

Is Spheroid a Relevant Model to Address Fibrogenesis in Keloid Research?


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Clinical Study Approval
2.2. Human Normal Dermal (NDFs) and Keloid Fibroblasts (KFs) Collection
2.3. Monolayer and Spheroid Cultures
2.4. Histological and Immuno-Histological Characterization of Spheroids
2.5. Terminal Deoxynucleotidyl Trasferas dUTP Nick End Labeling (TUNEL) Assay
2.6. RT-qPCR
2.7. α-Smooth Muscle Actin and Fibronectin Quantification
2.8. Statistical Analysis
3. Results
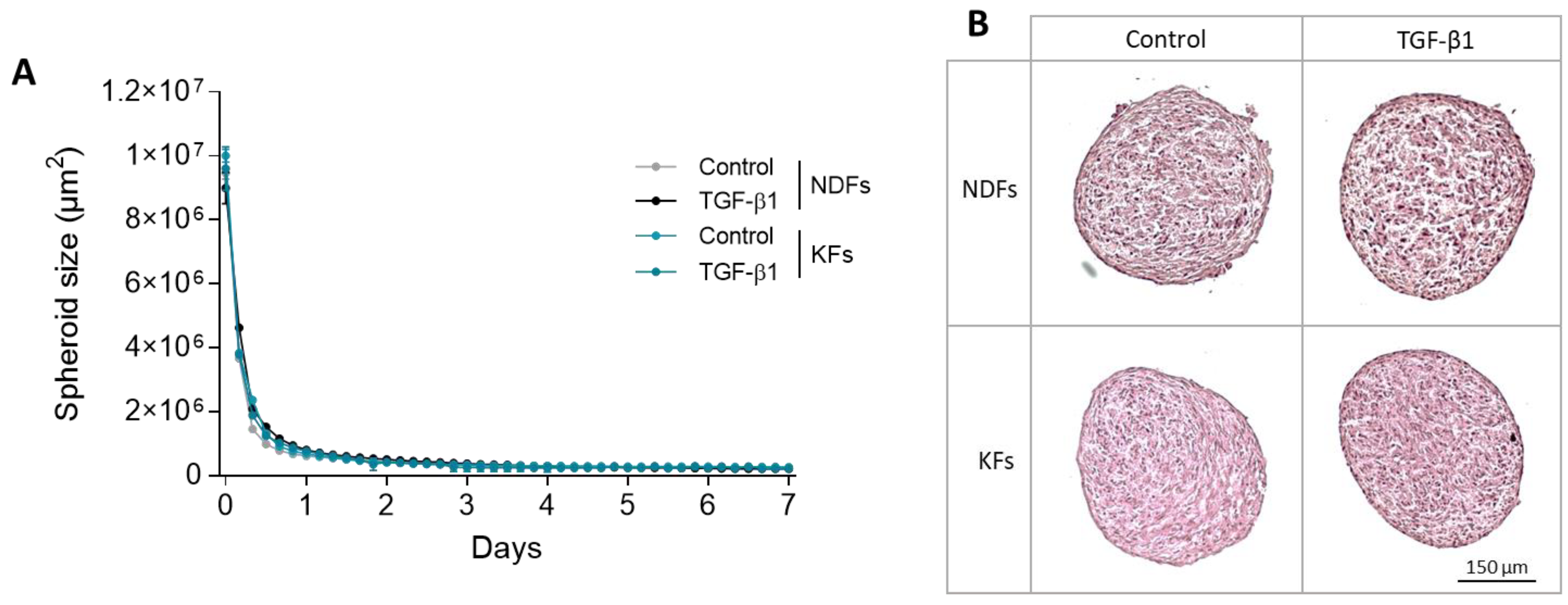
3.1. KFs and NDFs Diameter Equally Evolve Overtime Independently from TGF-β1 Activation
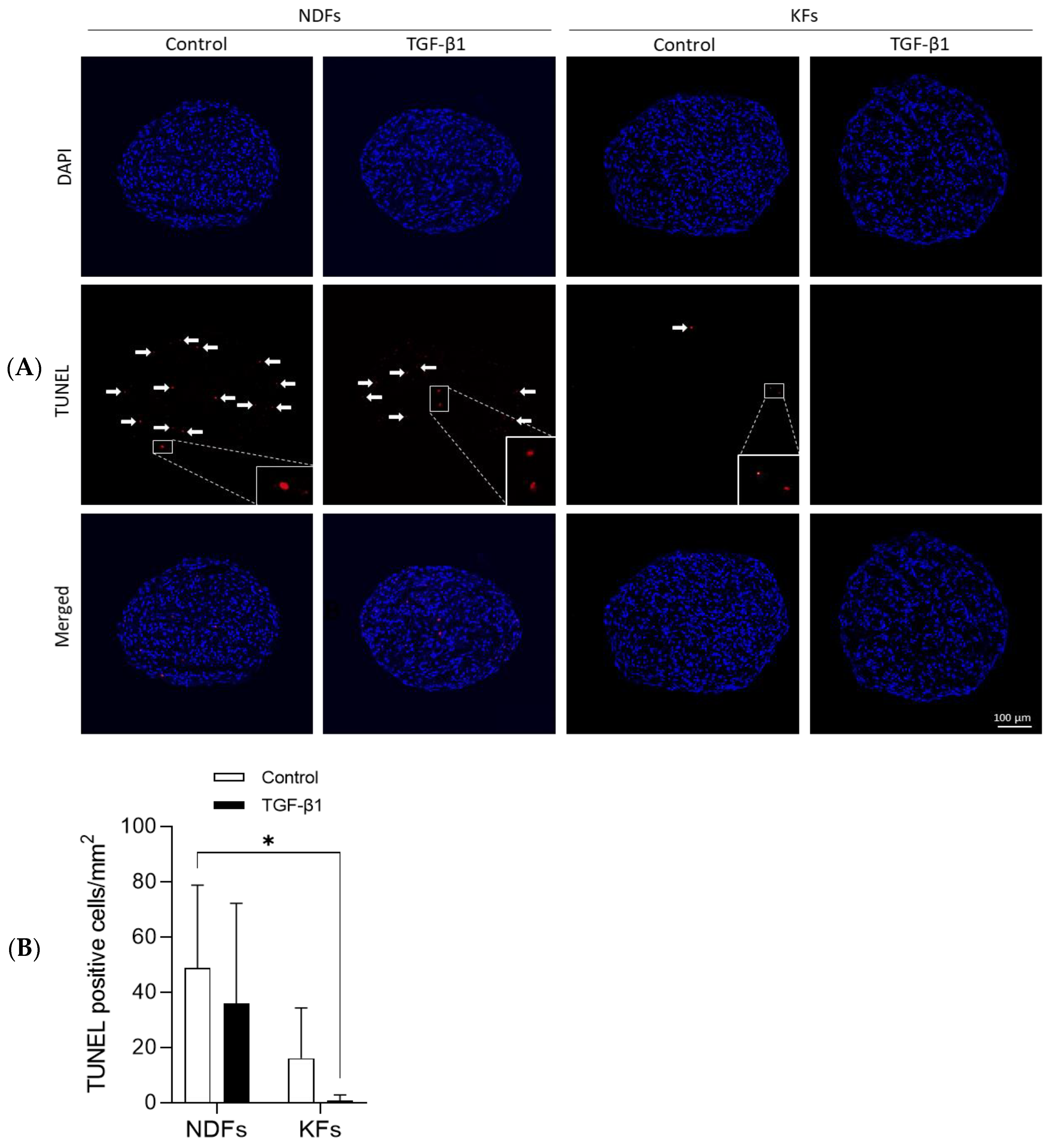
3.2. TGF-β1 Reduced Apoptotic Cells Rate in KFs Spheroids More Than in NDFs Ones
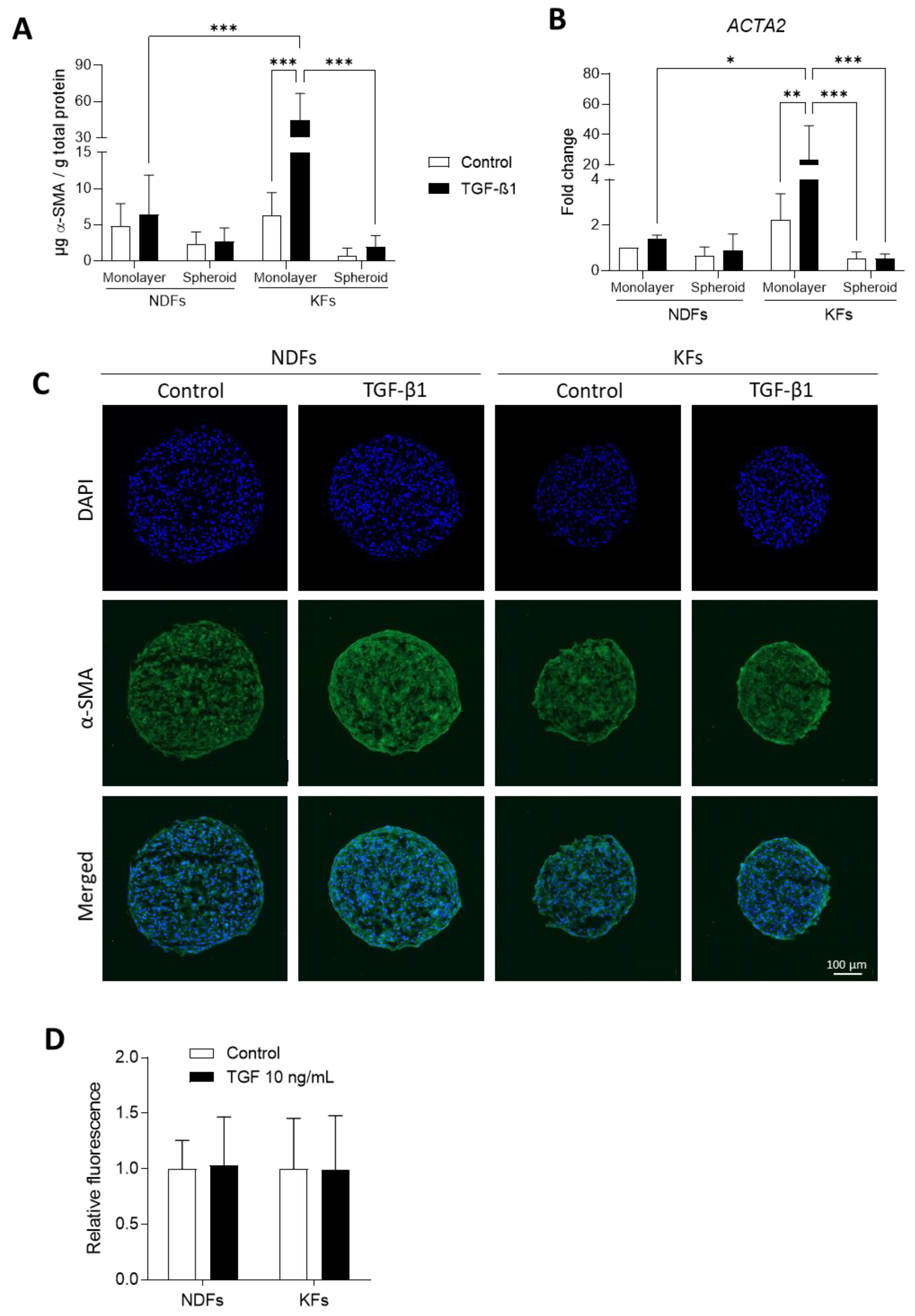
3.3. TGF-β1-Induced α-SMA Expression Discontinues in KFs Spheroids
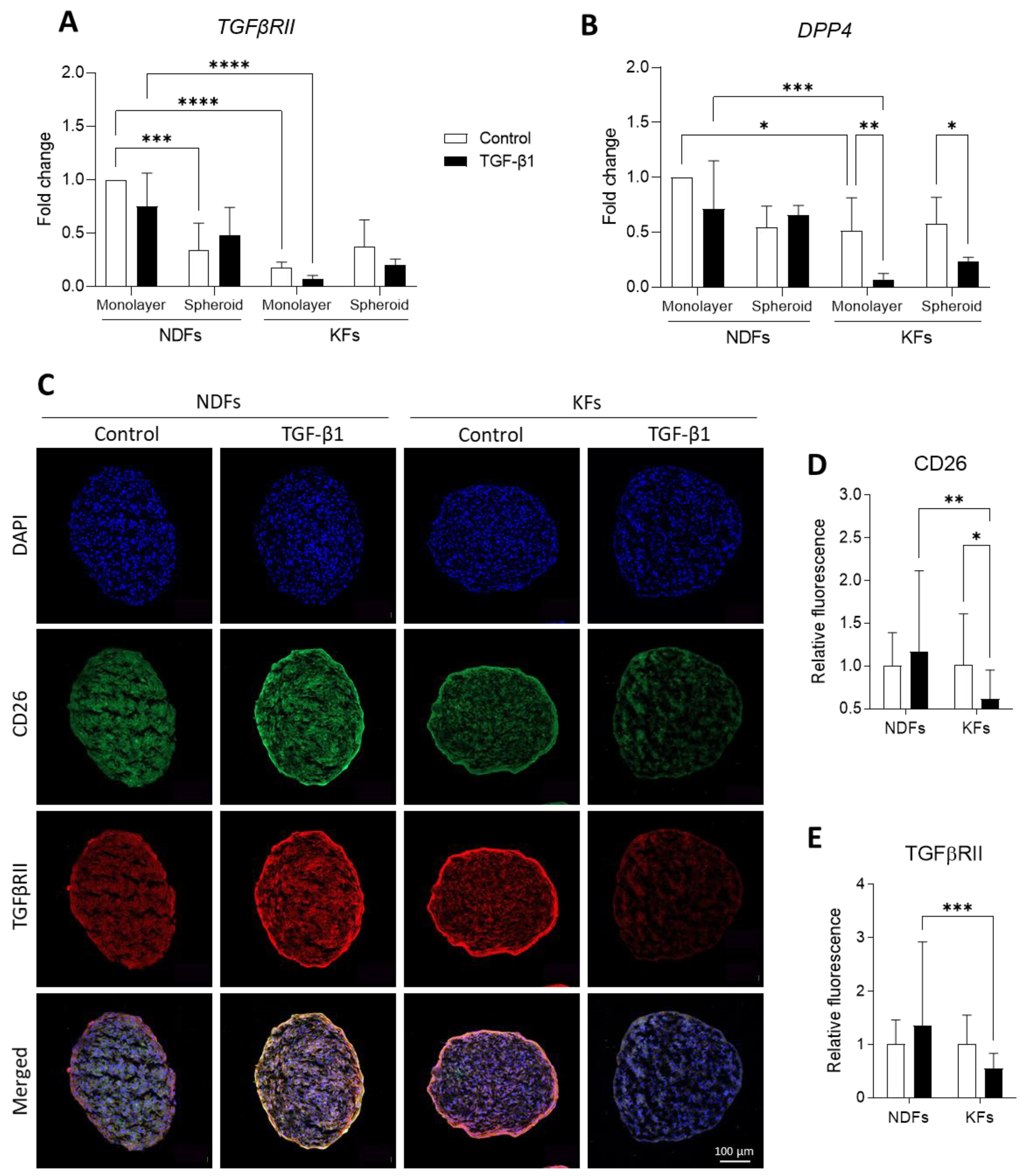
3.4. Three-Dimensional Culture and TGF-B1 Activation Converge to Downregulate CD26 and TGFβRII Expression
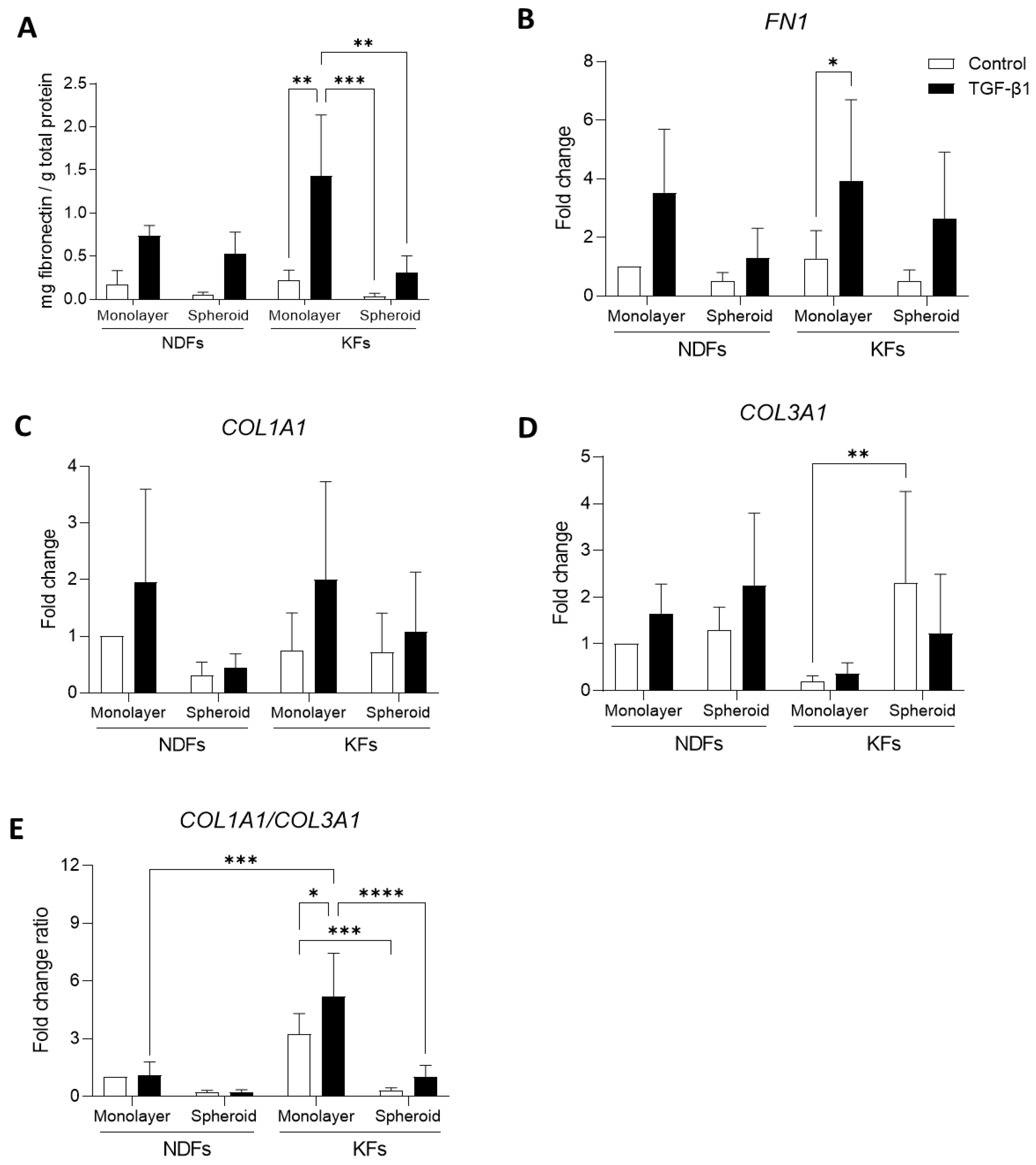
3.5. Overexpression of ECM Related Genes Discontinues When KFs Are Cultured from 2D to 3D
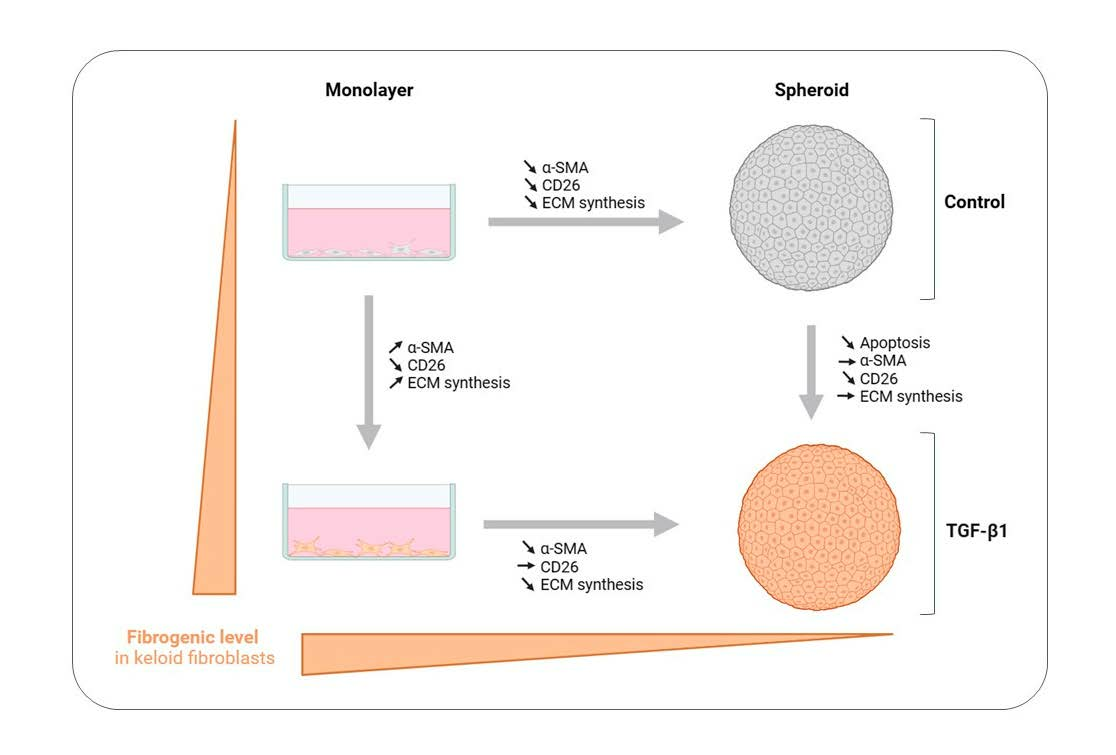
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. ICD-11 for Mortality and Morbidity Statistics. Available online: https://icd.who.int/browse11/l-m/en#/http%3a%2f%2fid.who.int%2ficd%2fentity%2f831995767 (accessed on 27 July 2023).
- Ogawa, R. Keloid and Hypertrophic Scars Are the Result of Chronic Inflammation in the Reticular Dermis. Int. J. Mol. Sci. 2017, 18, 606. [Google Scholar] [CrossRef]
- Tan, S.; Khumalo, N.; Bayat, A. Understanding Keloid Pathobiology from a Quasi-Neoplastic Perspective: Less of a Scar and More of a Chronic Inflammatory Disease with Cancer-Like Tendencies. Front. Immunol. 2019, 10, 1810. [Google Scholar] [CrossRef]
