
The Development of a Regulator of Human Serine Racemase for N-Methyl-D-aspartate Function
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Role of NMDA Receptor and Its Regulatory Enzyme Serine Racemase
1.2. NMDA Receptor Dysfunction and Its Potential Drug Target
1.3. Research Objectives
2. Materials and Methods
2.1. Analysis of D-Amino Acid (DAA) Concentration in C57BL/6J Mice Brain
2.2. Purification of Recombinant hSR
2.2.1. SDS-PAGE and Native Polyacrylamide Gel Electrophoresis
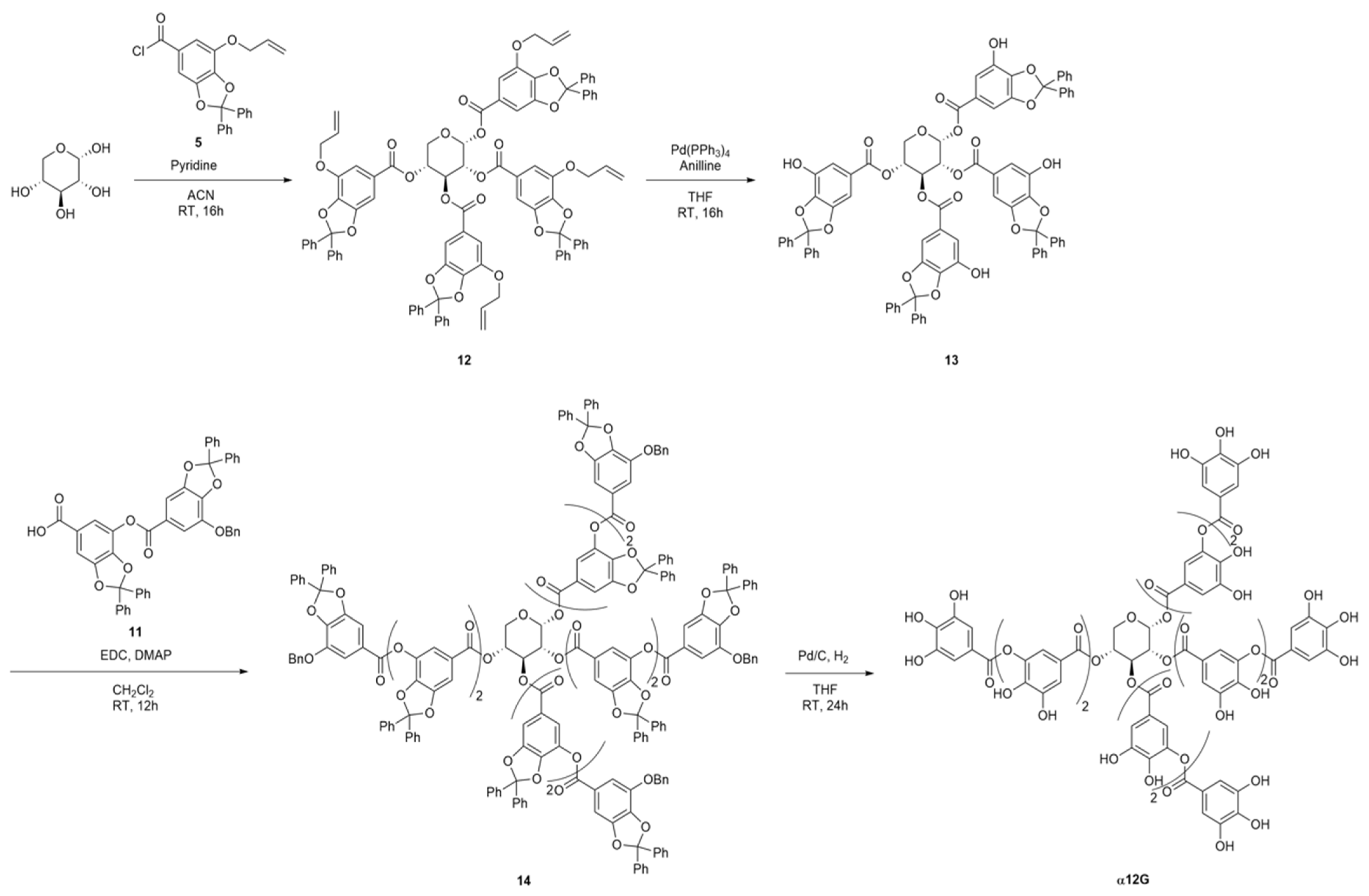
2.2.2. Preparation of α12G
2.3. Two-Step Enzymatic Assay
2.4. DAAO Enzymatic Assay
2.5. Molecular Modeling
2.6. Intrinsic Tryptophan Fluorescence (ITF)
2.7. Animals
2.8. Drug Administration
2.9. Open Field Test
2.10. Prepulse Inhibition Test
2.11. Novel Object Recognition
3. Results
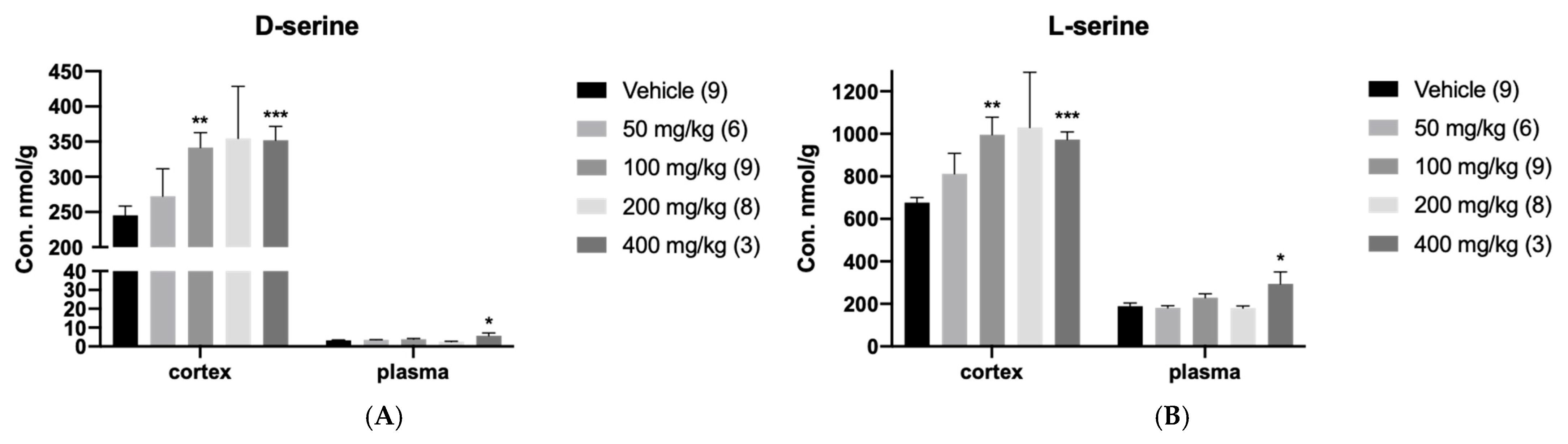
3.1. In Vivo Regulation of hSR by DMM
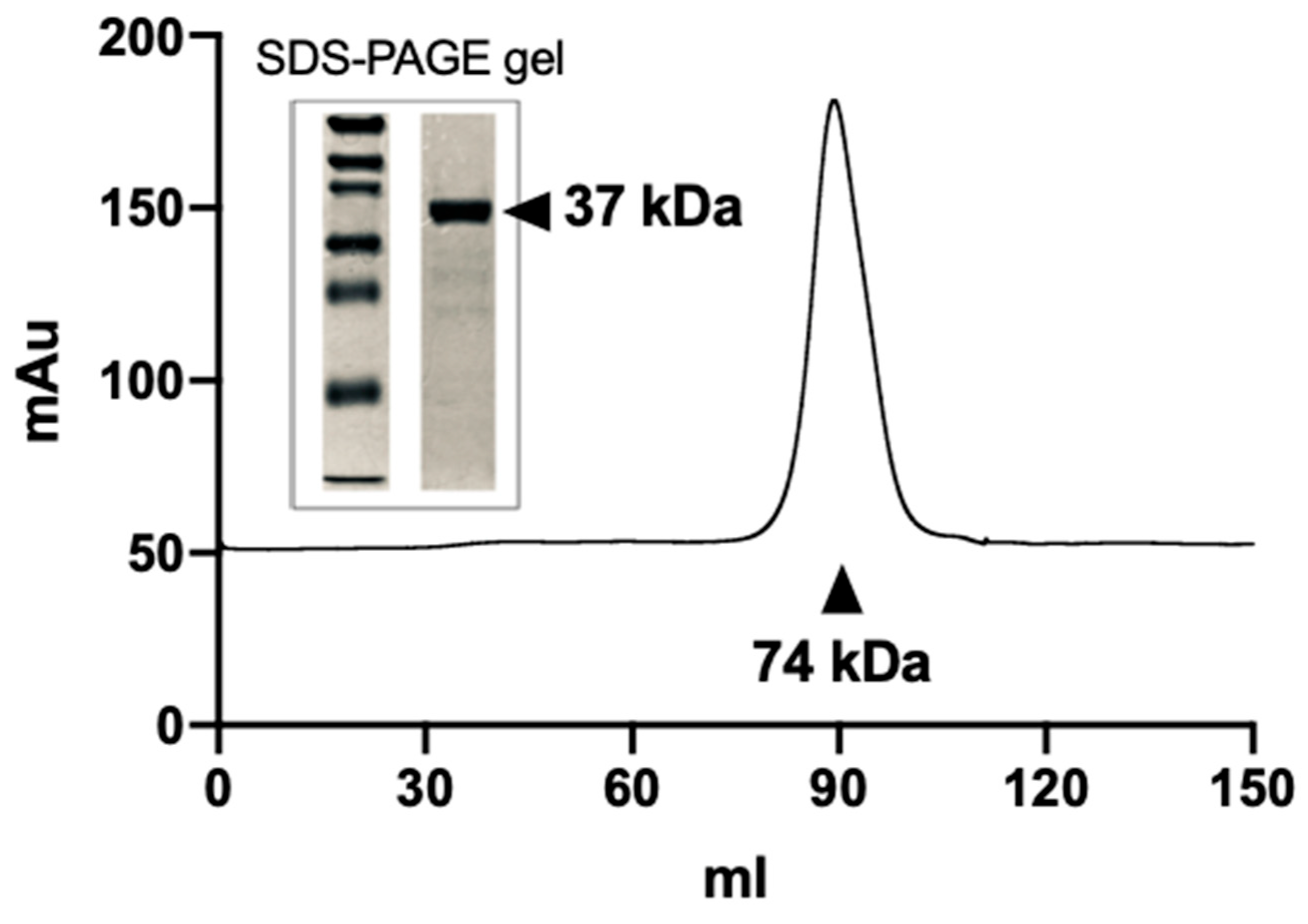
3.2. Purification of the Recombinant hSR Protein
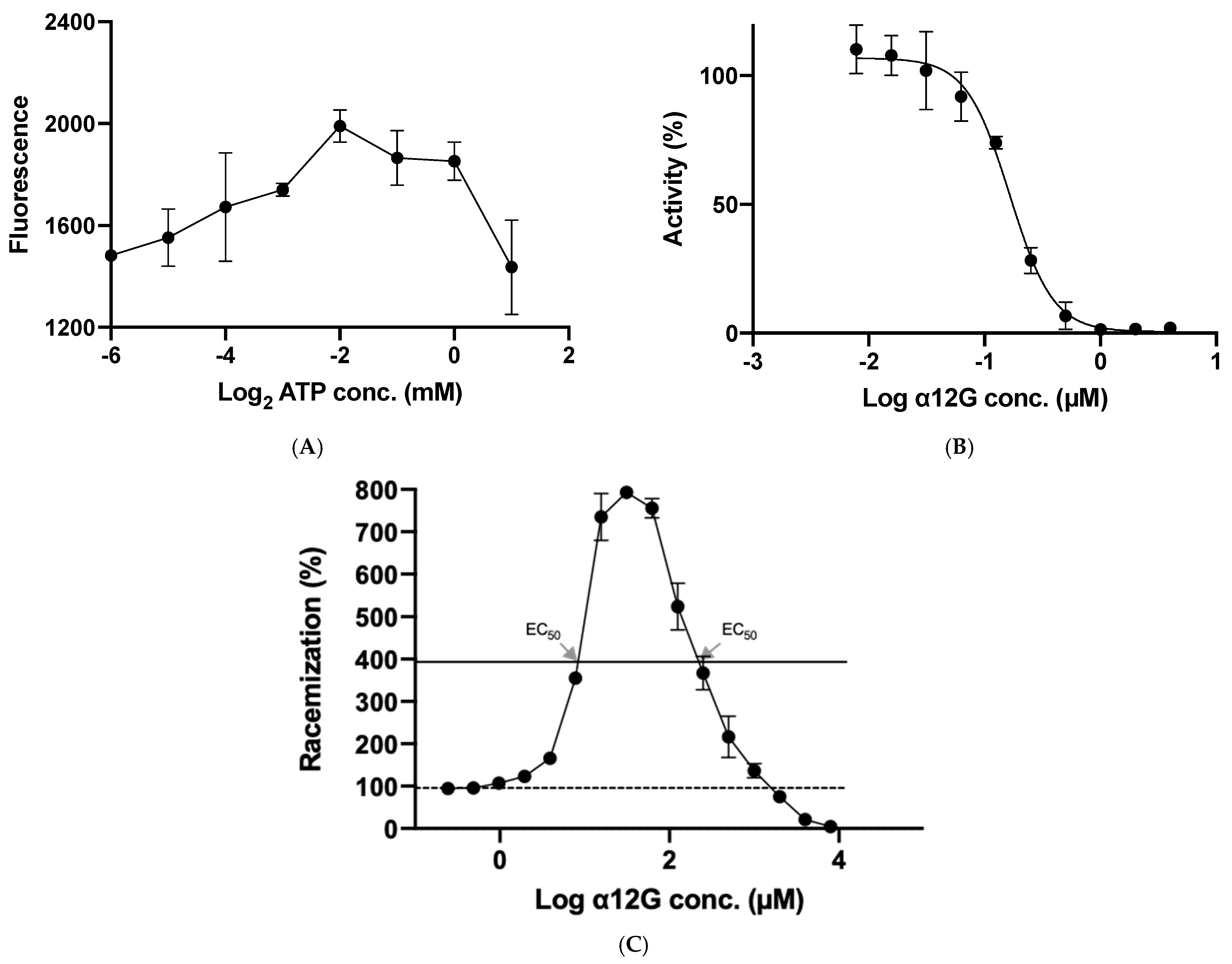
3.3. Enzymatic Characterization of hSR
3.4. In Vitro Regulatory Function of α12G on hSR
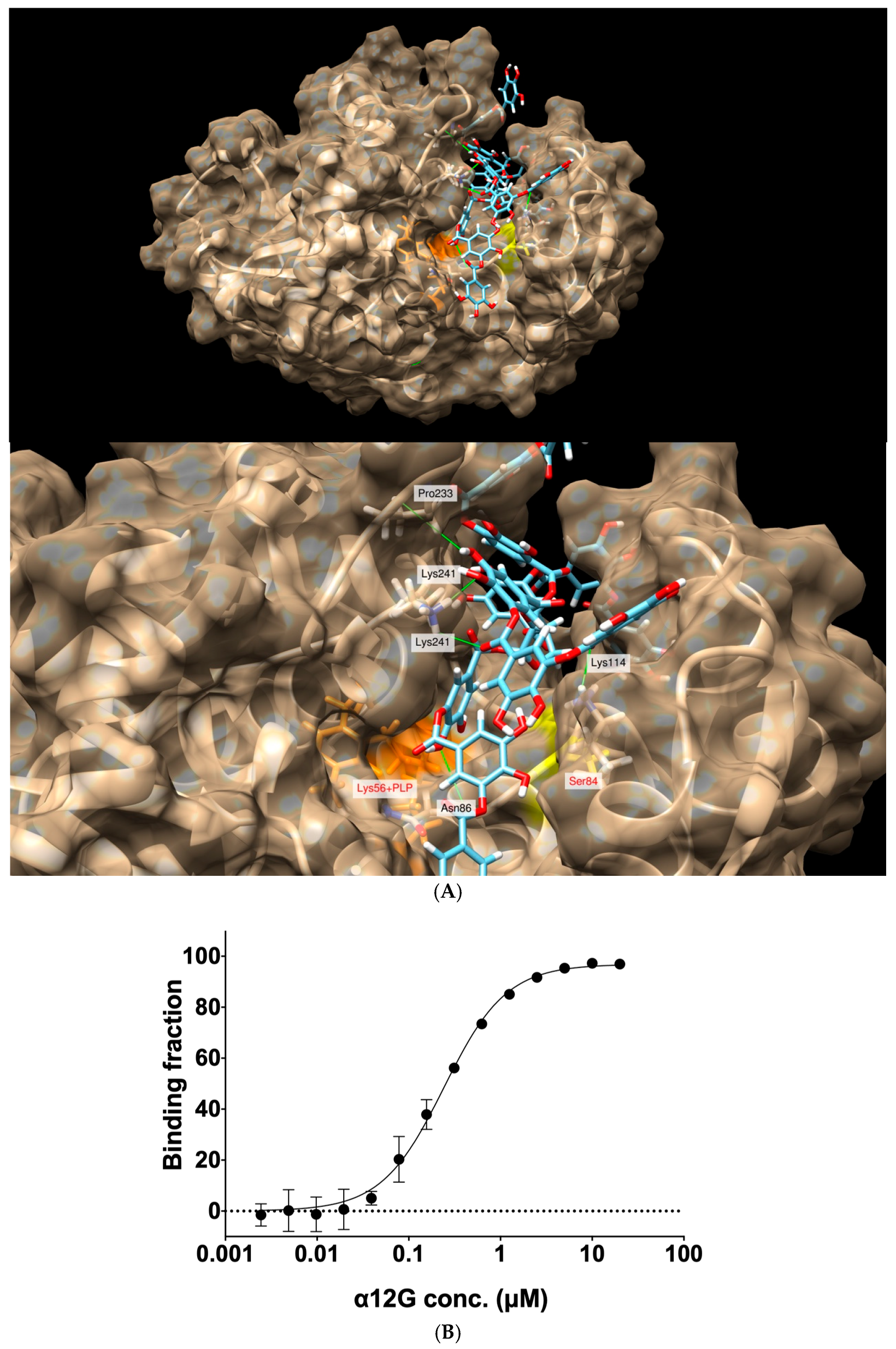
3.5. Interactions between the Active Site Residues of hSR with α12G
3.6. The Binding Affinity of α12G for hSR
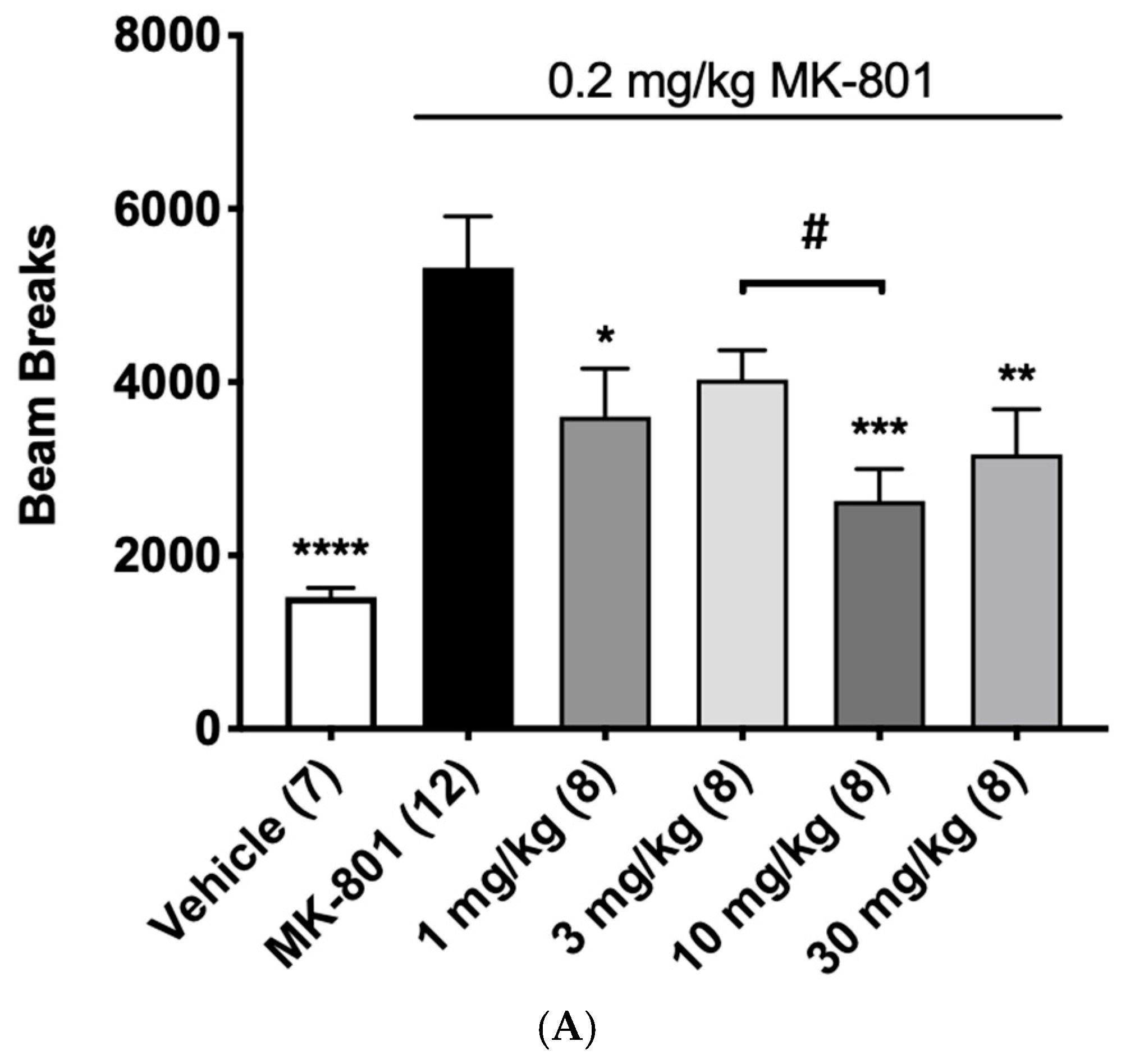
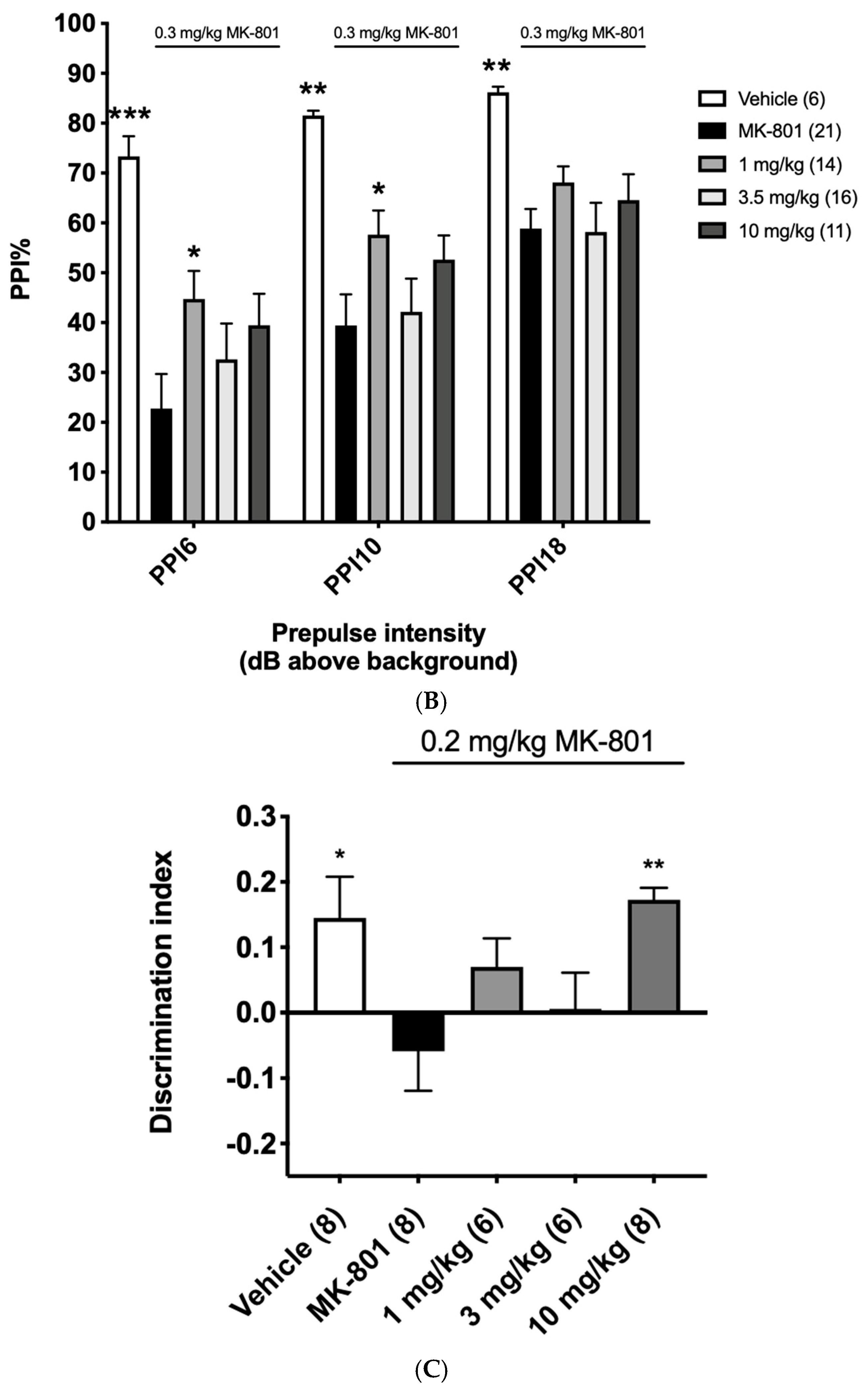
3.7. Pharmacodynamic Effects of α12G on Rodent NMDA Hypofunction Models
