
T Cell-Induced Colitis Is Exacerbated by Prolonged Stress: A Comparison in Male and Female Mice
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Induction of Chronic Colitis
2.3. Experimental Design
2.4. Histopathology
2.5. Semiquantitative Real-Time PCR
2.6. Serum and Colonic Measurement of Cytokines
2.7. Statistical Analysis
3. Results
3.1. RST Does Not Lead to Colitis in the Absence of T Cell Transfer in Male or Female Rag1tm1Mom Mice
3.2. Male Mice Lose Significantly More Weight Than Female Mice Regardless of RST Exposure
3.3. Sex and RST Exposure Lead to Significantly Altered Colonic Cytokine Expression and Histopathology
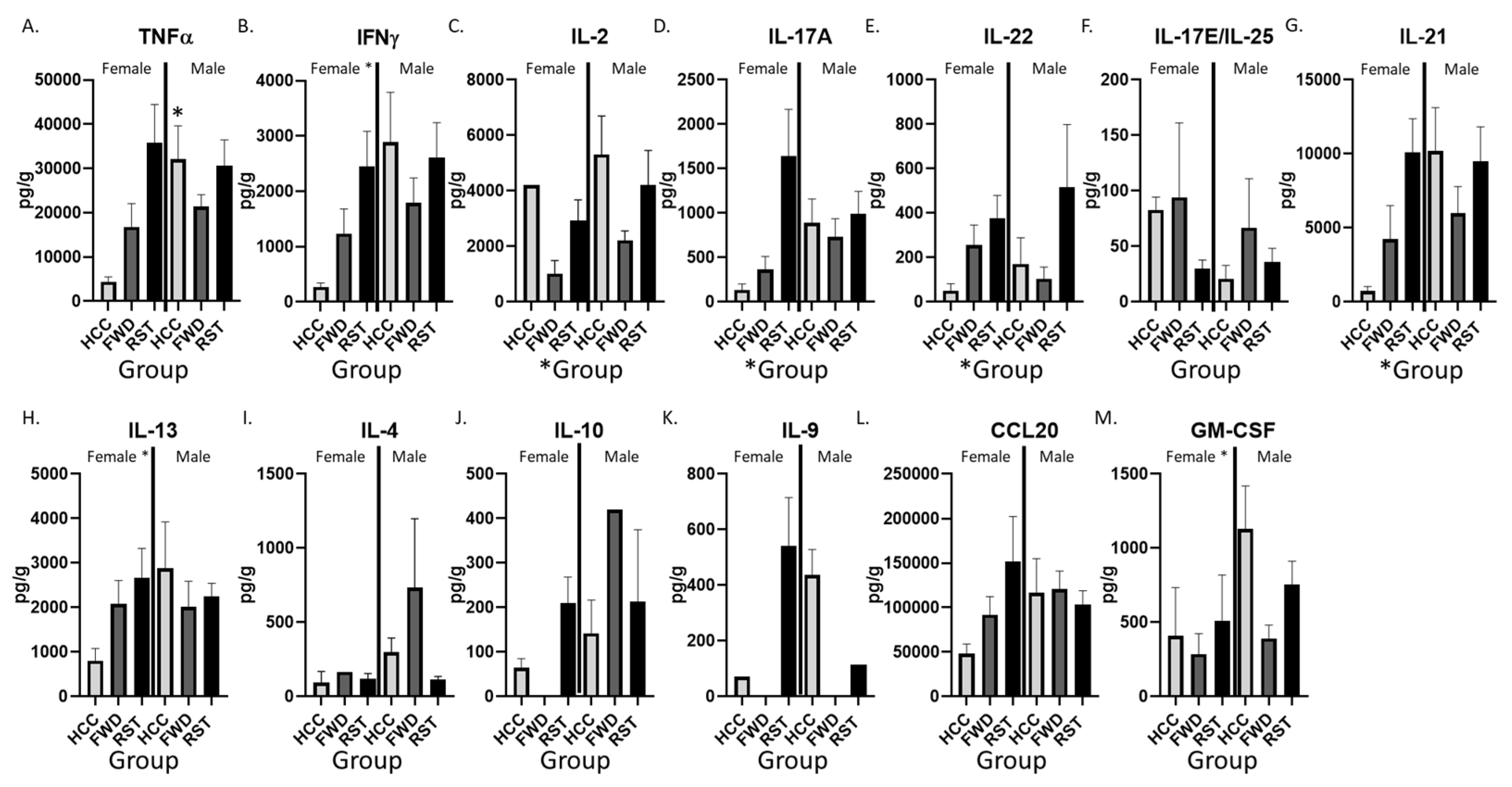
3.4. RST Exposure and Sex Significantly Alter Cytokine Protein Levels in the Colon
3.5. RST and Sex Significantly Alter Cytokine Levels in Serum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dahlhamer, J.M.; Zammitti, E.P.; Ward, B.W.; Wheaton, A.G.; Croft, J.B. Prevalence of Inflammatory Bowel Disease Among Adults Aged ≥18 Years—United States, 2015. MMWR Morb. Mortal. Weekly Rep. 2016, 65, 1166–1169. [Google Scholar]
- Sartor, R.B. Genetics and environmental interactions shape the intestinal microbiome to promote inflammatory bowel disease versus mucosal homeostasis. Gastroenterology 2010, 139, 1816–1819. [Google Scholar] [PubMed]
- Ostanin, D.V.; Bao, J.; Koboziev, I.; Gray, L.; Robinson-Jackson, S.A.; Kosloski-Davidson, M.; Price, V.H.; Grisham, M.B.; Steury, M.D.; Kang, H.J.; et al. T cell transfer model of chronic colitis: Concepts, considerations, and tricks of the trade. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, G135-46. [Google Scholar] [PubMed]
- Ostanin, D.V.; Furr, K.L.; Pavlick, K.P.; Gray, L.; Kevil, C.G.; Shukla, D.; D’Souza, D.; Hoffman, J.M.; Grisham, M.B.; Chidlow, J.H.; et al. T cell-associated CD18 but not CD62L, ICAM-1, or PSGL-1 is required for the induction of chronic colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1706-14. [Google Scholar] [PubMed]
- Pavlick, K.P.; Ostanin, D.V.; Furr, K.L.; Laroux, F.S.; Brown, C.M.; Gray, L.; Kevil, C.G.; Grisham, M.B. Role of T-cell-associated lymphocyte function-associated antigen-1 in the pathogenesis of experimental colitis. Int. Immunol. 2006, 18, 389–398. [Google Scholar] [PubMed]
