

Mechanisms of Action and Limitations of Monoclonal Antibodies and Single Chain Fragment Variable (scFv) in the Treatment of Cancer
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Cancer Overview
3. Monoclonal Antibodies: Structure and Function
3.1. Monoclonal Antibodies Production Methods
3.2. Monoclonal Antibodies in Cancer Treatment
3.2.1. Effector Mechanisms of mAb in Therapy
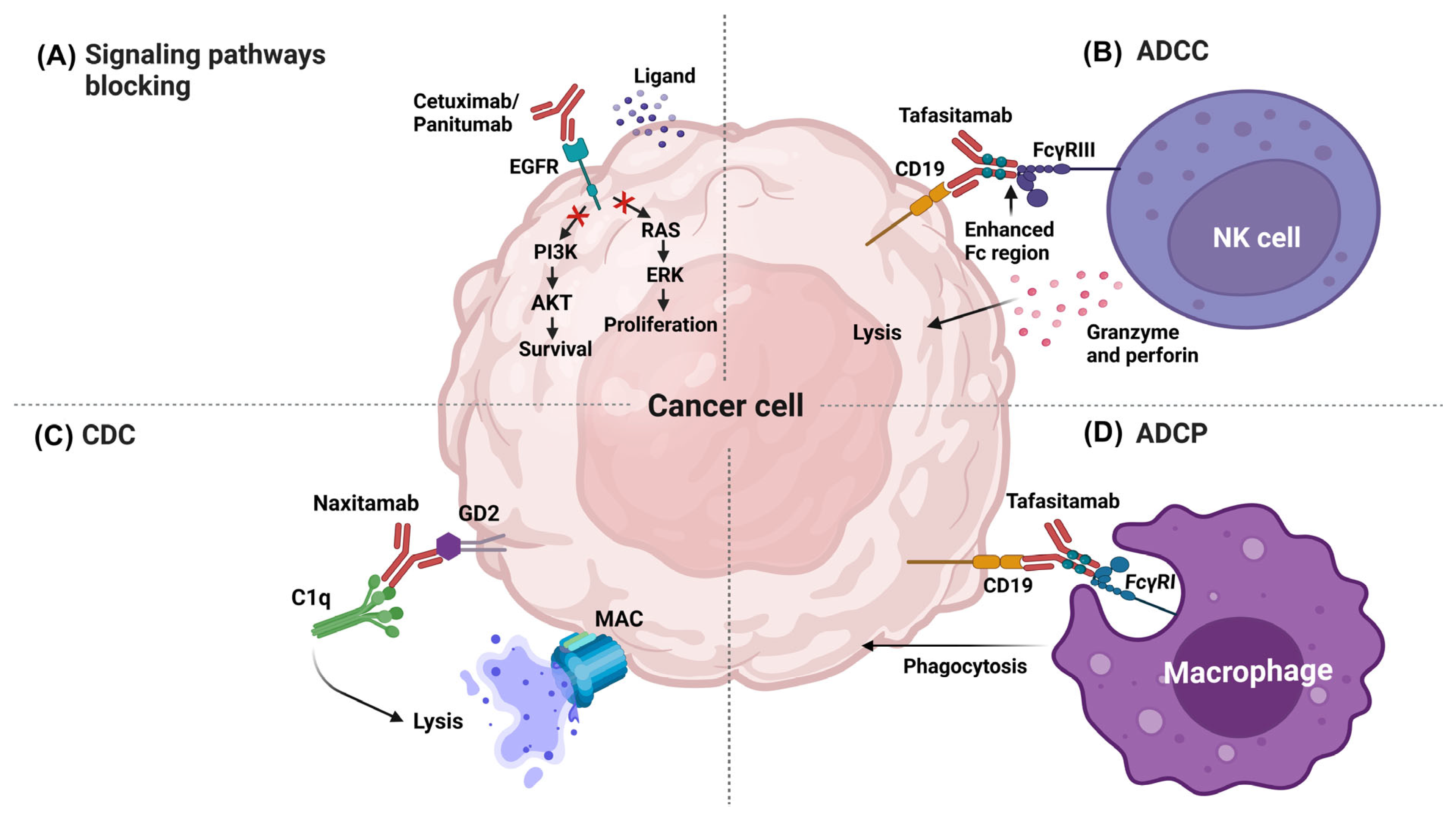
Blocking Signaling Pathways
Antibody-Dependent Cellular Cytotoxicity
Complement-Dependent Cytotoxicity
Antibody-Dependent Cellular Phagocytosis
3.2.2. Conjugated Antibodies
3.2.3. Disadvantages of mAb-Based Therapy
4. Single-Chain Variable Fragments (scFv)
4.1. Structure and Function
4.2. Mechanisms of Action of scFv in Cancer Therapy
4.2.1. T-Cell—Engaging CD3-Bispecific scFv Antibodies
4.2.2. Toxin-Conjugate scFv
4.2.3. Chimeric Antigen Receptor (CAR) T-Cells
4.2.4. Nanoparticle-Conjugate scFv
4.2.5. Blockade of Signaling Pathways and Biological Activity
4.3. In Situ Delivery of scFv Using Vectors
4.4. Advantages and Limitations of scFv
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Casals, E.; Gusta, M.F.; Cobaleda-Siles, M.; Garcia-Sanz, A.; Puntes, V.F. Cancer Resistance to Treatment and Antiresistance Tools Offered by Multimodal Multifunctional Nanoparticles. Cancer Nanotechnol. 2017, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Malone, E.R.; Oliva, M.; Sabatini, P.J.B.; Stockley, T.L.; Siu, L.L. Molecular Profiling for Precision Cancer Therapies. Genome Med. 2020, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Zahavi, D.; Weiner, L. Monoclonal Antibodies in Cancer Therapy. Antibodies 2020, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Vallina, L. Anticuerpos Monoclonales. Realidades y Perspectivas; Editorial Complutense: Madrid, Spain, 2004; ISBN 978-8-47491-759-8. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular Principles of Metastasis: A Hallmark of Cancer Revisited. Signal. Transduct. Target. Ther. 2020, 5, 28. [Google Scholar] [CrossRef]
- Cao, Y. Tumorigenesis as a Process of Gradual Loss of Original Cell Identity and Gain of Properties of Neural Precursor/Progenitor Cells. Cell Biosci. 2017, 7, 61. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Majcherek, D.; Weresa, M.A.; Ciecierski, C. A Cluster Analysis of Risk Factors for Cancer across EU Countries: Health Policy Recommendations for Prevention. Int. J. Environ. Res. Public. Health 2021, 18, 8142. [Google Scholar] [CrossRef]
- Barbet, J.; Bardiès, M.; Bourgeois, M.; Chatal, J.-F.; Chérel, M.; Davodeau, F.; Faivre-Chauvet, A.; Gestin, J.-F.; Kraeber-Bodéré, F. Radiolabeled Antibodies for Cancer Imaging and Therapy. Methods Mol. Biol. 2012, 907, 681–697. [Google Scholar] [CrossRef]
- Bournazos, S.; Ravetch, J.V. Fcγ Receptor Pathways during Active and Passive Immunization. Immunol. Rev. 2015, 268, 88–103. [Google Scholar] [CrossRef]
- Köhler, G.; Milstein, C. Derivation of Specific Antibody-Producing Tissue Culture and Tumor Lines by Cell Fusion. Eur. J. Immunol. 1976, 6, 511–519. [Google Scholar] [CrossRef]
- Holzlöhner, P.; Hanack, K. Generation of Murine Monoclonal Antibodies by Hybridoma Technology. J. Vis. Exp. 2017, 119, 54832. [Google Scholar] [CrossRef]
- Waldmann, H. Human Monoclonal Antibodies: The Benefits of Humanization. In Human Monoclonal Antibodies: Methods and Protocols; Steinitz, M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; pp. 1–10. ISBN 978-1-49398-958-4. [Google Scholar]
- Zhang, J.-H.; Shan, L.-L.; Liang, F.; Du, C.-Y.; Li, J.-J. Strategies and Considerations for Improving Recombinant Antibody Production and Quality in Chinese Hamster Ovary Cells. Front. Bioeng. Biotechnol. 2022, 10, 856049. [Google Scholar] [CrossRef]
- Carrara, S.C.; Fiebig, D.; Bogen, J.P.; Grzeschik, J.; Hock, B.; Kolmar, H. Recombinant Antibody Production Using a Dual-Promoter Single Plasmid System. Antibodies 2021, 10, 18. [Google Scholar] [CrossRef]
- Jyothilekshmi, I.; Jayaprakash, N.S. Trends in Monoclonal Antibody Production Using Various Bioreactor Systems. J. Microbiol. Biotechnol. 2021, 31, 349–357. [Google Scholar] [CrossRef]
- Rogers, L.M.; Veeramani, S.; Weiner, G.J. Complement in Monoclonal Antibody Therapy of Cancer. Immunol. Res. 2014, 59, 203–210. [Google Scholar] [CrossRef]
- Pardoll, D.M. The Blockade of Immune Checkpoints in Cancer Immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef]
- McLaughlin, P.; Grillo-López, A.J.; Link, B.K.; Levy, R.; Czuczman, M.S.; Williams, M.E.; Heyman, M.R.; Bence-Bruckler, I.; White, C.A.; Cabanillas, F.; et al. Rituximab Chimeric Anti-CD20 Monoclonal Antibody Therapy for Relapsed Indolent Lymphoma: Half of Patients Respond to a Four-Dose Treatment Program. J. Clin. Oncol. 1998, 16, 2825–2833. [Google Scholar] [CrossRef]
- Bar, L.; Nguyen, C.; Galibert, M.; Santos-Schneider, F.; Aldrian, G.; Dejeu, J.; Lartia, R.; Coche-Guérente, L.; Molina, F.; Boturyn, D. Determination of the Rituximab Binding Site to the CD20 Epitope Using SPOT Synthesis and Surface Plasmon Resonance Analyses. Anal. Chem. 2021, 93, 6865–6872. [Google Scholar] [CrossRef]
- Asano, T.; Suzuki, H.; Kaneko, M.K.; Kato, Y. Epitope Mapping of Rituximab Using HisMAP Method. Monoclon. Antibodies Immunodiagn. Immunother. 2022, 41, 8–14. [Google Scholar] [CrossRef]
- Casan, J.M.L.; Wong, J.; Northcott, M.J.; Opat, S. Anti-CD20 Monoclonal Antibodies: Reviewing a Revolution. Hum. Vaccin. Immunother. 2018, 14, 2820–2841. [Google Scholar] [CrossRef] [PubMed]
- FDA—Food & Drug Administration. FDA Grants Accelerated Approval to Sacituzumab Govitecan for Advanced Urothelial Cancer. 2021. Available online: https://www.fda.gov/drugs/resources-information-approved-drugs/fda-grants-accelerated-approval-sacituzumab-govitecan-advanced-urothelial-cancer (accessed on 20 January 2023).
