
MicroRNA-21 Silencing in Diabetic Nephropathy: Insights on Therapeutic Strategies



Abstract
:1. Introduction
2. Development of DN and Current Therapeutics
3. MicroRNAs in Diabetic Nephropathy
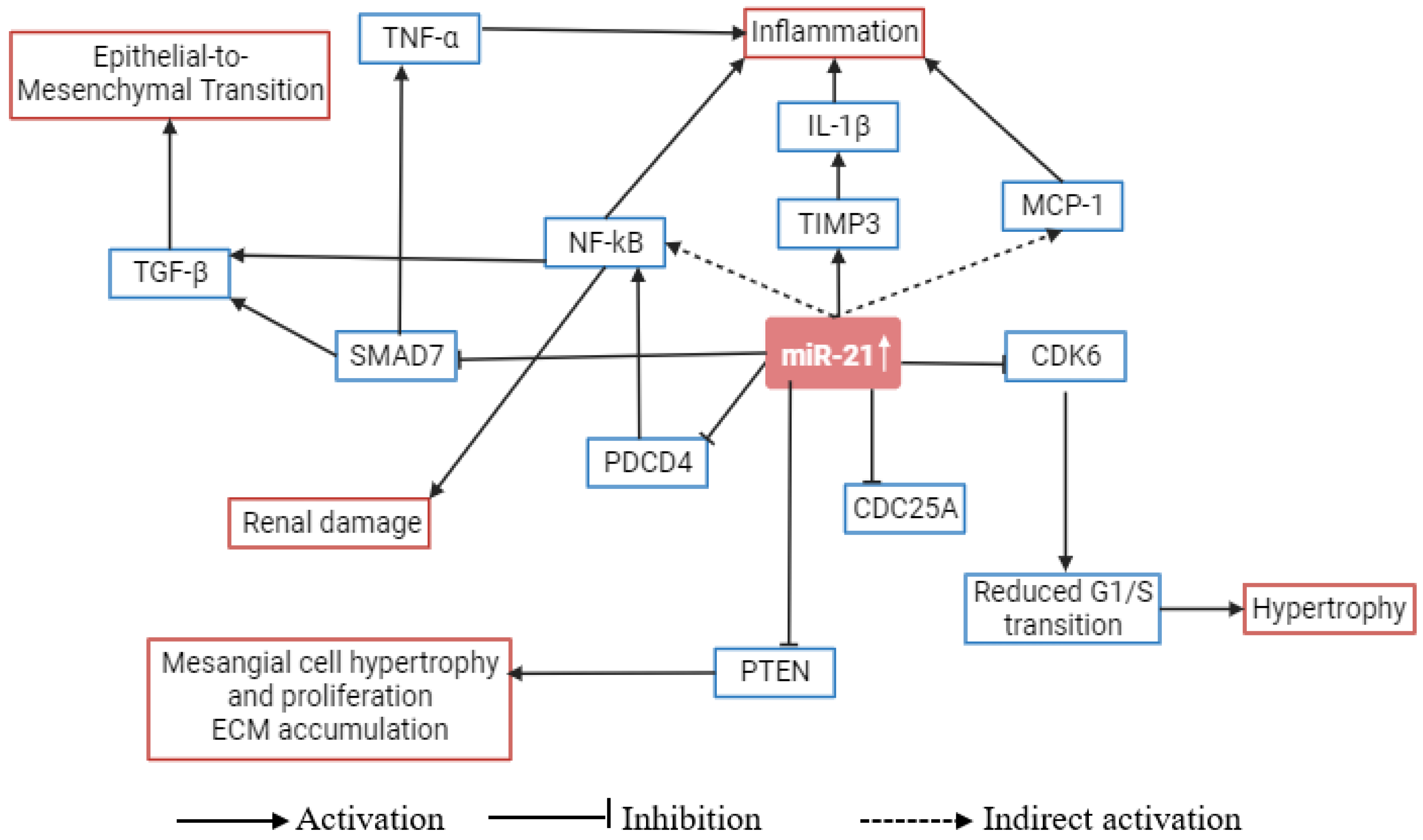
4. Role of microRNA-21 in Metabolic Pathways Related to Diabetic Nephropathy
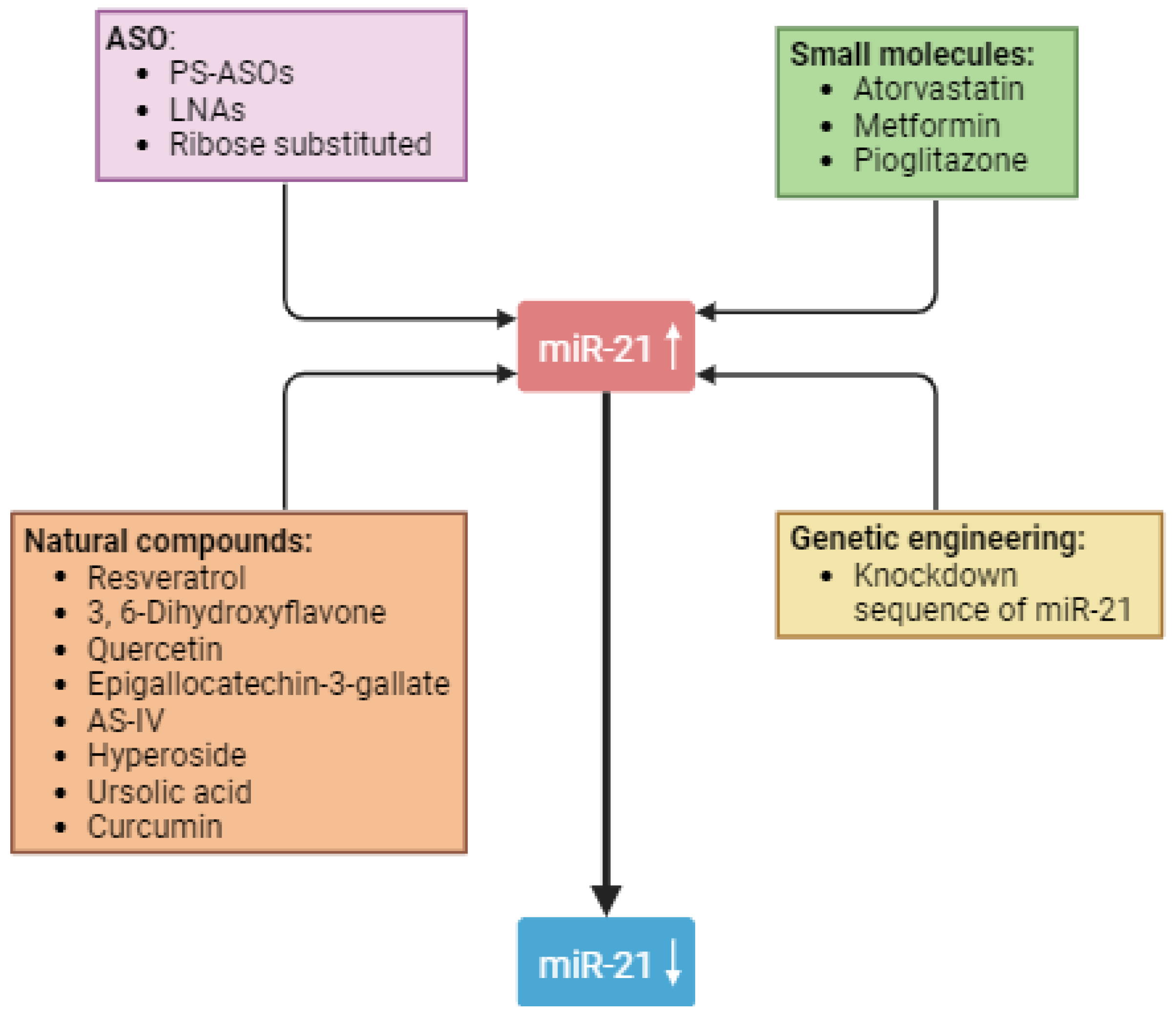
5. Strategies to Use miR-21 as a Therapeutic Target in Diabetic Nephropathy
5.1. Antisense Oligonucleotides
5.2. Natural Compounds
5.3. Small Molecules
5.4. Genetic Engineering
6. Future Possibilities in miRNA-Based Therapy
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Abarca-Gómez, L.; Abdeen, Z.A.; Hamid, Z.A.; Abu-Rmeileh, N.M.; Acosta-Cazares, B.; Acuin, C.; Adams, R.J.; Aekplakorn, W.; Afsana, K.; Aguilar-Salinas, C.A.; et al. Worldwide Trends in Body-Mass Index, Underweight, Overweight, and Obesity from 1975 to 2016: A Pooled Analysis of 2416 Population-Based Measurement Studies in 128·9 Million Children, Adolescents, and Adults. Lancet 2017, 390, 2627–2642. [Google Scholar] [CrossRef]
- Koye, D.N.; Magliano, D.J.; Nelson, R.G.; Pavkov, M.E. The Global Epidemiology of Diabetes and Kidney Disease. Adv. Chronic Kidney Dis. 2018, 25, 121–132. [Google Scholar] [CrossRef]
- Manne-Goehler, J.; Atun, R.; Stokes, A.; Goehler, A.; Houinato, D.; Houehanou, C.; Hambou, M.M.S.; Mbenza, B.L.; Sobngwi, E.; Balde, N.; et al. Diabetes Diagnosis and Care in Sub-Saharan Africa: Pooled Analysis of Individual Data from 12 Countries. Lancet Diabetes Endocrinol. 2016, 4, 903–912. [Google Scholar] [CrossRef]
- de Boer, I.H.; Caramori, M.L.; Chan, J.C.N.; Heerspink, H.J.L.; Hurst, C.; Khunti, K.; Liew, A.; Michos, E.D.; Navaneethan, S.D.; Olowu, W.A.; et al. Executive Summary of the 2020 KDIGO Diabetes Management in CKD Guideline: Evidence-Based Advances in Monitoring and Treatment. Kidney Int. 2020, 98, 839–848. [Google Scholar] [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, Regional and Country-Level Diabetes Prevalence Estimates for 2021 and Projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Chen, L.; Magliano, D.J.; Zimmet, P.Z. The Worldwide Epidemiology of Type 2 Diabetes Mellitus—Present and Future Perspectives. Nat. Rev. Endocrinol. 2012, 8, 228–236. [Google Scholar] [CrossRef]
- Shaw, J.E.; Sicree, R.A.; Zimmet, P.Z. Global Estimates of the Prevalence of Diabetes for 2010 and 2030. Diabetes Res. Clin. Pract. 2010, 87, 4–14. [Google Scholar] [CrossRef]
- Piwkowska, A.; Zdrojewski, Ł.; Heleniak, Z.; Dębska-Ślizień, A. Novel Markers in Diabetic Kidney Disease—Current State and Perspectives. Diagnostics 2022, 12, 1205. [Google Scholar] [CrossRef]
- Tang, J.; Yao, D.; Yan, H.; Chen, X.; Wang, L.; Zhan, H. The Role of MicroRNAs in the Pathogenesis of Diabetic Nephropathy. Int. J. Endocrinol. 2019, 2019, 8719060. [Google Scholar] [CrossRef]
- Liu, S.; Wu, W.; Liao, J.; Tang, F.; Gao, G.; Peng, J.; Fu, X.; Zhan, Y.; Chen, Z.; Xu, W.; et al. MicroRNA-21: A Critical Pathogenic Factor of Diabetic Nephropathy. Front. Endocrinol. 2022, 13, 895010. [Google Scholar] [CrossRef]