- Betarbet, U.; Blalock, T.W. Keloids: A Review of Etiology, Prevention, and Treatment. J. Clin. Aesthet. Dermatol. 2020, 13, 33–43. [Google Scholar]
- Supp, D.M. Animal Models for Studies of Keloid Scarring. Adv. Wound Care 2019, 8, 77–89. [Google Scholar] [CrossRef]
- Lebeko, M.; Khumalo, N.P.; Bayat, A. Multi-dimensional models for functional testing of keloid scars: In silico, in vitro, organoid, organotypic, ex vivo organ culture, and in vivo models. Wound Repair Regen. 2019, 27, 298–308. [Google Scholar] [CrossRef]
- Marty, P.; Chatelain, B.; Lihoreau, T.; Tissot, M.; Dirand, Z.; Humbert, P.; Senez, C.; Secomandi, E.; Isidoro, C.; Rolin, G. Halofuginone regulates keloid fibroblast fibrotic response to TGF-β induction. Biomed. Pharmacother. 2021, 135, 111182. [Google Scholar] [CrossRef]
- Feng, C.; Shan, M.; Xia, Y.; Zheng, Z.; He, K.; Wei, Y.; Song, K.; Meng, T.; Liu, H.; Hao, Y.; et al. Single-cell RNA sequencing reveals distinct immunology profiles in human keloid. Front. Immunol. 2022, 13, 940645. [Google Scholar] [CrossRef]
- DeLeon-Pennell, K.Y.; Barker, T.H.; Lindsey, M.L. Fibroblasts: The arbiters of extracellular matrix remodeling. Matrix Biol. 2020, 91–92, 1–7. [Google Scholar] [CrossRef]
- Shaker, S.A.; Ayuob, N.N.; Hajrah, N.H. Cell talk: A phenomenon observed in the keloid scar by immunohistochemical study. Appl. Immunohistochem. Mol. Morphol. 2011, 19, 153–159. [Google Scholar] [CrossRef]
- Deng, Z.; Subilia, M.; Chin, I.L.; Hortin, N.; Stevenson, A.W.; Wood, F.M.; Prêle, C.M.; Choi, Y.S.; Fear, M.W. Keloid fibroblasts have elevated and dysfunctional mechanotransduction signaling that is independent of TGF-β. J. Dermatol. Sci. 2021, 104, 11–20. [Google Scholar] [CrossRef]
