

Adiponectin C1q/Tumor Necrosis Factor-Related Protein 13 (CTRP13) Protects against Renal Inflammation and Fibrosis in Obstructive Nephropathy

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Establishment of the UUO-Induced CKD Model
2.2. Histology Staining
2.3. Biochemical Detection
2.4. Cell Culture and Treatment
2.5. Western Blot Analysis
2.6. Reverse Transcription Quantitative (RT-q) PCR Analysis
2.7. Statistical Analysis
3. Results
3.1. Plasma CTRP13 Levels Are Lower during the Progression of Renal Fibrosis
3.2. CTRP13 Treatment Leads to an Improvement in UUO-Mediated Renal Dysfunction
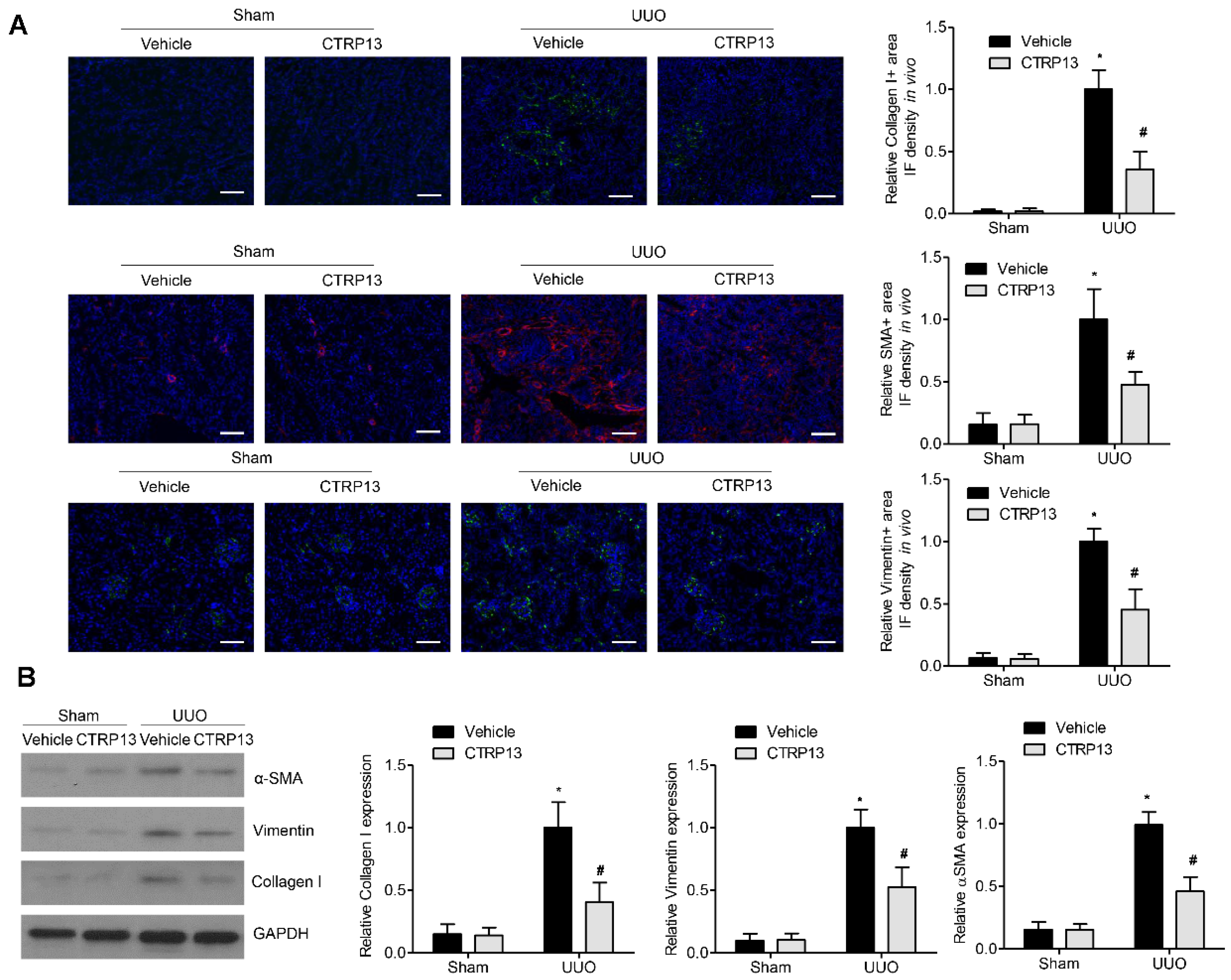
3.3. CTRP13 Ameliorates Kidney Injury and Fibrosis in UUO-Model Mice
3.4. CTRP13 Inhibits UUO-Induced ECM Accumulation
3.5. CTRP13 Regulates the TGF-β1/Smad Signaling Pathway in UUO-Model Mice
3.6. CTRP13 Suppresses Renal Inflammation in UUO-Model Mice
3.7. CTRP13 Suppresses NF-κB p65 Signaling Pathway in UUO-Model Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, H.T. Progression of chronic renal failure. Arch. Intern. Med. 2003, 163, 1417–1429. [Google Scholar] [CrossRef]
- Tonelli, M.; Karumanchi, S.A.; Thadhani, R. Epidemiology and Mechanisms of Uremia-Related Cardiovascular Disease. Circulation 2016, 133, 518–536. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Renal fibrosis: New insights into the pathogenesis and therapeutics. Kidney Int. 2006, 69, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Shabaka, A.; Casecs-Corona, C.; Fernandez-Juarez, G. Therapeutic Insights in Chronic Kidney Disease Progression. Front. Med. 2021, 8, 645187. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. Cellular and molecular mechanisms of renal fibrosis. Nat. Rev. Nephrol. 2011, 7, 684–696. [Google Scholar] [CrossRef]
- Zheng, Q.; Yuan, Y.; Yi, W.; Lau, W.B.; Wang, Y.; Wang, X.; Sun, Y.; Lopez, B.L.; Christopher, T.A.; Peterson, J.M.; et al. C1q/TNF-related proteins, a family of novel adipokines, induce vascular relaxation through the adiponectin receptor-1/AMPK/eNOS/nitric oxide signaling pathway. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2616–2623. [Google Scholar] [CrossRef]
- Xie, Y.; Meng, Z.; Gao, J.; Liu, C.; Wang, J.; Guo, R.; Zhao, J.; Lopez, B.; Christopher, T.; Lee, D.; et al. C1q Complement/Tumor Necrosis Factor-Associated Proteins in Cardiovascular Disease and COVID-19. Proteomes 2021, 9, 12. [Google Scholar] [CrossRef]