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Foltyn, V.N.; Bendikov, I.; De Miranda, J.; Panizzutti, R.; Dumin, E.; Shleper, M.; Li, P.; Toney, M.D.; Kartvelishvily, E.; Wolosker, H. Serine racemase modulates intracellular D-serine levels through an alpha, beta-elimination activity. J. Biol. Chem. 2005, 280, 1754–1763. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Yamauchi, T.; Kamiya, N.; Miyahara, I.; Yoshimura, T.; Mihara, H.; Kurihara, T.; Hirotsu, K.; Esaki, N. Crystal structure of a homolog of mammalian serine racemase from Schizosaccharomyces pombe. J. Biol. Chem. 2009, 284, 25944–25952. [Google Scholar] [CrossRef] [PubMed]
- Constantine-Paton, M.; Cline, H.T. LTP and activity-dependent synaptogenesis: The more alike they are, the more different they become. Curr. Opin. Neurobiol. 1998, 8, 139–148. [Google Scholar] [CrossRef]
- Cull-Candy, S.; Brickley, S.; Farrant, M. NMDA receptor subunits: Diversity, development and disease. Curr. Opin. Neurobiol. 2001, 11, 327–335. [Google Scholar] [CrossRef]
- Bowery, N.G.; Smart, T.G. GABA and glycine as neurotransmitters: A brief history. Br. J. Pharmacol. 2006, 147 (Suppl. S1), S109–S119. [Google Scholar] [CrossRef]
- Papouin, T.; Ladépêche, L.; Ruel, J.; Sacchi, S.; Labasque, M.; Hanini, M.; Groc, L.; Pollegioni, L.; Mothet, J.P.; Oliet, S.H. Synaptic and extrasynaptic NMDA receptors are gated by different endogenous coagonists. Cell 2012, 150, 633–646. [Google Scholar] [CrossRef]
- Tsai, G.; Yang, P.; Chung, L.C.; Lange, N.; Coyle, J.T. D-serine added to antipsychotics for the treatment of schizophrenia. Biol. Psychiatry 1998, 44, 1081–1089. [Google Scholar] [CrossRef]