- Chipev, C.C.; Simman, R.; Hatch, G.; Katz, A.E.; Siegel, D.M.; Simon, M. Myofibroblast phenotype and apoptosis in keloid and palmar fibroblasts in vitro. Cell Death. Differ. 2000, 7, 166–176. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Dong, X.H.; Guan, E.L.; Si, L.B.; Zhuge, R.Q.; Zhao, P.X.; Zhang, X.; Liu, M.Y.; Adzavon, Y.M.; Long, X.; et al. A comparison of apoptosis levels in keloid tissue, physiological scars and normal skin. Am. J. Transl. Res. 2017, 9, 5548–5557. [Google Scholar]
- Tsuchihashi, H.; Hasegawa, T.; Sumiyoshi, K.; Ikeda, S.; Ando, T.; Nakao, A.; Okumura, K.; Ogawa, H. TWEAK inhibits TGF-beta-induced contraction of normal and keloid fibroblast-embedded collagen gel. J. Dermatol. Sci. 2007, 45, 216–218. [Google Scholar] [CrossRef]
- Hasegawa, T.; Nakao, A.; Sumiyoshi, K.; Tsuchihashi, H.; Ogawa, H. SB-431542 inhibits TGF-beta-induced contraction of collagen gel by normal and keloid fibroblasts. J. Dermatol. Sci. 2005, 39, 33–38. [Google Scholar] [CrossRef]
- Bock, O.; Yu, H.; Zitron, S.; Bayat, A.; Ferguson, M.; Mrowietz, U. Studies of Transforming Growth Factors Beta 1–3 and their Receptors I and II in Fibroblast of Keloids and Hypertrophic Scars. Acta Dermato. Venereol. 2005, 85, 216–220. [Google Scholar] [CrossRef]
- Babu, M.; Diegelmann, R.; Oliver, N. Keloid fibroblasts exhibit an altered response to TGF-beta. J. Investig. Dermatol. 1992, 99, 650–655. [Google Scholar] [CrossRef]
- Bettinger, D.A.; Yager, D.R.; Diegelmann, R.F.; Cohen, I.K. The effect of TGF-beta on keloid fibroblast proliferation and collagen synthesis. Plast. Reconstr. Surg. 1996, 98, 827–833. [Google Scholar] [CrossRef]
- Haisa, M.; Okochi, H.; Grotendorst, G.R. Elevated levels of PDGF alpha receptors in keloid fibroblasts contribute to an enhanced response to PDGF. J. Investig. Dermatol. 1994, 103, 560–563. [Google Scholar] [CrossRef]
