

Different Effects of Phototherapy for Rat Glioma during Sleep and Wakefulness

 , , , , ,
, , , , ,  , , add
Show full author list
, , add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Model of Rat Glioma
2.3. PBM under EEG Control
2.4. Optical Monitoring of Brain’s Drainage System
2.5. MRI of Rat Glioma
2.6. Immunohistochemistry (IHC)
2.7. Statistical Analysis
3. Results
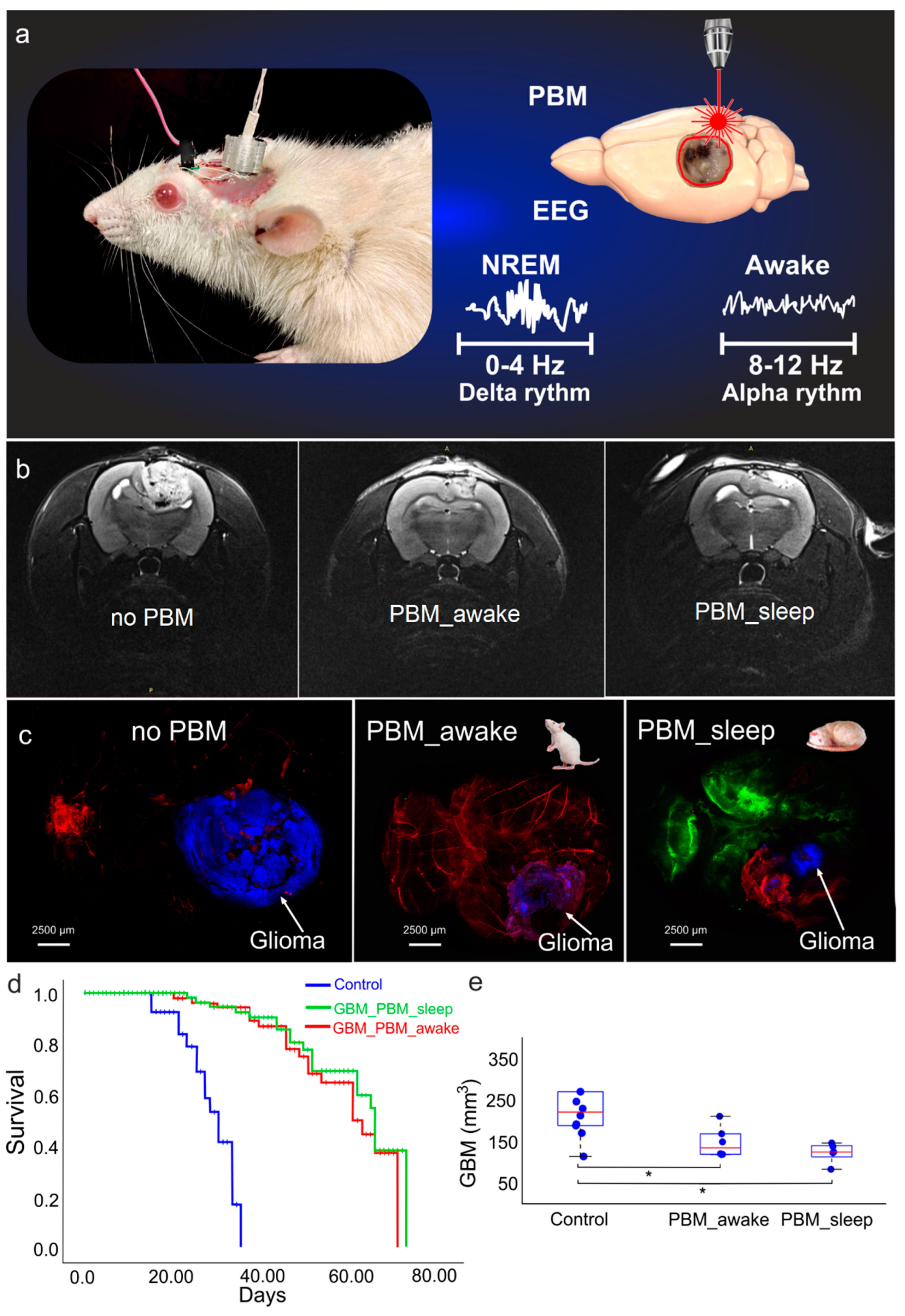
3.1. The Effects of the PBM Course during Sleep or Awake State on Survival and Glioma Progression
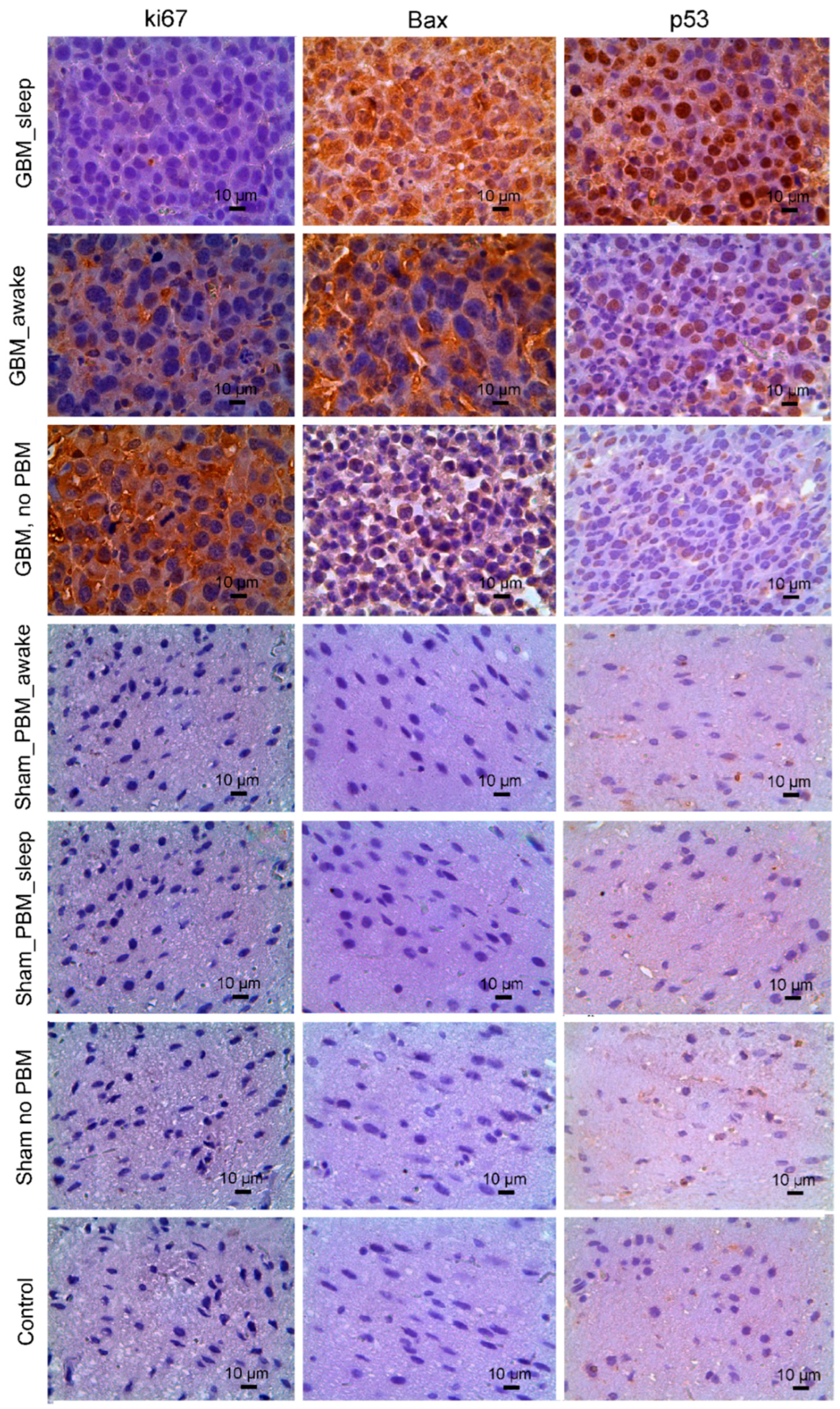
3.2. The Effects of the PBM Course during Sleep or Awake State on Apoptosis and Proliferation of Glioma Cells
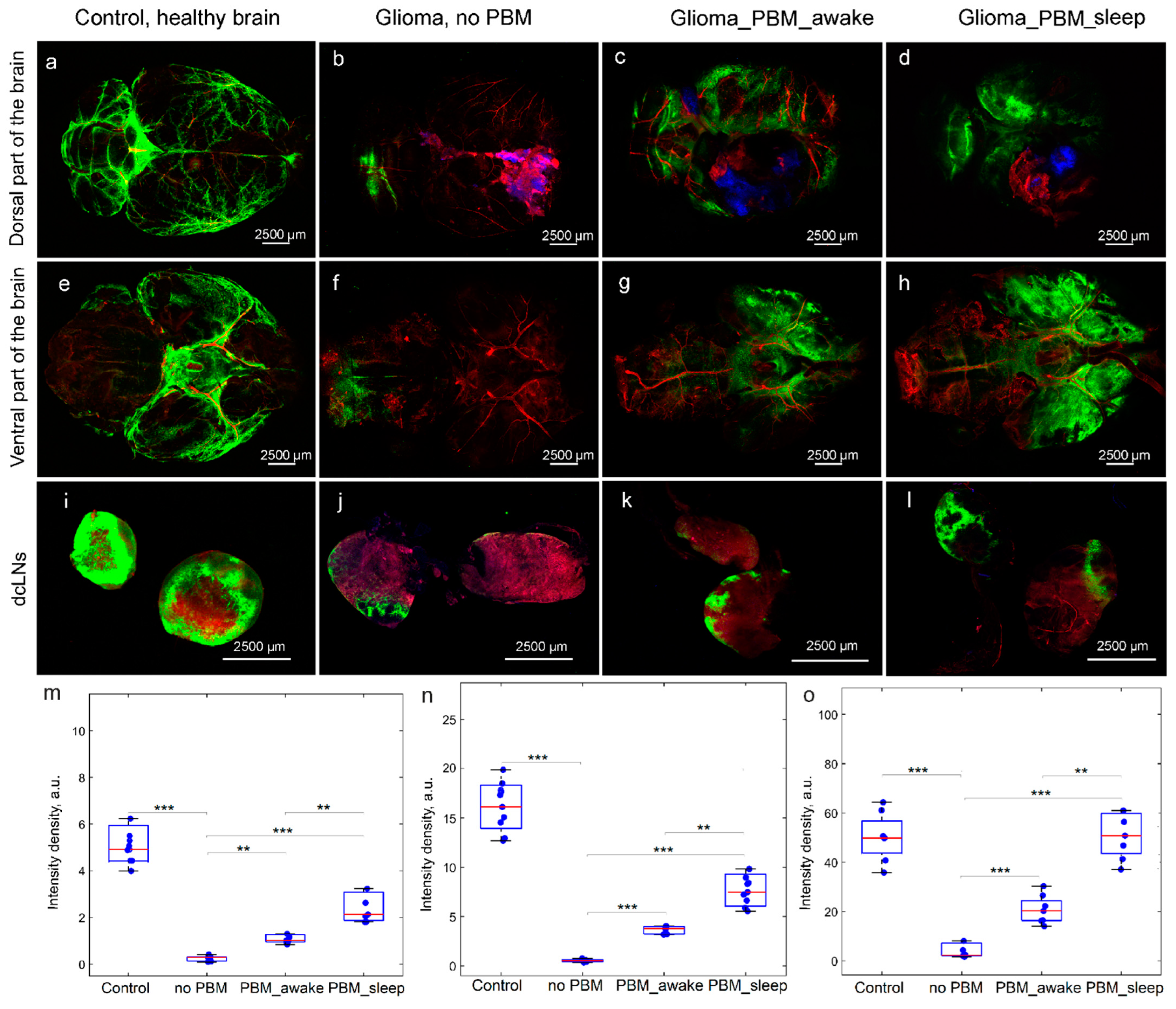
3.3. The Effects of the PBM Course during Sleep or Awake State on BD
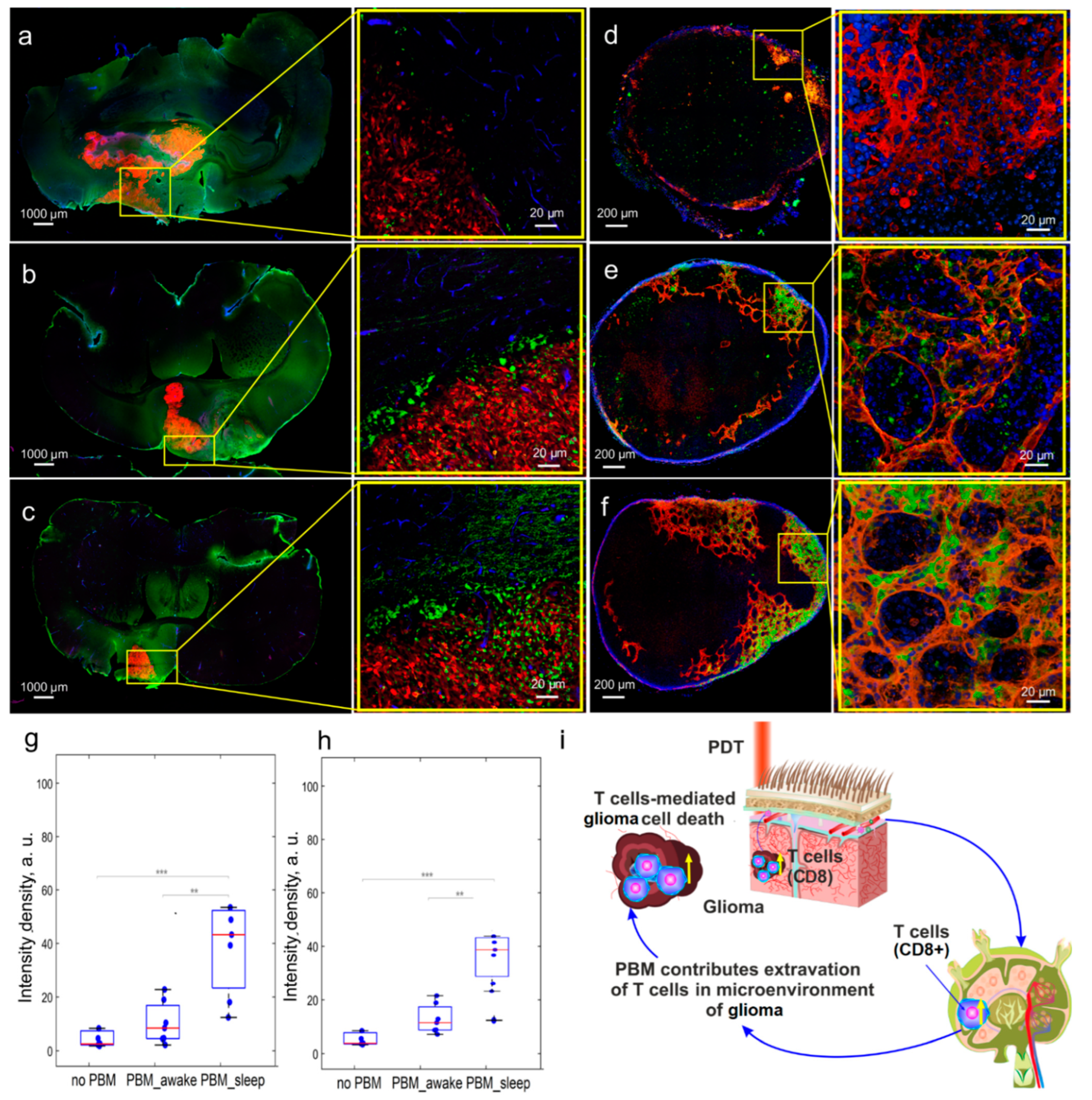
3.4. The Effects of the PBM Course during Sleep or Wakefulness on Brain’s Tumor Immunity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Noorvash, B.; Amaral, L.J.; Nelson, T.; Rudnick, J.D.; Irwin, S.A.; Freedland, S.J.; Gresham, G.; Hu, J.L. Remote monitoring of sleep activity in patients diagnosed with glioblastoma. J. Clin. Oncol. 2022, 16, e14036. [Google Scholar] [CrossRef]
- Martin, J.A.; Hart, N.H.; Bradford, N. Prevalence and management of sleep disturbance in adults with primary brain tumours and their caregivers: A systematic review. J. Neurooncol. 2023, 162, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Büttner-Teleagă, A.; Kim, Y.T.; Osel, T.; Richter, K. Sleep disorders in cancer—A systematic review. Int. J. Env. Res. Public. Health 2021, 18, 11696–11734. [Google Scholar] [CrossRef] [PubMed]
- Otte, J.L.; Carpenter, J.S.; Manchanda, S.; Rand, K.L.; Skaar, T.C.; Weaver, M. Systematic review of sleep disorders in cancer patients: Can the prevalence of sleep disorders be ascertained? Cancer Med. 2015, 4, 183–200. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.S.; Dhillon, H.M.; Agar, M.R. Sleep disturbance of adults with a brain tumor and their family caregivers: A systematic review. Neuro Oncol. 2017, 19, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, T.S.; Shade, M.Y.; Breton, G.; Gilbert, M.R.; Mahajan, A.; Scheurer, M.E. Sleep-wake disturbance in patients with brain tumors. Neuro Oncol. 2016, 19, 323–335. [Google Scholar] [CrossRef]