- Eri, R.; McGuckin, M.A.; Wadley, R. T cell transfer model of colitis: A great tool to assess the contribution of T cells in chronic intestinal inflammation. Methods Mol. Biol. 2012, 844, 261–275. [Google Scholar]
- Greuter, T.; Manser, C.; Pittet, V.; Vavricka, S.R.; Biedermann, L.; on behalf of Swiss IBDnet; an official working group of the Swiss Society of Gastroenterology. Gender Differences in Inflammatory Bowel Disease. Digestion 2020, 101 (Suppl. S1), 98–104. [Google Scholar]
- Assad, S.; Khan, H.H.; Ghazanfar, H.; Khan, Z.H.; Mansoor, S.; Rahman, M.A.; Khan, G.H.; Zafar, B.; Tariq, U.; Malik, S.A. Role of Sex Hormone Levels and Psychological Stress in the Pathogenesis of Autoimmune Diseases. Cureus 2017, 9, e1315. [Google Scholar]
- Park, H.J.; Choi, J.M. Sex-specific regulation of immune responses by PPARs. Exp. Mol. Med. 2017, 49, e364. [Google Scholar]
- Pierdominici, M.; Maselli, A.; Varano, B.; Barbati, C.; Cesaro, P.; Spada, C.; Zullo, A.; Lorenzetti, R.; Rosati, M.; Rainaldi, G.; et al. Linking estrogen receptor β expression with inflammatory bowel disease activity. Oncotarget 2015, 6, 40443–40451. [Google Scholar]
- Xu, L.; Huang, G.; Cong, Y.; Yu, Y.; Li, Y. Sex-related Differences in Inflammatory Bowel Diseases: The Potential Role of Sex Hormones. Inflamm. Bowel Dis. 2022, 28, 1766–1775. [Google Scholar] [PubMed]
- van der Giessen, J.; van der Woude, C.J.; Peppelenbosch, M.P.; Fuhler, G.M. A Direct Effect of Sex Hormones on Epithelial Barrier Function in Inflammatory Bowel Disease Models. Cells 2019, 8, 261. [Google Scholar] [PubMed]
- Byrne, G.; Rosenfeld, G.; Leung, Y.; Qian, H.; Raudzus, J.; Nunez, C.; Bressler, B. Prevalence of Anxiety and Depression in Patients with Inflammatory Bowel Disease. Can. J. Gastroenterol. Hepatol. 2017, 2017, 6496727. [Google Scholar] [PubMed]
- Ma, Y.; Liu, T.; Li, X.; Kong, A.; Xiao, R.; Xie, R.; Gao, J.; Wang, Z.; Cai, Y.; Zou, J.; et al. Estrogen receptor β deficiency impairs gut microbiota: A possible mechanism of IBD-induced anxiety-like behavior. Microbiome 2022, 10, 160. [Google Scholar]
- Rustgi, S.D.; Kayal, M.; Shah, S.C. Sex-based differences in inflammatory bowel diseases: A review. Therap Adv. Gastroenterol. 2020, 13, 1756284820915043. [Google Scholar]
- Haro, C.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Landa, B.B.; Navas-Cortés, J.A.; Tena-Sempere, M.; et al. Intestinal Microbiota Is Influenced by Gender and Body Mass Index. PLoS ONE 2016, 11, e0154090. [Google Scholar]