- Jagadeesan, J.; Bayat, A. Transforming growth factor beta (TGFβ) and keloid disease. Int. J. Surg. 2007, 5, 278–285. [Google Scholar] [CrossRef]
- Deng, C.-C.; Hu, Y.-F.; Zhu, D.-H.; Cheng, Q.; Gu, J.-J.; Feng, Q.-L.; Zhang, L.-X.; Xu, Y.-P.; Wang, D.; Rong, Z.; et al. Single-cell RNA-seq reveals fibroblast heterogeneity and increased mesenchymal fibroblasts in human fibrotic skin diseases. Nat. Commun. 2021, 12, 3709. [Google Scholar] [CrossRef]
- Lim, K.H.; Itinteang, T.; Davis, P.F.; Tan, S.T. Stem Cells in Keloid Lesions: A Review. Plast. Reconstr. Surg. Glob. Open 2019, 7, e2228. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Choi, I.-K.; Lee, J.H.; Kim, Y.O.; Yun, C.-O. A novel three-dimensional model system for keloid study: Organotypic multicellular scar model. Wound Repair Regen. 2013, 21, 155–165. [Google Scholar] [CrossRef]
- Lee, W.J.; Ahn, H.M.; Na, Y.; Wadhwa, R.; Hong, J.; Yun, C.-O. Mortalin deficiency suppresses fibrosis and induces apoptosis in keloid spheroids. Sci. Rep. 2017, 7, 12957. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Ahn, H.M.; Roh, H.; Na, Y.; Choi, I.-K.; Lee, J.H.; Kim, Y.O.; Lew, D.H.; Yun, C.-O. Decorin-expressing adenovirus decreases collagen synthesis and upregulates MMP expression in keloid fibroblasts and keloid spheroids. Exp. Dermatol. 2015, 24, 591–597. [Google Scholar] [CrossRef]
- Białkowska, K.; Komorowski, P.; Bryszewska, M.; Miłowska, K. Spheroids as a Type of Three-Dimensional Cell Cultures-Examples of Methods of Preparation and the Most Important Application. Int. J. Mol. Sci. 2020, 21, 6225. [Google Scholar] [CrossRef]
- Ryu, N.-E.; Lee, S.-H.; Park, H. Spheroid Culture System Methods and Applications for Mesenchymal Stem Cells. Cells 2019, 8, 1620. [Google Scholar] [CrossRef]