- Fox, S.W.; Lyon, D.; Farace, E. Symptom clusters in patients with high-grade glioma. J. Nurs. Sch. 2007, 39, 61–67. [Google Scholar] [CrossRef]
- Huang, Y.; Jiang, Z.-J.; Deng, J.; Qi, Y.-J. Sleep quality of patients with postoperative glioma at home. World J. Clin. Case 2020, 8, 4735–4742. [Google Scholar] [CrossRef]
- Lin, M.R.; Chen, P.Y.; Wang, H.C.; Lin, P.C.; Lee, H.C.; Chiu, H.Y. Prevalence of sleep disturbances and their effects on quality of life in adults with untreated pituitary tumor and meningioma. J. Neurooncol. 2021, 154, 179–186. [Google Scholar] [CrossRef]
- Willis, K.D.; Ravyts, S.G.; Lanoye, A.; Loughan, A.R. Sleep disturbance in primary brain tumor: Prevalence, risk factors, and patient preferences. Support. Care Cancer 2022, 30, 741–748. [Google Scholar] [CrossRef]
- Allgood, J.; Roe, A.; Sparks, B.; Castillo, M.; Cruz, A.; Brooks, A.; Brooks, B. The Correlation of Sleep Disturbance and Location of Glioma Tumors: A Narrative Review. J. Clin. Med. 2023, 12, 4058. [Google Scholar] [CrossRef] [PubMed]
- Sadighi, Z.; Curtis, E.; Zabrowksi, J.; Billups, C.; Gajjar, A.; Khan, R.; Qaddoumi, I. Neurologic impairments from pediatric low-grade glioma by tumor location and timing of diagnosis. Pediatr. Blood Cancer 2018, 65, 27063. [Google Scholar] [CrossRef]
- Subramanian, S.; Gholami, A.; Biros, G. Simulation of glioblastoma growth using a 3D multispecies tumor model with mass effect. J. Math. Biol. 2019, 79, 941–967. [Google Scholar] [CrossRef]
- Gapstur, R.; Gross, C.; Ness, K. Factors associated with sleep-wake disturbances in child and adult survivors of pediatric brain tumors: A review. Oncol. Nurs. Forum 2009, 36, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Parker, K.; Young-McCaughan, S.; Mallory, G.; Barsevick, A.; Beck, S.; Carpenter, J.; Carter, P.; Farr, L.; Hinds, P.; et al. Sleep wake disturbances in people with cancer and their caregivers: State of the science. Oncol. Nurs. Forum 2005, 32, E98–E126. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Schlegel, F.; Bachmann, S.B. Lymphatic outflow of cerebrospinal fluid is reduced in glioma. Sci. Rep. 2019, 9, 14815. [Google Scholar] [CrossRef]