- Thomas, M.C.; Brownlee, M.; Susztak, K.; Sharma, K.; Jandeleit-Dahm, K.A.M.; Zoungas, S.; Rossing, P.; Groop, P.-H.; Cooper, M.E. Diabetic Kidney Disease. Nat. Rev. Dis. Primers 2015, 1, 15018. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, C.E.; Christensen, C.K.; Vittinghus, E. The Stages in Diabetic Renal Disease. With Emphasis on the Stage of Incipient Diabetic Nephropathy. Diabetes 1983, 32 (Suppl. S2), 64–78. [Google Scholar] [CrossRef] [PubMed]
- Rezazadeh-Gavgani, E.; Oladghaffari, M.; Bahramian, S.; Majidazar, R.; Dolati, S. MicroRNA-21: A Critical Underestimated Molecule in Diabetic Retinopathy. Gene 2023, 859, 147212. [Google Scholar] [CrossRef]
- Fan, B.; Chopp, M.; Zhang, Z.G.; Liu, X.S. Emerging Roles of MicroRNAs as Biomarkers and Therapeutic Targets for Diabetic Neuropathy. Front. Neurol. 2020, 11, 558758. [Google Scholar] [CrossRef]
- Chawla, A.; Chawla, R.; Jaggi, S. Microvasular and Macrovascular Complications in Diabetes Mellitus: Distinct or Continuum? Indian J. Endocrinol. Metab. 2016, 20, 546. [Google Scholar] [CrossRef]
- Lundbæk, K. Diabetic Angiopathy: A Specific Vascular Disease. Lancet 1954, 263, 377–379. [Google Scholar] [CrossRef]
- Root, H.F.; Pote, W.H.; Frehner, H. Triopathy of Diabetes; Sequence of Neuropathy, Retinopathy, and Nephropathy in One Hundred Fifty-Five Patients. AMA Arch. Intern. Med. 1954, 94, 931–941. [Google Scholar] [CrossRef]
- Stehouwer, C.D.A. Endothelial Dysfunction in Diabetic Nephropathy: State of the Art and Potential Significance for Non-Diabetic Renal Disease. Nephrol. Dial. Transplant. 2004, 19, 778–781. [Google Scholar] [CrossRef]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Simpson, K.; Wonnacott, A.; Fraser, D.J.; Bowen, T. MicroRNAs in Diabetic Nephropathy: From Biomarkers to Therapy. Curr. Diabetes Rep. 2016, 16, 35. [Google Scholar] [CrossRef]
- Zhong, X.; Chung, A.C.K.; Chen, H.Y.; Dong, Y.; Meng, X.M.; Li, R.; Yang, W.; Hou, F.F.; Lan, H.Y. MiR-21 Is a Key Therapeutic Target for Renal Injury in a Mouse Model of Type 2 Diabetes. Diabetologia 2013, 56, 663–674. [Google Scholar] [CrossRef]
- Gomez, I.G.; MacKenna, D.A.; Johnson, B.G.; Kaimal, V.; Roach, A.M.; Ren, S.; Nakagawa, N.; Xin, C.; Newitt, R.; Pandya, S.; et al. Anti-MicroRNA-21 Oligonucleotides Prevent Alport Nephropathy Progression by Stimulating Metabolic Pathways. J. Clin. Investig. 2015, 125, 141–156. [Google Scholar] [CrossRef]
- Kölling, M.; Kaucsar, T.; Schauerte, C.; Hübner, A.; Dettling, A.; Park, J.-K.; Busch, M.; Wulff, X.; Meier, M.; Scherf, K.; et al. Therapeutic MiR-21 Silencing Ameliorates Diabetic Kidney Disease in Mice. Mol. Ther. 2017, 25, 165–180. [Google Scholar] [CrossRef] [PubMed]
- Gembillo, G.; Ingrasciotta, Y.; Crisafulli, S.; Luxi, N.; Siligato, R.; Santoro, D.; Trifirò, G. Kidney Disease in Diabetic Patients: From Pathophysiology to Pharmacological Aspects with a Focus on Therapeutic Inertia. Int. J. Mol. Sci. 2021, 22, 4824. [Google Scholar] [CrossRef] [PubMed]
