Vasodilators in Rat Small Mesenteric Artery. Br. J. Pharm. 2000, 129, 381–387. [Google Scholar] [CrossRef]
- Jackson, W.F.; Boerman, E.M.; Lange, E.J.; Lundback, S.S.; Cohen, K.D. Smooth Muscle A1D-Adrenoceptors Mediate Phenylephrine-Induced Vasoconstriction and Increases in Endothelial Cell Ca2+ in Hamster Cremaster Arterioles. Br. J. Pharmacol. 2008, 155, 514–524. [Google Scholar] [CrossRef] [Green Version]
- Lagaud, G.J.L.; Masih-Khan, E.; Kai, S.; Breemen, C.v.; Dubé, G.P. Influence of Type II Diabetes on Arterial Tone and Endothelial Function in Murine Mesenteric Resistance Arteries. JVR 2001, 38, 578–589. [Google Scholar] [CrossRef]
- Jarajapu, Y.P.; Guberski, D.L.; Grant, M.B.; Knot, H.J. Myogenic Tone and Reactivity of Cerebral Arteries in Type II Diabetic BBZDR/Wor Rat. Eur. J. Pharmacol. 2008, 579, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Frisbee, J.C.; Maier, K.G.; Stepp, D.W. Oxidant Stress-Induced Increase in Myogenic Activation of Skeletal Muscle Resistance Arteries in Obese Zucker Rats. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H2160–H2168. [Google Scholar] [CrossRef] [Green Version]
- Bagi, Z.; Erdei, N.; Toth, A.; Li, W.; Hintze, T.H.; Koller, A.; Kaley, G. Type 2 Diabetic Mice Have Increased Arteriolar Tone and Blood Pressure. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1610–1616. [Google Scholar] [CrossRef] [Green Version]
- Leo, M.D.; Peixoto-Nieves, D.; Yin, W.; Raghavan, S.; Muralidharan, P.; Mata-Daboin, A.; Jaggar, J.H. TMEM16A Channel Upregulation in Arterial Smooth Muscle Cells Produces Vasoconstriction during Diabetes. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1089–H1101. [Google Scholar] [CrossRef]
- Spinetti, G.; Kraenkel, N.; Emanueli, C.; Madeddu, P. Diabetes and Vessel Wall Remodelling: From Mechanistic Insights to Regenerative Therapies. Cardiovasc. Res. 2008, 78, 265–273. [Google Scholar] [CrossRef] [Green Version]
- Konukoglu, D.; Uzun, H. Endothelial Dysfunction and Hypertension. Adv. Exp. Med. Biol. 2017, 956, 511–540. [Google Scholar] [CrossRef]
- Gallo, G.; Volpe, M.; Savoia, C. Endothelial Dysfunction in Hypertension: Current Concepts and Clinical Implications. Front. Med. 2021, 8, 798958. [Google Scholar] [CrossRef] [PubMed]
- de Groot, E.; Hovingh, G.K.; Wiegman, A.; Duriez, P.; Smit, A.J.; Fruchart, J.-C.; Kastelein, J.J.P. Measurement of Arterial Wall Thickness as a Surrogate Marker for Atherosclerosis. Circulation 2004, 109, III33–III38. [Google Scholar] [CrossRef] [Green Version]
- Briones, A.M.; Salaices, M.; Vila, E. Mechanisms Underlying Hypertrophic Remodeling and Increased Stiffness of Mesenteric Resistance Arteries from Aged Rats. J. Gerontol. A Biol. Sci. Med. Sci. 2007, 62, 696–706. [Google Scholar] [CrossRef] [Green Version]
- Savoia, C.; D’Agostino, M.; Lauri, F.; Volpe, M. Angiotensin Type 2 Receptor in Hypertensive Cardiovascular Disease. Curr. Opin. Nephrol. Hypertens. 2011, 20, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.; Schiffrin, E.L. Small Artery Remodeling Is the Most Prevalent (Earliest?) Form of Target Organ Damage in Mild Essential Hypertension. J. Hypertens. 2001, 19, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Luan, Y.; Zhang, B.; Zhang, S.; Wang, X. Morphological Study of the Effect of Aerobic Exercise on Organs and Arteries in Spontaneously Hypertensive Rats. Healthcare 2021, 9, 1066. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric Oxide: Physiology, Pathophysiology, and Pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar]
- Parkington, H.C.; Chow, J.A.M.; Evans, R.G.; Coleman, H.A.; Tare, M. Role for Endothelium-Derived Hyperpolarizing Factor in Vascular Tone in Rat Mesenteric and Hindlimb Circulations in Vivo. J. Physiol. 2002, 542, 929–937. [Google Scholar] [CrossRef]