- Le Douce, J.; Maugard, M.; Veran, J.; Matos, M.; Jégo, P.; Vigneron, P.A.; Faivre, E.; Toussay, X.; Vandenberghe, M.; Balbastre, Y.; et al. Impairment of Glycolysis-Derived l-Serine Production in Astrocytes Contributes to Cognitive Deficits in Alzheimer’s Disease. Cell Metab. 2020, 31, 503–517.e8. [Google Scholar] [CrossRef]
- Lee, A.; Arachchige, B.J.; Henderson, R.; Pow, D.; Reed, S.; Aylward, J.; McCombe, P.A. Elevated plasma levels of D-serine in some patients with amyotrophic lateral sclerosis. Amyotroph. Lateral. Scler. Frontotemporal. Degener. 2021, 22, 206–210. [Google Scholar] [CrossRef] [PubMed]
- Madeira, C.; Lourenco, M.V.; Vargas-Lopes, C.; Suemoto, C.K.; Brandão, C.O.; Reis, T.; Leite, R.E.; Laks, J.; Jacob-Filho, W.; Pasqualucci, C.A.; et al. d-serine levels in Alzheimer’s disease: Implications for novel biomarker development. Transl. Psychiatry 2015, 5, e561. [Google Scholar] [CrossRef] [PubMed]
- Martinez, F.J.; Pratt, G.A.; Van Nostrand, E.L.; Batra, R.; Huelga, S.C.; Kapeli, K.; Freese, P.; Chun, S.J.; Ling, K.; Gelboin-Burkhart, C.; et al. Protein-RNA Networks Regulated by Normal and ALS-Associated Mutant HNRNPA2B1 in the Nervous System. Neuron 2016, 92, 780–795. [Google Scholar] [CrossRef]
- Orzylowski, M.; Fujiwara, E.; Mousseau, D.D.; Baker, G.B. An Overview of the Involvement of D-Serine in Cognitive Impairment in Normal Aging and Dementia. Front. Psychiatry 2021, 12, 754032. [Google Scholar] [CrossRef] [PubMed]