- Sheen, Y.-J.; Sheu, W.H. Risks of Rapid Decline Renal Function in Patients with Type 2 Diabetes. World J. Diabetes 2014, 5, 835–846. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Jin, L.; Zhang, F.; Sarnow, P.; Kay, M.A. Biological Basis for Restriction of MicroRNA Targets to the 3′ Untranslated Region in Mammalian MRNAs. Nat. Struct. Mol. Biol. 2009, 16, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, H.; Gross, N.; Wei, D.; Qian, Y.; Li, W.; Wei, P.; Li, G.; Zhang, F.; Yang, Z.; et al. Overexpression of MiRNA 4451 Is Associated With a Poor Survival of Patients With Hypopharyngeal Cancer After Surgery with Postoperative Radiotherapy. Transl. Oncol. 2018, 11, 1244–1250. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. MicroRNAs as Oncogenes and Tumor Suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef]
- McClelland, A.D.; Herman-Edelstein, M.; Komers, R.; Jha, J.C.; Winbanks, C.E.; Hagiwara, S.; Gregorevic, P.; Kantharidis, P.; Cooper, M.E. MiR-21 Promotes Renal Fibrosis in Diabetic Nephropathy by Targeting PTEN and SMAD7. Clin. Sci. 2015, 129, 1237–1249. [Google Scholar] [CrossRef]
- Zhang, Z.; Peng, H.; Chen, J.; Chen, X.; Han, F.; Xu, X.; He, X.; Yan, N. MicroRNA-21 Protects from Mesangial Cell Proliferation Induced by Diabetic Nephropathy in Db/Db Mice. FEBS Lett. 2009, 583, 2009–2014. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Gao, Y.-B.; Zhang, N.; Zou, D.-W.; Wang, P.; Zhu, Z.-Y.; Li, J.-Y.; Zhou, S.-N.; Wang, S.-C.; Wang, Y.-Y.; et al. MiR-21 Overexpression Enhances TGF-Β1-Induced Epithelial-to-Mesenchymal Transition by Target Smad7 and Aggravates Renal Damage in Diabetic Nephropathy. Mol. Cell. Endocrinol. 2014, 392, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Kriegel, A.J.; Liu, Y.; Usa, K.; Mladinov, D.; Liu, H.; Fang, Y.; Ding, X.; Liang, M. Delayed Ischemic Preconditioning Contributes to Renal Protection by Upregulation of MiR-21. Kidney Int. 2012, 82, 1167–1175. [Google Scholar] [CrossRef] [PubMed]
- MIR21 MicroRNA 21 [Homo Sapiens (Human)]—Gene—NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/406991 (accessed on 2 March 2023).
- Silahtaroglu, A.; Stenvang, J. MicroRNAs, Epigenetics and Disease. Essays Biochem. 2010, 48, 165–185. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.J.; Varani, G. Design of RNA-Targeting Macrocyclic Peptides. Methods Enzymol. 2019, 623, 339–372. [Google Scholar] [CrossRef]
- Buscaglia, L.E.B.; Li, Y. Apoptosis and the Target Genes of MicroRNA-21. Chin. J. Cancer 2011, 30, 371. [Google Scholar] [CrossRef]
- Su, Z.; Yang, Z.; Xu, Y.; Chen, Y.; Yu, Q. MicroRNAs in Apoptosis, Autophagy and Necroptosis. Oncotarget 2015, 6, 8474–8490. [Google Scholar] [CrossRef]
- Hashemi, M.; Mirdamadi, M.S.A.; Talebi, Y.; Khaniabad, N.; Banaei, G.; Daneii, P.; Gholami, S.; Ghorbani, A.; Tavakolpournegari, A.; Farsani, Z.M.; et al. Pre-Clinical and Clinical Importance of MiR-21 in Human Cancers: Tumorigenesis, Therapy Response, Delivery Approaches and Targeting Agents. Pharmacol. Res. 2023, 187, 106568. [Google Scholar] [CrossRef]
- An, F.; Liu, Y.; Hu, Y. MiR-21 Inhibition of LATS1 Promotes Proliferation and Metastasis of Renal Cancer Cells and Tumor Stem Cell Phenotype. Oncol. Lett. 2017, 14, 4684–4688. [Google Scholar] [CrossRef]
