

Effect of Bariatric Surgery on Plasma Cell-Free Mitochondrial DNA, Insulin Sensitivity and Metabolic Changes in Obese Patients
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Metabolic Parameters and Muscle Biopsy
2.3. Isolation of Plasma and PBM Cells
2.4. Isolation of Plasma cf-DNA
2.5. Analysis of Plasma cf-mtDNA
2.6. Analysis of mtDNA Content
2.7. Statistical Analyses
3. Results
3.1. Metabolic Parameters
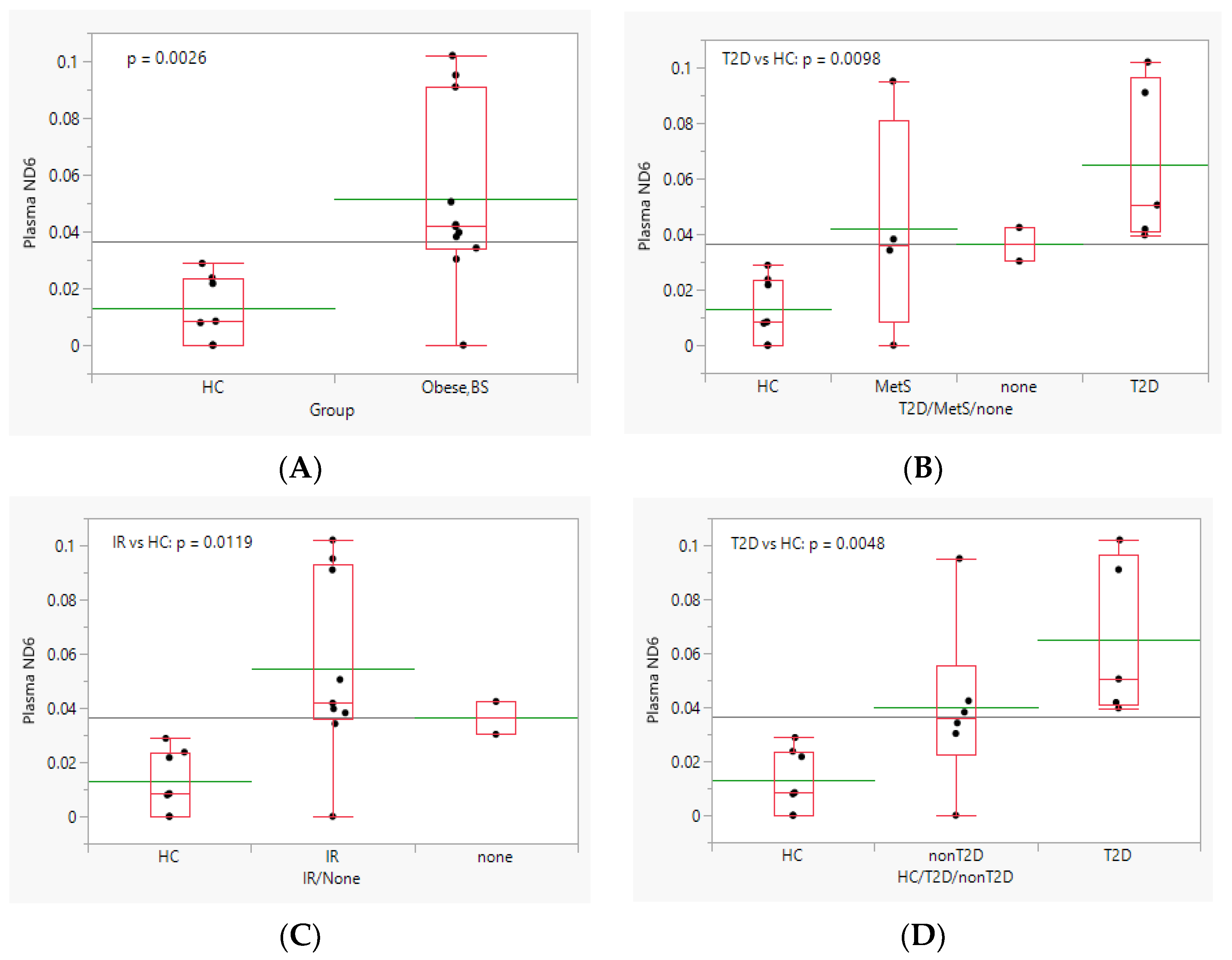
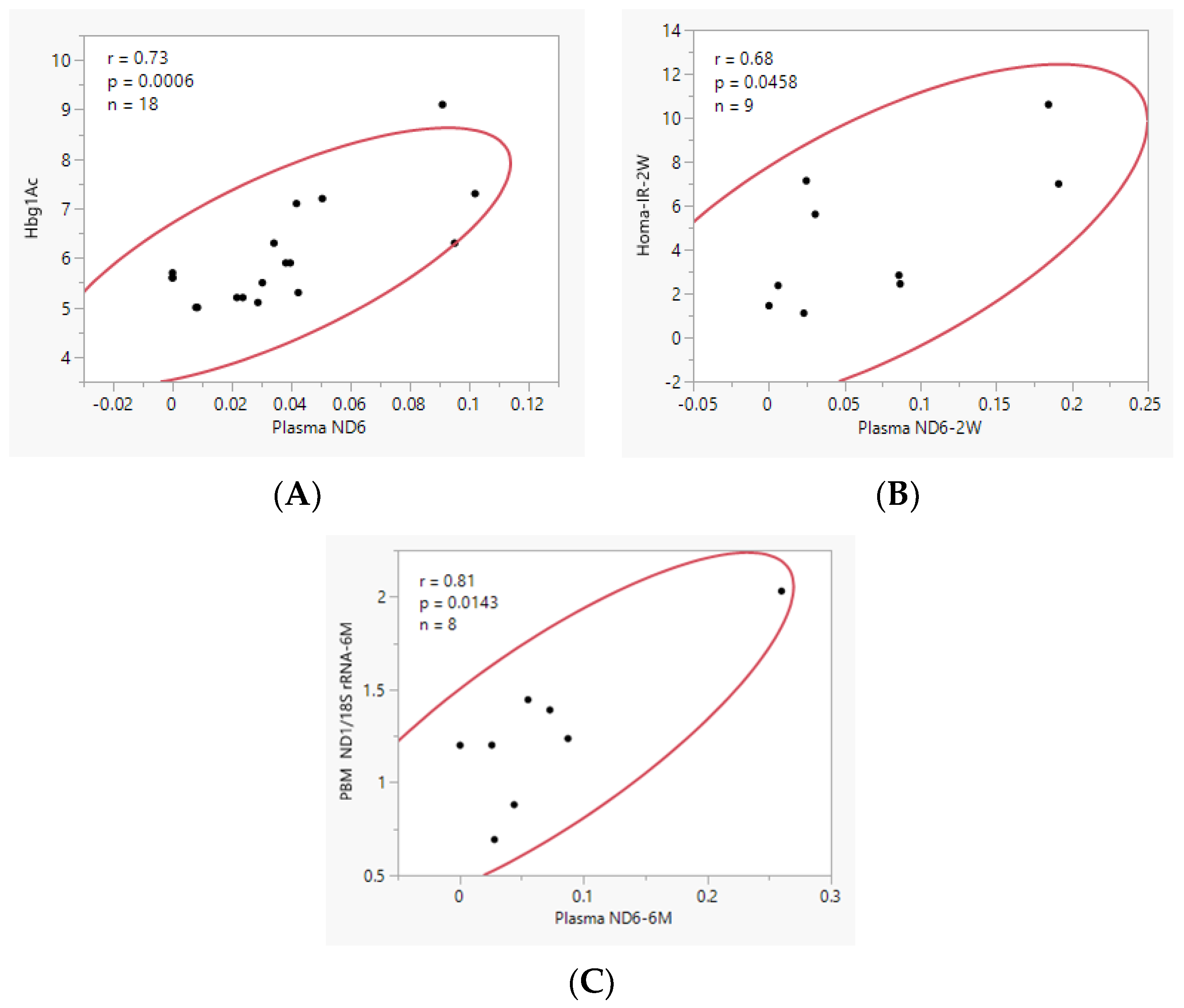
3.2. Effect of Obesity and Bariatric Surgery on Plasma cf-mtDNA
3.3. Effect of Obesity and Bariatric Surgery on mtDNA Content in PBM Cells and Skeletal Muscle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Reaven, G.M. Insulin resistance: The link between obesity and cardiovascular disease. Med. Clin. N. Am. 2011, 95, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Hruby, A.; Manson, J.A.E.; Qi, L.; Malik, V.S.; Rimm, E.B.; Sun, Q.; Walter, C.W.; Frank, B.H. Determinants and consequences of obesity. Am. J. Public Health 2016, 106, 1656–1662. [Google Scholar] [CrossRef]
- Bonnard, C.; Durand, A.; Peyrol, S.; Chanseaume, E.; Chauvin, M.A.; Morio, B.; Vidal, H.; Rieusset, J. Mitochondrial dysfunction results from oxidative stress in the skeletal muscle of diet-induced insulin-resistant mice. J. Clin. Investig. 2008, 118, 789–800. [Google Scholar] [CrossRef]
- Anderson, E.J.; Lustig, M.E.; Boyle, K.E.; Woodlief, T.L.; Kane, D.A.; Lin, C.T.; Price, J.W., III; Kang, L.; Rabinovitch, P.S.; Szeto, H.H.; et al. Mitochondrial H2O2 emission and cellular redox state link excess fat intake to insulin resistance in both rodents and humans. J. Clin. Investig. 2009, 119, 573–581. [Google Scholar] [CrossRef]
- Hoehn, K.L.; Salmon, A.B.; Hohnen-Behrens, C.; Turner, N.; Hoy, A.J.; Maghzal, G.J.; Stocker, R.; Van Remmen, H.; Kraegen, E.W.; Cooney, G.J.; et al. Insulin resistance is a cellular antioxidant defense mechanism. Proc. Natl. Acad. Sci. USA 2009, 106, 17787–17792. [Google Scholar] [CrossRef] [PubMed]
