

Adipocyte-Derived Adipokines and Other Obesity-Associated Molecules in Feline Mammary Cancer

Abstract
:1. Introduction
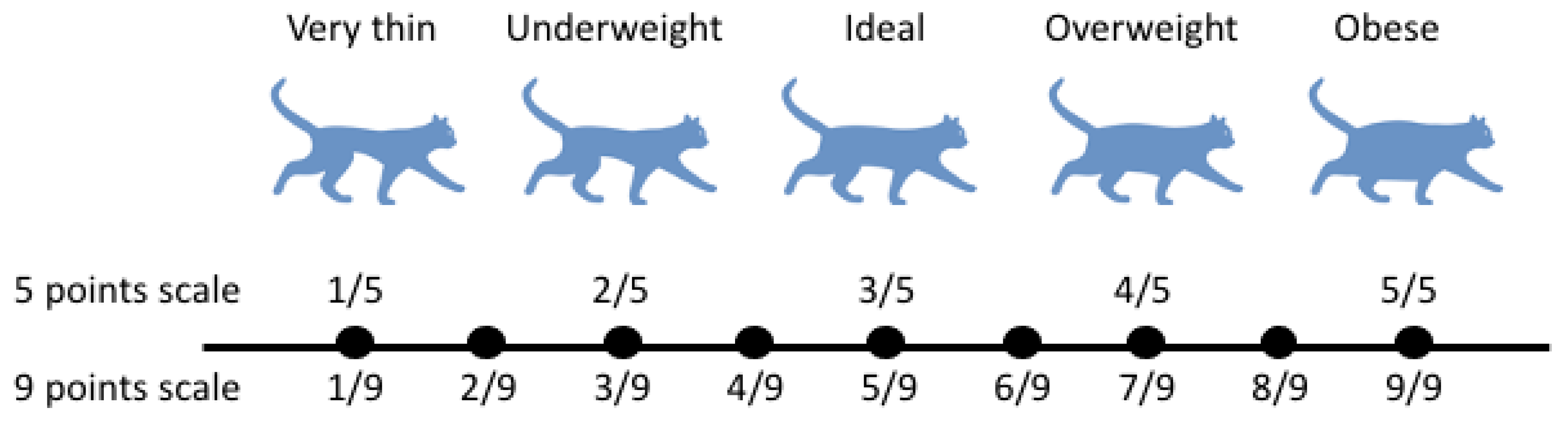
2. Obesity Scoring in Domestic Cats
3. White Adipose Tissue as an Endocrine Organ
4. Obesity as a Chronic Inflammatory Disorder
5. Sex Hormone Modulation of White Adipose Tissue
6. Feline Mammary Cancer
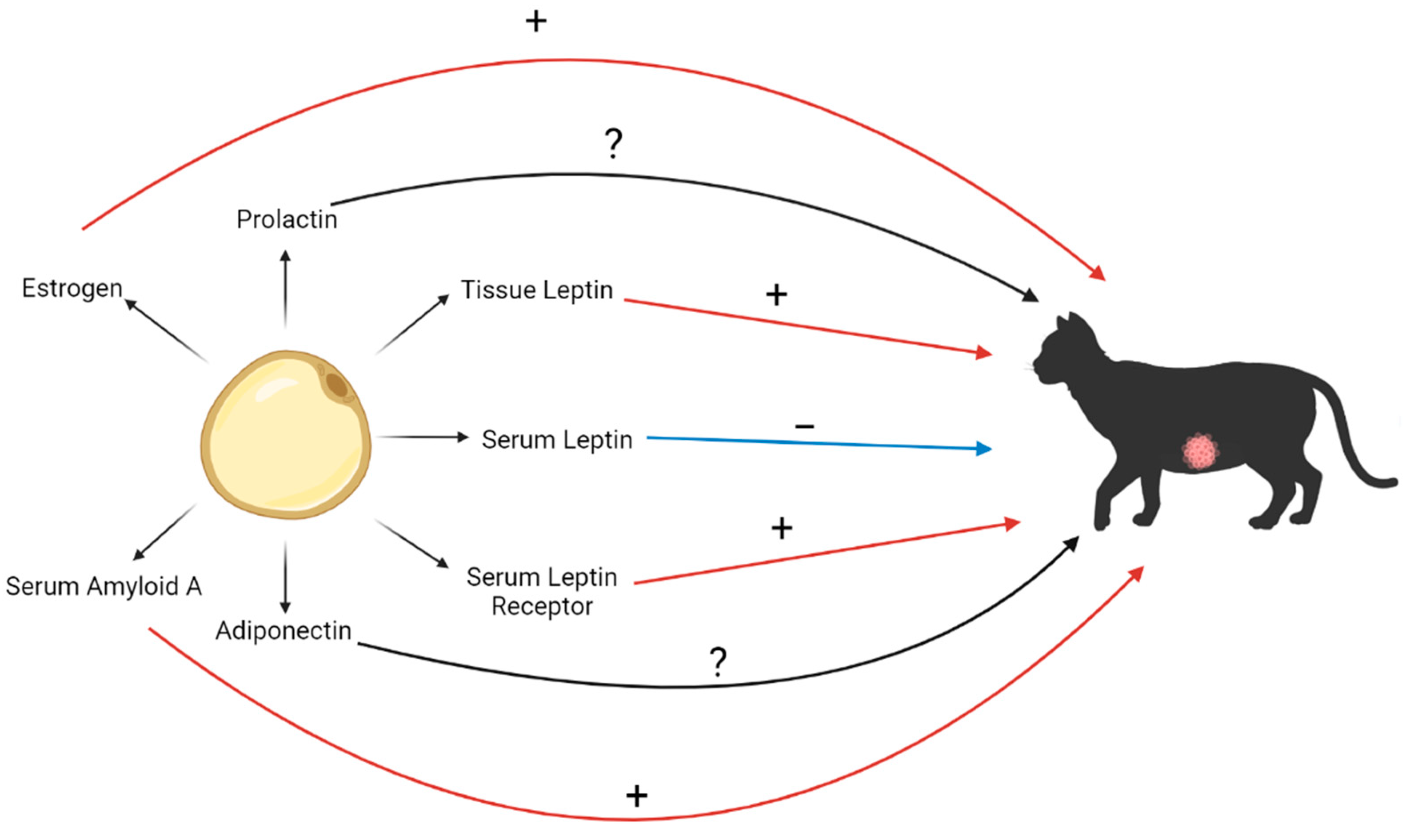
7. Mediators of Obesity and their Association with FMC
7.1. Leptin
7.2. Adiponectin
7.3. Serum Amyloid A
7.4. Estrogen
7.5. Prolactin
8. Conclusions and Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Okada, Y.; Ueno, H.; Mizorogi, T.; Ohara, K.; Kawasumi, K.; Arai, T. Diagnostic Criteria for Obesity Disease in Cats. Front. Vet. Sci. 2019, 6, 284. [Google Scholar] [CrossRef] [PubMed]
- German, A.J.; Ryan, V.H.; German, A.C.; Wood, I.S.; Trayhurn, P. Obesity, its associated disorders and the role of inflammatory adipokines in companion animals. Vet. J. 2010, 185, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, C.; Ferreira, F. Tumor microenvironment of human breast cancer, and feline mammary carcinoma as a potential study model. Biochim. Biophys. Acta Rev. Cancer 2021, 1876, 188587. [Google Scholar] [CrossRef] [PubMed]
- Lorincz, A.M.; Sukumar, S. Molecular links between obesity and breast cancer. Endocr. Relat. Cancer 2006, 13, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Gameiro, A.; Nascimento, C.; Urbano, A.C.; Correia, J.; Ferreira, F. Serum and Tissue Expression Levels of Leptin and Leptin Receptor Are Putative Markers of Specific Feline Mammary Carcinoma Subtypes. Front. Vet. Sci. 2021, 8, 625147. [Google Scholar] [CrossRef]
- Cave, N.J.; Backus, R.C.; Marks, S.L.; Klasing, K.C. Oestradiol and genistein reduce food intake in male and female overweight cats after gonadectomy. N. Z. Vet. J. 2007, 55, 113–119. [Google Scholar] [CrossRef]
- Montoya, M.; Morrison, J.A.; Arrignon, F.; Spofford, N.; Charles, H.; Hours, M.A.; Biourge, V. Life expectancy tables for dogs and cats derived from clinical data. Front. Vet. Sci. 2023, 10, 1082102. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.; Madeira, S.; Correia, J.; Peleteiro, M.; Cardoso, F.; Ferreira, F. Molecular based subtyping of feline mammary carcinomas and clinicopathological characterization. Breast 2016, 27, 44–51. [Google Scholar] [CrossRef]
- Adega, F.; Borges, A.; Chaves, R. Cat Mammary Tumors: Genetic Models for the Human Counterpart. Vet. Sci. 2016, 3, 17. [Google Scholar] [CrossRef]
- Teng, K.T.; McGreevy, P.D.; Toribio, J.A.L.M.L.; Raubenheimer, D.; Kendall, K.; Dhand, N.K. Associations of body condition score with health conditions related to overweight and obesity in cats. J. Small Anim. Pract. 2018, 59, 603–615. [Google Scholar] [CrossRef]
