

Verapamil Attenuates the Severity of Tendinopathy by Mitigating Mitochondrial Dysfunction through the Activation of the Nrf2/HO-1 Pathway

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Animals
2.3. Materials and Reagents
2.4. Isolation and Culture of Tenocytes
2.5. Cell Experiments
2.6. Cell Viability Assay
2.7. Live/Dead Cell Staining
2.8. RNA Extraction and Quantitative Real-Time PCR Analysis
2.9. RNA Sequencing and Differentially Expressed Gene Analysis
2.10. Protein Extraction and Western Blot Analysis
2.11. Immunofluorescence
2.12. Reactive Oxygen Species Assay
2.13. Dihydroethidium Assay
2.14. Mitochondrial Membrane Potential Assay
2.15. MitoTracker Green and MitoSOX Red Assay
2.16. Histology Analysis
2.17. Immunohistochemistry
2.18. Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick-End Labeling
2.19. Statistical Analysis
3. Results
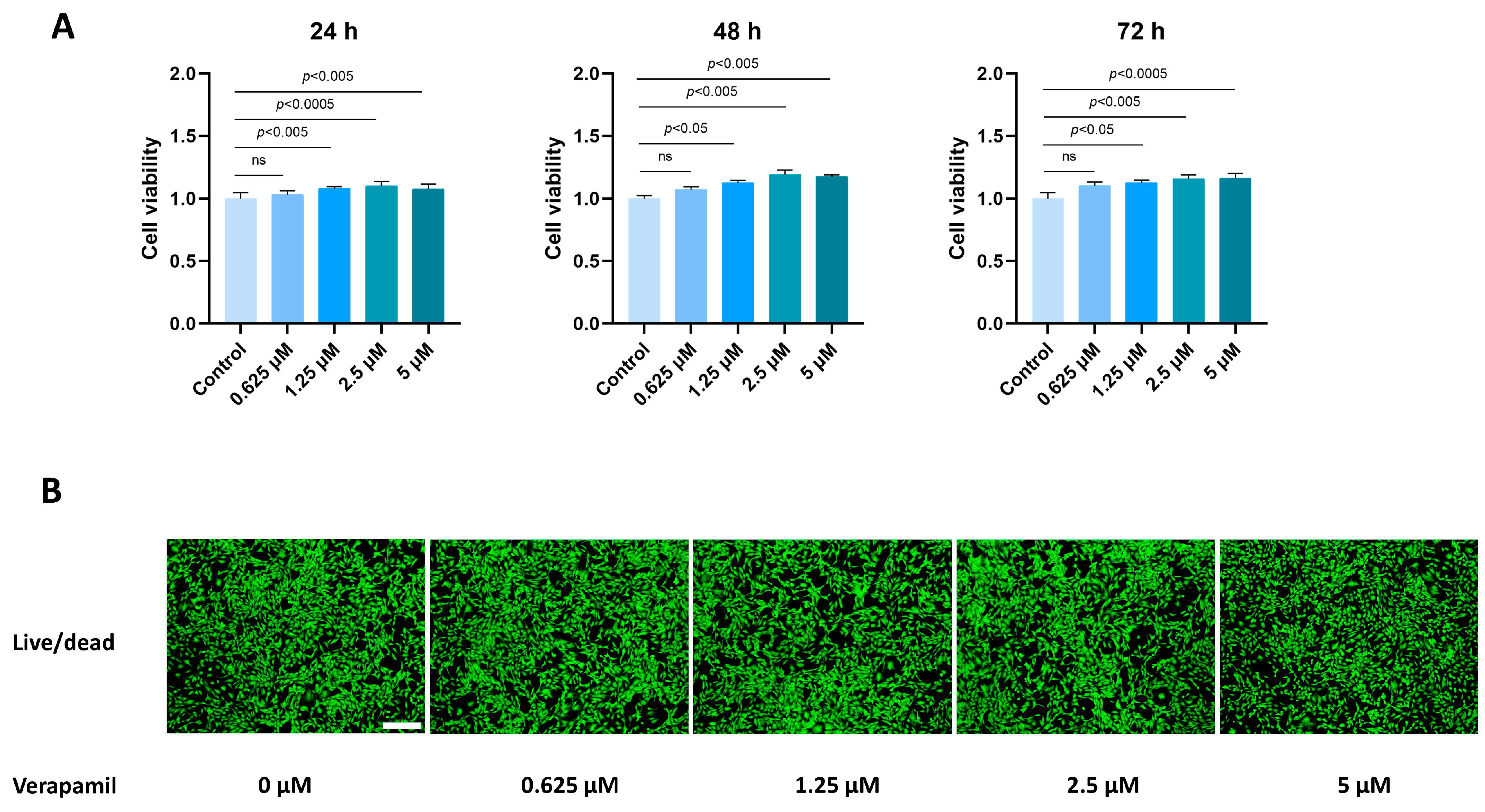
3.1. Verapamil Has Little Toxicity in Tenocytes
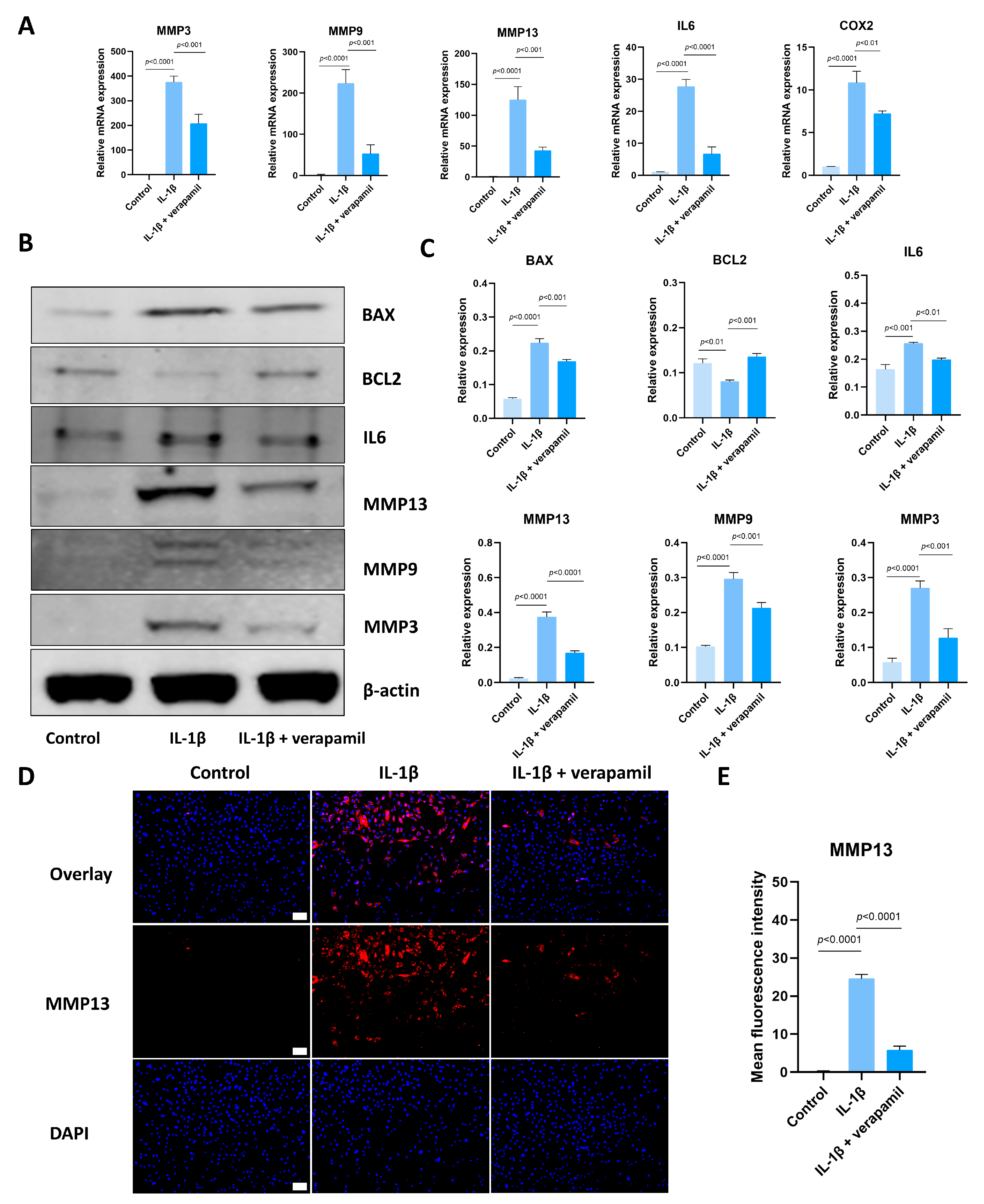
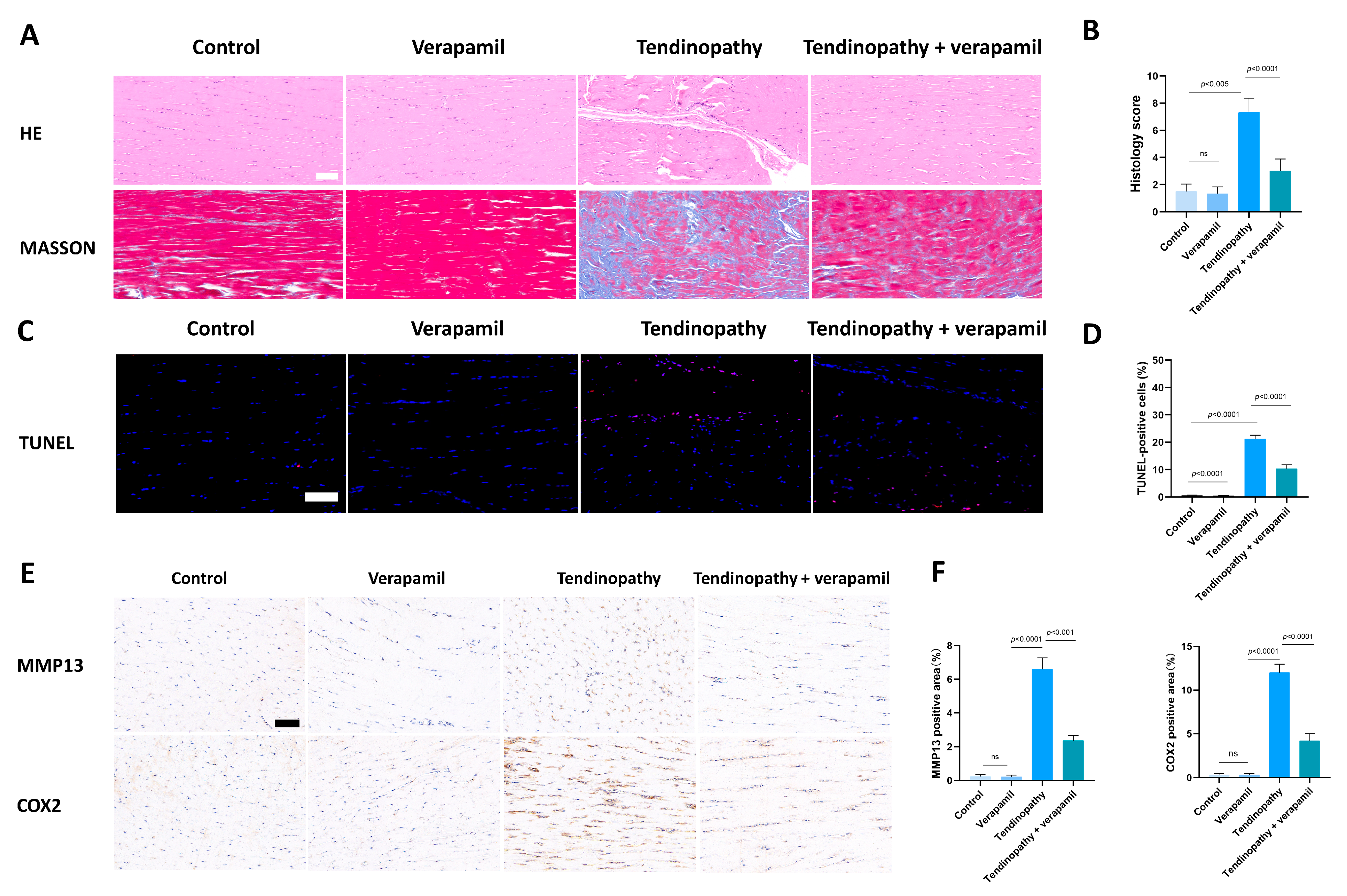
3.2. Verapamil Inhibits Extracellular Matrix Degradation, Inflammation, and Apoptosis
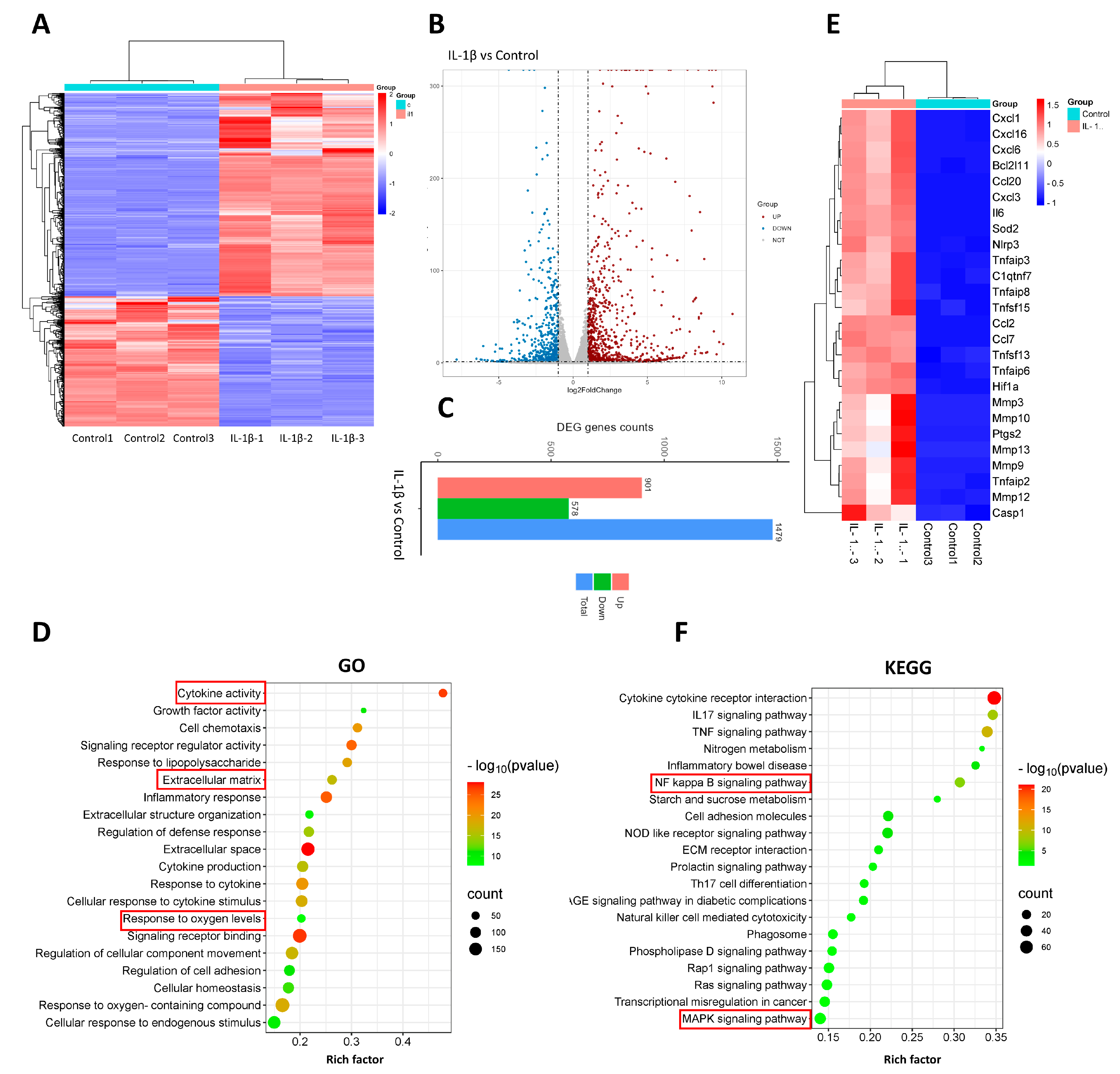
3.3. The Transcriptomic Evaluation of Tendinopathy
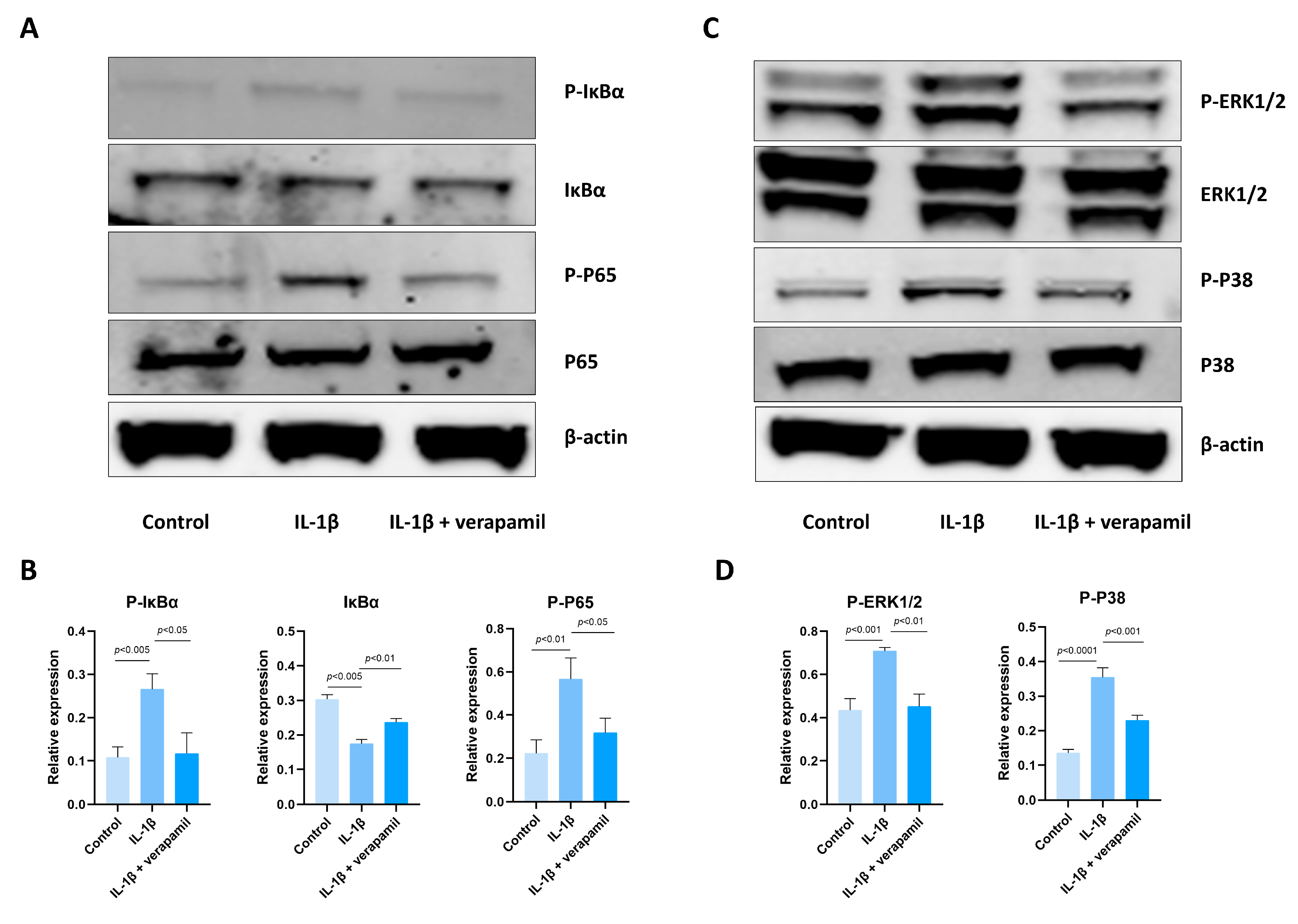
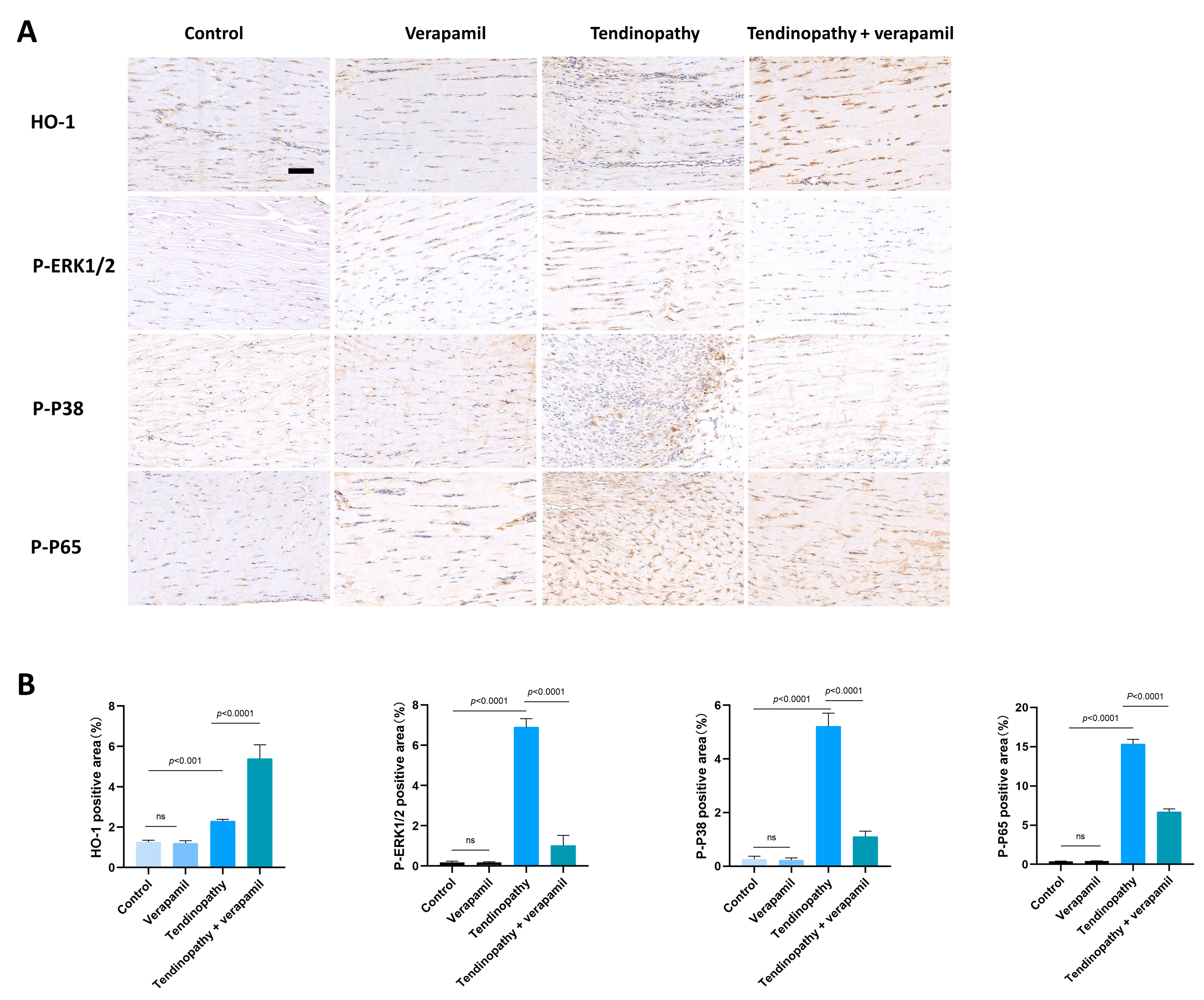
3.4. The Effect of Verapamil on the NFκb and MAPK Pathways
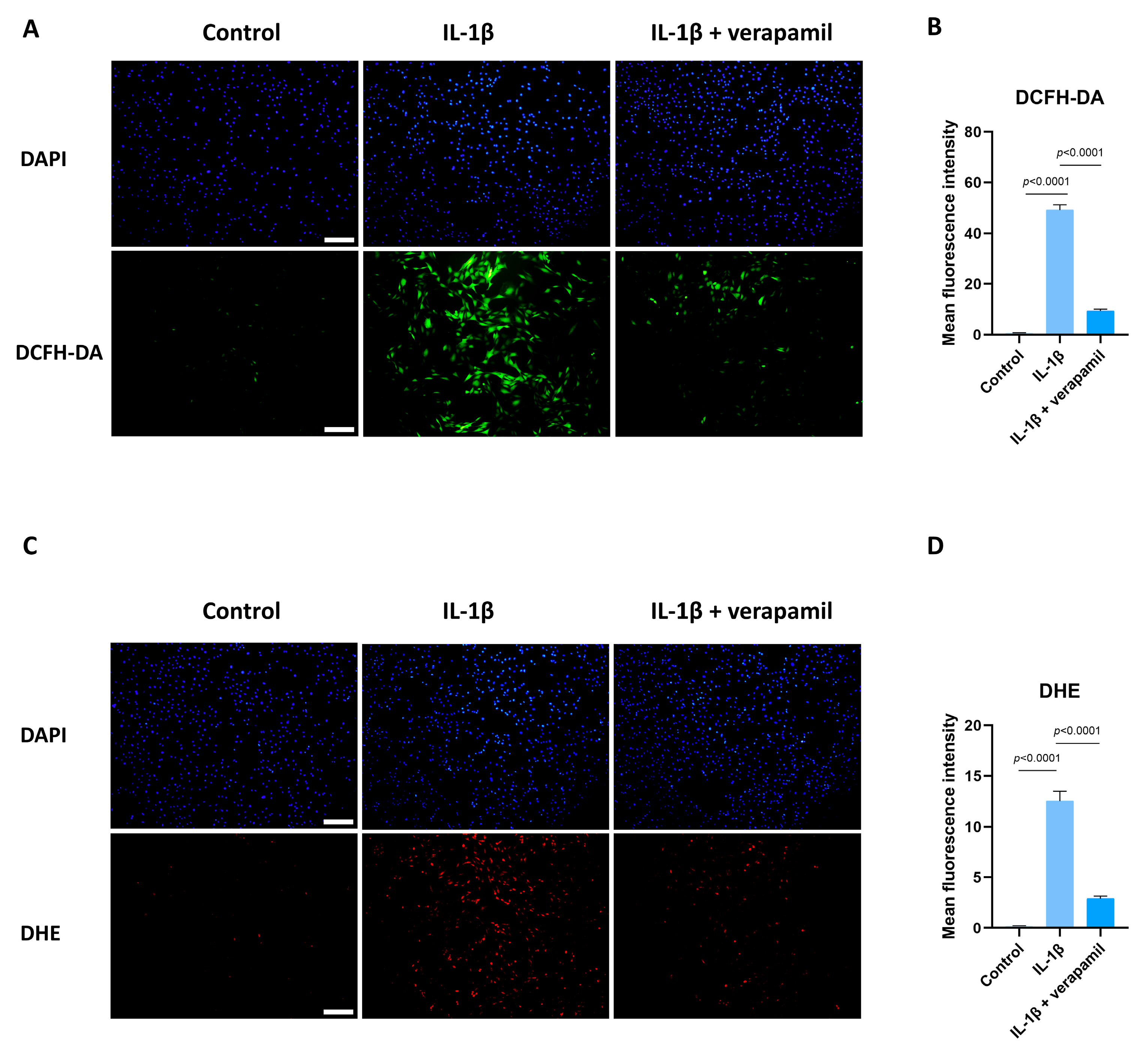
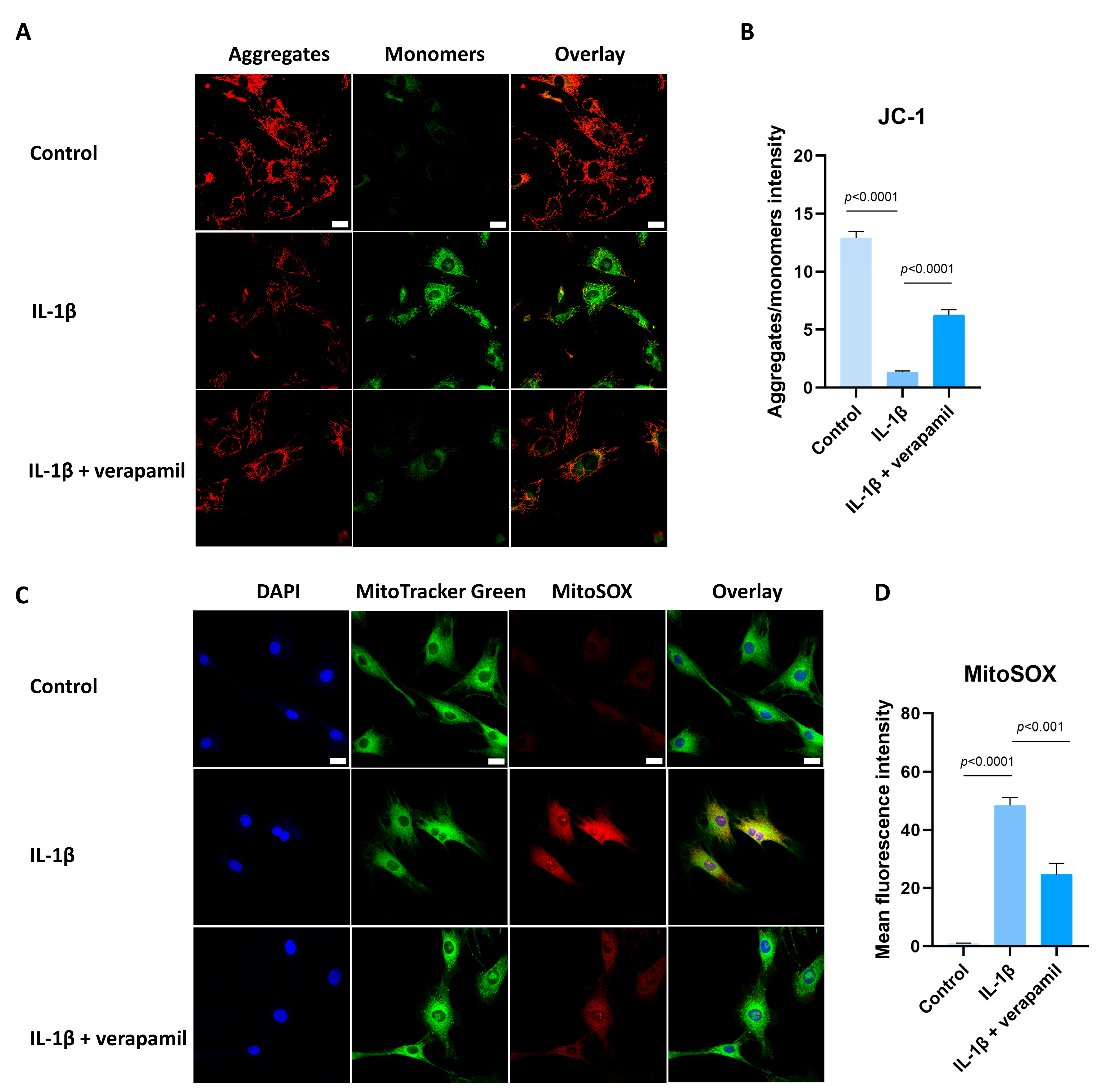
3.5. Verapamil Rescues Mitochondrial Dysfunction and Reactive Oxygen Species Production in Tendinopathy
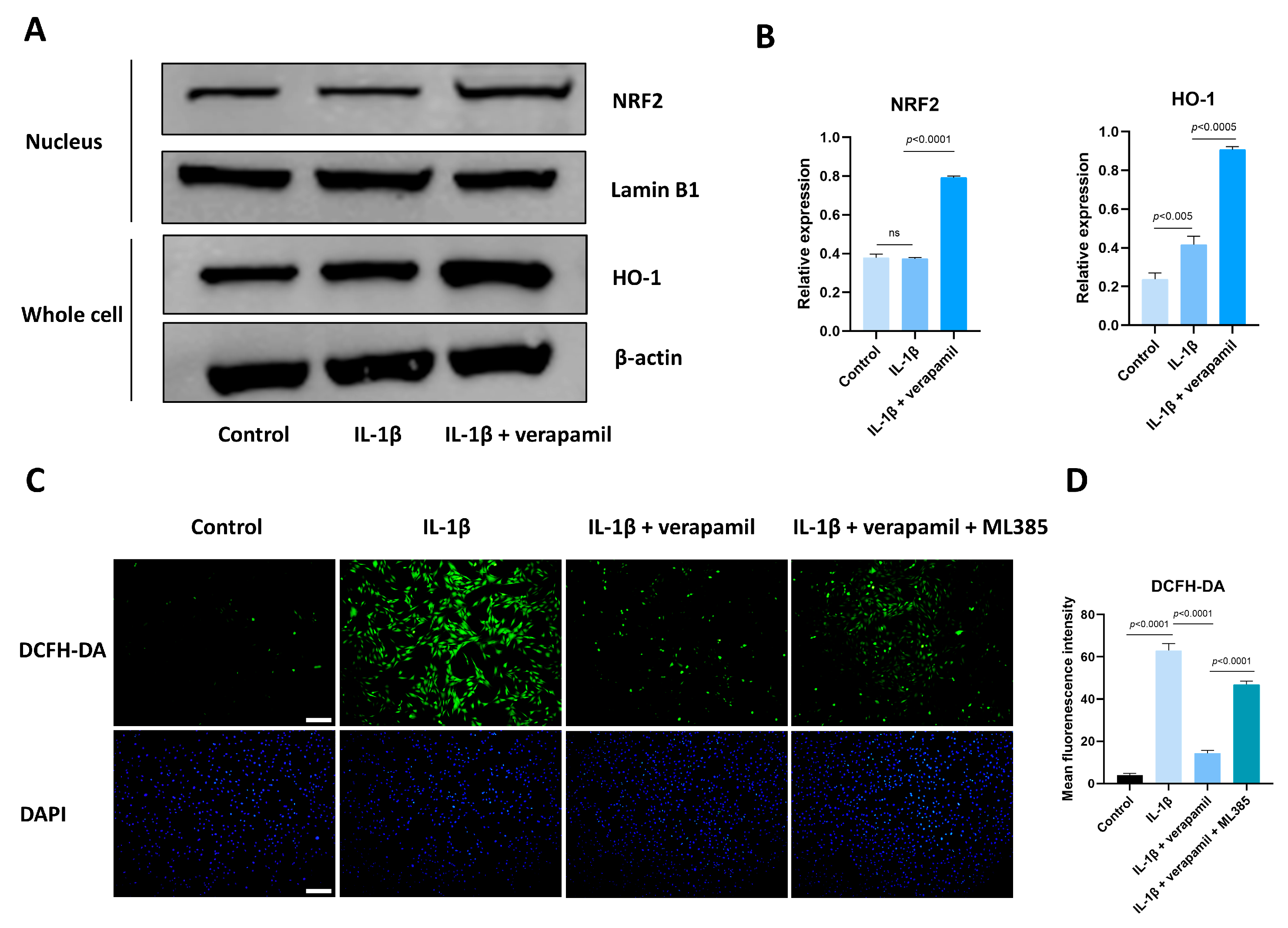
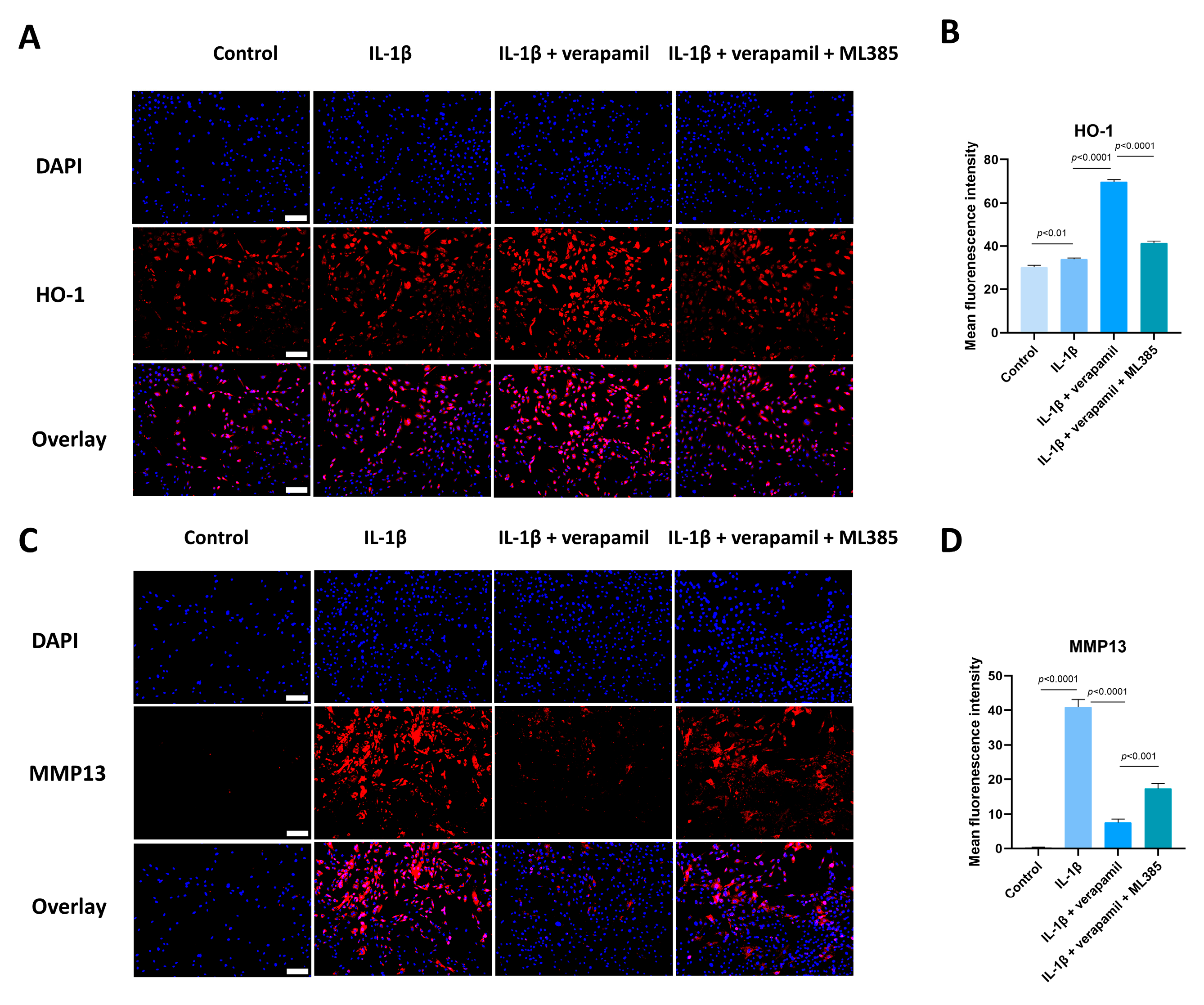
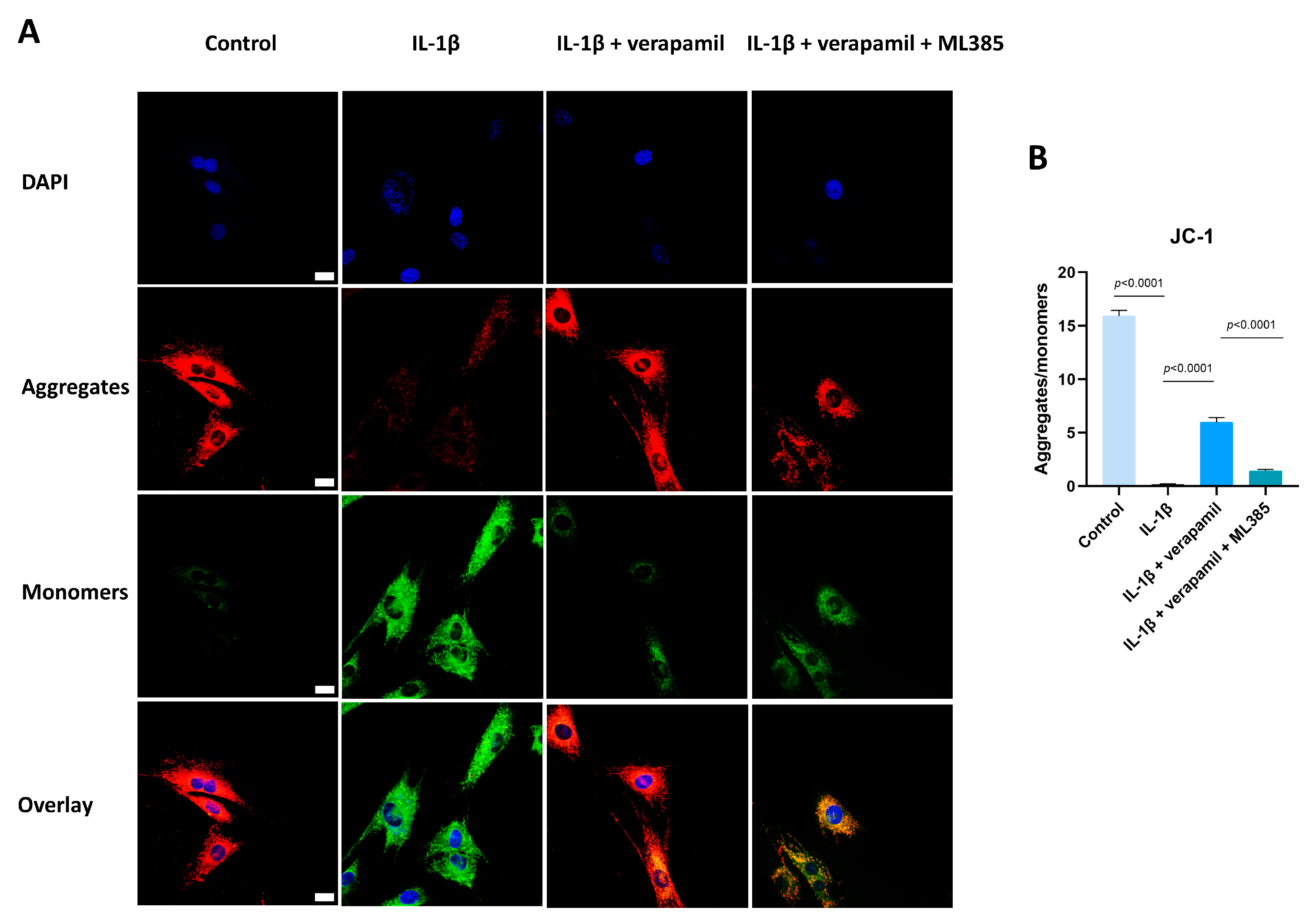
3.6. The Nrf2/HO-1 Pathway Plays an Important Role in Verapamil Protective Effects
3.7. Verapamil Rescues the Severity of Achilles Tendinopathy In Vivo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaux, J.F.; Forthomme, B.; Goff, C.L.; Crielaard, J.M.; Croisier, J.L. Current opinions on tendinopathy. J. Sports Sci. Med. 2011, 10, 238–253. [Google Scholar]
- Riel, H.; Lindstrøm, C.F.; Rathleff, M.S.; Jensen, M.B.; Olesen, J.L. Prevalence and incidence rate of lower-extremity tendinopathies in a Danish general practice: A registry-based study. BMC Musculoskelet. Disord. 2019, 20, 239. [Google Scholar] [CrossRef]