- Mayneris-Perxachs, J.; Arnoriaga-Rodríguez, M.; Luque-Córdoba, D.; Priego-Capote, F.; Pérez-Brocal, V.; Moya, A.; Burokas, A.; Maldonado, R.; Fernández-Real, J.-M. Gut microbiota steroid sexual dimorphism and its impact on gonadal steroids: Influences of obesity and menopausal status. Microbiome 2020, 8, 136. [Google Scholar]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar]
- Kim, Y.S.; Unno, T.; Kim, B.-Y.; Park, M.-S. Sex Differences in Gut Microbiota. World J. Mens Health 2020, 38, 48–60. [Google Scholar]
- Fuyuno, Y.; Yamazaki, K.; Takahashi, A.; Esaki, M.; Kawaguchi, T.; Takazoe, M.; Matsumoto, T.; Matsui, T.; Tanaka, H.; Motoya, S.; et al. Genetic characteristics of inflammatory bowel disease in a Japanese population. J. Gastroenterol. 2016, 51, 672–681. [Google Scholar]
- Ananthakrishnan, A.N. Epidemiology and risk factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [PubMed]
- Camara, R.J.; Ziegler, R.; Begré, S.; Schoepfer, A.M.; von Känel, R. The role of psychological stress in inflammatory bowel disease: Quality assessment of methods of 18 prospective studies and suggestions for future research. Digestion 2009, 80, 129–139. [Google Scholar]
- Collins, S.M.; McHugh, K.; Jacobson, K.; Khan, I.; Riddell, R.; Murase, K.; Weingarten, H. Previous inflammation alters the response of the rat colon to stress. Gastroenterology 1996, 111, 1509–1515. [Google Scholar] [PubMed]
- Qiu, B.S.; Vallance, B.; Blennerhassett, P.; Collins, S. The role of CD4+ lymphocytes in the susceptibility of mice to stress-induced reactivation of experimental colitis. Nat. Med. 1999, 5, 1178–1182. [Google Scholar] [PubMed]
- Maltz, R.M.; Keirsey, J.; Kim, S.C.; Mackos, A.R.; Gharaibeh, R.Z.; Moore, C.C.; Xu, J.; Bakthavatchalu, V.; Somogyi, A.; Bailey, M.T. Prolonged restraint stressor exposure in outbred CD-1 mice impacts microbiota, colonic inflammation, and short chain fatty acids. PLoS ONE 2018, 13, e0196961. [Google Scholar]
- Bailey, M.T.; Dowd, S.E.; Parry, N.M.A.; Galley, J.D.; Schauer, D.B.; Lyte, M. Stressor exposure disrupts commensal microbial populations in the intestines and leads to increased colonization by Citrobacter rodentium. Infect. Immun. 2010, 78, 1509–1519. [Google Scholar]
- Mackos, A.R.; Maltz, R.; Bailey, M.T. The role of the commensal microbiota in adaptive and maladaptive stressor-induced immunomodulation. Horm. Behav. 2017, 88, 70–78. [Google Scholar]