- Hu, X.; Deng, Q.; Ma, L. Meningeal lymphatic vessels regulate brain tumor drainage and immunity. Cell Res. 2020, 30, 229–243. [Google Scholar] [CrossRef]
- Song, E.; Mao, T.; Dong, H. VEGF-C-driven lymphatic drainage enables immunosurveillance of brain tumours. Nature 2020, 577, 689–694. [Google Scholar] [CrossRef]
- Xie, L.; Kand, H.; Xu, Q. Sleep drives metabolite clearance from the adult brain. Science 2013, 342, 373–377. [Google Scholar] [CrossRef]
- Fultz, N.E.; Bonmassar, G.; Setsompop, K. Coupled electrophysiological, hemodynamic, and cerebrospinal fluid oscillations in human sleep. Science 2019, 366, 628–631. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Penzel, T.; Blokhina, I.; Khorovodov, A.; Fedosov, I.; Yu, T.; Karandin, G.; Evsukova, A.; Elovenko, D.; Adushkina, V.; et al. Night photostimulation of clearance of beta-amyloid from mouse brain: New strategies in preventing Alzheimer’s disease. Cells 2021, 10, 3289. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.; Shirokov, A.; Blokhina, I.; Fedosov, I.; Terskov, A.; Dubrovsky, A.; Tsoy, M.; Zlatogosrskaya, D.; Adushkina, V.; Evsukova, A.; et al. Mechanisms of phototherapy of Alzheimer’s disease during sleep and wakefulness: The role of the meningeal lymphatics. Front. Optoelectron. 2023, 16, 22. [Google Scholar]
- Qin, X.; Liu, R.; Akter, F.; Qin, L.; Xie, Q.; Li, Y.; Qiao, H.; Zhao, W.; Jian, Z.; Liu, R.; et al. Peri-tumoral brain edema associated with glioblastoma correlates with tumor recurrence. J. Cancer 2021, 12, 2073–2082. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Sokolovski, S.; Fedosov, I.; Shirokov, A.; Navolokin, N.; Bucharskaya, A.; Blokhina, I.; Terskov, A.; Dubrovski, A.; Telnova, V.; et al. Transcranial photosensitiser-free laser treatment of glioblastoma in rat brain. Int. J. Mol. Sci. 2023, 24, 13696. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.; Abdurashitov, A.; Dubrovsky, A.; Klimova, M.; Agranovich, I.; Terskov, A.; Shirokov, A.; Vinnik, V.; Kuznecova, A.; Lezhnev, N.; et al. Photobiomodulation of lymphatic drainage and clearance: Perspective strategy for augmentation of meningeal lymphatic functions. Biomed Opt. Express 2020, 11, 725–734. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Abdurashitov, A.; Klimova, M.; Dubrovsky, A.; Shirokov, A.; Fomin, A.; Terskov, A.; Agranovich, I.; Mamedova, A.; Khorovodov, A.; et al. Photostimulation of cerebral and peripheral lymphatic functions. Transl. Biophotonics 2020, 2, e201900036. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Fedosov, I.; Shirokov, A.; Vodovozova, E.; Alekseeva, A.; Khorovodov, A.; Blokhina, I.; Terskov, A.; Mamedova, A.; Klimova, M.; et al. Photomodulation of lymphatic delivery of liposomes to the brain bypassing the blood-brain barrier: New perspectives for glioma therapy. Nanophotonics 2021, 12, 3215–3227. [Google Scholar] [CrossRef]
- Li, D.; Liu, S.; Yu, T.-T.; Liu, Z.; Sub, S.; Bragin, D.; Shirokov, A.; Navolokin, N.; Bragina, O.; Zheng-Wu, H.; et al. Photostimulation of brain lymphatics in male newborn and adult rodents for therapy of intraventricular hemorrhage. Nat. Comm. 2023, 14, 6104. [Google Scholar] [CrossRef]
- Bohlen, H.; Gasheva, O.; Zawieja, D. Nitric oxide formation by lymphatic bulb and valves is a major regulatory component of lymphatic pumping. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H1897–H1906. [Google Scholar] [CrossRef]
- Bohlen, H.; Wang, W.; Gashev, A.; Gasheva, O.; Zawieja, D. Phasic contractions of rat mesenteric lymphatics increase basal and phasic nitric oxide generation in vivo. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1319–H1328. [Google Scholar] [CrossRef]
- Kunert, C.; Baish, J.W.; Liao, S.; Padera, T.P.; Munn, L.L. Mechanobiological oscillators control lymph flow. Proc. Natl. Acad. Sci. USA 2015, 112, 10938–10943. [Google Scholar] [CrossRef]
- Petrelli, F.; De Stefani, A.; Ghidini, A.; Bruschieri, L.; Riboldi, V.; Dottorini, L.; Iaculli, A.; Zaniboni, A.; Trevisan, F. Steroids use and survival in patients with glioblastoma multiforme: A pooled analysis. J. Neurol. 2021, 268, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Zoccarato, M.; Nardetto, L.; Basile, A.M.; Giometto, B.; Zagonel, V.; Lombardi, G. Seizures, Edema, Thrombosis, and Hemorrhages: An Update Review on the Medical Management of Gliomas. Front. Oncol. 2021, 11, 617966. [Google Scholar] [CrossRef]