- FDA—Food & Drug Administration. FDA Grants Regular Approval to Sacituzumab Govitecan for Triple-Negative Breast Cancer. 2021. Available online: https://www.fda.gov/drugs/resources-information-approved-drugs/fda-grants-regular-approval-sacituzumab-govitecan-triple-negative-breast-cancer (accessed on 20 January 2023).
- FDA—Food & Drug Administration. FDA Approves Sacituzumab Govitecan-Hziy for HR-Positive Breast Cancer. 2023. Available online: https://www.fda.gov/drugs/resources-information-approved-drugs/fda-approves-sacituzumab-govitecan-hziy-hr-positive-breast-cancer (accessed on 20 January 2023).
- FDA—Food & Drug Administration. FDA Approves Atezolizumab Plus Bevacizumab for Unresectable Hepatocellular Carcinoma. 2021. Available online: https://www.fda.gov/drugs/resources-information-approved-drugs/fda-approves-atezolizumab-plus-bevacizumab-unresectable-hepatocellular-carcinoma (accessed on 20 January 2023).
- Koh, J.; Nam, S.K.; Lee, Y.W.; Kim, J.W.; Lee, K.-W.; Ock, C.-Y.; Oh, D.-Y.; Ahn, S.-H.; Kim, H.-H.; Kang, K.-W.; et al. Trastuzumab Specific Epitope Evaluation as a Predictive and Prognostic Biomarker in Gastric Cancer Patients. Biomolecules 2019, 9, 782. [Google Scholar] [CrossRef] [PubMed]
- Holgate, R.G.E.; Weldon, R.; Jones, T.D.; Baker, M.P. Characterisation of a Novel Anti-CD52 Antibody with Improved Efficacy and Reduced Immunogenicity. PLoS ONE 2015, 10, e0138123. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Lammens, A.; Schäfer, W.; Georges, G.; Schwaiger, M.; Mössner, E.; Hopfner, K.-P.; Umaña, P.; Niederfellner, G. Epitope Interactions of Monoclonal Antibodies Targeting CD20 and Their Relationship to Functional Properties. MAbs 2013, 5, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Wang, Z.; Fan, J.; Bai, X.; Xu, Y.; Chou, J.J.; Hou, T.; Chen, S.; Pan, L. Structure-Guided and Phage-Assisted Evolution of a Therapeutic Anti-EGFR Antibody to Reverse Acquired Resistance. Nat. Commun. 2022, 13, 4431. [Google Scholar] [CrossRef]
- Wentink, M.Q.; Hackeng, T.M.; Tabruyn, S.P.; Puijk, W.C.; Schwamborn, K.; Altschuh, D.; Meloen, R.H.; Schuurman, T.; Griffioen, A.W.; Timmerman, P. Targeted Vaccination against the Bevacizumab Binding Site on VEGF Using 3D-Structured Peptides Elicits Efficient Antitumor Activity. Proc. Natl. Acad. Sci. USA 2016, 113, 12532–12537. [Google Scholar] [CrossRef]
- García-Foncillas, J.; Sunakawa, Y.; Aderka, D.; Wainberg, Z.; Ronga, P.; Witzler, P.; Stintzing, S. Distinguishing Features of Cetuximab and Panitumumab in Colorectal Cancer and Other Solid Tumors. Front. Oncol. 2019, 9, 849. [Google Scholar] [CrossRef]
- Cotchett, K.R.; Dittel, B.N.; Obeidat, A.Z. Comparison of the Efficacy and Safety of Anti-CD20 B Cells Depleting Drugs in Multiple Sclerosis. Mult. Scler. Relat. Disord. 2021, 49, 102787. [Google Scholar] [CrossRef]
- Gao, H.; Cai, H.; Liu, J.; Wang, X.; Zheng, P.; Devenport, M.; Xu, T.; Dou, F.; Liu, Y.; Zhou, A. Structure of CTLA-4 Complexed with a PH-Sensitive Cancer Immunotherapeutic Antibody. Cell Discov. 2020, 6, 79. [Google Scholar] [CrossRef]
- Deng, C.; Pan, B.; O’Connor, O.A. Brentuximab Vedotin. Clin. Cancer Res. 2013, 19, 22–27. [Google Scholar] [CrossRef]
- Wang, H.; Wang, Y.; Xiao, Z.; Li, W.; Dimitrov, D.S.; Chen, W. Human Domain Antibodies to Conserved Epitopes on HER2 Potently Inhibit Growth of HER2-Overexpressing Human Breast Cancer Cells In Vitro. Antibodies 2019, 8, 25. [Google Scholar] [CrossRef]
- Lu, R.-M.; Chiu, C.-Y.; Liu, I.-J.; Chang, Y.-L.; Liu, Y.-J.; Wu, H.-C. Novel Human Ab against Vascular Endothelial Growth Factor Receptor 2 Shows Therapeutic Potential for Leukemia and Prostate Cancer. Cancer Sci. 2019, 110, 3773–3787. [Google Scholar] [CrossRef]
- Teplyakov, A.; Obmolova, G.; Luo, J.; Gilliland, G.L. Crystal Structure of B-Cell Co-Receptor CD19 in Complex with Antibody B43 Reveals an Unexpected Fold. Proteins 2018, 86, 495–500. [Google Scholar] [CrossRef]
- Zhang, M.M.; Huang, R.Y.-C.; Beno, B.R.; Deyanova, E.G.; Li, J.; Chen, G.; Gross, M.L. Epitope and Paratope Mapping of PD-1/Nivolumab by Mass Spectrometry-Based Hydrogen/Deuterium Exchange, Cross-Linking, and Molecular Docking. Anal. Chem. 2020, 92, 9086–9094. [Google Scholar] [CrossRef] [PubMed]
- Córdova-Bahena, L.; Velasco-Velázquez, M.A. Anti-PD-1 And Anti-PD-L1 Antibodies as Immunotherapy Against Cancer: A Structural Perspective. RIC 2021, 73, 5139. [Google Scholar] [CrossRef]
- Makabe, K.; Yokoyama, T.; Uehara, S.; Uchikubo-Kamo, T.; Shirouzu, M.; Kimura, K.; Tsumoto, K.; Asano, R.; Tanaka, Y.; Kumagai, I. Anti-EGFR Antibody 528 Binds to Domain III of EGFR at a Site Shifted from the Cetuximab Epitope. Sci. Rep. 2021, 11, 5790. [Google Scholar] [CrossRef]
- Zirngibl, F.; Ivasko, S.M.; Grunewald, L.; Klaus, A.; Schwiebert, S.; Ruf, P.; Lindhofer, H.; Astrahantseff, K.; Andersch, L.; Schulte, J.H.; et al. GD2-Directed Bispecific Trifunctional Antibody Outperforms Dinutuximab Beta in a Murine Model for Aggressive Metastasized Neuroblastoma. J. Immunother. Cancer 2021, 9, e002923. [Google Scholar] [CrossRef]
- Lee, H.T.; Kim, Y.; Park, U.B.; Jeong, T.J.; Lee, S.H.; Heo, Y.-S. Crystal Structure of CD38 in Complex with Daratumumab, a First-in-Class Anti-CD38 Antibody Drug for Treating Multiple Myeloma. Biochem. Biophys. Res. Commun. 2021, 536, 26–31. [Google Scholar] [CrossRef]
- Ishibashi, M.; Morita, R.; Tamura, H. Immune Functions of Signaling Lymphocytic Activation Molecule Family Molecules in Multiple Myeloma. Cancers 2021, 13, 279. [Google Scholar] [CrossRef]
- Moroncini, G.; Maccaroni, E.; Fiordoliva, I.; Pellei, C.; Gabrielli, A.; Berardi, R. Developments in the Management of Advanced Soft-Tissue Sarcoma—Olaratumab in Context. Oncotargets Ther. 2018, 11, 833–842. [Google Scholar] [CrossRef]