- Radin, M.J.; Sharkey, L.C.; Holycross, B.J. Adipokines: A review of biological and analytical principles and an update in dogs, cats, and horses. Vet. Clin. Pathol. 2009, 38, 136–156. [Google Scholar] [CrossRef]
- Tarkosova, D.; Story, M.M.; Rand, J.S.; Svoboda, M. Feline obesity—Prevalence, risk factors, pathogenesis, associated conditions and assessment: A review. Veterinární Medicína 2016, 61, 295–307. [Google Scholar] [CrossRef]
- Stenberg, K.; Novotny, G.W.; Lutz, T.A.; Mandrup-Poulsen, T.; Bjornvad, C.R. Obesity-induced changes in gene expression in feline adipose and skeletal muscle tissue. J. Anim. Physiol. Anim. Nutr. 2023, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Takashima, S.; Nishii, N.; Kobatake, Y.; Kiyosue, M.; Kimura, S.; Kitagawa, H. Concentrations of leptin, adiponectin, and resistin in the serum of obese cats during weight loss. J. Vet. Med Sci. 2019, 81, 1294–1300. [Google Scholar] [CrossRef] [PubMed]
- Chandler, M.; Cunningham, S.; Lund, E.M.; Khanna, C.; Naramore, R.; Patel, A.; Day, M.J. Obesity and Associated Comorbidities in People and Companion Animals: A One Health Perspective. J. Comp. Pathol. 2017, 156, 296–309. [Google Scholar] [CrossRef]
- Winkel, V.M.; Pavan, T.L.; Wirthl, V.A.; Alves, A.L.; Lucas, S.R. Serum alpha-1 acid glycoprotein and serum amyloid A concentrations in cats receiving antineoplastic treatment for lymphoma. Am. J. Vet. Res. 2015, 76, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Ceron, J.J.; Eckersall, P.D.; Martynez-Subiela, S. Acute phase proteins in dogs and cats: Current knowledge and future perspectives. Vet. Clin. Pathol. 2005, 34, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Tamamoto, T.; Ohno, K.; Goto-Koshino, Y.; Tsujimoto, H. Serum amyloid A promotes invasion of feline mammary carcinoma cells. J. Vet. Med. Sci. 2014, 76, 1183–1188. [Google Scholar] [CrossRef]
- Kil, D.Y.; Swanson, K.S. Endocrinology of obesity. Vet. Clin. N. Am. Small Anim. Pract. 2010, 40, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Belsito, K.R.; Vester, B.M.; Keel, T.; Graves, T.K.; Swanson, K.S. Impact of ovariohysterectomy and food intake on body composition, physical activity, and adipose gene expression in cats. J. Anim. Sci. 2009, 87, 594–602. [Google Scholar] [CrossRef]
- Larsen, J.A. Risk of obesity in the neutered cat. J. Feline Med. Surg. 2017, 19, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, J.; Beeraka, N.M.; Tang, C.; Babayeva, Y.V.; Sinelnikov, M.Y.; Zhang, X.; Zhang, J.; Liu, J.; Reshetov, I.V.; et al. Advances in the Prevention and Treatment of Obesity-Driven Effects in Breast Cancers. Front. Oncol. 2022, 12, 820968. [Google Scholar] [CrossRef] [PubMed]