- Nijhawan, S.; Richards, W.; O’Hea, M.F.; Audia, J.P.; Alvarez, D.F. Bariatric surgery rapidly improves mitochondrial respiration in morbidly obese patients. Surg. Endosc. 2013, 27, 4569–4573. [Google Scholar] [CrossRef]
- Fernstrom, M.; Bakkman, L.; Loogna, P.; Rooyackers, O.; Svensson, M.; Jakobsson, T.; Brandt, L.; Lagerros, Y.T. Improved Muscle Mitochondrial Capacity Following Gastric Bypass Surgery in Obese Subjects. Obes. Surg. 2016, 26, 1391–1397. [Google Scholar] [CrossRef]
- Toledo, F.G.; Menshikova, E.V.; Ritov, V.B.; Azuma, K.; Radikova, Z.; DeLany, J.; Kelley, D.E. Effects of physical activity and weight loss on skeletal muscle mitochondria and relationship with glucose control in type 2 diabetes. Diabetes 2007, 56, 2142–2147. [Google Scholar] [CrossRef]
- Lee, H.; Oh, S.; Yang, W.; Park, R.; Kim, H.; Jeon, J.S.; Noh, H.; Han, D.C.; Cho, K.W.; Kim, Y.J.; et al. Bariatric surgery reduces elevated urinary mitochondrial DNA copy number in patients with obesity. J. Clin. Endocrinol. Metab. 2019, 104, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Kim, H.; Noh, H.; Jeon, J.S.; Byun, D.W.; Kim, S.H.; Kim, H.J.; Suh, K.; Park, H.K.; Kwon, S.H. Effect of bariatric surgery on circulating and urinary mitochondrial DNA copy numbers in obesity with or without diabetes. BMJ Open Diabetes Res. Care 2020, 8, e001372. [Google Scholar] [CrossRef]
- Chandru, S.; Prabhu, P.; Balasubramanyam, M.; Subhashini, R.; Tiwaskar, M.; Pramodkumar, T.A.; Pradeepa, R.; Anjana, R.M.; Mohan, V. Beneficial Primary Outcomes of Metabolic Surgery with Changes in Telomere Length and Mitochondrial DNA in Obese Asian Indians with Dysglycemia. J. Assoc. Physicians Ind. 2021, 69, 11–12. [Google Scholar]
- Skuratovskaia, D.; Litvinova, L.; Vulf, M.; Zatolokin, P.; Popadin, K.; Mazunin, I. From normal to obesity and back: The associations between mitochondrial DNA copy number, gender, and body mass index. Cells 2019, 8, 430. [Google Scholar] [CrossRef]