- Reinoso Webb, C.; Bakker, H.D.; Koboziev, I.; Jones-Hall, Y.; Kottapalli, K.R.; Ostanin, D.; Furr, K.L.; Mu, Q.; Luo, X.M.; Grisham, M.B. Differential Susceptibility to T Cell-Induced Colitis in Mice: Role of the Intestinal Microbiota. Inflamm. Bowel Dis. 2018, 24, 361–379. [Google Scholar]
- Rath, H.C.; Herfarth, H.H.; Ikeda, J.S.; Grenther, W.B.; Jr, T.E.H.; Balish, E.; Taurog, J.D.; Hammer, R.E.; Wilson, K.H.; Sartor, R.B. Normal luminal bacteria, especially Bacteroides species, mediate chronic colitis, gastritis, and arthritis in HLA-B27/human beta2 microglobulin transgenic rats. J. Clin. Investig. 1996, 98, 945–953. [Google Scholar]
- Head, C.C.; Farrow, M.J.; Sheridan, J.F.; Padgett, D.A. Androstenediol reduces the anti-inflammatory effects of restraint stress during wound healing. Brain Behav. Immun. 2006, 20, 590–596. [Google Scholar]
- Tizard, I. Sickness behavior, its mechanisms and significance. Anim. Health Res. Rev. 2008, 9, 87–99. [Google Scholar] [PubMed]
- Knopp, R.C.; Baumann, K.K.; Wilson, M.L.; Banks, W.A.; Erickson, M.A. Amyloid Beta Pathology Exacerbates Weight Loss and Brain Cytokine Responses following Low-Dose Lipopolysaccharide in Aged Female Tg2576 Mice. Int. J. Mol. Sci. 2022, 23, 2377. [Google Scholar] [PubMed]
- Vichaya, E.G.; Malik, S.; Sominsky, L.; Ford, B.G.; Spencer, S.J.; Dantzer, R. Microglia depletion fails to abrogate inflammation-induced sickness in mice and rats. J. Neuroinflammation 2020, 17, 172. [Google Scholar]
- Veenema, A.H.; Reber, S.O.; Selch, S.; Obermeier, F.; Neumann, I.D. Early life stress enhances the vulnerability to chronic psychosocial stress and experimental colitis in adult mice. Endocrinology 2008, 149, 2727–2736. [Google Scholar]
- Babickova, J.; Tóthová, Ľ.; Lengyelová, E.; Bartoňová, A.; Hodosy, J.; Gardlík, R.; Celec, P. Sex Differences in Experimentally Induced Colitis in Mice: A Role for Estrogens. Inflammation 2015, 38, 1996–2006. [Google Scholar] [PubMed]
- Maite, C.B.; Roy, M.; Emilie, V. The Effect of Sex-Specific Differences on IL-10−/− Mouse Colitis Phenotype and Microbiota. Int. J. Mol. Sci. 2023, 24, 10364. [Google Scholar]
- Goodman, W.A.; Garg, R.R.; Reuter, B.K.; Mattioli, B.; Rissman, E.F.; Pizarro, T.T. Loss of estrogen-mediated immunoprotection underlies female gender bias in experimental Crohn’s-like ileitis. Mucosal Immunol. 2014, 7, 1255–1265. [Google Scholar]