- Bannister, D.; Popovic, B.; Sridharan, S.; Giannotta, F.; Filée, P.; Yilmaz, N.; Minter, R. Epitope Mapping and Key Amino Acid Identification of Anti-CD22 Immunotoxin CAT-8015 Using Hybrid β-Lactamase Display. Protein Eng. Des. Sel. 2011, 24, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Fenwarth, L.; Fournier, E.; Cheok, M.; Boyer, T.; Gonzales, F.; Castaigne, S.; Boissel, N.; Lambert, J.; Dombret, H.; Preudhomme, C.; et al. Biomarkers of Gemtuzumab Ozogamicin Response for Acute Myeloid Leukemia Treatment. Int. J. Mol. Sci. 2020, 21, 5626. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Xu, Z.; Zhang, D.; Jiang, M.; Liu, K.; He, J.; Ma, D.; Ma, X.; Tan, S.; Gao, G.F.; et al. PD-1 N58-Glycosylation-Dependent Binding of Monoclonal Antibody Cemiplimab for Immune Checkpoint Therapy. Front. Immunol. 2022, 13, 826045. [Google Scholar] [CrossRef]
- Li, D.; Lee, D.; Dere, R.C.; Zheng, B.; Yu, S.; Fuh, F.K.; Kozak, K.R.; Chung, S.; Bumbaca Yadav, D.; Nazzal, D.; et al. Evaluation and Use of an Anti-cynomolgus Monkey CD79b Surrogate Antibody–Drug Conjugate to Enable Clinical Development of Polatuzumab Vedotin. Br. J. Pharmacol. 2019, 176, 3805–3818. [Google Scholar] [CrossRef]
- Jain, R.K.; Skelton, W.P.; Zhang, J. Emerging Treatment Options for the Treatment of Metastatic Urothelial Cancer: Therapeutic Potential of Enfortumab Vedotin. Cancer Manag. Res. 2020, 12, 8379–8386. [Google Scholar] [CrossRef]
- Sun, M.; Zhang, H.; Jiang, M.; Chai, Y.; Qi, J.; Gao, G.F.; Tan, S. Structural Insights into the Cis and Trans Assembly of Human Trophoblast Cell Surface Antigen 2. iScience 2021, 24, 103190. [Google Scholar] [CrossRef]
- Düll, J.; Topp, M.; Salles, G. The Use of Tafasitamab in Diffuse Large B-Cell Lymphoma. Ther. Adv. Hematol. 2021, 12, 20406207211027458. [Google Scholar] [CrossRef]
- Offidani, M.; Corvatta, L.; Morè, S.; Olivieri, A. Belantamab Mafodotin for the Treatment of Multiple Myeloma: An Overview of the Clinical Efficacy and Safety. Drug. Des. Dev. Ther. 2021, 15, 2401–2415. [Google Scholar] [CrossRef]
- Chan, G.C.-F.; Chan, C.M. Anti-GD2 Directed Immunotherapy for High-Risk and Metastatic Neuroblastoma. Biomolecules 2022, 12, 358. [Google Scholar] [CrossRef]
- Furqan, F.; Hamadani, M. Loncastuximab Tesirine in Relapsed or Refractory Diffuse Large B-Cell Lymphoma: A Review of Clinical Data. Ther. Adv. Hematol. 2022, 13, 20406207221087511. [Google Scholar] [CrossRef]
- Neijssen, J.; Cardoso, R.M.F.; Chevalier, K.M.; Wiegman, L.; Valerius, T.; Anderson, G.M.; Moores, S.L.; Schuurman, J.; Parren, P.W.H.I.; Strohl, W.R.; et al. Discovery of Amivantamab (JNJ-61186372), a Bispecific Antibody Targeting EGFR and MET. J. Biol. Chem. 2021, 296, 100641. [Google Scholar] [CrossRef] [PubMed]
- Jeong, T.-J.; Lee, H.-T.; Gu, N.; Jang, Y.-J.; Choi, S.-B.; Park, U.-B.; Lee, S.-H.; Heo, Y.-S. The High-Resolution Structure Reveals Remarkable Similarity in PD-1 Binding of Cemiplimab and Dostarlimab, the FDA-Approved Antibodies for Cancer Immunotherapy. Biomedicines 2022, 10, 3154. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Li, S.; Han, S.; Shi, C.; Zhang, Y. Antibody Drug Conjugate: The “Biological Missile” for Targeted Cancer Therapy. Signal Transduct. Target. Ther. 2022, 7, 93. [Google Scholar] [CrossRef] [PubMed]
- Pillarisetti, K.; Powers, G.; Luistro, L.; Babich, A.; Baldwin, E.; Li, Y.; Zhang, X.; Mendonça, M.; Majewski, N.; Nanjunda, R.; et al. Teclistamab Is an Active T Cell–Redirecting Bispecific Antibody against B-Cell Maturation Antigen for Multiple Myeloma. Blood Adv. 2020, 4, 4538–4549. [Google Scholar] [CrossRef] [PubMed]
- Hartnett, E.G.; Knight, J.; Radolec, M.; Buckanovich, R.J.; Edwards, R.P.; Vlad, A.M. Immunotherapy Advances for Epithelial Ovarian Cancer. Cancers 2020, 12, 3733. [Google Scholar] [CrossRef]
- Kang, C. Mosunetuzumab: First Approval. Drugs 2022, 82, 1229–1234. [Google Scholar] [CrossRef]
- Tintelnot, J.; Baum, N.; Schultheiß, C.; Braig, F.; Trentmann, M.; Finter, J.; Fumey, W.; Bannas, P.; Fehse, B.; Riecken, K.; et al. Nanobody Targeting of Epidermal Growth Factor Receptor (EGFR) Ectodomain Variants Overcomes Resistance to Therapeutic EGFR Antibodies. Mol. Cancer Ther. 2019, 18, 823–833. [Google Scholar] [CrossRef]
- Arena, S.; Bellosillo, B.; Siravegna, G.; Martínez, A.; Cañadas, I.; Lazzari, L.; Ferruz, N.; Russo, M.; Misale, S.; González, I.; et al. Emergence of Multiple EGFR Extracellular Mutations during Cetuximab Treatment in Colorectal Cancer. Clin. Cancer Res. 2015, 21, 2157–2166. [Google Scholar] [CrossRef]
- Braig, F.; März, M.; Schieferdecker, A.; Schulte, A.; Voigt, M.; Stein, A.; Grob, T.; Alawi, M.; Indenbirken, D.; Kriegs, M.; et al. Epidermal Growth Factor Receptor Mutation Mediates Cross-Resistance to Panitumumab and Cetuximab in Gastrointestinal Cancer. Oncotarget 2015, 6, 12035–12047. [Google Scholar] [CrossRef]
- Bagchi, A.; Haidar, J.N.; Eastman, S.W.; Vieth, M.; Topper, M.; Iacolina, M.D.; Walker, J.M.; Forest, A.; Shen, Y.; Novosiadly, R.D.; et al. Molecular Basis for Necitumumab Inhibition of EGFR Variants Associated with Acquired Cetuximab Resistance. Mol. Cancer Ther. 2018, 17, 521–531. [Google Scholar] [CrossRef]
- Yu, J.; Song, Y.; Tian, W. How to Select IgG Subclasses in Developing Anti-Tumor Therapeutic Antibodies. J. Hematol. Oncol. 2020, 13, 45. [Google Scholar] [CrossRef]
- Shuptrine, C.; Surana, R.; Weiner, L.M. Monoclonal Antibodies for the Treatment of Cancer. Semin. Cancer Biol. 2012, 22, 3–13. [Google Scholar] [CrossRef]
- Ma, H.; O’Kennedy, R. The Structure of Natural and Recombinant Antibodies. In Peptide Antibodies: Methods and Protocols; Houen, G., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; pp. 7–11. ISBN 978-1-49392-999-3. [Google Scholar]
- Rajaram, P.; Chandra, P.; Ticku, S.; Pallavi, B.K.; Rudresh, K.B.; Mansabdar, P. Epidermal Growth Factor Receptor: Role in Human Cancer. Indian Journal. Dent. Res. 2017, 28, 687. [Google Scholar] [CrossRef]
- Liu, X.; Wang, P.; Zhang, C.; Ma, Z. Epidermal growth factor receptor (EGFR): A rising star in the era of precision medicine of lung cancer. Oncotarget 2017, 8, 50209–50220. [Google Scholar] [CrossRef]
- Chidharla, A.; Parsi, M.; Kasi, A. Cetuximab [Updated 2022 May 8]. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK459293/ (accessed on 20 January 2023).