- Zhang, Y.; Jin, D.; Kang, X.; Zhou, R.; Sun, Y.; Lian, F.; Tong, X. Signaling Pathways Involved in Diabetic Renal Fibrosis. Front. Cell Dev. Biol. 2021, 9, 696542. [Google Scholar] [CrossRef]
- Simonson, M.S. Phenotypic Transitions and Fibrosis in Diabetic Nephropathy. Kidney Int. 2007, 71, 846–854. [Google Scholar] [CrossRef]
- Mason, R.M.; Wahab, N.A. Extracellular Matrix Metabolism in Diabetic Nephropathy. J. Am. Soc. Nephrol. 2003, 14, 1358–1373. [Google Scholar] [CrossRef]
- Singh, V.P.; Bali, A.; Singh, N.; Jaggi, A.S. Advanced Glycation End Products and Diabetic Complications. Korean J. Physiol. Pharmacol. 2014, 18, 1–14. [Google Scholar] [CrossRef]
- Heilig, C.W.; Concepcion, L.A.; Riser, B.L.; Freytag, S.; Zhu, M.; Cortes, P.; Hassett, C.C.; Gilbert, J.D.; Sastry, K.S.S.; Heilig, K.; et al. Overexpression of Glucose Transporters in Rat Mesangial Cells Cultured in a Normal Glucose Milieu Mimics the Diabetic Phenotype. J. Clin. Investig. 1995, 96, 1802. [Google Scholar] [CrossRef]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive Oxygen Species in Metabolic and Inflammatory Signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Panizo, S.; Martínez-Arias, L.; Alonso-Montes, C.; Cannata, P.; Martín-Carro, B.; Fernández-Martín, J.L.; Naves-Díaz, M.; Carrillo-López, N.; Cannata-Andía, J.B. Fibrosis in Chronic Kidney Disease: Pathogenesis and Consequences. Int. J. Mol. Sci. 2021, 22, 408. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, L.; Peng, W.; Liu, H.; Liang, L.; Zhang, X.; Mao, Y.; Zhou, X.; Shi, M.; Xiao, Y.; et al. Ski-related Novel Protein Suppresses the Development of Diabetic Nephropathy by Modulating Transforming Growth Factor-β Signaling and MicroRNA-21 Expression. J. Cell. Physiol. 2019, 234, 17925–17936. [Google Scholar] [CrossRef]
- Isaka, Y. Targeting TGF-β Signaling in Kidney Fibrosis. Int. J. Mol. Sci. 2018, 19, 2532. [Google Scholar] [CrossRef]
- Xu, Q.; Norman, J.T.; Shrivastav, S.; Lucio-Cazana, J.; Kopp, J.B. In Vitro Models of TGF-Beta-Induced Fibrosis Suitable for High-Throughput Screening of Antifibrotic Agents. Am. J. Physiol. Ren. Physiol. 2007, 293, F631–F640. [Google Scholar] [CrossRef]
- Chau, B.N.; Xin, C.; Hartner, J.; Ren, S.; Castano, A.P.; Linn, G.; Li, J.; Tran, P.T.; Kaimal, V.; Huang, X.; et al. MicroRNA-21 Promotes Fibrosis of the Kidney by Silencing Metabolic Pathways. Sci. Transl. Med. 2012, 4, 121ra18. [Google Scholar] [CrossRef]
- Roggli, E.; Britan, A.; Gattesco, S.; Lin-Marq, N.; Abderrahmani, A.; Meda, P.; Regazzi, R. Involvement of MicroRNAs in the Cytotoxic Effects Exerted by Proinflammatory Cytokines on Pancreatic β-Cells. Diabetes 2010, 59, 978–986. [Google Scholar] [CrossRef]
- Liu, R.; Liu, C.; He, X.; Sun, P.; Zhang, B.; Yang, H.; Shi, W.; Ruan, Q. MicroRNA-21 Promotes Pancreatic β Cell Function through Modulating Glucose Uptake. Nat. Commun. 2022, 13, 3545. [Google Scholar] [CrossRef]
- Zhang, J.; Xing, Q.; Zhou, X.; Li, J.; Li, Y.; Zhang, L.; Zhou, Q.; Tang, B. Circulating MiRNA-21 Is a Promising Biomarker for Heart Failure. Mol. Med. Rep. 2017, 16, 7766–7774. [Google Scholar] [CrossRef]