- Pedersen, J.S.; Rygg, M.O.; Chrøis, K.; Sustarsic, E.G.; Gerhart-Hines, Z.; Wever Albrechtsen, N.J.; Serizawa, R.R.; Kristiansen, V.B.; Basse, A.L.; Boilesen, A.E.B.; et al. Influence of NAFLD and bariatric surgery on hepatic and adipose tissue mitochondrial biogenesis and respiration. Nat. Commun. 2022, 13, 2931. [Google Scholar] [CrossRef] [PubMed]
- Yuzefovych, L.V.; Pastukh, V.M.; Ruchko, M.V.; Simmons, J.D.; Richards, W.O.; Rachek, L.I. Plasma mitochondrial DNA is elevated in obese type 2 diabetes patients and correlates positively with insulin resistance. PLoS ONE 2019, 14, e0222278. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.L. A comprehensive definition for metabolic syndrome. Dis. Model Mech. 2009, 2, 231–237. [Google Scholar] [CrossRef]
- Simmons, J.D.; Lee, Y.L.; Mulekar, S.; Kuck, J.L.; Brevard, S.B.; Gonzalez, R.P.; Gillespie, M.N.; Richards, W.O. Elevated levels of plasma mitochondrial DNA DAMPs are linked to clinical outcome in severely injured human subjects. Ann. Surg. 2013, 258, 591–596. [Google Scholar] [CrossRef]
- Dankel, S.N.; Staalesen, V.; Bjørndal, B.; Berge, R.K.; Mellgren, G.; Burri, L. Tissue-specific effects of bariatric surgery including mitochondrial function. J. Obes. 2011, 2011, 435245. [Google Scholar] [CrossRef]
- Gancheva, S.; Ouni, M.; Jelenik, T.; Koliaki, C.; Szendroedi, J.; Toledo, F.G.S.; Markgraf, D.F.; Pesta, D.H.; Mastrototaro, L.; De Filippo, E.; et al. Dynamic changes of muscle insulin sensitivity after metabolic surgery. Nat. Commun. 2019, 10, 4179. [Google Scholar] [CrossRef] [PubMed]
- Abad-Jiménez, Z.; López-Domènech, S.; García-Gargallo, C.; Vezza, T.; Gómez-Abril, S.Á.; Morillas, C.; Díaz-Pozo, P.; Falcón, R.; Bañuls, C.; Víctor, V.M.; et al. Roux-en-Y Gastric Bypass Modulates AMPK, Autophagy and Inflammatory Response in Leukocytes of Obese Patients. Biomedicines 2022, 10, 430. [Google Scholar] [CrossRef]
- Liu, J.; Zou, Y.; Tang, Y.; Xi, M.; Xie, L.; Zhang, Q.; Gong, J. Circulating cell-free mitochondrial deoxyribonucleic acid is increased in coronary heart disease patients with diabetes mellitus. J. Diabetes Investig. 2016, 7, 109–114. [Google Scholar] [CrossRef]