- Dubois, E.A.; Cohen, A.F. Panitumumab. Br. J. Clin. Pharmacol. 2009, 68, 482–483. [Google Scholar] [CrossRef]
- Sulica, A.; Morel, R.; Metes, D.; Herberman, R.B. Ig-Binding Receptors on Human NK Cells as Effector and Regulatory Surface Molecules. Int. Rev. Immunol. 2001, 20, 371–414. [Google Scholar] [CrossRef]
- Zinzani, P.L.; Minotti, G. Anti-CD19 Monoclonal Antibodies for the Treatment of Relapsed or Refractory B-Cell Malignancies: A Narrative Review with Focus on Diffuse Large B-Cell Lymphoma. J. Cancer Res. Clin. Oncol. 2022, 148, 177–190. [Google Scholar] [CrossRef]
- Salles, G.; Długosz-Danecka, M.; Ghesquières, H.; Jurczak, W. Tafasitamab for the Treatment of Relapsed or Refractory Diffuse Large B-Cell Lymphoma. Expert. Opin. Biol. Ther. 2021, 21, 455–463. [Google Scholar] [CrossRef]
- Golay, J.; Taylor, R.P. The Role of Complement in the Mechanism of Action of Therapeutic Anti-Cancer MAbs. Antibodies 2020, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Salles, G.; Barrett, M.; Foà, R.; Maurer, J.; O’Brien, S.; Valente, N.; Wenger, M.; Maloney, D.G. Rituximab in B-Cell Hematologic Malignancies: A Review of 20 Years of Clinical Experience. Adv. Ther. 2017, 34, 2232–2273. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.P.; Lindorfer, M.A. Cytotoxic Mechanisms of Immunotherapy: Harnessing Complement in the Action of Anti-Tumor Monoclonal Antibodies. Semin. Immunol. 2016, 28, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Kamen, L.; Myneni, S.; Langsdorf, C.; Kho, E.; Ordonia, B.; Thakurta, T.; Zheng, K.; Song, A.; Chung, S. A Novel Method for Determining Antibody-Dependent Cellular Phagocytosis. J. Immunol. Methods 2019, 468, 55–60. [Google Scholar] [CrossRef]
- Nooka, A.K.; Kaufman, J.L.; Hofmeister, C.C.; Joseph, N.S.; Heffner, T.L.; Gupta, V.A.; Sullivan, H.C.; Neish, A.S.; Dhodapkar, M.V.; Lonial, S. Daratumumab in Multiple Myeloma. Cancer 2019, 125, 2364–2382. [Google Scholar] [CrossRef]
- Thomas, A.; Teicher, B.A.; Hassan, R. Antibody–Drug Conjugates for Cancer Therapy. Lancet Oncol. 2016, 17, e254–e262. [Google Scholar] [CrossRef]
- Coleman, R.L.; Lorusso, D.; Gennigens, C.; González-Martín, A.; Randall, L.; Cibula, D.; Lund, B.; Woelber, L.; Pignata, S.; Forget, F.; et al. Efficacy and Safety of Tisotumab Vedotin in Previously Treated Recurrent or Metastatic Cervical Cancer (InnovaTV 204/GOG-3023/ENGOT-Cx6): A Multicentre, Open-Label, Single-Arm, Phase 2 Study. Lancet Oncol. 2021, 22, 609–619. [Google Scholar] [CrossRef]
- Torka, P.; Barth, M.; Ferdman, R.; Hernandez-Ilizaliturri, F.J. Mechanisms of Resistance to Monoclonal Antibodies (MAbs) in Lymphoid Malignancies. Curr. Hematol. Malig. Rep. 2019, 14, 426–438. [Google Scholar] [CrossRef]
- Reslan, L.; Dalle, S.; Dumontet, C. Understanding and Circumventing Resistance to Anticancer Monoclonal Antibodies. MAbs 2009, 1, 222–229. [Google Scholar] [CrossRef]
- Wallace, Z.S.; Harkness, T.; Blumenthal, K.G.; Choi, H.K.; Stone, J.H.; Walensky, R.P. Increasing Operational Capacity and Reducing Costs of Rituximab Administration: A Costing Analysis. ACR Open. Rheumatol. 2020, 2, 261–268. [Google Scholar] [CrossRef]
- Ahmadzadeh, V.; Farajnia, S.; Feizi, M.A.H.; Nejad, R.A.K. Antibody Humanization Methods for Development of Therapeutic Applications. Monoclon. Antibodies Immunodiagn. Immunother. 2014, 33, 67–73. [Google Scholar] [CrossRef]
- Winter, G.; Milstein, C. Man-Made Antibodies. Nature 1991, 349, 293–299. [Google Scholar] [CrossRef]
- Chames, P.; Baty, D. Antibody Engineering and Its Applications in Tumor Targeting and Intracellular Immunization. FEMS Microbiol. Lett. 2000, 189, 1–8. [Google Scholar] [CrossRef]
- Brofelth, M.; Städe, L.W.; Ekstrand, A.I.; Edfeldt, L.P.; Kovačič, R.; Nielsen, T.T.; Larsen, K.L.; Duroux, L.; Wingren, C. Site-Specific Photocoupling of PBpa Mutated ScFv Antibodies for Use in Affinity Proteomics. Biochim. Et Biophys. Acta (BBA)-Proteins Proteom. 2017, 1865, 985–996. [Google Scholar] [CrossRef]
- Gu, X.; Jia, X.; Feng, J.; Shen, B.; Huang, Y.; Geng, S.; Sun, Y.; Wang, Y.; Li, Y.; Long, M. Molecular Modeling and Affinity Determination of ScFv Antibody: Proper Linker Peptide Enhances Its Activity. Ann. Biomed. Eng. 2010, 38, 537–549. [Google Scholar] [CrossRef]
- Chen, X.; Zaro, J.; Shen, W.-C. Fusion Protein Linkers: Property, Design and Functionality. Adv. Drug. Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef]
- Li, Z.; Krippendorff, B.-F.; Sharma, S.; Walz, A.C.; Lavé, T.; Shah, D.K. Influence of Molecular Size on Tissue Distribution of Antibody Fragments. MAbs 2016, 8, 113–119. [Google Scholar] [CrossRef]
- Kang, T.H.; Seong, B.L. Solubility, Stability, and Avidity of Recombinant Antibody Fragments Expressed in Microorganisms. Front. Microbiol. 2020, 11, 1927. [Google Scholar] [CrossRef]
- Sandomenico, A.; Sivaccumar, J.P.; Ruvo, M. Evolution of Escherichia Coli Expression System in Producing Antibody Recombinant Fragments. Int. J. Mol. Sci. 2020, 21, 6324. [Google Scholar] [CrossRef]
- Jefferis, R. Recombinant Antibody Therapeutics: The Impact of Glycosylation on Mechanisms of Action. Trends Pharmacol. Sci. 2009, 30, 356–362. [Google Scholar] [CrossRef]
- Spadiut, O.; Capone, S.; Krainer, F.; Glieder, A.; Herwig, C. Microbials for the Production of Monoclonal Antibodies and Antibody Fragments. Trends Biotechnol. 2014, 32, 54–60. [Google Scholar] [CrossRef]
- Pan, K.; Wang, H.; Zhang, H.; Liu, H.; Lei, H.; Huang, L.; Sun, Y. Production and Characterization of Single Chain Fv Directed against Β2-Agonist Clenbuterol. J. Agric. Food Chem. 2006, 54, 6654–6659. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, H.C.; Duan, C.F.; Dong, J.; Zhao, G.X.; Wang, J.P.; Li, N.; Liu, J.Z.; Li, Y.W. Production of Anti-Amoxicillin ScFv Antibody and Simulation Studying Its Molecular Recognition Mechanism for Penicillins. J. Environ. Sci. Health B 2016, 51, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, S.; Kobashigawa, Y.; Fukuda, N.; Teramoto, M.; Toyota, Y.; Liu, C.; Ikeguchi, Y.; Sato, T.; Sato, Y.; Kimura, H.; et al. Cyclization of Single-Chain Fv Antibodies Markedly Suppressed Their Characteristic Aggregation Mediated by Inter-Chain VH-VL Interactions. Molecules 2019, 24, 2620. [Google Scholar] [CrossRef] [PubMed]
- Bates, A.; Power, C.A. David vs. Goliath: The Structure, Function, and Clinical Prospects of Antibody Fragments. Antibodies 2019, 8, 28. [Google Scholar] [CrossRef] [PubMed]
- Przepiorka, D.; Ko, C.-W.; Deisseroth, A.; Yancey, C.L.; Candau-Chacon, R.; Chiu, H.-J.; Gehrke, B.J.; Gomez-Broughton, C.; Kane, R.C.; Kirshner, S.; et al. FDA Approval: Blinatumomab. Clin. Cancer Res. 2015, 21, 4035–4039. [Google Scholar] [CrossRef]
- Iizuka, A.; Nonomura, C.; Ashizawa, T.; Kondou, R.; Ohshima, K.; Sugino, T.; Mitsuya, K.; Hayashi, N.; Nakasu, Y.; Maruyama, K.; et al. A T-Cell–Engaging B7-H4/CD3-Bispecific Fab-ScFv Antibody Targets Human Breast Cancer. Clin. Cancer Res. 2019, 25, 2925–2934. [Google Scholar] [CrossRef]
- Hall, W.A.; Fodstad, Ø. Immunotoxins and Central Nervous System Neoplasia. J. Neurosurg. 1992, 76, 1–12. [Google Scholar] [CrossRef]
- Dhillon, S. Moxetumomab Pasudotox: First Global Approval. Drugs 2018, 78, 1763–1767. [Google Scholar] [CrossRef]
- Philogen S.p.A. A Dose Finding Pharmacokinetic Study of the Tumour-Targeting Human L19IL2 Monoclonal Antibody-Cytokine Fusion Protein in Patients with Advanced Solid Tumours. Available online: https://clinicaltrials.gov/ct2/show/NCT02957019 (accessed on 20 January 2023).