- Piubelli, L.; Pollegioni, L.; Rabattoni, V.; Mauri, M.; Cariddi, L.P.; Versino, M.; Sacchi, S. Serum D-serine levels are altered in early phases of Alzheimer’s disease: Towards a precocious biomarker. Transl. Psychiatry 2021, 11, 77. [Google Scholar] [CrossRef]
- Watanabe, A.; Sasaki, T.; Yukami, T.; Kanki, H.; Sakaguchi, M.; Takemori, H.; Kitagawa, K.; Mochizuki, H. Serine racemase inhibition induces nitric oxide-mediated neurovascular protection during cerebral ischemia. Neuroscience 2016, 339, 139–149. [Google Scholar] [CrossRef]
- Brito, D.V.C.; Esteves, F.; Rajado, A.T.; Silva, N.; Andrade, R.; Apolónio, J.; Calado, S.; Faleiro, L.; Matos, C.; Marques, N.; et al. Assessing cognitive decline in the aging brain: Lessons from rodent and human studies. NPJ Aging 2023, 9, 23. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Chen, P.K.; Chang, Y.C.; Chuo, L.J.; Chen, Y.S.; Tsai, G.E.; Lane, H.Y. Benzoate, a D-amino acid oxidase inhibitor, for the treatment of early-phase Alzheimer disease: A randomized, double-blind, placebo-controlled trial. Biol. Psychiatry 2014, 75, 678–685. [Google Scholar] [CrossRef]
- Long, K.D.; Mastropaolo, J.; Rosse, R.B.; Manaye, K.F.; Deutsch, S.I. Modulatory effects of d-serine and sarcosine on NMDA receptor-mediated neurotransmission are apparent after stress in the genetically inbred BALB/c mouse strain. Brain Res. Bull. 2006, 69, 626–630. [Google Scholar] [CrossRef]
- Nava-Gómez, L.; Calero-Vargas, I.; Higinio-Rodríguez, F.; Vázquez-Prieto, B.; Olivares-Moreno, R.; Ortiz-Retana, J.; Aranda, P.; Hernández-Chan, N.; Rojas-Piloni, G.; Alcauter, S.; et al. Aging-Associated Cognitive Decline is Reversed