- Wei, Z.; Peterson, J.M.; Wong, G.W. Metabolic regulation by C1q/TNF-related protein-13 (CTRP13): Activation OF AMP-activated protein kinase and suppression of fatty acid-induced JNK signaling. J. Biol. Chem. 2011, 286, 15652–15665. [Google Scholar] [CrossRef]
- Bai, B.; Ban, B.; Liu, Z.; Zhang, M.M.; Tan, B.K.; Chen, J. Circulating C1q complement/TNF-related protein (CTRP); CTRP9, CTRP12 and CTRP13 concentrations in Type 2 diabetes mellitus: In vivo regulation by glucose. PLoS ONE 2017, 12, e0172271. [Google Scholar] [CrossRef]
- Li, Y.; Wang, W.; Chao, Y.; Zhang, F.; Wang, C. CTRP13 attenuates vascular calcification by regulating Runx2. FASEB J. 2019, 33, 9627–9637. [Google Scholar] [CrossRef]
- Kim, D.; Lee, A.S.; Jung, Y.J.; Yang, K.H.; Lee, S.; Park, S.K.; Kim, W.; Kang, K.P. Tamoxifen ameliorates renal tubulointerstitial fibrosis by modulation of estrogen receptor alpha-mediated transforming growth factor-beta1/Smad signaling pathway. Nephrol. Dial. Transplant. 2014, 29, 2043–2053. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.E.; Jeong, J.Y.; Lim, B.J.; Chang, Y.K.; Na, K.R.; Shin, Y.T.; Lee, K.W. Aliskiren ameliorates renal inflammation and fibrosis induced by unilateral ureteral obstruction in mice. J. Urol. 2011, 186, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ge, S.; Wang, Y.; Liu, Y.; Qiu, L.; Li, J.; Huang, X.; Sun, L. Puerarin Alleviates UUO-Induced Inflammation and Fibrosis by Regulating the NF-kappaB P65/STAT3 and TGFbeta1/Smads Signaling Pathways. Drug. Des. Devel. Ther. 2021, 15, 3697–3708. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, X.; Peng, W.; Liu, H.; Zhou, X.; Liang, L.; Xiang, J.; Zhang, H.; Wang, D.; Liu, L.; et al. EI24 alleviates renal interstitial fibrosis through inhibition of epithelial-mesenchymal transition and fibroblast activation. FASEB J. 2021, 35, e21239. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, W.; An, J.; Liang, M.; Li, Y.; Zhang, F.; Tong, Q.; Huang, K. Poly(ADP-ribose) polymerase 1 accelerates vascular calcification by upregulating Runx2. Nat. Commun. 2019, 10, 1203. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, F.; Wang, L.; Zhang, Y.; Li, X.; Huang, K.; Du, M.; Liu, F.; Huang, S.; Guan, Y.; et al. Poly(ADP-ribose) polymerase 1 promotes oxidative-stress-induced liver cell death via suppressing farnesoid X receptor alpha. Mol. Cell Biol. 2013, 33, 4492–4503. [Google Scholar] [CrossRef]