- Albers, I.S.; Zwerver, J.; Diercks, R.L.; Dekker, J.H.; Van den Akker-Scheek, I. Incidence and prevalence of lower extremity tendinopathy in a Dutch general practice population: A cross sectional study. BMC Musculoskelet. Disord. 2016, 17, 16. [Google Scholar] [CrossRef]
- Burry, H.C. Pathogenesis of some traumatic and degenerative disorders of soft tissue. Aust. N. Z. J. Med. 1978, 8 (Suppl. S1), 163–167. [Google Scholar] [CrossRef]
- Wang, J.H.; Jia, F.; Yang, G.; Yang, S.; Campbell, B.H.; Stone, D.; Woo, S.L. Cyclic mechanical stretching of human tendon fibroblasts increases the production of prostaglandin E2 and levels of cyclooxygenase expression: A novel in vitro model study. Connect. Tissue Res. 2003, 44, 128–133. [Google Scholar] [CrossRef]
- Yuan, J.; Murrell, G.A.; Wei, A.Q.; Wang, M.X. Apoptosis in rotator cuff tendonopathy. J. Orthop. Res. 2002, 20, 1372–1379. [Google Scholar] [CrossRef]
- Rees, J.D.; Stride, M.; Scott, A. Tendons—Time to revisit inflammation. Br. J. Sports Med. 2014, 48, 1553–1557. [Google Scholar] [CrossRef]
- Cook, J.L.; Rio, E.; Purdam, C.R.; Docking, S.I. Revisiting the continuum model of tendon pathology: What is its merit in clinical practice and research? Br. J. Sports Med. 2016, 50, 1187–1191. [Google Scholar] [CrossRef]
- Liu, Y.C.; Wang, H.L.; Huang, Y.Z.; Weng, Y.H.; Chen, R.S.; Tsai, W.C.; Yeh, T.H.; Lu, C.S.; Chen, Y.L.; Lin, Y.W.; et al. Alda-1, an activator of ALDH2, ameliorates Achilles tendinopathy in cellular and mouse models. Biochem. Pharmacol. 2020, 175, 113919. [Google Scholar] [CrossRef]
- Kan, T.; Ran, Z.; Sun, L.; Jiang, X.; Hou, L.; Yang, Y.; Jia, Z.; Zhang, W.; Wang, L.; Yan, M.; et al. Cell-free fat extract-loaded microneedles attenuate inflammation-induced apoptosis and mitochondrial damage in tendinopathy. Mater. Today Bio 2023, 22, 100738. [Google Scholar] [CrossRef]
- Jiao, X.; Wang, Z.; Li, Y.; Wang, T.; Xu, C.; Zhou, X.; Gan, Y. Fullerenol inhibits tendinopathy by alleviating inflammation. Front. Bioeng. Biotechnol. 2023, 11, 1171360. [Google Scholar] [CrossRef]
- Nunnari, J.; Suomalainen, A. Mitochondria: In sickness and in health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef]