by D-Serine Supplementation. eNeuro 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Kantrowitz, J.T.; Epstein, M.L.; Lee, M.; Lehrfeld, N.; Nolan, K.A.; Shope, C.; Petkova, E.; Silipo, G.; Javitt, D.C. Improvement in mismatch negativity generation during d-serine treatment in schizophrenia: Correlation with symptoms. Schizophr. Res. 2018, 191, 70–79. [Google Scholar] [CrossRef]
- Kantrowitz, J.T.; Malhotra, A.K.; Cornblatt, B.; Silipo, G.; Balla, A.; Suckow, R.F.; D’Souza, C.; Saksa, J.; Woods, S.W.; Javitt, D.C. High dose D-serine in the treatment of schizophrenia. Schizophr. Res. 2010, 121, 125–130. [Google Scholar] [CrossRef]
- Krug, A.W.; Völker, K.; Dantzler, W.H.; Silbernagl, S. Why is D-serine nephrotoxic and alpha-aminoisobutyric acid protective? Am. J. Physiol. Renal. Physiol. 2007, 293, 382–390. [Google Scholar] [CrossRef]
- Williams, R.E.; Lock, E.A. D-serine-induced nephrotoxicity: Possible interaction with tyrosine metabolism. Toxicology 2004, 201, 231–238. [Google Scholar] [CrossRef]
- Benneyworth, M.A.; Li, Y.; Basu, A.C.; Bolshakov, V.Y.; Coyle, J.T. Cell selective conditional null mutations of serine racemase demonstrate a predominate localization in cortical glutamatergic neurons. Cell. Mol. Neurobiol. 2012, 32, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Ploux, E.; Bouet, V.; Radzishevsky, I.; Wolosker, H.; Freret, T.; Billard, J.M. Serine Racemase Deletion Affects the Excitatory/Inhibitory Balance of the Hippocampal CA1 Network. Int. J. Mol. Sci. 2020, 21, 9447. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.; Salamon, R.J.; Brandt, E.B.; Paltzer, W.G.; Zhang, Z.; Britt, E.C.; Hacker, T.A.; Fan, J.; Mahmoud, A.I. Malonate Promotes Adult Cardiomyocyte Proliferation and Heart Regeneration. Circulation 2021, 143, 1973–1986. [Google Scholar] [CrossRef]
- Prag, H.A.; Pala, L.; Kula-Alwar, D.; Mulvey, J.F.; Luping, D.; Beach, T.E.; Booty, L.M.; Hall, A.R.; Logan, A.; Sauchanka, V.; et al. Ester Prodrugs of Malonate with Enhanced Intracellular Delivery Protect Against Cardiac Ischemia-Reperfusion Injury In Vivo. Cardiovasc. Drugs Ther. 2022, 36, 1–13. [Google Scholar] [CrossRef]
- Vorlová, B.; Nachtigallová, D.; Jirásková-Vaníčková, J.; Ajani, H.; Jansa, P.; Rezáč, J.; Fanfrlík, J.; Otyepka, M.; Hobza, P.; Konvalinka, J.; et al. Malonate-based inhibitors of mammalian serine racemase: Kinetic characterization and structure-based computational study. Eur. J. Med. Chem. 2015, 89, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Pan, H.; Xie, X.; Zhang, J.; Wang, Y.; Yang, G. Inhibiting Succinate Dehydrogenase by Dimethyl Malonate Alleviates Brain Damage in a Rat Model of Cardiac Arrest. Neuroscience 2018, 393, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Rani, K.; Tyagi, M.; Mazumder, M.; Singh, A.; Shanmugam, A.; Dalal, K.; Pillai, M.; Samudrala, G.; Kumar, S.; Srinivasan, A. Accelerated identification of serine racemase inhibitor from Centella asiatica. Sci. Rep. 2020, 10, 4640. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.W.; Lu, L.P.; Shih, P.C.; Hsiao, J.F.; Wu, V.; Tsai, G.E. Ultrapure and potent tannic acid (UPPTA) is a novel inhibitor of D-amino acid oxidase to improve the N-methyl-D-aspartate function of CNS disorders. Phytomedicine Plus 2023, 3, 100399. [Google Scholar] [CrossRef]