- Ohmura, K.; Tomita, H.; Hara, A. Peritumoral Edema in Gliomas: A Review of Mechanisms and Management. Biomedicines 2023, 11, 2731. [Google Scholar] [CrossRef] [PubMed]
- Semyachkina-Glushkovskaya, O.; Fedosov, I.; Zaikin, A.; Ageev, V.; Ilyukov, E.; Myagkov, D.; Tuktarov, D.; Blokhina, I.; Shirokov, A.; Terskov, A.; et al. Technology of the photobiostimulation of the brain’s drainage system during sleep for improvement of learning and memory in male mice. Biomed. Opt. Express 2024, 15, 44–58. [Google Scholar] [CrossRef]
- Hablitz, L.M.; Vinitsky, H.S.; Sun, Q. Increased glymphatic influx is correlated with high EEG delta power and low heart rate in mice under anesthesia. Sci. Adv. 2019, 5, eaav5447. [Google Scholar] [CrossRef]
- Iber, C. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications, 1st ed.; American Academy of Sleep Medicine: Darien, IL, USA, 2007. [Google Scholar]
- Devos, S.L.; Miller, T.M. Direct intraventricular delivery of drugs to the rodent central nervous system. J. Vis. Exp. 2013, 12, e50326. [Google Scholar]
- Ahn, J.H.; Cho, H.; Kim, J. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature 2019, 572, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]
- Bragin, D.; Kameneva, M.; Bragina, O.; Thomson, S.; Statom, G.; Lara, D.; Yang, Y.; Nemoto, E. Rheological effects of drag-reducing polymers improve cerebral blood flow and oxygenation after traumatic brain injury in rats. J. Cereb. Blood Flow. Metab. 2017, 37, 762–775. [Google Scholar] [CrossRef]
- Mauldin, I.S.; Jo, J.; Wages, N.A.; Yogendran, L.V.; Mahmutovic, A.; Young, S.J.; Lopes, M.B.; Slingluff, C.L.; Erickson, L.D.; Fadul, C.E. Proliferating CD8+ T cell infiltrates are associated with improved survival in glioblastoma. Cells 2021, 10, 3378. [Google Scholar] [CrossRef]
- Ferrantelli, F.; Manfredil, F.; Chiozzini, C.; Leone, P.; Giovannelli, A.; Olivetta, E.; Federico, M. Long-term antitumor CD8+ T cell immunity induced by endogenously engineered extracellular vesicles. Cancers 2021, 13, 2263. [Google Scholar] [CrossRef] [PubMed]