- Liu, X.; Hussain, R.; Mehmood, K.; Tang, Z.; Zhang, H.; Li, Y. Mitochondrial-Endoplasmic Reticulum Communication-Mediated Oxidative Stress and Autophagy. Biomed. Res. Int. 2022, 2022, 6459585. [Google Scholar] [CrossRef]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef]
- Saha, S.; Buttari, B.; Panieri, E.; Profumo, E.; Saso, L. An Overview of Nrf2 Signaling Pathway and Its Role in Inflammation. Molecules 2020, 25, 5474. [Google Scholar] [CrossRef]
- Liu, S.; Pi, J.; Zhang, Q. Signal amplification in the KEAP1-NRF2-ARE antioxidant response pathway. Redox Biol. 2022, 54, 102389. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, Z.; Liu, L.; Xiao, Y. Natural compounds protect against the pathogenesis of osteoarthritis by mediating the NRF2/ARE signaling. Front. Pharmacol. 2023, 14, 1188215. [Google Scholar] [CrossRef]
- Saw, C.L.; Yang, A.Y.; Huang, M.T.; Liu, Y.; Lee, J.H.; Khor, T.O.; Su, Z.Y.; Shu, L.; Lu, Y.; Conney, A.H.; et al. Nrf2 null enhances UVB-induced skin inflammation and extracellular matrix damages. Cell Biosci. 2014, 4, 39. [Google Scholar] [CrossRef]
- Singh, B.N.; Ellrodt, G.; Peter, C.T. Verapamil: A review of its pharmacological properties and therapeutic use. Drugs 1978, 15, 169–197. [Google Scholar] [CrossRef]
- Baky, S.H.; Singh, B.N. Verapamil hydrochloride: Pharmacological properties and role in cardiovascular therapeutics. Pharmacotherapy 1982, 2, 328–353. [Google Scholar] [CrossRef]
- Bao, M.; Huang, W.; Zhao, Y.; Fang, X.; Zhang, Y.; Gao, F.; Huang, D.; Wang, B.; Shi, G. Verapamil Alleviates Myocardial Ischemia/Reperfusion Injury by Attenuating Oxidative Stress via Activation of SIRT1. Front. Pharmacol. 2022, 13, 822640. [Google Scholar] [CrossRef]
- Ismael, S.; Patrick, D.; Salman, M.; Parveen, A.; Stanfill, A.G.; Ishrat, T. Verapamil inhibits TXNIP-NLRP3 inflammasome activation and preserves functional recovery after intracerebral hemorrhage in mice. Neurochem. Int. 2022, 161, 105423. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, X.; Pan, B.; Du, H.; Li, B.; Yang, X.; Chen, X.; Wang, X.; Zhou, T.; Qin, A.; et al. Verapamil attenuates intervertebral disc degeneration by suppressing ROS overproduction and pyroptosis via targeting the Nrf2/TXNIP/NLRP3 axis in four-week puncture-induced rat models both in vivo and in vitro. Int. Immunopharmacol. 2023, 123, 110789. [Google Scholar] [CrossRef]