- Canosa, A.V.; Faggiano, S.; Marchetti, M.; Armao, S.; Bettati, S.; Bruno, S.; Percudani, R.; Campanini, B.; Mozzarelli, A. Glutamine 89 is a key residue in the allosteric modulation of human serine racemase activity by ATP. Sci. Rep. 2018, 8, 9016. [Google Scholar] [CrossRef] [PubMed]
- Koulouris, C.R.; Gardiner, S.E.; Harris, T.K.; Elvers, K.T.; Mark Roe, S.; Gillespie, J.A.; Ward, S.E.; Grubisha, O.; Nicholls, R.A.; Atack, J.R.; et al. Tyrosine 121 moves revealing a ligandable pocket that couples catalysis to ATP-binding in serine racemase. Commun. Biol. 2022, 5, 346. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.H.; Chang, H.T.; Hsu, L.F.; Lee, M.H.; Cheng, J.; Wu, D.C.; Lin, W.Y. In Silico and In Vitro Screening of Serine Racemase Agonist and In Vivo Efficacy on Alzheimer’s Disease Drosophila melanogaster. Pharmaceuticals 2023, 16, 280. [Google Scholar] [CrossRef]
- Lopina, O.D. Enzyme Inhibitors and Activators; InTech Open: London, UK, 2017. [Google Scholar] [CrossRef]
- Baldi, E.; Bucherelli, C. The inverted “u-shaped” dose-effect relationships in learning and memory: Modulation of arousal and consolidation. Nonlinearity Biol. Toxicol. Med. 2005, 3, 9–21. [Google Scholar] [CrossRef]
- Boondam, Y.; Songvut, P.; Tantisira, M.H.; Tapechum, S.; Tilokskulchai, K.; Pakaprot, N. Inverted U-shaped response of a standardized extract of Centella asiatica (ECa 233) on memory enhancement. Sci. Rep. 2019, 9, 8404. [Google Scholar] [CrossRef] [PubMed]