- Overley, B.; Shofer, F.S.; Goldschmidt, M.H.; Sherer, D.; Sorenmo, K.U. Association between Ovarihysterectomy and Feline Mammary Carcinoma. J. Vet. Intern. Med. 2005, 19, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Trummel, D.K. Expression of Prolactin in Feline Mammary Adenomas and Adenocarcinomas. Bachelor’s Thesis, Oregon State University, Corvallis, OR, USA, 2007. [Google Scholar]
- Martin, L.J.; Siliart, B.; Dumon, H.J.; Nguyen, P. Spontaneous hormonal variations in male cats following gonadectomy. J. Feline Med. Surg. 2006, 8, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Gimenez, F.; Hecht, S.; Craig, L.E.; Legendre, A.M. Early detection, aggressive therapy: Optimizing the management of feline mammary masses. J. Feline Med. Surg. 2010, 12, 214–224. [Google Scholar] [CrossRef]
- Tomlinson, M.J.; Barteaux, L.; Ferns, L.E.; Angelopoulos, E. Feline mammary carcinoma: A retrospective evaluation of 17 cases. Can. Vet. J. 1984, 25, 435–439. [Google Scholar]
- Hayes, H.M., Jr.; Milne, K.L.; Mandell, C.P. Epidemiological features of feline mammary carcinoma. Vet. Rec. 1981, 108, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Cannon, C.M. Cats, Cancer and Comparative Oncology. Vet. Sci. 2015, 2, 111–126. [Google Scholar] [CrossRef]
- Courcier, E.A.; Mellor, D.J.; Pendlebury, E.; Evans, C.; Yam, P.S. An investigation into the epidemiology of feline obesity in Great Britain: Results of a cross-sectional study of 47 companion animal practises. Vet. Rec. 2012, 171, 560. [Google Scholar] [CrossRef]
- O’Neill, D.G.; Church, D.B.; McGreevy, P.D.; Thomson, P.C.; Brodbelt, D.C. Longevity and mortality of cats attending primary care veterinary practices in England. J. Feline Med. Surg. 2015, 17, 125–133. [Google Scholar] [CrossRef]
- Novosad, C.A. Principles of treatment for mammary gland tumors. Clin. Tech. Small Anim. Pract. 2003, 18, 107–109. [Google Scholar] [CrossRef] [PubMed]
- Petrucci, G.; Henriques, J.; Gregorio, H.; Vicente, G.; Prada, J.; Pires, I.; Lobo, L.; Medeiros, R.; Queiroga, F. Metastatic feline mammary cancer: Prognostic factors, outcome and comparison of different treatment modalities—A retrospective multicentre study. J. Feline Med. Surg. 2021, 23, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.; Soares, M.; Correia, J.; Adega, F.; Ferreira, F.; Chaves, R. Assessment of ERBB2 and TOP2alpha gene status and expression profile in feline mammary tumors: Findings and guidelines. Aging 2019, 11, 4688–4705. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.; Correia, J.; Rodrigues, P.; Simoes, M.; de Matos, A.; Ferreira, F. Feline HER2 protein expression levels and gene status in feline mammary carcinoma: Optimization of immunohistochemistry (IHC) and in situ hybridization (ISH) techniques. Microsc. Microanal. 2013, 19, 876–882. [Google Scholar] [CrossRef]