- Fiorentino, L.; Cavalera, M.; Mavilio, M.; Conserva, F.; Menghini, R.; Gesualdo, L.; Federici, M. Regulation of TIMP3 in Diabetic Nephropathy: A Role for MicroRNAs. Acta Diabetol. 2013, 50, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, Y.; Ma, M.; Li, M.; Zou, D.; Yang, J.; Zhu, Z.; Zhao, X. Effect of MiR-21 on Renal Fibrosis by Regulating MMP-9 and TIMP1 in Kk-Ay Diabetic Nephropathy Mice. Cell Biochem. Biophys. 2013, 67, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Gan, H.; Zhang, H.; Tang, W.; Sun, Y.; Tang, X.; Kong, D.; Zhou, J.; Wang, Y.; Zhu, Y. MicroRNA-21 Inhibits SMAD7 Expression through a Target Sequence in the 3′ Untranslated Region and Inhibits Proliferation of Renal Tubular Epithelial Cells. Mol. Med. Rep. 2014, 10, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Dey, N.; Das, F.; Mariappan, M.M.; Mandal, C.C.; Ghosh-Choudhury, N.; Kasinath, B.S.; Choudhury, G.G. MicroRNA-21 Orchestrates High Glucose-Induced Signals to TOR Complex 1, Resulting in Renal Cell Pathology in Diabetes. J. Biol. Chem. 2011, 286, 25586–25603. [Google Scholar] [CrossRef]
- Baker, M.A.; Davis, S.J.; Liu, P.; Pan, X.; Williams, A.M.; Iczkowski, K.A.; Gallagher, S.T.; Bishop, K.; Regner, K.R.; Liu, Y.; et al. Tissue-Specific MicroRNA Expression Patterns in Four Types of Kidney Disease. J. Am. Soc. Nephrol. 2017, 28, 2985–2992. [Google Scholar] [CrossRef]
- Huang, C.-K.; Bär, C.; Thum, T. MiR-21, Mediator, and Potential Therapeutic Target in the Cardiorenal Syndrome. Front. Pharmacol. 2020, 11, 726. [Google Scholar] [CrossRef]
- Gura, T. Antisense Has Growing Pains. Science 1995, 270, 575–577. [Google Scholar] [CrossRef]
- Kurreck, J.; Wyszko, E.; Gillen, C.; Erdmann, V.A. Design of Antisense Oligonucleotides Stabilized by Locked Nucleic Acids. Nucleic Acids Res. 2002, 30, 1911–1918. [Google Scholar] [CrossRef]
- Judge, A.; Bola, G.; Lee, A.; Maclachlan, I. Design of Noninflammatory Synthetic SiRNA Mediating Potent Gene Silencing in Vivo. Mol. Ther. 2006, 13, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Marcucci, G.; Croce, C.M. Targeting MicroRNAs in Cancer: Rationale, Strategies and Challenges. Nat. Rev. Drug Discov. 2010, 9, 775–789. [Google Scholar] [CrossRef] [PubMed]
- Rubel, D.; Boulanger, J.; Craciun, F.; Xu, E.Y.; Zhang, Y.; Phillips, L.; Callahan, M.; Weber, W.; Song, W.; Ngai, N.; et al. Anti-MicroRNA-21 Therapy on Top of ACE Inhibition Delays Renal Failure in Alport Syndrome Mouse Models. Cells 2022, 11, 594. [Google Scholar] [CrossRef]
- Genzyme, a Sanofi Company. A Phase 2, Randomized, Double-Blind, Placebo-Controlled Study to Evaluate the Safety, Efficacy, Pharmacodynamics, and Pharmacokinetics of Lademirsen (SAR339375) for Subcutaneous Injection Administered Every Week in Patients With Alport Syndrome. 2022. Available online: https://clinicaltrials.gov/ (accessed on 2 March 2023).
- Alnuqaydan, A.M. Targeting Micro-RNAs by Natural Products: A Novel Future Therapeutic Strategy to Combat Cancer. Am. J. Transl. Res. 2020, 12, 3531–3556. [Google Scholar] [PubMed]