- Van Horn, M.R.; Sild, M.; Ruthazer, E.S. D-serine as a gliotransmitter and its roles in brain development and disease. Front. Cell. Neurosci. 2013, 7, 39. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, L.-P.; Chang, W.-H.; Mao, Y.-W.; Cheng, M.-C.; Zhuang, X.-Y.; Kuo, C.-S.; Lai, Y.-A.; Shih, T.-M.; Chou, T.-Y.; Tsai, G.E. The Development of a Regulator of Human Serine Racemase for N-Methyl-D-aspartate Function. Biomedicines 2024, 12, 853. https://doi.org/10.3390/biomedicines12040853
Lu L-P, Chang W-H, Mao Y-W, Cheng M-C, Zhuang X-Y, Kuo C-S, Lai Y-A, Shih T-M, Chou T-Y, Tsai GE. The Development of a Regulator of Human Serine Racemase for N-Methyl-D-aspartate Function. Biomedicines. 2024; 12(4):853. https://doi.org/10.3390/biomedicines12040853
Chicago/Turabian StyleLu, Lu-Ping, Wei-Hua Chang, Yi-Wen Mao, Min-Chi Cheng, Xiao-Yi Zhuang, Chi-Sheng Kuo, Yi-An Lai, Tsai-Miao Shih, Teh-Ying Chou, and Guochuan Emil Tsai. 2024. "The Development of a Regulator of Human Serine Racemase for N-Methyl-D-aspartate Function" Biomedicines 12, no. 4: 853. https://doi.org/10.3390/biomedicines12040853
APA StyleLu, L. -P., Chang, W. -H., Mao, Y. -W., Cheng, M. -C., Zhuang, X. -Y., Kuo, C. -S., Lai, Y. -A., Shih, T. -M., Chou, T. -Y., & Tsai, G. E. (2024). The Development of a Regulator of Human Serine Racemase for N-Methyl-D-aspartate Function. Biomedicines, 12(4), 853. https://doi.org/10.3390/biomedicines12040853