- Soares, M.; Correia, J.; Peleteiro, M.C.; Ferreira, F. St Gallen molecular subtypes in feline mammary carcinoma and paired metastases-disease progression and clinical implications from a 3-year follow-up study. Tumor Biol. 2016, 37, 4053–4064. [Google Scholar] [CrossRef]
- Gameiro, A.; Urbano, A.C.; Ferreira, F. Emerging Biomarkers and Targeted Therapies in Feline Mammary Carcinoma. Vet. Sci. 2021, 8, 164. [Google Scholar] [CrossRef]
- Granados-Soler, J.L.; Bornemann-Kolatzki, K.; Beck, J.; Brenig, B.; Schutz, E.; Betz, D.; Junginger, J.; Hewicker-Trautwein, M.; Murua Escobar, H.; Nolte, I. Analysis of Copy-Number Variations and Feline Mammary Carcinoma Survival. Sci. Rep. 2020, 10, 1003. [Google Scholar] [CrossRef]
- Nascimento, C.; Gameiro, A.; Correia, J.; Ferreira, J.; Ferreira, F. The Landscape of Tumor-Infiltrating Immune Cells in Feline Mammary Carcinoma: Pathological and Clinical Implications. Cells 2022, 11, 2578. [Google Scholar] [CrossRef]
- Urbano, A.C.; Nascimento, C.; Soares, M.; Correia, J.; Ferreira, F. Clinical Relevance of the serum CTLA-4 in Cats with Mammary Carcinoma. Sci. Rep. 2020, 10, 3822. [Google Scholar] [CrossRef]
- Frenel, J.S.; Nguyen, F. Mammary carcinoma: Comparative oncology between small animals and humans—New therapeutic tools. Reprod. Domest. Anim. 2023, 1–7. [Google Scholar] [CrossRef]
- Vilhena, H.; Tvarijonaviciute, A.; Ceron, J.J.; Figueira, A.C.; Miranda, S.; Ribeiro, A.; Canadas, A.; Dias-Pereira, P.; Rubio, C.P.; Franco, L.; et al. Acute phase proteins and biomarkers of oxidative status in feline spontaneous malignant mammary tumours. Vet. Comp. Oncol. 2019, 17, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Wiese, D.A.; Thaiwong, T.; Yuzbasiyan-Gurkan, V.; Kiupel, M. Feline mammary basal-like adenocarcinomas: A potential model for human triple-negative breast cancer (TNBC) with basal-like subtype. BMC Cancer 2013, 13, 403. [Google Scholar] [CrossRef] [PubMed]
- Carre, N.; Binart, N. Prolactin and adipose tissue. Biochimie 2014, 97, 16–21. [Google Scholar] [CrossRef]
- Hugo, E.R.; Borcherding, D.C.; Gersin, K.S.; Loftus, J.; Ben-Jonathan, N. Prolactin release by adipose explants, primary adipocytes, and LS14 adipocytes. J. Clin. Endocrinol. Metab. 2008, 93, 4006–4012. [Google Scholar] [CrossRef] [PubMed]
- Nathanson, K.L.; Wooster, R.; Weber, B.L. Breast cancer genetics: What we know and what we need. Nat. Med. 2001, 7, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Baptista, C.S.; Santos, S.; Laso, A.; Bastos, E.; Avila, S.; Guedes-Pinto, H.; Gartner, F.; Gut, I.G.; Castrillo, J.L.; Chaves, R. Sequence variation and mRNA expression of the TWIST1 gene in cats with mammary hyperplasia and neoplasia. Vet. J. 2012, 191, 203–207. [Google Scholar] [CrossRef]