- Nath, S.; Devi, G.R. Three-dimensional culture systems in cancer research: Focus on tumor spheroid model. Pharmacol. Ther. 2016, 163, 94–108. [Google Scholar] [CrossRef]
- Lee, N.-H.; Bayaraa, O.; Zechu, Z.; Kim, H.S. Biomaterials-assisted spheroid engineering for regenerative therapy. BMB Rep. 2021, 54, 356–367. [Google Scholar] [CrossRef]
- Tan, Y.; Suarez, A.; Garza, M.; Khan, A.A.; Elisseeff, J.; Coon, D. Human fibroblast-macrophage tissue spheroids demonstrate ratio-dependent fibrotic activity for in vitro fibrogenesis model development. Biomater. Sci. 2020, 8, 1951–1960. [Google Scholar] [CrossRef]
- Granato, G.; Ruocco, M.R.; Iaccarino, A.; Masone, S.; Calì, G.; Avagliano, A.; Russo, V.; Bellevicine, C.; Di Spigna, G.; Fiume, G.; et al. Generation and analysis of spheroids from human primary skin myofibroblasts: An experimental system to study myofibroblasts deactivation. Cell Death Discov. 2017, 3, 17038. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Limandjaja, G.C.; Niessen, F.B.; Scheper, R.J.; Gibbs, S. The Keloid Disorder: Heterogeneity, Histopathology, Mechanisms and Models. Front. Cell Dev. Biol. 2020, 8, 360. [Google Scholar] [CrossRef] [PubMed]
- Supp, D.M.; Hahn, J.M.; Glaser, K.; McFarland, K.L.; Boyce, S.T. Deep and superficial keloid fibroblasts contribute differentially to tissue phenotype in a novel in vivo model of keloid scar. Plast. Reconstr. Surg. 2012, 129, 1259–1271. [Google Scholar] [CrossRef] [PubMed]
- Duong, H.S.; Zhang, Q.; Kobi, A.; Le, A.; Messadi, D.V. Assessment of morphological and immunohistological alterations in long-term keloid skin explants. Cells Tissues Organs 2005, 181, 89–102. [Google Scholar] [CrossRef]
- Bagabir, R.; Syed, F.; Paus, R.; Bayat, A. Long-term organ culture of keloid disease tissue. Exp. Dermatol. 2012, 21, 376–381. [Google Scholar] [CrossRef]