- Celec, P.; Janovičová, Ĺ.; Gurecká, R.; Koborová, I.; Gardlík, R.; Šebeková, K. Circulating extracellular DNA is in association with continuous metabolic syndrome score in healthy adolescents. Physiol. Genom. 2021, 53, 309–318. [Google Scholar] [CrossRef]
- Jylhävä, J.; Lehtimäki, T.; Jula, A.; Moilanen, L.; Kesäniemi, Y.A.; Nieminen, M.S.; Kähönen, M.; Hurme, M. Circulating cell free DNA is associated with cardiometabolic risk factors: The health 2000 survey. Atherosclerosis 2014, 233, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Camuzi Zovico, P.V.; Gasparini Neto, V.H.; Venâncio, F.A.; Soares Miguel, G.P.; Graça Pedrosa, R.; Kenji Haraguchi, F.; Barauna, V.G. Cell-free DNA as an obesity biomarker. Physiol. Res. 2020, 69, 515–520. [Google Scholar] [CrossRef] [PubMed]
- Kesner, E.E.; Saada-Reich, A.; Lorberboum-Galski, H. Characteristics of Mitochondrial Transformation into Human Cells. Sci. Rep. 2016, 6, 26057. [Google Scholar] [CrossRef]
- Kim, M.J.; Hwang, J.W.; Yun, C.K.; Lee, Y.; Choi, Y.S. Delivery of exogenous mitochondria via centrifugation enhances cellular metabolic function. Sci. Rep. 2018, 8, 3330. [Google Scholar] [CrossRef]
- Berridge, M.V.; McConnell, M.J.; Grasso, C.; Bajzikova, M.; Kovarova, J.; Neuzil, J. Horizontal transfer of mitochondria between mammalian cells: Beyond co-culture approaches. Curr. Opin. Genet. Dev. 2016, 38, 75–82. [Google Scholar] [CrossRef]
- Alexander, J.F.; Seua, A.V.; Arroyo, L.D.; Ray, P.R.; Wangzhou, A.; Heibeta-Luckemann, L.; Schedlowski, M.; Price, T.J.; Kavelaars, A.; Heijnen, C.J. Nasal administration of mitochondria reverses chemotherapy-induced cognitive deficits. Theranostics 2021, 11, 3109–3130. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C.; Wu, S.L.; Liu, K.H.; Chen, Y.H.; Chuang, C.S.; Cheng, F.C.; Su, H.L.; Wei, Y.H.; Kuo, S.J.; Liu, C.S. Allogeneic/xenogeneic transplantation of peptide-labeled mitochondria in Parkinson’s disease: Restoration of mitochondria functions and attenuation of 6-hydroxydopamine-induced neurotoxicity. Transl. Res. 2016, 170, 40–56.e3. [Google Scholar] [CrossRef]