- Lee, J.M.; Hwang, J.W.; Kim, M.J.; Jung, S.Y.; Kim, K.S.; Ahn, E.H.; Min, K.; Choi, Y.S. Mitochondrial Transplantation Modulates Inflammation and Apoptosis, Alleviating Tendinopathy Both In Vivo and In Vitro. Antioxidants 2021, 10, 696. [Google Scholar] [CrossRef]
- Jiao, X.; Zhang, Y.; Li, W.; Zhou, X.; Chu, W.; Li, Y.; Wang, Z.; Sun, X.; Xu, C.; Gan, Y. HIF-1α inhibition attenuates severity of Achilles tendinopathy by blocking NF-κB and MAPK pathways. Int. Immunopharmacol. 2022, 106, 108543. [Google Scholar] [CrossRef]
- Chen, J.; Yu, Q.; Wu, B.; Lin, Z.; Pavlos, N.J.; Xu, J.; Ouyang, H.; Wang, A.; Zheng, M.H. Autologous tenocyte therapy for experimental Achilles tendinopathy in a rabbit model. Tissue Eng. Part. A 2011, 17, 2037–2048. [Google Scholar] [CrossRef]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef]
- Ren, Q.; Guo, F.; Tao, S.; Huang, R.; Ma, L.; Fu, P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-κB p65 and MAPK signaling pathways in septic AKI mice. Biomed. Pharmacother. 2020, 122, 109772. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- He, R.; Cui, M.; Lin, H.; Zhao, L.; Wang, J.; Chen, S.; Shao, Z. Melatonin resists oxidative stress-induced apoptosis in nucleus pulposus cells. Life Sci. 2018, 199, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Millar, N.L.; Murrell, G.A.; McInnes, I.B. Inflammatory mechanisms in tendinopathy—Towards translation. Nat. Rev. Rheumatol. 2017, 13, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Pufe, T.; Petersen, W.J.; Mentlein, R.; Tillmann, B.N. The role of vasculature and angiogenesis for the pathogenesis of degenerative tendons disease. Scand. J. Med. Sci. Sports 2005, 15, 211–222. [Google Scholar] [CrossRef]
- Fredberg, U.; Stengaard-Pedersen, K. Chronic tendinopathy tissue pathology, pain mechanisms, and etiology with a special focus on inflammation. Scand. J. Med. Sci. Sports 2008, 18, 3–15. [Google Scholar] [CrossRef]
- Fu, S.C.; Rolf, C.; Cheuk, Y.C.; Lui, P.P.; Chan, K.M. Deciphering the pathogenesis of tendinopathy: A three-stages process. Sports Med. Arthrosc. Rehabil. Ther. Technol. 2010, 2, 30. [Google Scholar] [CrossRef] [PubMed]