- Uppsala University. CD19-Targeting 3rd Generation CAR T Cells for Refractory B Cell Malignancy—A Phase I/IIa Trial. Available online: https://clinicaltrials.gov/ct2/show/NCT02132624 (accessed on 20 January 2023).
- Xu, K.L.; Zheng, J.N. Humanized CAR-T Therapy for Treatment of Recurrent or Refractory B Cell Malignancy by Targeting CD19. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02782351 (accessed on 20 January 2023).
- University of Pennsylvania. Pilot Study of Redirected Autologous T Cells Engineered to Contain Anti-CD19 Attached to TCRζ and 4-1BB Signaling Domains Coupled with Salvage Autologous Stem-Cell Transplantation (ASCT) In Multiple Myeloma Patients with Early Relapse/Progression after Initial ASCT. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02135406 (accessed on 20 January 2023).
- Zhou, M.-T. Pilot Study of Autologous Chimeric Antigen Receptor Cells against Mesothelin and CD19 in Patients with Pancreatic Cancer. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03497819 (accessed on 20 January 2023).
- Shenzhen Second People’s Hospital the Study of Autologous T Cells Expressing CD19 Chimeric Antigen Receptors Treatment of Minimal Residual Disease (MRD) of B Cell Malignancies and Then Autologous Hematopoietic Stem Cell Transplantation (Auto-HSCT). Available online: https://clinicaltrials.gov/ct2/show/study/NCT03685786 (accessed on 20 January 2023).
- University of Pennsylvania. Phase II Study of Redirected Autologous T Cells Engineered to Contain Anti-CD19 Attached to TCR and 4-1BB Signaling Domains in Patients with Chemotherapy Resistant or Refractory Acute Lymphoblastic Leukemia. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02030847 (accessed on 20 January 2023).
- Maastricht University. Stereotactic Ablative Body Radiotherapy (SABR) Combined with Immunotherapy (L19-IL2) in Stage IV NSCLC Patients, ImmunoSABR: A Multicentre, Randomised Controlled Open-Label Phase II Trial. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03705403 (accessed on 20 January 2023).
- Ligen, L. CD19-Targeting CAR T Cell Therapy in the Treatment of Relapsed or Refractory CD19 Positive B-Cell Malignancies. Available online: https://clinicaltrials.gov/ct2/show/NCT03559439 (accessed on 20 January 2023).
- Zhang, L. Safety and Toxicity of CAR-T Cell Immunotherapy in Patients with Advanced Lung Cancer after Standard Treatment Failure: A Single-Arm and Single-Center Phase I Clinical Study. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03330834 (accessed on 20 January 2023).
- PersonGen BioTherapeutics (Suzhou) Co., Ltd. CD276 CAR-T for Patients with Advanced CD276+ Solid Tumors. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04691713 (accessed on 20 January 2023).
- University of Pennsylvania. Pilot Study of Redirected Autologous T Cells Engineered to Contain an Anti-BCMA ScFv Coupled to TCRζ and 4-1BB Signaling Domains in Patients with Relapsed and/or Refractory Multiple Myeloma. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02546167 (accessed on 20 January 2023).
- PersonGen BioTherapeutics (Suzhou) Co., Ltd. Single Arm, Single Center, Open Label Clinical Trial of BCMA Autologous Chimeric Antigen Receptor T Cell Infusion in Patients with BCMA Positive Recurrent or Refractory Multiple Myeloma. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04650724 (accessed on 20 January 2023).
- University of Pennsylvania. Phase 2 Study of CD19-Directed Chimeric Antigen Receptor-Modified T Cells (CART19) for Adult Patients with Minimal Residual Disease during Upfront Treatment for Acute Lymphoblastic Leukemia. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02935543 (accessed on 20 January 2023).
- Zhang, Z.; Zhaotai, G. Engineered TILs/CAR-TILs to Treat Advanced Solid Tumors. Available online: https://clinicaltrials.gov/ct2/show/record/NCT04842812 (accessed on 25 August 2022).
- PersonGen BioTherapeutics (Suzhou) Co., Ltd. Chimeric Antigen Receptor T Cells Therapy for r/r Malignant Tumors in Children. Available online: https://clinicaltrials.gov/ct2/show/NCT04691349 (accessed on 20 January 2023).
- Cohen, R. Phase I Trial of GFRα4 CAR T Cells in Adult Patients with Recurrent or Metastatic Medullary Thyroid Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT04877613 (accessed on 20 January 2023).
- Shenzhen BinDeBio Ltd. Study of Autologous T-Cells Redirected to Mesothelin with a Chimeric Antigen Receptor in Patients with Metastatic Pancreatic Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT03638193 (accessed on 20 January 2023).
- Minerva Biotechnologies Corporation. Adoptive Immunotherapy for Advanced MUC1* Positive Breast Cancer with Autologous T Cells Engineered to Express a Chimeric Antigen Receptor, HuMNC2-CAR44 Specific for a Cleaved Form of MUC1 (MUC1*). Available online: https://clinicaltrials.gov/ct2/show/study/NCT04020575 (accessed on 20 January 2023).
- Second Affiliated Hospital of Guangzhou Medical University. Intratumor CpG-ODN Boosters Immune Killing against in Situ Tumor Antigen Released by Interventional Approaches for Advanced Solid Tumors. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04952272 (accessed on 20 January 2023).
- Second Affiliated Hospital of Guangzhou Medical University. CAR-T Cells Targeting PSCA, MUC1, TGFβ, HER2, Mesothelin, Lewis-Y, GPC3, AXL, EGFR, B7-H3 or Claudin18.2 for Immunotherapy of Lung Cancer: Phase I Clinical Trial. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03198052 (accessed on 20 January 2023).
- Salman, H. A Phase I, Multicenter Study of CD4- Directed Chimeric Antigen Receptor Engineered T-Cells (CD4CAR) in Patients with Relapsed or Refractory CD4+ Hematological Malignancies. Available online: https://www.clinicaltrials.gov/ct2/show/NCT03829540 (accessed on 20 January 2023).
- Nanjing IASO Biotherapeutics Co., Ltd. Phase 1/2 Clinical Study on Fully Human BCMA Chimeric Antigen Receptor Autologous T Cell Injection (CT103A) in the Treatment of Patients with Relapsed/Refractory Multiple Myeloma. Available online: https://clinicaltrials.gov/ct2/show/NCT05066646 (accessed on 20 January 2023).
- Tmunity Therapeutics. A Phase 1/2, Open-Label, Multi-Center Study of Dually Armored Chimeric Antigen Receptor (CAR) T-Cells (TmPSMA-02) in Patients with Metastatic Castration Resistant Prostate Cancer (MCRPC). Available online: https://clinicaltrials.gov/ct2/show/study/NCT05489991 (accessed on 20 January 2023).
- Second Affiliated Hospital of Guangzhou Medical University. Phase I/IIa Trial to Evaluate Safety and Preliminary Efficacy of CLDN6-CAR-NK in Patients with CLDN6-Positive Advanced Solid Tumors. Available online: https://clinicaltrials.gov/ct2/show/study/NCT05410717 (accessed on 20 January 2023).
- Shah, N. Phase 1/1b Study of Redirected Autologous T Cells Engineered to Contain an Anti CD19 and Anti CD20 ScFv Coupled to CD3ζ and 4-1BB Signaling Domains in Patients with Relapsed and/or Refractory CD19 or CD20 Positive B Cell Malignancies. Available online: https://clinicaltrials.gov/ct2/show/study/NCT03019055 (accessed on 20 January 2023).
- I-Mab Biopharma Co., Ltd. A Phase 1 Study of TJ033721 in Subjects with Advanced or Metastatic Solid Tumors. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04900818 (accessed on 20 January 2023).
- Bigner, D. Phase I Single-Center, Dose Escalation Study of D2C7-IT Administered Intratumorally via Convection-Enhanced Delivery for Adult Patients with Recurrent Malignant Glioma. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02303678 (accessed on 20 January 2023).
- Philogen S.p.A. A Study to Evaluate the Safety and Efficacy of the Tumor-Targeting Human Antibody-Cytokine Fusion Protein L19TNF Plus Lomustine in Patients with Glioblastoma at First Progression. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04573192 (accessed on 20 January 2023).