- Alzeeb, G.; Arzur, D.; Trichet, V.; Talagas, M.; Corcos, L.; Le Jossic-Corcos, C. Gastric cancer cell death analyzed by live cell imaging of spheroids. Sci. Rep. 2022, 12, 1488. [Google Scholar] [CrossRef]
- Chen, G.; Liu, W.; Yan, B. Breast Cancer MCF-7 Cell Spheroid Culture for Drug Discovery and Development. J. Cancer Ther. 2022, 13, 117–130. [Google Scholar] [CrossRef]
- Gallegos-Martínez, S.; Lara-Mayorga, I.M.; Samandari, M.; Mendoza-Buenrostro, C.; Flores-Garza, B.G.; Reyes-Cortés, L.M.; Segoviano-Ramírez, J.C.; Zhang, Y.S.; Santiago, G.T.-D.; Álvarez, M.M. Culture of cancer spheroids and evaluation of anti-cancer drugs in 3D-printed miniaturized continuous stirred tank reactors (mCSTRs). Biofabrication 2022, 14, 035007. [Google Scholar] [CrossRef]
- Bochaton-Piallat, M.-L.; Gabbiani, G.; Hinz, B. The myofibroblast in wound healing and fibrosis: Answered and unanswered questions. F1000Research 2016, 5, 752. [Google Scholar] [CrossRef]
- Salmenperä, P.; Karhemo, P.-R.; Räsänen, K.; Laakkonen, P.; Vaheri, A. Fibroblast spheroids as a model to study sustained fibroblast quiescence and their crosstalk with tumor cells. Exp. Cell Res. 2016, 345, 17–24. [Google Scholar] [CrossRef]
- Messadi, D.V.; Doung, H.S.; Zhang, Q.; Kelly, A.P.; Tuan, T.-L.; Reichenberger, E.; Le, A.D. Activation of NFkappaB signal pathways in keloid fibroblasts. Arch. Dermatol. Res. 2004, 296, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Chodon, T.; Sugihara, T.; Igawa, H.H.; Funayama, E.; Furukawa, H. Keloid-derived fibroblasts are refractory to Fas-mediated apoptosis and neutralization of autocrine transforming growth factor-beta1 can abrogate this resistance. Am. J. Pathol. 2000, 157, 1661–1669. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, H.; Yoshimoto, H.; Fujioka, M.; Murakami, R.; Hirano, A.; Fujii, T.; Ohtsuru, A.; Namba, H.; Yamashita, S. Keloid Fibroblasts Resist Ceramide-Induced Apoptosis by Overexpression of Insulin-Like Growth Factor I Receptor. J. Investig. Dermatol. 2000, 115, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Lei, R.; Li, J.; Liu, F.; Li, W.; Zhang, S.; Wang, Y.; Chu, X.; Xu, J. HIF-1α promotes the keloid development through the activation of TGF-β/Smad and TLR4/MyD88/NF-κB pathways. Cell Cycle 2019, 18, 3239–3250. [Google Scholar] [CrossRef]
- Kang, Y.; Roh, M.R.; Rajadurai, S.; Rajadurai, A.; Kumar, R.; Njauw, C.-N.; Zheng, Z.; Tsao, H. Hypoxia and HIF-1α Regulate Collagen Production in Keloids. J. Investig. Dermatol. 2020, 140, 2157–2165. [Google Scholar] [CrossRef]
- Xin, Y.; Wang, X.; Zhu, M.; Qu, M.; Bogari, M.; Lin, L.; Aung, Z.M.; Chen, W.; Chen, X.; Chai, G.; et al. Expansion of CD26 positive fibroblast population promotes keloid progression. Exp. Cell Res. 2017, 356, 104–113. [Google Scholar] [CrossRef]
- Ohm, B.; Moneke, I.; Jungraithmayr, W. Targeting cluster of differentiation 26/dipeptidyl peptidase 4 (CD26/DPP4) in organ fibrosis. Br. J. Pharmacol. 2022; early view. [Google Scholar] [CrossRef]
- Huang, X.; Khoong, Y.; Han, C.; Su, D.; Ma, H.; Gu, S.; Li, Q.; Zan, T. Targeting Dermal Fibroblast Subtypes in Antifibrotic Therapy: Surface Marker as a Cellular Identity or a Functional Entity? Front. Physiol. 2022, 12, 694605. Available online: https://www.frontiersin.org/article/10.3389/fphys.2021.694605 (accessed on 30 May 2022). [CrossRef]
- Lu, Y.-Y.; Fang, C.-C.; Hong, C.-H.; Wu, C.-H.; Lin, Y.-H.; Chang, K.-L.; Lee, C.-H. Nonmuscle Myosin II Activation Regulates Cell Proliferation, Cell Contraction, and Myofibroblast Differentiation in Keloid-Derived Fibroblasts. Adv. Wound Care 2020, 9, 491–501. [Google Scholar] [CrossRef]