- Legerlotz, K.; Jones, E.R.; Screen, H.R.; Riley, G.P. Increased expression of IL-6 family members in tendon pathology. Rheumatology 2012, 51, 1161–1165. [Google Scholar] [CrossRef]
- Millar, N.L.; Silbernagel, K.G.; Thorborg, K.; Kirwan, P.D.; Galatz, L.M.; Abrams, G.D.; Murrell, G.A.C.; McInnes, I.B.; Rodeo, S.A. Tendinopathy. Nat. Rev. Dis. Primers 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Zhang, Y.; Chen, J.; Hu, Y.; Xu, Y. Verapamil Ameliorates Hepatic Metaflammation by Inhibiting Thioredoxin-Interacting Protein/NLRP3 Pathways. Front. Endocrinol. 2018, 9, 640. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Li, S.; Li, L. Verapamil inhibits early acute liver failure through suppressing the NLRP3 inflammasome pathway. J. Cell Mol. Med. 2021, 25, 5963–5975. [Google Scholar] [CrossRef]
- Ismael, S.; Nasoohi, S.; Yoo, A.; Mirzahosseini, G.; Ahmed, H.A.; Ishrat, T. Verapamil as an Adjunct Therapy to Reduce tPA Toxicity in Hyperglycemic Stroke: Implication of TXNIP/NLRP3 Inflammasome. Mol. Neurobiol. 2021, 58, 3792–3804. [Google Scholar] [CrossRef]
- Abraham, A.C.; Shah, S.A.; Golman, M.; Song, L.; Li, X.; Kurtaliaj, I.; Akbar, M.; Millar, N.L.; Abu-Amer, Y.; Galatz, L.M.; et al. Targeting the NF-κB signaling pathway in chronic tendon disease. Sci. Transl. Med. 2019, 11, aav4319. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP kinase signalling pathways in cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef]
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-kB in development and progression of human cancer. Virchows Arch. 2005, 446, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Lai, J.L.; Liu, Y.H.; Liu, C.; Qi, M.P.; Liu, R.N.; Zhu, X.F.; Zhou, Q.G.; Chen, Y.Y.; Guo, A.Z.; Hu, C.M. Indirubin Inhibits LPS-Induced Inflammation via TLR4 Abrogation Mediated by the NF-kB and MAPK Signaling Pathways. Inflammation 2017, 40, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yuan, L.; Liu, Y.; Wang, R.; Zhang, Y.; Yang, Y.; Wei, H.; Ma, J. Integrated Cascade Nanozyme Remodels Chondrocyte Inflammatory Microenvironment in Temporomandibular Joint Osteoarthritis via Inhibiting ROS-NF-κB and MAPK Pathways. Adv. Healthc. Mater. 2023, 12, e2203195. [Google Scholar] [CrossRef] [PubMed]