- Huang, R.; Fu, P.; Ma, L. Kidney fibrosis: From mechanisms to therapeutic medicines. Signal Transduct. Target. Ther. 2023, 8, 129. [Google Scholar] [CrossRef]
- Meran, S.; Steadman, R. Fibroblasts and myofibroblasts in renal fibrosis. Int. J. Exp. Pathol. 2011, 92, 158–167. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.C. NF-kappaB in inflammation and renal diseases. Cell Biosci. 2015, 5, 63. [Google Scholar] [CrossRef]
- Chen, T.K.; Knicely, D.H.; Grams, M.E. Chronic Kidney Disease Diagnosis and Management: A Review. JAMA 2019, 322, 1294–1304. [Google Scholar] [CrossRef]
- Wuhl, E.; Schaefer, F. Therapeutic strategies to slow chronic kidney disease progression. Pediatr. Nephrol. 2008, 23, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Wilmer, W.A.; Rovin, B.H.; Hebert, C.J.; Rao, S.V.; Kumor, K.; Hebert, L.A. Management of glomerular proteinuria: A commentary. J. Am. Soc. Nephrol. 2003, 14, 3217–3232. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Sarver, D.C.; Saqib, M.; Zhou, M.; Aja, S.; Seldin, M.M.; Wong, G.W. CTRP13 ablation improves systemic glucose and lipid metabolism. Mol. Metab. 2023, 78, 101824. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chao, Y.; Xu, W.; Liang, M.; Deng, S.; Zhang, D.; Huang, K. CTRP13 Preserves Endothelial Function by Targeting GTP Cyclohydrolase 1 in Diabetes. Diabetes 2020, 69, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, W.; Liang, M.; Huang, D.; Huang, K. CTRP13 inhibits atherosclerosis via autophagy-lysosome-dependent degradation of CD36. FASEB J. 2019, 33, 2290–2300. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Chao, Y.; Liang, M.; Huang, K.; Wang, C. CTRP13 Mitigates Abdominal Aortic Aneurysm Formation via NAMPT1. Mol. Ther. 2021, 29, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Schaffler, A.; Buechler, C. CTRP family: Linking immunity to metabolism. Trends Endocrinol. Metab. 2012, 23, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Seldin, M.M.; Tan, S.Y.; Wong, G.W. Metabolic function of the CTRP family of hormones. Rev. Endocr. Metab. Disord. 2014, 15, 111–123. [Google Scholar] [CrossRef]
- Xiang, H.; Xue, W.; Li, Y.; Zheng, J.; Ding, C.; Dou, M.; Wu, X. C1q/TNF-related protein 6 (CTRP6) attenuates renal ischaemia-reperfusion injury through the activation of PI3K/Akt signalling pathway. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1030–1040. [Google Scholar] [CrossRef]