- Li, C. An Exploratory Study of Fully Human Anti-GPRC5D Chimeric Antigen Receptor T Cells (CAR-GPRC5D) in Patients with Relapsed/Refractory Multiple Myeloma or Plasma Cell Leukemia. Available online: https://clinicaltrials.gov/ct2/show/study/NCT05219721 (accessed on 20 January 2023).
- Duke University. A Phase I Study of HEGFRvIII-CD3 Bi-ScFv (BRiTE) in Patients with WHO Grade IV Malignant Glioma. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04903795 (accessed on 20 January 2023).
- University of Alberta. A Phase 1b/2 Multi-Center, De-Centralized, Dose Selection Study of Autologous CD19-Directed Chimeric Antigen Receptor (CAR) T-Cells in Patients with Relapsed/Refractory Aggressive Lymphoma or Acute Lymphoblastic Leukemia (ALL). Available online: https://clinicaltrials.gov/ct2/show/study/NCT03938987 (accessed on 20 January 2023).
- M.D. Anderson Cancer Center. Phase I Study of an Active Immunotherapy for Asymptomatic Phase Lymphoplasmacytic Lymphoma with DNA Vaccines Encoding Antigen-Chemokine Fusion. Available online: https://clinicaltrials.gov/ct2/show/study/NCT01209871 (accessed on 20 January 2023).
- University of Pennsylvania. Phase 1 Study of Lentivirally Transduced T Cells Engineered to Contain Anti-CD123 Linked to TCRζ and 4-1BB Signaling Domains in Pediatric Subjects with Refractory or Relapsed Acute Myeloid Leukemia. Available online: https://clinicaltrials.gov/ct2/show/study/NCT04678336 (accessed on 20 January 2023).
- He, S.; Zhao, C.; Tao, H.; Sheng, W.; Gao, R.; Liu, X.; Zhen, Y. A Recombinant ScFv Antibody-Based Fusion Protein That Targets EGFR Associated with IMPDH2 Downregulation and Its Drug Conjugate Show Therapeutic Efficacy against Esophageal Cancer. Drug Deliv. 2022, 29, 1243–1256. [Google Scholar] [CrossRef]
- Lee, H.H.; Kim, I.; Kim, U.K.; Choi, S.S.; Kim, T.Y.; Lee, D.; Lee, Y.; Lee, J.; Jo, J.; Lee, Y.-T.; et al. Therapeutic Effiacy of T Cells Expressing Chimeric Antigen Receptor Derived from a Mesothelin-Specific ScFv in Orthotopic Human Pancreatic Cancer Animal Models. Neoplasia 2021, 24, 98–108. [Google Scholar] [CrossRef]
- Zhu, H.; Fang, X.; Tuhin, I.J.; Tan, J.; Ye, J.; Jia, Y.; Xu, N.; Kang, L.; Li, M.; Lou, X.; et al. CAR T Cells Equipped with a Fully Human ScFv Targeting Trop2 Can Be Used to Treat Pancreatic Cancer. J. Cancer Res. Clin. Oncol. 2022, 148, 2261–2274. [Google Scholar] [CrossRef]
- Fu, J.; Shang, Y.; Qian, Z.; Hou, J.; Yan, F.; Liu, G.; Dehua, L.; Tian, X. Chimeric Antigen Receptor-T (CAR-T) Cells Targeting Epithelial Cell Adhesion Molecule (EpCAM) Can Inhibit Tumor Growth in Ovarian Cancer Mouse Model. J. Vet. Med. Sci. 2021, 83, 241–247. [Google Scholar] [CrossRef]
- Yang, M.; Tang, X.; Zhang, Z.; Gu, L.; Wei, H.; Zhao, S.; Zhong, K.; Mu, M.; Huang, C.; Jiang, C.; et al. Tandem CAR-T Cells Targeting CD70 and B7-H3 Exhibit Potent Preclinical Activity against Multiple Solid Tumors. Theranostics 2020, 10, 7622–7634. [Google Scholar] [CrossRef]
- Sauer, T.; Parikh, K.; Sharma, S.; Omer, B.; Sedloev, D.; Chen, Q.; Angenendt, L.; Schliemann, C.; Schmitt, M.; Müller-Tidow, C.; et al. CD70-Specific CAR T Cells Have Potent Activity against Acute Myeloid Leukemia without HSC Toxicity. Blood 2021, 138, 318–330. [Google Scholar] [CrossRef]
- Ochi, T.; Maruta, M.; Tanimoto, K.; Kondo, F.; Yamamoto, T.; Kurata, M.; Fujiwara, H.; Masumoto, J.; Takenaka, K.; Yasukawa, M. A Single-Chain Antibody Generation System Yielding CAR-T Cells with Superior Antitumor Function. Commun. Biol. 2021, 4, 273. [Google Scholar] [CrossRef]
- Wutti-In, Y.; Sujjitjoon, J.; Sawasdee, N.; Panya, A.; Kongkla, K.; Yuti, P.; Yongpitakwattana, P.; Thepmalee, C.; Junking, M.; Chieochansin, T.; et al. Development of a Novel Anti-CD19 CAR Containing a Fully Human ScFv and Three Costimulatory Domains. Front. Oncol. 2021, 11, 802876. [Google Scholar] [CrossRef]
- Li, Z.; Liu, C.; Li, C.; Wang, F.; Liu, J.; Zheng, Z.; Wu, J.; Zhang, B. Irinotecan/ScFv Co-Loaded Liposomes Coaction on Tumor Cells and CAFs for Enhanced Colorectal Cancer Therapy. J. Nanobiotechnology 2021, 19, 421. [Google Scholar] [CrossRef]
- Xu, W.; Liu, L.; Brown, N.J.; Christian, S.; Hornby, D. Quantum Dot- Conjugated Anti-GRP78 ScFv Inhibits Cancer Growth in Mice. Molecules 2012, 17, 796–808. [Google Scholar] [CrossRef]
- Darvishi, B.; Salehi, M.; Boroumandieh, S.; Majidzadeh-A, K.; Jalili, N.; Moradi-Kalbolandi, S.; Farahmand, L. Dual in Vitro Invasion/Migration Suppressing and Tamoxifen Response Modulating Effects of a Recombinant Anti-ALCAM ScFv on Breast Cancer Cells. Cell. Biochem. Funct. 2020, 38, 651–659. [Google Scholar] [CrossRef]
- Du, P.; Wang, X.; Yin, T.; Zhang, X.; Zhang, Z.; Yu, W.; Wang, M.; Luo, C.; Yu, L. Anti-Tumor Effect of Single-Chain Antibody to Reg3a in Colorectal Cancer. Exp. Cell Res. 2020, 396, 112278. [Google Scholar] [CrossRef]
- Yamada, S. Specific Functions of Exostosin-like 3 (EXTL3) Gene Products. Cell. Mol. Biol. Lett. 2020, 25, 39. [Google Scholar] [CrossRef]
- Kalim, M.; Liang, K.; Khan, M.S.I.; Zhan, J. Efficient Development and Expression of ScFv Recombinant Proteins against PD-L1 Surface Domain and Potency in Cancer Therapy. Cytotechnology 2019, 71, 705–722. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.H.; Nowicki, C.; Giurini, E.F.; Marzo, A.L.; Zloza, A. Bacterial-Based Cancer Therapy (BBCT): Recent Advances, Current Challenges, and Future Prospects for Cancer Immunotherapy. Vaccines 2021, 9, 1497. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Shimizu, H.; Akiyama, Y.; Taniguchi, S. In Situ Delivery and Production System of Trastuzumab ScFv with Bifidobacterium. Biochem. Biophys. Res. Commun. 2017, 493, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liu, H.; Pan, X.; Feng, Q.; Yang, J. A Method of Delivering an Anti-P21Ras Single-Chain Antibody Fragment to Tumor Sites in Vivo. J. Drug. Deliv. Sci. Technol. 2022, 68, 103024. [Google Scholar] [CrossRef]
- Liu, F.-R.; Bai, S.; Feng, Q.; Pan, X.-Y.; Song, S.-L.; Fang, H.; Cui, J.; Yang, J.-L. Anti-Colorectal Cancer Effects of Anti-P21Ras ScFv Delivered by the Recombinant Adenovirus KGHV500 and Cytokine-Induced Killer Cells. BMC Cancer 2018, 18, 1087. [Google Scholar] [CrossRef]
- Lin, X.-R.; Zhou, X.-L.; Feng, Q.; Pan, X.-Y.; Song, S.-L.; Fang, H.; Lei, J.; Yang, J.-L. CIK Cell-Based Delivery of Recombinant Adenovirus KGHV500 Carrying the Anti-P21Ras ScFv Gene Enhances the Anti-Tumor Effect and Safety in Lung Cancer. J. Cancer Res. Clin. Oncol. 2019, 145, 1123–1132. [Google Scholar] [CrossRef]
- Dai, F.; Zhang, P.-B.; Feng, Q.; Pan, X.-Y.; Song, S.-L.; Cui, J.; Yang, J.-L. Cytokine-Induced Killer Cells Carrying Recombinant Oncolytic Adenovirus Expressing P21Ras ScFv Inhibited Liver Cancer. J. Cancer 2021, 12, 2768–2776. [Google Scholar] [CrossRef]
- Falanga, A.; Lombardi, L.; Galdiero, E.; Genio, V.D.; Galdiero, S. The World of Cell Penetrating: The Future of Medical Applications. Future Med. Chem. 2020, 12, 1431–1446. [Google Scholar] [CrossRef]