- Longo, U.G.; Oliva, F.; Denaro, V.; Maffulli, N. Oxygen species and overuse tendinopathy in athletes. Disabil. Rehabil. 2008, 30, 1563–1571. [Google Scholar] [CrossRef]
- Yuan, T.; Qian, H.; Yu, X.; Meng, J.; Lai, C.T.; Jiang, H.; Zhao, J.N.; Bao, N.R. Proteomic analysis reveals rotator cuff injury caused by oxidative stress. Ther. Adv. Chronic Dis. 2021, 12, 2040622320987057. [Google Scholar] [CrossRef]
- Li, X.; Su, Z.; Shen, K.; Wang, Q.; Xu, C.; Wang, F.; Zhang, Y.; Jiang, D. Eugenol-Preconditioned Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Antioxidant Capacity of Tendon Stem Cells In Vitro and In Vivo. Oxid. Med. Cell Longev. 2022, 2022, 3945195. [Google Scholar] [CrossRef]
- Itoigawa, Y.; Yoshida, K.; Nojiri, H.; Morikawa, D.; Kawasaki, T.; Wada, T.; Koga, A.; Maruyama, Y.; Ishijima, M. Association of Recurrent Tear After Arthroscopic Rotator Cuff Repair and Superoxide-Induced Oxidative Stress. Am. J. Sports Med. 2021, 49, 2048–2055. [Google Scholar] [CrossRef]
- Sinha, K.; Das, J.; Pal, P.B.; Sil, P.C. Oxidative stress: The mitochondria-dependent and mitochondria-independent pathways of apoptosis. Arch. Toxicol. 2013, 87, 1157–1180. [Google Scholar] [CrossRef]
- Pang, Z.; Jiang, Z.; Zhu, R.; Song, C.; Tang, H.; Cao, L.; Guo, C. Bardoxolone-Methyl Prevents Oxidative Stress-Mediated Apoptosis and Extracellular Matrix Degradation in vitro and Alleviates Osteoarthritis in vivo. Drug Des. Dev. Ther. 2021, 15, 3735–3747. [Google Scholar] [CrossRef]
- Tang, Y.; Zhou, X.; Cao, T.; Chen, E.; Li, Y.; Lei, W.; Hu, Y.; He, B.; Liu, S. Endoplasmic Reticulum Stress and Oxidative Stress in Inflammatory Diseases. DNA Cell Biol. 2022, 41, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Scialò, F.; Fernández-Ayala, D.J.; Sanz, A. Role of Mitochondrial Reverse Electron Transport in ROS Signaling: Potential Roles in Health and Disease. Front. Physiol. 2017, 8, 428. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Zhang, J.; Li, M.; Zhao, Y.; Guo, Y.; Xu, J.; Liu, R.; Li, Z.; Ren, D.; Liu, P. Liquiritigenin Confers Liver Protection by Enhancing NRF2 Signaling through Both Canonical and Non-canonical Signaling Pathways. J. Med. Chem. 2023, 66, 11324–11334. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, M.; Kurokawa, H.; Waguri, S.; Taguchi, K.; Kobayashi, A.; Ichimura, Y.; Sou, Y.S.; Ueno, I.; Sakamoto, A.; Tong, K.I.; et al. The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nat. Cell Biol. 2010, 12, 213–223. [Google Scholar] [CrossRef]