- Guo, H.; Wang, Y.; Zhang, X.; Zang, Y.; Zhang, Y.; Wang, L.; Wang, H.; Wang, Y.; Cao, A.; Peng, W. Astragaloside IV Protects against Podocyte Injury via SERCA2-Dependent ER Stress Reduction and AMPKα-Regulated Autophagy Induction in Streptozotocin-Induced Diabetic Nephropathy. Sci. Rep. 2017, 7, 6852. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, Y.; Tian, N.; Zou, D.; Shi, Y.; Zhang, N. Astragaloside IV Improves Renal Function and Fibrosis via Inhibition of MiR-21-Induced Podocyte Dedifferentiation and Mesangial Cell Activation in Diabetic Mice. Drug Des. Dev. Ther. 2018, 12, 2431–2442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; He, S.; Yang, F.; Yu, H.; Xie, W.; Dai, Q.; Zhang, D.; Liu, X.; Zhou, S.; Zhang, K. Hyperoside Ameliorates Glomerulosclerosis in Diabetic Nephropathy by Downregulating MiR-21. Can. J. Physiol. Pharmacol. 2016, 94, 1249–1256. [Google Scholar] [CrossRef]
- Dong, X.; Liu, S.; Zhang, L.; Yu, S.; Huo, L.; Qile, M.; Liu, L.; Yang, B.; Yu, J. Downregulation of MiR-21 Is Involved in Direct Actions of Ursolic Acid on the Heart: Implications for Cardiac Fibrosis and Hypertrophy. Cardiovasc. Ther. 2015, 33, 161–167. [Google Scholar] [CrossRef]
- Wang, J.; Li, Y.; Wang, X.; Jiang, C. Ursolic Acid Inhibits Proliferation and Induces Apoptosis in Human Glioblastoma Cell Lines U251 by Suppressing TGF-Β1/MiR-21/PDCD4 Pathway. Basic. Clin. Pharmacol. Toxicol. 2012, 111, 106–112. [Google Scholar] [CrossRef]
- Xu, L.; Fan, Q.; Wang, X.; Li, L.; Lu, X.; Yue, Y.; Cao, X.; Liu, J.; Zhao, X.; Wang, L. Ursolic Acid Improves Podocyte Injury Caused by High Glucose. Nephrol. Dial. Transplant. 2017, 32, 1285–1293. [Google Scholar] [CrossRef]
- Cao, Y.; Hu, J.; Sui, J.; Jiang, L.; Cong, Y.; Ren, G. Quercetin Is Able to Alleviate TGF-β-Induced Fibrosis in Renal Tubular Epithelial Cells by Suppressing MiR-21. Exp. Ther. Med. 2018, 16, 2442–2448. [Google Scholar] [CrossRef] [PubMed]
- Kassiri, Z.; Oudit, G.Y.; Kandalam, V.; Awad, A.; Wang, X.; Ziou, X.; Maeda, N.; Herzenberg, A.M.; Scholey, J.W. Loss of TIMP3 Enhances Interstitial Nephritis and Fibrosis. J. Am. Soc. Nephrol. 2009, 20, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Lan, R.; Geng, H.; Polichnowski, A.J.; Singha, P.K.; Saikumar, P.; McEwen, D.G.; Griffin, K.A.; Koesters, R.; Weinberg, J.M.; Bidani, A.K.; et al. PTEN Loss Defines a TGF-β-Induced Tubule Phenotype of Failed Differentiation and JNK Signaling during Renal Fibrosis. Am. J. Physiol.-Ren. Physiol. 2012, 302, F1210–F1223. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Zhang, X.; Wang, Y.; Cai, L.; Ren, L.; Tang, L.; Wang, J.; Zhao, Y.; Wang, Y.; Liu, Q.; et al. Targeting JNK by a New Curcumin Analog to Inhibit NF-KB-Mediated Expression of Cell Adhesion Molecules Attenuates Renal Macrophage Infiltration and Injury in Diabetic Mice. PLoS ONE 2013, 8, e79084. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, Y.; Zhao, Y.; Peng, K.; Li, W.; Wang, Y.; Zhang, J.; Zhou, S.; Liu, Q.; Li, X.; et al. Inhibition of JNK Phosphorylation by a Novel Curcumin Analog Prevents High Glucose-Induced Inflammation and Apoptosis in Cardiomyocytes and the Development of Diabetic Cardiomyopathy. Diabetes 2014, 63, 3497–3511. [Google Scholar] [CrossRef]