- Huang, C.-C.; Liu, F.-R.; Feng, Q.; Pan, X.-Y.; Song, S.-L.; Yang, J.-L. RGD4C Peptide Mediates Anti-P21Ras ScFv Entry into Tumor Cells and Produces an Inhibitory Effect on the Human Colon Cancer Cell Line SW480. BMC Cancer 2021, 21, 321. [Google Scholar] [CrossRef]
- Du, Y.; Lin, X.; Feng, Q.; Pan, X.; Song, S.; Yang, J. Inhibition of Human Lung Cancer Cells by Anti-P21Ras ScFv Mediated by the Activatable Cell-Penetrating Peptide. Anticancer Drugs 2022, 33, e562–e572. [Google Scholar] [CrossRef]
- Klee, G.G. Human Anti-Mouse Antibodies. Arch. Pathol. Lab. Med. 2000, 124, 921–923. [Google Scholar] [CrossRef]
- Kuramochi, T.; Igawa, T.; Tsunoda, H.; Hattori, K. Humanization and Simultaneous Optimization of Monoclonal Antibody. Methods Mol. Biol. 2019, 1904, 213–230. [Google Scholar] [CrossRef]
- Vallabhajosula, S. Molecular Imaging: Radiopharmaceuticals for PET and SPECT; Springer Science & Business Media: Hoboken, NJ, USA, 2009; ISBN 978-3-54076-735-0. [Google Scholar]
- Behr, T.M.; Goldenberg, D.M.; Becker, W. Reducing the Renal Uptake of Radiolabeled Antibody Fragments and Peptides for Diagnosis and Therapy: Present Status, Future Prospects and Limitations. Eur. J. Nucl. Med. 1998, 25, 201–212. [Google Scholar] [CrossRef]
- Peltomaa, R.; Barderas, R.; Benito-Peña, E.; Moreno-Bondi, M.C. Recombinant Antibodies and Their Use for Food Immunoanalysis. Anal. Bioanal. Chem. 2022, 414, 193–217. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mAb Name | Target/Epitope | Antibody Kind | Cancer Kind | Year Approval |
|---|---|---|---|---|
| Rituximab | CD20/169-PANPSE-174 and 183-CYSIQ-187 [22] | Chimeric IgG1 | Non-Hodgkin lymphoma | 1997 |
| Trastuzumab | HER2 1/extracellular domain [28] | Humanized IgG1 | Breast | 1998 |
| Alemtuzumab | CD52/C-terminal with part of the GPI anchor [29] | Humanized IgG1 | Chronic myeloid leukemia | 2001 |
| Ibritumomab tiuxetan | CD20/Same as Rituximab [30] | Murine IgG1 | Non-Hodgkin lymphoma | 2002 |
| Cetuximab | EGFR/Domain III amino acids 334–504 [31] | Chimeric IgG1 | Colorectal | 2004 |
| Bevacizumab | VEGF-A/Hairpin loop (β5–turn–β6) and β2–α2–β3 [32] | Humanized IgG1 | Colorectal | 2004 |
| Panitumumab | EGFR/Domain III, P349, P362, D355, F412 and I438 [33] | Human IgG2 | Colorectal | 2006 |
| Ofatumumab | CD20/FLKMESLNFIRAHT region [34] | Human IgG1 | Chronic lymphocytic leukemia | 2009 |
| Ipilimumab | CTLA-4/front β-sheet [35] | Human IgG1 | Metastatic melanoma | 2011 |
| Brentuximab vedotin | CD30/Extracellular domain [36] | Chimeric IgG1 | Hodgkin lymphoma, systemic anaplastic large cell lymphoma | 2011 |
| Pertuzumab | HER2/Extracellular domain II [37] | Humanized IgG1 | Breast | 2012 |
| Obinutuzumab | CD20/Large extracellular loop (172–176 region) [30] | Humanized IgG1 Glycoengineered | Chronic lymphocytic leukemia | 2013 |
| Ramucirumab | VEGFR2/Domain III [38] | Human IgG1 | Gastric | 2014 |
| Blinatumomab | CD19, CD3/Residues 97–107, 155–166, and 216–224 [39] | Murine bispecific tandem scFv | Acute lymphoblastic leukemia | 2014 |
| Nivolumab | PD-1/BC-loop [40] | Human IgG4 | Melanoma, non-small cell lung | 2014 |
| Pembrolizumab | PD-1/C, C′, and G antiparallel beta sheets and C-C′ and F-G loops [41] | Humanized IgG4 | Melanoma | 2014 |
| Necitumumab | EGFR/Domain III [42] | Human IgG1 | Non-small cell lung cancer | 2015 |
| Dinutuximab | GD2 [43] | Chimeric IgG1 | Neuroblastoma | 2015 |
| Daratumumab | CD38/C-terminal loop (residues 189–202 and 223–236) [44] | Human IgG1 | Multiple myeloma | 2015 |
| Elotuzumab | SLAMF7/IgC2 domain [45] | Humanized IgG1 | Multiple myeloma | 2015 |
| Olaratumab | PDGFRα 2/Extracellular domain [46] | Human IgG1 | Soft tissue sarcoma | 2016 |
| Atezolizumab | PD-L1 3/Beta-sheet C′ and B-C loop [41] | Humanized IgG1 | Bladder | 2016 |
| Inotuzumab ozogamicin | CD22/V-like domain [47] | Humanized IgG4 | Acute lymphoblastic leukemia | 2017 |
| Avelumab | PD-L1/Central beta-sheets C and F [41] | Human IgG1 | Merkel cell carcinoma | 2017 |
| Gemtuzumab ozogamicin | CD33/Ig-like V-set domain [48] | Humanized IgG4 | Acute myeloid leukemia | 2017 |
| Durvalumab | PD-L1/Central beta-sheets C and F [41] | Human IgG1 | Bladder | 2017 |
| Cemiplimab | PD-1/BC and FG loops (N58 Glycan) [49] | Human mAb | Cutaneous squamous cell carcinoma | 2018 |
| Polatuzumab vedotin-piiq | CD79β/ARSEDRYRNPKGS [50] | Humanized IgG1 | Diffuse large B-cell lymphoma | 2019 |
| Enfortumab vedotin-ejfv | Nectin-4/V-domain [51] | Human IgG1 | Cancers expressing Nectin-4 | 2019 |
| Sacituzumab govitecan | Trop-2/Q237-Q252 [52] | Humanized IgG1 | Solid tumors | 2020 |
| Isatuximab-irfc | CD38/C-terminal loop (residues 81–90) [44] | Chimeric IgG1 | Multiple myeloma | 2020 |
| Tafasitamab-cxix | CD19 [53] | Fc-modified IgG1 | Diffuse large B-cell lymphoma | 2020 |
| Belantamab mafodotin-blmf | BCMA 4 [54] | Afucosylated IgG1 | Multiple myeloma | 2020 |
| Naxitamab | GD2 [55] | Recombinant humanized IgG1 | Neuroblastoma | 2020 |
| Margetuximab-cmkb | HER2/Extracellular domain [28] | Chimeric Fc-engineered IgG1 | Metastatic HER2-positive breast | 2020 |
| Loncastuximab tesirine-lpyl | CD19/RB4 [56] | Humanized IgG1 | Large B-cell lymphoma | 2021 |
| Amivantamab-vmjw | EGFR/Residues K443, K465, I467, S468 [57] and MET | Human Ig G1-based bispecific antibody | Metastatic non-small cell lung | 2021 |
| Dostarlimab-gxly | PD-1/PD-L1/BC, C′D and FG loops [58] | IgG4 humanized | Advanced solid tumors | 2021 |
| Tisotumab vedotin-tftv | Tissue Factor [59] | IgG1 | Cervical | 2021 |
| Teclistamab-cqyv | BCMA [60] | Humanized Ig G4-proline, alanine, alanine | Multiple myeloma | 2022 |
| Mirvetuximab soravtansine-gynx | FRα 5 [61] | IgG1, Antibody-drug conjugate | Epithelial ovarian, fallopian tube, or peritoneal | 2022 |
| Mosunetuzumab-axgb | CD20/CD3 [62] | Bispecific CD20-directed CD3 T-cell engager | Relapsed or refractory follicular lymphoma | 2022 |
| Kind/Treatment Name | Description | Clinical Phase | Evidence |
|---|---|---|---|
| L19-IL2 is a tumor-directed immunocytokine consisting of IL2 and scFv directed against the ED-B domain of fibronectin. | Advanced solid tumors. | I/II | Register: NCT01058538 [106]. |
| Blinatumomab. | Relapsed or refractory B-cell precursor Philadelphia chromosome-negative acute lymphoblastic leukemia (R/R ALL). | Approved | Register: BLA 125557 [102]. |
| CAR-T cells, where the CAR consists of a scFv directed against CD19, with three intracellular signaling domains derived from CD3 zeta, CD28, and 4-1BB. | B cell lymphoma B cell leukemia. | I/II | Register: NCT02132624 [107]. |
| CAR-T cells targeted to CD19 by a humanized scFv. | B-cell chronic lymphocytic leukemia treatment. | I/II | Register: NCT02782351 [108]. |
| Moxetumomab pasudotox. | Hairy cell leukemia. | Approved | Register: 1020748-57-5 [105]. |
| CAR-T cells expressing an anti-CD19 scFv bound to TCRζ and 4-1BB signaling domains. | Multiple myeloma. | I | Register: NCT02135406 [109]. |
| CAR-T cells expressing scFv with 41BB costimulatory domain and CD3ζ signaling domain targeting mesothelin or CD19. | Pancreatic cancer. | I | Register: NCT03497819 [110]. |