- Raskov, H.; Orhan, A.; Christensen, J.P. Cytotoxic CD8+ T cells in cancer and cancer immunotherapy. Br. J. Cancer 2021, 124, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Philip, M.; Schietinger, A. CD8+ T cell differentiation and dysfunction in cancer. Nat. Rev. Immunol. 2022, 22, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A guide to cancer immunotherapy: From T cell basic science to clinical practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Zhang, Y.; Xu, J.; Yin, T.; Lu, X.J. T cell dysfunction in cancer immunity and immunotherapy. Front. Immunol. 2019, 10, 1719. [Google Scholar] [CrossRef]
- Nakano, O.; Sato, M.; Naito, Y.; Suzuki, K.; Orikasa, S.; Aizawa, M.; Suzuki, Y.; Shintaku, I.; Nagura, H.; Ohtani, H. Proliferative activity of intratumoral CD8+ T-lymphocytes as a prognostic factor in human renal cell carcinoma: Clinicopathologic demonstration of antitumor immunity. Cancer Res. 2001, 61, 5132–5136. [Google Scholar]
- Xie, Q.; Ding, J.; Chen, Y. Role of CD8+ T lymphocyte cells: Interplay with stromal cells in tumor microenvironment. Acta Pharm. Sin. B 2021, 11, 1365–1378. [Google Scholar] [CrossRef]
- Domka, W.; Bartusik-Aebisher, D.; Rudy, I.; Dynarowicz, K.; Pięta, K.; Aebisher, D. Photodynamic therapy in brain cancer: Mechanisms, clinical and preclinical studies and therapeutic challenges. Front. Chem. 2023, 11, 1250621. [Google Scholar] [CrossRef]
- Abdurashitov, A.; Tuchin, V.; Semyachkina-Glushkovskaya, O. Photodynamic therapy of brain tumors and novel optical coherence tomography strategies for in vivo monitoring of cerebral fluid dynamics. J. Innov. Opt. Health Sci. 2020, 13, 2030004. [Google Scholar] [CrossRef]
- Bartusik-Aebisher, D.; Żołyniak, A.; Barnaś, E.; Machorowska-Pieniążek, A.; Oleś, P.; Kawczyk-Krupka, A.; Aebisher, D. The Use of Photodynamic Therapy in the Treatment of Brain Tumors-A Review of the Literature. Molecules 2022, 27, 6847. [Google Scholar] [CrossRef] [PubMed]