- Hast, B.E.; Goldfarb, D.; Mulvaney, K.M.; Hast, M.A.; Siesser, P.F.; Yan, F.; Hayes, D.N.; Major, M.B. Proteomic analysis of ubiquitin ligase KEAP1 reveals associated proteins that inhibit NRF2 ubiquitination. Cancer Res. 2013, 73, 2199–2210. [Google Scholar] [CrossRef] [PubMed]
- Gorrini, C.; Baniasadi, P.S.; Harris, I.S.; Silvester, J.; Inoue, S.; Snow, B.; Joshi, P.A.; Wakeham, A.; Molyneux, S.D.; Martin, B.; et al. BRCA1 interacts with Nrf2 to regulate antioxidant signaling and cell survival. J. Exp. Med. 2013, 210, 1529–1544. [Google Scholar] [CrossRef] [PubMed]
- Silva-Islas, C.A.; Maldonado, P.D. Canonical and non-canonical mechanisms of Nrf2 activation. Pharmacol. Res. 2018, 134, 92–99. [Google Scholar] [CrossRef]
- Li, Y.; Ma, K.; Han, Z.; Chi, M.; Sai, X.; Zhu, P.; Ding, Z.; Song, L.; Liu, C. Immunomodulatory Effects of Heme Oxygenase-1 in Kidney Disease. Front. Med. 2021, 8, 708453. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Hu, X.C.; Zhang, G.Z.; Liu, M.Q.; Chen, H.W.; Kang, X.W. Role of Nrf2 and HO-1 in intervertebral disc degeneration. Connect. Tissue Res. 2022, 63, 559–576. [Google Scholar] [CrossRef] [PubMed]
- Severance, S.; Hamza, I. Trafficking of heme and porphyrins in metazoa. Chem. Rev. 2009, 109, 4596–4616. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhong, H.; Wei, J.; Lin, S.; Zong, Z.; Gong, F.; Huang, X.; Sun, J.; Li, P.; Lin, H.; et al. Inhibition of Nrf2/HO-1 signaling leads to increased activation of the NLRP3 inflammasome in osteoarthritis. Arthritis Res. Ther. 2019, 21, 300. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, L.; Chen, J.; Li, Q.; Huo, L.; Wang, Y.; Wang, H.; Du, J. Pharmacological Modulation of Nrf2/HO-1 Signaling Pathway as a Therapeutic Target of Parkinson's Disease. Front. Pharmacol. 2021, 12, 757161. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.; Wang, H.; Jiao, D.; Geng, Y.; Wu, Q.; Yan, H.; Li, C. Curcumin Restrains Oxidative Stress of After Intracerebral Hemorrhage in Rat by Activating the Nrf2/HO-1 Pathway. Front. Pharmacol. 2022, 13, 889226. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Li, H.; Hu, F.; Luo, P. Stevioside attenuates osteoarthritis via regulating Nrf2/HO-1/NF-κB pathway. J. Orthop. Transl. 2023, 38, 190–202. [Google Scholar] [CrossRef] [PubMed]