- Valenti, D.; Vacca, R.A.; Moro, L.; Atlante, A. Mitochondria Can Cross Cell Boundaries: An Overview of the Biological Relevance, Pathophysiological Implications and Therapeutic Perspectives of Intercellular Mitochondrial Transfer. Int. J. Mol. Sci. 2021, 22, 8312. [Google Scholar] [CrossRef]
- Lee, S.-E.; Kang, Y.C.; Kim, Y.; Kim, S.; Yu, S.-H.; Park, J.H.; Kim, I.-H.; Kim, H.-Y.; Han, K.; Lee, H.K.; et al. Preferred Migration of Mitochondria toward Cells and Tissues with Mitochondrial Damage. Int. J. Mol. Sci. 2022, 23, 15734. [Google Scholar] [CrossRef]
- Al Amir Dache, Z.; Otandault, A.; Tanos, R.; Pastor, B.; Meddeb, R.; Sanchez, C.; Arena, G.; Lasorsa, L.; Bennett, A.; Grange, T.; et al. Blood contains circulating cell-free respiratory competent mitochondria. FASEB J. 2020, 34, 3616–3630. [Google Scholar] [CrossRef]
- Sansone, P.; Savini, C.; Kurelac, I.; Chang, Q.; Amato, L.B.; Strillacci, A.; Stepanova, A.; Iommarini, L.; Mastroleo, C.; Daly, L.; et al. Packaging and transfer of mitochondrial DNA via exosomes regulate escape from dormancy in hormonal therapy-resistant breast cancer. Proc. Natl. Acad. Sci. USA 2017, 114, E9066–E9075. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, N.M.; Mikhalchenko, A.; Ma, H.; Koski, A.; Li, Y.; Van Dyken, C.; Tippner-Hedges, R.; Yoon, D.; Liang, D.; Hayama, T.; et al. Horizontal mtDNA transfer between cells is common during mouse development. iScience 2022, 25, 103901. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.; Shirihai, O.S. Mitochondrial signal transduction. Cell Metab. 2022, 34, 1620–1653. [Google Scholar] [CrossRef]
- Litvinova, L.; Zatolokin, P.; Vulf, M.; Mazunin, I.; Skuratovskaia, D. The relationship between the mtDNA copy number in insulin-dependent tissues and markers of endothelial dysfunction and inflammation in obese patients. BMC Med. Genom. 2019, 12, 41. [Google Scholar] [CrossRef]