- Fuller-Thomson, E.; Sulman, J. Depression and inflammatory bowel disease: Findings from two nationally representative Canadian surveys. Inflamm. Bowel Dis. 2006, 12, 697–707. [Google Scholar]
- Severs, M.; Spekhorst, L.M.; Mangen, M.-J.J.; Dijkstra, G.; Löwenberg, M.; Hoentjen, F.; Jong, A.E.v.d.M.-D.; Pierik, M.; Ponsioen, C.Y.; Bouma, G.; et al. Sex-Related Differences in Patients With Inflammatory Bowel Disease: Results of 2 Prospective Cohort Studies. Inflamm. Bowel Dis. 2018, 24, 1298–1306. [Google Scholar]
- Zelinkova, Z.; Bultman, E.; Vogelaar, L.; Bouziane, C.; Kuipers, E.J.; van der Woude, C.J. Sex-dimorphic adverse drug reactions to immune suppressive agents in inflammatory bowel disease. World J. Gastroenterol. 2012, 18, 6967–6973. [Google Scholar]
- Graff, L.A.; Walker, J.R.; Bernstein, C.N. Depression and anxiety in inflammatory bowel disease: A review of comorbidity and management. Inflamm. Bowel Dis. 2009, 15, 1105–1118. [Google Scholar] [PubMed]
- Hauser, W.; Janke, K.-H.; Klump, B.; Hinz, A. Anxiety and depression in patients with inflammatory bowel disease: Comparisons with chronic liver disease patients and the general population. Inflamm. Bowel Dis. 2011, 17, 621–632. [Google Scholar] [PubMed]
- Araki, M.; Shinzaki, S.; Yamada, T.; Arimitsu, S.; Komori, M.; Shibukawa, N.; Mukai, A.; Nakajima, S.; Kinoshita, K.; Kitamura, S.; et al. Psychologic stress and disease activity in patients with inflammatory bowel disease: A multicenter cross-sectional study. PLoS ONE 2020, 15, e0233365. [Google Scholar]
- Fuller-Thomson, E.; Lateef, R.; Sulman, J. Robust Association Between Inflammatory Bowel Disease and Generalized Anxiety Disorder: Findings from a Nationally Representative Canadian Study. Inflamm. Bowel Dis. 2015, 21, 2341–2348. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cytokines | HCC Female | FWD Female | RST Female | HCC Male | FWD Male | RST Male |
|---|---|---|---|---|---|---|
| TNFα | 416 ± 90.6 (6) | 690.8 ± 189.4 (7) | 1122.2 ± 254.1 (7) | 758.3 ± 195.3 (7) | 1087.8 ± 138.6 (7) | 878.9 ± 100.2 (7) |
| IFNγ | 134.1 ± 54.3 (6) | 290.1 ±105.0 (7) | 237.9 ± 52.9 (7) | 257.3 ± 82.5 (7) | 316.5 ± 37.9 (7) | 305.5 ± 45.0 (7) |
| IL-2 | 126.9 ± 39.1 (7) | 97.9 ± 14.7 (7) | 112.2 ± 22.5 (7) | 112.1 ± 41.9 (7) | 136.9 ± 13.3 (7) | 157.5 ± 10.6 (7) |