- Stepp, H.; Beck, T.; Pongraz, T.; Meinel, F.; Kreth, W.; Tonn, J.; Stummer, W. ALA and malignant glioma: Fluorescence-guided resection and photodynamic treatment. J. Environ. Pathol. Toxicol. Oncol. 2007, 21, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, K.; Garvey, K.; Bouras, A.; Cramer, G.; Stepp, H.; Jesu Raj, J.; Bozec, D.; Busch, T.; Hadjipanayis, C. 5-aminolevulinic acid photodynamic therapy for the treatment of high-grade gliomas. J. Neurooncol. 2019, 141, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Correia, J.; Rodrigues, J.; Pimenta, S.; Dong, T.; Yang, Z. Photodynamic Therapy Review: Principles, Photosensitizers, Applications, and Future Directions. Pharmaceutics 2021, 13, 1332. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, P.; Xie, Y.; Yang, J.; Yang, Y.; Shi, L.; Wu, W.; Li, Z. Photosensitizers with multiple degradation modes for efficient and postoperatively safe photodynamic therapy. Biomaterials 2023, 299, 122182. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lovell, J.; Yoon, J.; Chen, X. Clinical development and potential of photothermal and photodynamic therapies for cancer. Nat. Rev. Clin. Oncol. 2020, 17, 657–674. [Google Scholar] [CrossRef] [PubMed]
- Wojewoda, K.; Gillstedt, M.; Tovi, J.; Salah, L.; Wennberg Larko, A.; Sjoholm, A.; Sandberg, C. Optimizing treatment of acne with photodynamic therapy (PDT) to achieve long-term remission and reduce side effects. A prospective randomized controlled trial. J. Photochem. Photobiol. B 2021, 223, 112299. [Google Scholar] [CrossRef]
- Ibbotson, S.; Wong, T.; Morton, C.; Collier, N.; Haylett, A.; McKenna, K.; Mallipeddi, R.; Moseley, H.; Rhodes, L.E.; Seukeran, D.C.; et al. Adverse effects of topical photodynamic therapy: A consensus review and approach to management. Br. J. Dermatol. 2019, 180, 715–729. [Google Scholar] [CrossRef]
- Morton, C.; Szeimies, R.; Basset-Seguin, N.; Calzavara-Pinton, P.; Gilaberte, Y.; Haedersdal, M.; Hofbauer, G.F.L.; Hunger, R.E.; Karrer, S.; Piaserico, S.; et al. European dermatology forum guidelines on topical photodynamic therapy 2019 part 2: Emerging indications—Field cancerization, photorejuvenation and inflammatory/infective dermatoses. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 17–29. [Google Scholar] [CrossRef]