- Hepokoski, M.L.; Odish, M.; Lam, M.T.; Coufal, N.G.; Rolfsen, M.L.; Shadel, G.S.; Moyzis, A.G.; Sainz, A.G.; Takiar, P.G.; Patel, S.; et al. Absolute quantification of plasma mitochondrial DNA by droplet digital PCR marks COVID-19 severity over time during intensive care unit admissions. Am. J. Physiol. Lung Cell. Mol. Physiol. 2022, 323, L84–L92. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Obese Only | Time | n | Mean | Std Dev | R2 | p (Patient) | p (Time) | |||
|---|---|---|---|---|---|---|---|---|---|---|
| BMI | BS | 13 | 49.90 | 7.91 | 0.88 | <0.0001 | <0.0001 | A | ||
| 2 W | 12 | 44.49 | 7.59 | B | ||||||
| 3 M | 10 | 39.54 | 6.99 | C | ||||||
| 6 M | 10 | 36.61 | 6.62 | C | ||||||
| Glucose | BS | 13 | 125.31 | 43.64 | 0.78 | <0.0001 | 0.1317 | |||
| 2 W | 12 | 117.17 | 72.08 | |||||||
| 3 M | 10 | 89.30 | 9.68 | |||||||
| 6 M | 10 | 85.30 | 20.24 | |||||||
| HgbA1c | BS | 13 | 7.04 | 1.73 | 0.89 | 0.0023 | 0.0151 | A | ||
| 2 W | 0 | |||||||||
| 3 M | 8 | 5.39 | 0.31 | B | ||||||
| 6 M | 7 | 5.30 | 0.29 | B | ||||||
| Insulin | BS | 12 | 50.29 | 42.57 | 0.56 | 0.3424 | 0.0034 | A | ||
| 2 W | 12 | 16.35 | 10.14 | B | ||||||
| 3 M | 10 | 10.58 | 4.24 | B | ||||||
| 6 M | 10 | 14.40 | 14.90 | B | ||||||
| HOMA-IR | BS | 12 | 18.66 | 22.50 | 0.59 | 0.0780 | 0.0126 | A | ||
| 2 W | 12 | 4.52 | 3.07 | B | ||||||
| 3 M | 10 | 2.40 | 1.16 | B | ||||||
| 6 M | 10 | 3.58 | 5.01 | A | B | |||||
| Triglycerides | BS | 13 | 159.85 | 77.95 | 0.92 | 0.0002 | 0.0026 | A | ||
| 2 W | 0 | |||||||||
| 3 M | 8 | 99.50 | 13.77 | B | ||||||
| 6 M | 7 | 91.57 | 31.97 | B | ||||||
| Total Cholesterol | BS | 13 | 167.23 | 31.93 | 0.75 | 0.0213 | 0.9287 | |||
| 2 W | 0 | |||||||||
| 3 M | 8 | 168.13 | 27.53 | |||||||
| 6 M | 7 | 169.00 | 28.61 | |||||||
| HDL | BS | 13 | 40.08 | 6.91 | 0.91 | 0.0002 | 0.0113 | B | ||
| 2 W | 0 | |||||||||
| 3 M | 8 | 43.88 | 11.70 | B | ||||||