- Govoni, V.M.; Da Silva, T.C.; Guerra, J.M.; Pereira, I.V.A.; Queiroga, F.L.; Cogliati, B. Genetic variants of BRCA1 and BRCA2 genes in cats with mammary gland carcinoma. Vet. Comp. Oncol. 2021, 19, 404–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Subtypes of FMC | Associated Molecules | Clinical-Pathological Features | Prognostic and/or Predictive |
|---|---|---|---|
| Luminal A | ER and/or PR Low Ki-67 index | Less invasive and proliferative tumors Low biologic aggressiveness | Longest survival time and disease-free interval |
| Luminal B HER2-negative | ER and/or PR High Ki-67 index | Less aggressive than luminal B HER2-positive | Better prognosis than the HER2-positive type |
| Luminal B HER2-positive | ER and/or PR fHER2 | High biologic aggressiveness | May benefit from HER2 targeted therapy |
| HER2-positive | fHER2 | Highly proliferative | Second-shortest survival time and disease-free interval [36,38] May benefit from HER-2 targeted therapy [41] |
| Triple-negative basal-like | CK 5/6 | Associated with tissue necrosis High biologic aggressiveness Large tumors | Shortest survival time and disease-free interval No molecular-targeted therapy identified |
| Triple-negative normal-like | None | Associated with tissue necrosis High biologic aggressiveness Large tumors |
| Molecule | Correlation with FMC | Reference |
|---|---|---|
| Tissue leptin | Positively correlated | [5] |
| Serum leptin | Negatively correlated | [5] |
| Serum leptin receptor | Positively correlated | [5] |
| Adiponectin | None documented to date | N/A |
| Serum amyloid A (SAA) | Positively correlated | [42] |
| Estrogen | Positively correlated | [23] |
| Prolactin | Unclear | [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marshall, T.; Chen, J.; Viloria-Petit, A.M. Adipocyte-Derived Adipokines and Other Obesity-Associated Molecules in Feline Mammary Cancer. Biomedicines 2023, 11, 2309. https://doi.org/10.3390/biomedicines11082309
Marshall T, Chen J, Viloria-Petit AM. Adipocyte-Derived Adipokines and Other Obesity-Associated Molecules in Feline Mammary Cancer. Biomedicines. 2023; 11(8):2309. https://doi.org/10.3390/biomedicines11082309
Chicago/Turabian StyleMarshall, Taylor, Jing Chen, and Alicia M. Viloria-Petit. 2023. "Adipocyte-Derived Adipokines and Other Obesity-Associated Molecules in Feline Mammary Cancer" Biomedicines 11, no. 8: 2309. https://doi.org/10.3390/biomedicines11082309
APA StyleMarshall, T., Chen, J., & Viloria-Petit, A. M. (2023). Adipocyte-Derived Adipokines and Other Obesity-Associated Molecules in Feline Mammary Cancer. Biomedicines, 11(8), 2309. https://doi.org/10.3390/biomedicines11082309