- Semyachkina-Glushkovskaya, O.; Fedosov, I.; Penzel, T.; Li, D.; Yu, T.; Telnova, V.; Kaybeleva, E.; Saranceva, E.; Terskov, A.; Khorovodov, A.; et al. Brain Waste Removal System and Sleep: Photobiomodulation as an Innovative Strategy for Night Therapy of Brain Diseases. Int. J. Mol. Sci. 2023, 24, 3221. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Markers | Control (Healthy Rats) | Control (Sham Rats) | Control (Sham Rats PBM_Sleep) | Control (Sham Rats PBM_Awake) | Rats with Glioma, No PBM | Rats with Glioma, PBM_Awake | Rats with Glioma, PBM_Sleep |
|---|---|---|---|---|---|---|---|
| Ki67 | - | - | - | - | 98.87 ± 1.1 | 27.44 ± 6.0 *** | 11.69 ± 1.0 ***††† |
| Bax | - | - | - | - | 16.29 ± 4.3 | 65.33 ± 7.5 ** | 96.91 ± 3.8 ***†† |
| p53 | - | - | - | - | 20.25 ± 6.1 | 47.00 ± 4.4 ** | 87.33 ± 7.4 ***††† |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shirokov, A.; Blokhina, I.; Fedosov, I.; Ilyukov, E.; Terskov, A.; Myagkov, D.; Tuktarov, D.; Tzoy, M.; Adushkina, V.; Zlatogosrkaya, D.; et al. Different Effects of Phototherapy for Rat Glioma during Sleep and Wakefulness. Biomedicines 2024, 12, 262. https://doi.org/10.3390/biomedicines12020262
Shirokov A, Blokhina I, Fedosov I, Ilyukov E, Terskov A, Myagkov D, Tuktarov D, Tzoy M, Adushkina V, Zlatogosrkaya D, et al. Different Effects of Phototherapy for Rat Glioma during Sleep and Wakefulness. Biomedicines. 2024; 12(2):262. https://doi.org/10.3390/biomedicines12020262
Chicago/Turabian StyleShirokov, Alexander, Inna Blokhina, Ivan Fedosov, Egor Ilyukov, Andrey Terskov, Dmitry Myagkov, Dmitry Tuktarov, Maria Tzoy, Viktoria Adushkina, Daria Zlatogosrkaya, and et al. 2024. "Different Effects of Phototherapy for Rat Glioma during Sleep and Wakefulness" Biomedicines 12, no. 2: 262. https://doi.org/10.3390/biomedicines12020262
APA StyleShirokov, A., Blokhina, I., Fedosov, I., Ilyukov, E., Terskov, A., Myagkov, D., Tuktarov, D., Tzoy, M., Adushkina, V., Zlatogosrkaya, D., Evsyukova, A., Telnova, V., Dubrovsky, A., Dmitrenko, A., Manzhaeva, M., Krupnova, V., Tuzhilkin, M., Elezarova, I., Navolokin, N., ... Semyachkina-Glushkovskaya, O. (2024). Different Effects of Phototherapy for Rat Glioma during Sleep and Wakefulness. Biomedicines, 12(2), 262. https://doi.org/10.3390/biomedicines12020262