- Ralevic, V. Endothelial Nitric Oxide Modulates Perivascular Sensory Neurotransmission in the Rat Isolated Mesenteric Arterial Bed. Br. J. Pharmacol. 2002, 137, 19–28. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | mmHg | CS | CE | DS | DE |
|---|---|---|---|---|---|
| 0 week | SBP | 114 ± 3 | 119 ± 6 | 155 ± 8 *# | 153 ± 5 *# |
| DBP | 80 ± 2 | 86 ± 3 | 117 ± 8 *# | 112 ± 6 *# | |
| MAP | 95 ± 3 | 99 ± 4 | 129 ± 8 *# | 125 ± 6 *# | |
| 8 weeks | SBP | 114 ± 3 | 98 ± 4 a | 138 ± 5 *#$ | 115 ± 5 a |
| DBP | 76 ± 2 | 84 ± 10 | 93 ± 3 | 79 ± 4 b | |
| MAP | 91 ± 3 | 85 ± 8 | 104 ± 7 c | 91 ± 4 c |
| ACh | n | pD2 (-logEC50) | Rmax (%) |
|---|---|---|---|
| CS | 7 | 7.40 ± 0.2 | 98.91 ± 0.6 |
| CE | 5 | 7.22 ± 0.1 | 98.60 ± 1.0 |
| DS | 5 | 5.80 ± 0.2 *#$ | 59.85 ± 2.2 *#$ |
| DE | 8 | 6.72 ± 0.1 # | 90.74 ± 2.4 *# |
| Groups | No Drug | Indo | Indo + L-NAME | Indo + L-NAME + Apamin + TRAM-34 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pD2 | Rmax, (%) | ΔAUC | pD2 | Rmax (%) | ΔAUC | pD2 | Rmax (%) | ΔAUC | pD2 | Rmax (%) | ΔAUC | |
| CS | 7.63 ± 0.1 | 99.57 ± 0.1 | ND | 7.49 ± 0.1 | 99.41 ± 0.2 | 13.80 ± 5.71 | ND | 80.54 ± 7.3 | 93.90 ± 9.62 | ND | −1.71 ± 2.5 abc | 147.3 ± 8.34 |
| CE | 7.22 ± 0.12 | 98.60 ± 0.9 | ND | 7.26 ± 0.2 | 99.83 ± 0.2 | 1.0 ± 7.66 | ND | 93.88 ± 1.9 | 75.20 ± 8.13 | ND | −6.67 ± 1.4 abc | 144.7 ± 5.63 |
| DS | 5.80 ± 0.2 *#$ | 59.85 ± 2.2 *#$ | ND | 6.70 ± 0.2 #a | 98.61 ± 0.4 a*#$ | 47.20 ± 4.77 *#$ | ND | 10.27 ± 3.7 *#ab | 115.9 ± 3.45 * | ND | −7.36 ± 2.5 abc | 6.24 ± 1.83 *#$ |
| DE | 6.72 ± 0.1 # | 90.74 ± 2.4 *# | ND | 6.61 ± 0.1 *# | 88.92 ± 3.4 | 0.5 ± 7.49 | ND | 38.58 ± 13.9 *#ab | 107.7 ± 10.1 | ND | −6.5 ± 1.4 abc | 48.38 ± 9.1 *# |
| PE | n | pD2(-logEC50) | Tensionmax |
|---|---|---|---|
| CS | 6 | 6.49 ± 0.16 | 15.45 ± 1.24 |
| CE | 5 | 6.46 ± 0.12 | 12.23 ± 0.54 |
| DS | 5 | 6.46 ± 0.18 | 23.03 ± 2.74 *#$ |
| DE | 5 | 6.41 ± 0.11 | 16.20 ± 1.18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razan, M.R.; Amissi, S.; Islam, R.A.; Graham, J.L.; Stanhope, K.L.; Havel, P.J.; Rahimian, R. Moderate-Intensity Exercise Improves Mesenteric Arterial Function in Male UC Davis Type-2 Diabetes Mellitus (UCD-T2DM) Rats: A Shift in the Relative Importance of Endothelium-Derived Relaxing Factors (EDRF). Biomedicines 2023, 11, 1129. https://doi.org/10.3390/biomedicines11041129
Razan MR, Amissi S, Islam RA, Graham JL, Stanhope KL, Havel PJ, Rahimian R. Moderate-Intensity Exercise Improves Mesenteric Arterial Function in Male UC Davis Type-2 Diabetes Mellitus (UCD-T2DM) Rats: A Shift in the Relative Importance of Endothelium-Derived Relaxing Factors (EDRF). Biomedicines. 2023; 11(4):1129. https://doi.org/10.3390/biomedicines11041129
Chicago/Turabian StyleRazan, Md Rahatullah, Said Amissi, Rifat Ara Islam, James L. Graham, Kimber L. Stanhope, Peter J. Havel, and Roshanak Rahimian. 2023. "Moderate-Intensity Exercise Improves Mesenteric Arterial Function in Male UC Davis Type-2 Diabetes Mellitus (UCD-T2DM) Rats: A Shift in the Relative Importance of Endothelium-Derived Relaxing Factors (EDRF)" Biomedicines 11, no. 4: 1129. https://doi.org/10.3390/biomedicines11041129
APA StyleRazan, M. R., Amissi, S., Islam, R. A., Graham, J. L., Stanhope, K. L., Havel, P. J., & Rahimian, R. (2023). Moderate-Intensity Exercise Improves Mesenteric Arterial Function in Male UC Davis Type-2 Diabetes Mellitus (UCD-T2DM) Rats: A Shift in the Relative Importance of Endothelium-Derived Relaxing Factors (EDRF). Biomedicines, 11(4), 1129. https://doi.org/10.3390/biomedicines11041129