- Smith, J.C.; Boone, B.E.; Opalenik, S.R.; Williams, S.M.; Russell, S.B. Gene profiling of keloid fibroblasts shows altered expression in multiple fibrosis-associated pathways. J. Investig. Dermatol. 2008, 128, 1298–1310. [Google Scholar] [CrossRef]
- Chen, Z.; Gao, Z.; Xia, L.; Wang, X.; Lu, L.; Wu, X. Dysregulation of DPP4-CXCL12 Balance by TGF-β1/SMAD Pathway Promotes CXCR4+ Inflammatory Cell Infiltration in Keloid Scars. J. Inflamm. Res. 2021, 14, 4169–4180. [Google Scholar] [CrossRef] [PubMed]
- Kunz-Schughart, L.A.; Wenninger, S.; Neumeier, T.; Seidl, P.; Knuechel, R. Three-dimensional tissue structure affects sensitivity of fibroblasts to TGF-β1. Am. J. Physiol. Cell Physiol. 2003, 284, C209–C219. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Sex | Age | Phototype |
|---|---|---|---|
| Earlobe | F | 22 | III |
| Earlobe | F | 22 | II |
| Earlobe | F | 19 | II |
| Earlobe | F | 54 | III |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dirand, Z.; Tissot, M.; Chatelain, B.; Viennet, C.; Rolin, G. Is Spheroid a Relevant Model to Address Fibrogenesis in Keloid Research? Biomedicines 2023, 11, 2350. https://doi.org/10.3390/biomedicines11092350
Dirand Z, Tissot M, Chatelain B, Viennet C, Rolin G. Is Spheroid a Relevant Model to Address Fibrogenesis in Keloid Research? Biomedicines. 2023; 11(9):2350. https://doi.org/10.3390/biomedicines11092350
Chicago/Turabian StyleDirand, Zélie, Marion Tissot, Brice Chatelain, Céline Viennet, and Gwenaël Rolin. 2023. "Is Spheroid a Relevant Model to Address Fibrogenesis in Keloid Research?" Biomedicines 11, no. 9: 2350. https://doi.org/10.3390/biomedicines11092350
APA StyleDirand, Z., Tissot, M., Chatelain, B., Viennet, C., & Rolin, G. (2023). Is Spheroid a Relevant Model to Address Fibrogenesis in Keloid Research? Biomedicines, 11(9), 2350. https://doi.org/10.3390/biomedicines11092350