- Wu, B.; Bell, K.; Stanford, A.; Kern, D.M.; Tunceli, O.; Vupputuri, S.; Kalsekar, I.; Willey, V. Understanding CKD among Patients with T2DM: Prevalence, Temporal Trends, and Treatment Patterns—NHANES 2007–2012. BMJ Open Diabetes Res. Care 2016, 4, e000154. [Google Scholar] [CrossRef]
- Nozari, E.; Moradi, A.; Samadi, M. Effect of Atorvastatin, Curcumin, and Quercetin on MiR-21 and MiR-122 and Their Correlation with TGFβ1 Expression in Experimental Liver Fibrosis. Life Sci. 2020, 259, 118293. [Google Scholar] [CrossRef]
- Xiang, J.; Zhang, H.; Zhou, X.; Wang, D.; Chen, R.; Tan, W.; Liang, L.; Shi, M.; Zhang, F.; Xiao, Y.; et al. Atorvastatin Restores PPARα Inhibition of Lipid Metabolism Disorders by Downregulating MiR-21 Expression to Improve Mitochondrial Function and Alleviate Diabetic Nephropathy Progression. Front. Pharmacol. 2022, 13, 819787. [Google Scholar] [CrossRef]
- Khokhar, M.; Roy, D.; Bajpai, N.K.; Bohra, G.K.; Yadav, D.; Sharma, P.; Purohit, P. Metformin Mediates MicroRNA-21 Regulated Circulating Matrix Metalloproteinase-9 in Diabetic Nephropathy: An in-Silico and Clinical Study. Arch. Physiol. Biochem. 2021, 1–11. [Google Scholar] [CrossRef]
- Sun, L.; Yuan, Q.; Xu, T.; Yao, L.; Feng, J.; Ma, J.; Wang, L.; Lu, C.; Wang, D. Pioglitazone Improves Mitochondrial Function in the Remnant Kidney and Protects against Renal Fibrosis in 5/6 Nephrectomized Rats. Front. Pharmacol. 2017, 8, 545. [Google Scholar] [CrossRef]
- Toblli, J.E.; Cao, G.; Giani, J.F.; Angerosa, M.; Dominici, F.P.; Gonzalez-Cadavid, N.F. Antifibrotic Effects of Pioglitazone at Low Doses on the Diabetic Rat Kidney Are Associated with the Improvement of Markers of Cell Turnover, Tubular and Endothelial Integrity, and Angiogenesis. Kidney Blood Press Res. 2011, 34, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Xu, T.; Chen, Y.; Qu, W.; Sun, D.; Song, X.; Yuan, Q.; Yao, L. Pioglitazone Attenuates Kidney Fibrosis via MiR-21-5p Modulation. Life Sci. 2019, 232, 116609. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Model | miR-21 Expression | Target | Major Findings | Ref. |
|---|---|---|---|---|
| Human and mouse | Upregulated | CDC25A, CDK6 | miR-21 targets CDC25A and CDK6 in mesangial cells and results in mesangial cell hypertrophy by stimulating a G1-phase arrest. miR-21 targets PTEN, increasing podocyte motility as well as the formation and deposition of extracellular matrix. | [23] |
| MES13 cell line, mouse kidney, and human biopsies | Upregulated | TIMP3 | A considerable overexpression of miR-21 was seen in mesangial cells cultured in high glucose environments and in mouse kidney and human kidney biopsies. The glycemic burden can stimulate miR-21 expression and destroy TIMP3 mRNA. | [54] |