- Hu, T.Y.; Li, L.M.; Pan, Y.Z. CTRP3 inhibits high glucose-induced human glomerular mesangial cell dysfunction. J. Cell Biochem. 2019, 120, 5729–5736. [Google Scholar] [CrossRef]
- Xu, E.; Yin, C.; Yi, X.; Liu, Y. Knockdown of CTRP6 inhibits high glucose-induced oxidative stress, inflammation and extracellular matrix accumulation in mesangial cells through regulating the Akt/NF-kappaB pathway. Clin. Exp. Pharmacol. Physiol. 2020, 47, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Koinuma, D.; Shinozaki, M.; Komuro, A.; Goto, K.; Saitoh, M.; Hanyu, A.; Ebina, M.; Nukiwa, T.; Miyazawa, K.; Imamura, T.; et al. Arkadia amplifies TGF-beta superfamily signalling through degradation of Smad7. EMBO J. 2003, 22, 6458–6470. [Google Scholar] [CrossRef] [PubMed]
- Kavsak, P.; Rasmussen, R.K.; Causing, C.G.; Bonni, S.; Zhu, H.; Thomsen, G.H.; Wrana, J.L. Smad7 binds to Smurf2 to form an E3 ubiquitin ligase that targets the TGF beta receptor for degradation. Mol. Cell 2000, 6, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Feinberg, M.W.; Shimizu, K.; Lebedeva, M.; Haspel, R.; Takayama, K.; Chen, Z.; Frederick, J.P.; Wang, X.F.; Simon, D.I.; Libby, P.; et al. Essential role for Smad3 in regulating MCP-1 expression and vascular inflammation. Circ. Res. 2004, 94, 601–608. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wang, W.; Liu, C.; Zeng, M.; Xu, L.; Du, R.; Wang, C. Adiponectin C1q/Tumor Necrosis Factor-Related Protein 13 (CTRP13) Protects against Renal Inflammation and Fibrosis in Obstructive Nephropathy. Biomedicines 2024, 12, 51. https://doi.org/10.3390/biomedicines12010051
Li Y, Wang W, Liu C, Zeng M, Xu L, Du R, Wang C. Adiponectin C1q/Tumor Necrosis Factor-Related Protein 13 (CTRP13) Protects against Renal Inflammation and Fibrosis in Obstructive Nephropathy. Biomedicines. 2024; 12(1):51. https://doi.org/10.3390/biomedicines12010051
Chicago/Turabian StyleLi, Yongxia, Wenzhe Wang, Changxuan Liu, Min Zeng, Li Xu, Rong Du, and Cheng Wang. 2024. "Adiponectin C1q/Tumor Necrosis Factor-Related Protein 13 (CTRP13) Protects against Renal Inflammation and Fibrosis in Obstructive Nephropathy" Biomedicines 12, no. 1: 51. https://doi.org/10.3390/biomedicines12010051
APA StyleLi, Y., Wang, W., Liu, C., Zeng, M., Xu, L., Du, R., & Wang, C. (2024). Adiponectin C1q/Tumor Necrosis Factor-Related Protein 13 (CTRP13) Protects against Renal Inflammation and Fibrosis in Obstructive Nephropathy. Biomedicines, 12(1), 51. https://doi.org/10.3390/biomedicines12010051