- McTavish, D.; Sorkin, E.M. Verapamil. An updated review of its pharmacodynamic and pharmacokinetic properties, and therapeutic use in hypertension. Drugs 1989, 38, 19–76. [Google Scholar] [CrossRef]
- Ovalle, F.; Grimes, T.; Xu, G.; Patel, A.J.; Grayson, T.B.; Thielen, L.A.; Li, P.; Shalev, A. Verapamil and beta cell function in adults with recent-onset type 1 diabetes. Nat. Med. 2018, 24, 1108–1112. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| β-actin | GTC CAC CCG CGA GTA CAA C | GGA TGC CTC TCT TGC TCT GG |
| MMP3 | GCT GTC TTT GAA GCA TTT GGG TT | CCT CCA TGA AAA GAC TCA GAG GA |
| MMP9 | TCC AGC ATC TGT ATG GTC GTG | GCA GTG GGA CAC ATA GTG GG |
| MMP13 | CAA GCA GCT CCA AAG GCT AC | TGG CTT TTG CCA GTG TAG GT |
| IL6 | CCA GTT GCC TTC TTG GGA CT | TGC CAT TGC ACA ACT CTT TTC |
| COX2 | CTC AGC CAT GCA GCA AAT CC | GGG TGG GCT TCA GCA GTA AT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Dong, Z.; Li, Y.; Jiao, X.; Liu, Y.; Chang, H.; Gan, Y. Verapamil Attenuates the Severity of Tendinopathy by Mitigating Mitochondrial Dysfunction through the Activation of the Nrf2/HO-1 Pathway. Biomedicines 2024, 12, 904. https://doi.org/10.3390/biomedicines12040904
Wang Z, Dong Z, Li Y, Jiao X, Liu Y, Chang H, Gan Y. Verapamil Attenuates the Severity of Tendinopathy by Mitigating Mitochondrial Dysfunction through the Activation of the Nrf2/HO-1 Pathway. Biomedicines. 2024; 12(4):904. https://doi.org/10.3390/biomedicines12040904
Chicago/Turabian StyleWang, Zengguang, Zhenglin Dong, Yiming Li, Xin Jiao, Yihao Liu, Hanwen Chang, and Yaokai Gan. 2024. "Verapamil Attenuates the Severity of Tendinopathy by Mitigating Mitochondrial Dysfunction through the Activation of the Nrf2/HO-1 Pathway" Biomedicines 12, no. 4: 904. https://doi.org/10.3390/biomedicines12040904
APA StyleWang, Z., Dong, Z., Li, Y., Jiao, X., Liu, Y., Chang, H., & Gan, Y. (2024). Verapamil Attenuates the Severity of Tendinopathy by Mitigating Mitochondrial Dysfunction through the Activation of the Nrf2/HO-1 Pathway. Biomedicines, 12(4), 904. https://doi.org/10.3390/biomedicines12040904