| Diabetic kk-ay mice and C57BL mice (control) | Upregulated | MMP-9/TIMP1 | miR-21 expression was significantly higher in kk-ay mice. miR-21 expression positively correlated with TIMP1, collagen IV, urine albumin creatine ratio (ACR), and fibronectin; whereas negatively correlated with creatine clearance ratio (Ccr) and MMP-9 protein. Antagomir-21 improved Ccr and ACR and reduced collagen IV, TIMP1, and fibronectin. | [55] |
| Male kk-ay and C57BL/6J mice | Upregulated | SMAD7 | miR-21 overexpression accelerated TGF-β1-induced EMT by targeting SMAD7. Notably, miR-21 inhibitor improves the renal structure and function and inhibits fibrosis. | [31] |
| DN mouse models and cell models | Upregulated | FOXO1 | FOXO1 was recognized as a target of miR-21. By specifically targeting FOXO1 in high glucose cultured podocytes, miR-21 utilizes its pro-apoptosis and anti-autophagy effects. | [47] |
| Rat renal tubular epithelial cells and HEK 293T cells | Upregulated | SMAD7 | SMAD7 is a direct target of miR-21, and its overexpression may prevent rat renal tubular epithelial cells from proliferating. | [56] |
| db/db mice (a mouse model of T2D) | Upregulated | SMAD7 | Overexpression of miR-21 in kidney cells increased the generation of fibrotic and inflammatory markers driven by high glucose, whereas miR-21 knockdown decreased this production. Renal miR-21 knockdown restored Smad7 levels and reduced activation of the TGF-β and NF-κB signaling pathways. | [21] |
| OVE26 type 1 diabetic mouse | Upregulated | PTEN, PRAS40 | Upregulation of miR-21 resulted in the promotion of renal fibrosis. In high glucose-induced TORC1 activity, miR-21 increased renal cell hypertrophy and fibronectin expression. | [57] |
| Kidney biopsies of DN patients and normal kidney donors | Upregulated | PTEN-SMAD7 | Tubular miR-21 upregulation was seen in human kidney biopsies. miR-21 specifically targets the repressors of SMAD3-dependent and PI3K-dependent TGF-β1 signaling, SMAD7, and PTEN (known fibrotic signaling proteins), respectively. | [29] |
| DN patients | Upregulated | Not determined | Patients with DN had higher levels of hsa-miR-21-5p, and an inverse relationship between eGFR and miR-21-5p in the proximal tubules and glomeruli was found. | [58] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhas, Y.; Arshad, N.; Biswas, N.; Jones, L.D.; Ashili, S. MicroRNA-21 Silencing in Diabetic Nephropathy: Insights on Therapeutic Strategies. Biomedicines 2023, 11, 2583. https://doi.org/10.3390/biomedicines11092583
Dhas Y, Arshad N, Biswas N, Jones LD, Ashili S. MicroRNA-21 Silencing in Diabetic Nephropathy: Insights on Therapeutic Strategies. Biomedicines. 2023; 11(9):2583. https://doi.org/10.3390/biomedicines11092583
Chicago/Turabian StyleDhas, Yogita, Numair Arshad, Nupur Biswas, Lawrence D. Jones, and Shashaanka Ashili. 2023. "MicroRNA-21 Silencing in Diabetic Nephropathy: Insights on Therapeutic Strategies" Biomedicines 11, no. 9: 2583. https://doi.org/10.3390/biomedicines11092583
APA StyleDhas, Y., Arshad, N., Biswas, N., Jones, L. D., & Ashili, S. (2023). MicroRNA-21 Silencing in Diabetic Nephropathy: Insights on Therapeutic Strategies. Biomedicines, 11(9), 2583. https://doi.org/10.3390/biomedicines11092583