| CAR-T cells expressing an anti-CD19 scFv. | B-cell acute lymphocytic leukemia B-cell chronic lymphocytic leukemia B cell lymphoma. | I | Register: NCT03685786 [111]. |
| CART-19 cells transduced with a lentiviral vector to express anti-CD19 scFv. | Patients With B Cell ALL, Relapsed or Refractory, With no Available Curative Treatment Options. | II | Register: NCT02030847 [112]. |
| Combination of radiotherapy with Darleukin, which is a fusion protein L19-IL2, composed of two fractions: L19, a scFv, linked by a flexible linker to IL2. | Stage IV non-small cell lung cancer. | II | Register: NCT03705403 [113]. |
| Autologous T cells expressing anti-CD19 scFv chimeric antigen receptors. | B cell neoplasms, B cell lymphoma, B-cell acute lymphoblastic leukemia. | I | Register: NCT03559439 [114]. |
| CAR-T cells expressing anti-PD-L1 scFv. | Advanced lung cancer. | I | Register: NCT03330834 [115]. |
| CAR-T cells expressing an anti-CD276 scFv. | Solid tumors. | -- | Register: NCT04691713 [116]. |
| Autologous T cells expressing an anti-BCMA scFv coupled to TCRζ and 4-1BB signaling domains. | Multiple myeloma | I | Register: NCT02546167 [117]. |
| CAR T cells expressing anti-BCMA scFv. | Multiple myeloma | I | Register: NCT04650724 [118]. |
| CART19 cells transduced with a lentiviral vector to express anti-CD19 scFv. | Leukemia, Acute Lymphoblastic. | II | Register: NCT02935543 [119]. |
| TILs/CAR-TILs with PD1 knockout and Anti-PD1/CTLA4-scFv Secreting or CARs. | Solid tumors such as liver, breast, lung, colorectal, and brain. | I | Register: NCT04842812 [120]. |
| CAR-T cells expressing scFv with affinity for malignant tumors. | Malignant tumors in children. | I | Register: NCT04691349 [121]. |
| Autologous T cells expressing scFv with specificity against GFRα4. | Recurrent or metastatic medullary thyroid cancer. | I | Register: NCT04877613 [122]. |
| CART-meso cells expressing an anti-mesothelin scFv fused with TCRζ and 4-1BB costimulatory domains. | Pancreatic cancer. | -- | Register: NCT03638193 [123]. |
| CAR T cells (huMNC2-CAR44) that bind via a scFv to the extracellular domain of the cleaved form of MUC1 (called MUC1*). | Metastatic breast cancer. | I | Register: NCT04020575 [124]. |
| CAR-T cells secreting scFv against OX40. | Lung cancer, hepatocellular carcinoma and solid tumor. | I | Register: NCT04952272 [125]. |
| CAR-T cells that express scFv’s against PD1/CTLA4/Tigit. | Lung cancer. | I | Register: NCT03198052 [126]. |
| CAR-T cells with Ibalizumab-derived anti-CD4 scFv and the intracellular domains of CD28 and 4-1BB coactivators fused with the CD3ζ T cell activation signaling domain. | T cell lymphoma. T cell leukemia. | I | Register: NCT03829540 [127]. |
| CAR-T cells expressing an anti-BCMA scFv. | Multiple myeloma. | I/II | Register: NCT05066646 [128]. |
| Autologous T cells expressing an anti-PSMA scFv, CD2 costimulatory domain, and dual-shielded with a dominant TGFβ receptor-negative domain and PD1.CD28 switch. | Metastatic prostate cancer. | I/II | Register: NCT05489991 [129]. |
| AR-NK cells that express and secrete IL7/CCL19 and/or scFv against PD1/CTLA4/Lag3, targeting Claudin 6. | Stage IV Ovarian Cancer Treatment. Refractory testicular cancer. Recurrent endometrial cancer. | I/II | Register: NCT05410717 [130]. |
| Autologous T cells containing anti CD19 and anti CD20 scFv coupled to CD3ζ and co-stimulatory domain 4-1BB (4-1BB). | Non-Hodgkin’s lymphoma. B cell lymphoma. Chronic lymphocytic leukemia. Small lymphocyte lymphoma. | I | Register: NCT03019055 [131]. |
| Tetravalent IgG(H)-scFv fusion-type of bi-specific antibody (BsAb). | Solid tumor. Advanced cancer metastatic cancer. Gastric cancer. Gastroesophageal junction carcinoma adenocarcinoma of the esophagus. Pancreatic ductal adenocarcinoma. | I | Register: NCT04900818 [132]. |
| D2C7-IT is an immunotoxin composed of a scFv with high affinity for EGFRwt and EGFRvIII. | Malignant glioma recurrent brain tumor | I | Register: NCT02303678 [133]. |
| L19TNF is a fully human fusion protein consisting of human TNF-α fused to the L19 antibody in scFv format, specific for the extra B domain of fibronectin. | Glioblastoma. | I/II | Register: NCT04573192 [134]. |
| CAR-T cells expressing an anti-GPRC5D scFv. | Relapsed/Refractory Multiple Myeloma Plasma cell leukemia. | I | Register: NCT05219721 [135]. |
| BRITE is a bispecific hooker that has one scFv binding for the CD3 epsilon subunit while another scFv is directed against the hEGFRvIII epitope that is differentially expressed on the surface of tumor cells. | Malignant glioma Glioblastoma. | I | Register: NCT04903795 [136]. |
| CAR-T cells expressing an scFv that recognizes CD19 and dual co-stimulating intracellular signaling domains (4-1BB and CD3ζ). | Recurrent Non-Hodgkin’s lymphoma. relapsed adult ALL. Recurrent Pediatric ALL. | I/II | Register: NCT03938987 [137]. |
| Autologous Lymphoma Ig-derived scFv-chemokine DNA Vaccine. | Lymphoplasmacytic Lymphoma. | I | Register: NCT01209871 [138]. |
| T cells expressing anti-CD123 scFv chimeric antigen receptors linked to TCRζ and 4-1BB signaling domains. | Acute myeloid leukemia, relapsed. Acute myeloid leukemia, pediatric. Refractory acute myeloid leukemia. | I | Register: NCT04678336 [139]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Nava, C.; Ortuño-Pineda, C.; Illades-Aguiar, B.; Flores-Alfaro, E.; Leyva-Vázquez, M.A.; Parra-Rojas, I.; del Moral-Hernández, O.; Vences-Velázquez, A.; Cortés-Sarabia, K.; Alarcón-Romero, L.d.C. Mechanisms of Action and Limitations of Monoclonal Antibodies and Single Chain Fragment Variable (scFv) in the Treatment of Cancer. Biomedicines 2023, 11, 1610. https://doi.org/10.3390/biomedicines11061610
Rodríguez-Nava C, Ortuño-Pineda C, Illades-Aguiar B, Flores-Alfaro E, Leyva-Vázquez MA, Parra-Rojas I, del Moral-Hernández O, Vences-Velázquez A, Cortés-Sarabia K, Alarcón-Romero LdC. Mechanisms of Action and Limitations of Monoclonal Antibodies and Single Chain Fragment Variable (scFv) in the Treatment of Cancer. Biomedicines. 2023; 11(6):1610. https://doi.org/10.3390/biomedicines11061610
Chicago/Turabian StyleRodríguez-Nava, Cynthia, Carlos Ortuño-Pineda, Berenice Illades-Aguiar, Eugenia Flores-Alfaro, Marco Antonio Leyva-Vázquez, Isela Parra-Rojas, Oscar del Moral-Hernández, Amalia Vences-Velázquez, Karen Cortés-Sarabia, and Luz del Carmen Alarcón-Romero. 2023. "Mechanisms of Action and Limitations of Monoclonal Antibodies and Single Chain Fragment Variable (scFv) in the Treatment of Cancer" Biomedicines 11, no. 6: 1610. https://doi.org/10.3390/biomedicines11061610
APA StyleRodríguez-Nava, C., Ortuño-Pineda, C., Illades-Aguiar, B., Flores-Alfaro, E., Leyva-Vázquez, M. A., Parra-Rojas, I., del Moral-Hernández, O., Vences-Velázquez, A., Cortés-Sarabia, K., & Alarcón-Romero, L. d. C. (2023). Mechanisms of Action and Limitations of Monoclonal Antibodies and Single Chain Fragment Variable (scFv) in the Treatment of Cancer. Biomedicines, 11(6), 1610. https://doi.org/10.3390/biomedicines11061610