| IL-17A | 56.0 ± 16.9 (7) | 66.9 ± 14.0 (7) | 97.9 ± 28.1 (7) | 106.1 ± 30.4 (7) | 106.2 ± 20.4 (7) | 103.1 ± 19.0 (7) |
| IL-22 | 258.8 ± 57.8 (7) | 283.1 ± 109.2 (7) | 698.0 ± 263.4 (7) | 308.4 ± 136.2 (7) | 394.6 ± 109.1 (7) | 368.1 ± 76.6 (7) |
| IL-21 | 13.1 (1) | 33.9 (1) | 22.4 ± 5.3 (5) | 34.0 ± 19.1 (3) | 8.8 ± 0.9 (2) | 14.8 ± 5.0 (3) |
| IL-17E/IL-25 | 2.4 ± 1.1 (7) | 4.9 ± 2.4 (7) | 5.9 ± 1.8 (7) | 3.4 ± 0.7 (7) | 2.8 ± 0.9 (7) | 5.2 ± 0.7 (7) |
| IL-17F | 200.9 ± 73.7 (4) | 59.8 ± 46.8 (2) | 188.1 ± 49.3 (5) | 106.5 ± 34.8 (2) | 92.2 ± 42.8 (2) | 120.8 ± 103.9 (4) |
| IL-13 | 68.8 ± 27.2 (4) | 48.4 ± 33.5 (2) | 110.1 ± 14.2 (6) | 78.8 ± 23.3 (3) | 67.7 ± 12.0 (6) | 72.4 ± 12.9 (5) |
| IL-4 | 13.5 ± 3.5 (7) | 9.9 ± 4.4 (7) | 10.3 ± 3.8 (7) | 6.9 ± 3.5 (7) | 11.0 ± 1.9 (7) | 11.0 ± 4.5 (6) |
| IL-10 | 140.6 ± 20.4 (7) | 150.7 ± 29.2 (7) | 165.6 ± 17.3 (7) | 129.2 ±30.9 (7) | 139.7 ± 11.9 (7) | 120.7 ± 8.3 (7) |
| IL-9 | NDL (0) | 19.7 ± 16.1 (2) | 31.0 ± 8.6 (4) | 39.3 ± 0.6 (2) | 21.2 ± 11.5 (4) | 14.5 ± 4.2 (4) |
| CCL20 | 2178.6 ± 493.3 (7) | 2716.6 ± 605.8 (7) | 4473.8 ± 1212.2 (6) | 2946 ± 802.5 (6) | 9600 ± 3183.6 (7) | 11,045 ± 3883.5 (6) |
| GM-CSF | 3.8 ± 1.2 (7) | 3.5 ± 0.8 (7) | 3.7 ± 0.8 (7) | 3.6 ± 1.0 (7) | 4.3 ±0.4 (7) | 4.9 ± 1.3 (7) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maltz, R.M.; Marte-Ortiz, P.; McClinchie, M.G.; Hilt, M.E.; Bailey, M.T. T Cell-Induced Colitis Is Exacerbated by Prolonged Stress: A Comparison in Male and Female Mice. Biomedicines 2024, 12, 214. https://doi.org/10.3390/biomedicines12010214
Maltz RM, Marte-Ortiz P, McClinchie MG, Hilt ME, Bailey MT. T Cell-Induced Colitis Is Exacerbated by Prolonged Stress: A Comparison in Male and Female Mice. Biomedicines. 2024; 12(1):214. https://doi.org/10.3390/biomedicines12010214
Chicago/Turabian StyleMaltz, Ross M., Pedro Marte-Ortiz, Madeline G. McClinchie, Miranda E. Hilt, and Michael T. Bailey. 2024. "T Cell-Induced Colitis Is Exacerbated by Prolonged Stress: A Comparison in Male and Female Mice" Biomedicines 12, no. 1: 214. https://doi.org/10.3390/biomedicines12010214
APA StyleMaltz, R. M., Marte-Ortiz, P., McClinchie, M. G., Hilt, M. E., & Bailey, M. T. (2024). T Cell-Induced Colitis Is Exacerbated by Prolonged Stress: A Comparison in Male and Female Mice. Biomedicines, 12(1), 214. https://doi.org/10.3390/biomedicines12010214