| 6 M | 7 | 51.57 | 14.59 | A | ||||||
| LDL | BS | 13 | 95.08 | 29.76 | 0.69 | 0.0773 | 0.8800 | |||
| 2 W | 0 | |||||||||
| 3 M | 8 | 104.44 | 20.80 | |||||||
| 6 M | 7 | 99.14 | 12.62 | |||||||
| Plasma ND6 | BS | 6 | 0.06 | 0.03 | 0.66 | 0.0443 | 0.8539 | |||
| 2 W | 9 | 0.07 | 0.07 | |||||||
| 3 M | 7 | 0.08 | 0.06 | |||||||
| 6 M | 8 | 0.07 | 0.08 | |||||||
| Skeletal muscle ND1/18SrRNA | BS | 13 | 1.74 | 1.06 | 0.69 | 0.3104 | 0.0700 | |||
| 2 W | 0 | |||||||||
| 3 M | 9 | 1.14 | 0.42 | |||||||
| 6 M | 2 | 1.34 | 0.61 | |||||||
| PBM ND1/18S rRNA | BS | 13 | 0.97 | 0.28 | 0.39 | 0.3311 | 0.3759 | |||
| 2 W | 12 | 0.85 | 0.32 | |||||||
| 3 M | 10 | 0.87 | 0.46 | |||||||
| 6 M | 11 | 1.16 | 0.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuzefovych, L.V.; Pastukh, V.M.; Mulekar, M.S.; Ledbetter, K.; Richards, W.O.; Rachek, L.I. Effect of Bariatric Surgery on Plasma Cell-Free Mitochondrial DNA, Insulin Sensitivity and Metabolic Changes in Obese Patients. Biomedicines 2023, 11, 2514. https://doi.org/10.3390/biomedicines11092514
Yuzefovych LV, Pastukh VM, Mulekar MS, Ledbetter K, Richards WO, Rachek LI. Effect of Bariatric Surgery on Plasma Cell-Free Mitochondrial DNA, Insulin Sensitivity and Metabolic Changes in Obese Patients. Biomedicines. 2023; 11(9):2514. https://doi.org/10.3390/biomedicines11092514
Chicago/Turabian StyleYuzefovych, Larysa V., Viktor M. Pastukh, Madhuri S. Mulekar, Kate Ledbetter, William O. Richards, and Lyudmila I. Rachek. 2023. "Effect of Bariatric Surgery on Plasma Cell-Free Mitochondrial DNA, Insulin Sensitivity and Metabolic Changes in Obese Patients" Biomedicines 11, no. 9: 2514. https://doi.org/10.3390/biomedicines11092514
APA StyleYuzefovych, L. V., Pastukh, V. M., Mulekar, M. S., Ledbetter, K., Richards, W. O., & Rachek, L. I. (2023). Effect of Bariatric Surgery on Plasma Cell-Free Mitochondrial DNA, Insulin Sensitivity and Metabolic Changes in Obese Patients. Biomedicines, 11(9), 2514. https://doi.org/10.3390/biomedicines11092514