Salmonella-Based Targeted Cancer Therapy: Updates on A Promising and Innovative Tumor Immunotherapeutic Strategy


Abstract
:
1. Introduction
2. Salmonella Efficiently Targets and Invades Tumor
3. Modified Salmonella Bypasses Antibacterial Immune Response
4. Salmonella Promotes Activation of Antitumor Immunity
5. Salmonella Mediates Tumor Cell Self-Destruction
6. Salmonella Diminishes Tumor Metastasis
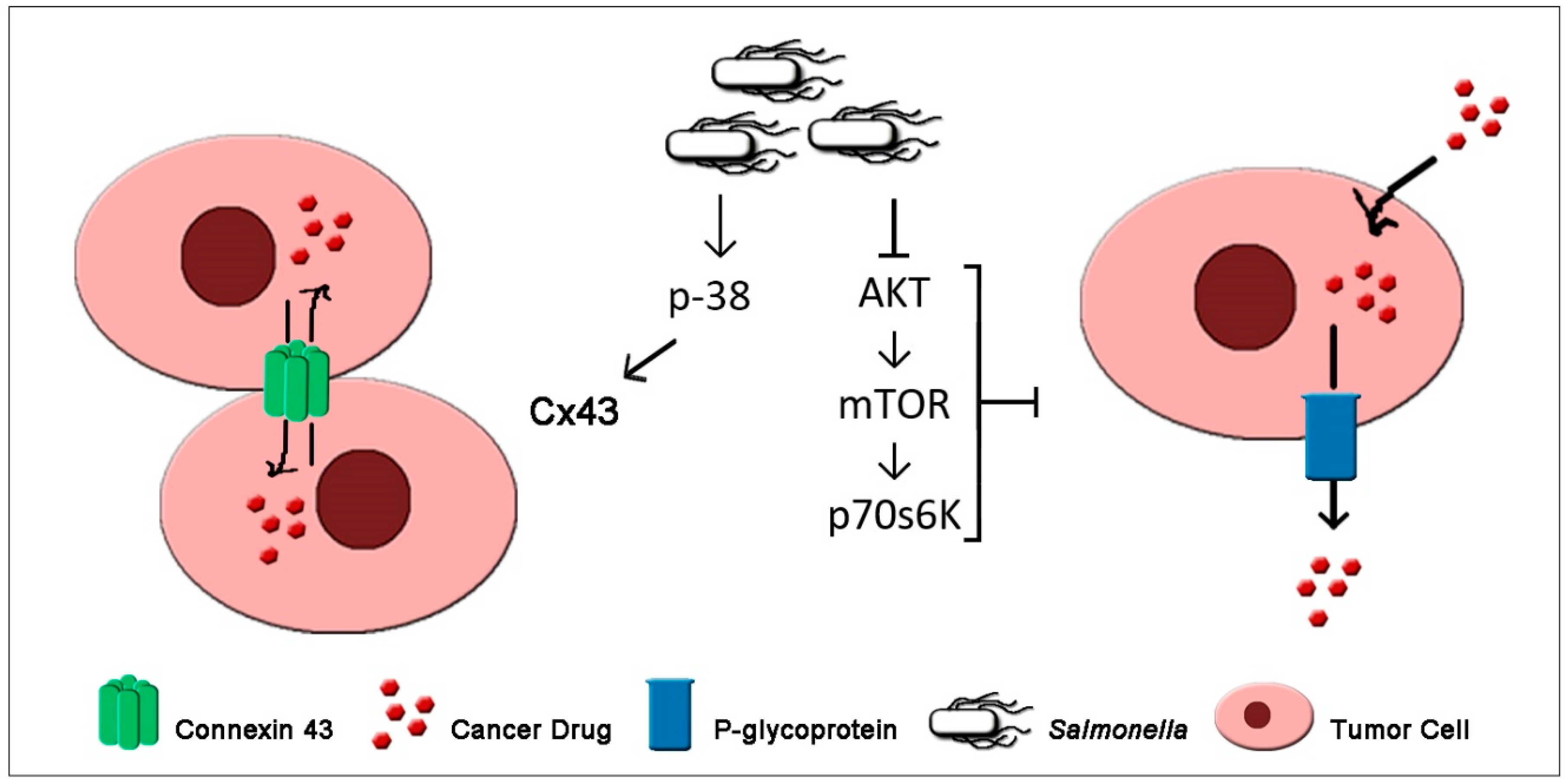
7. Salmonella Enhances Chemosensitivity of Tumor
8. Combination Therapy with Salmonella Further Improves Tumor-Regression
9. Clinical Trials, Challenges and Future Perspectives
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AKT | protein kinase B |
| asd | aspartate-semialdehyde dehydrogenase gene |
| B-NHL | B-cell non-Hodgkin lymphoma |
| CHOP | cyclophosphamide, doxorubicin, vincristine, and prednisone |
| Cx43 | connexin-43 |
| ECM | extracellular matrix |
| EMT | epithelium to mesenchymal transition |
| GJIC | gap junction intercellular communication |
| HIF-1 | hypoxia inducible factor-1 |
| HK | heat killed |
| IDO | indoleamine 2, 3-dioxygenase 1 |
| IFN-γ | interferon-γ |
| IL | interleukins |
| iNOS | inducible nitric oxide synthase |
| LPS | lipopolysaccharide |
| MMP | matrix metalloproteinase |
| mTOR | mammalian target of rapamycin |
| OmpA | outer membrane protein A |
| p70S6K | ribosomal protein S6 kinase beta-1 |
| P-gp | P glycoprotein |
| PAH | poly(allylamine hydrochloride) |
| pDA | polydopamine |
| PMN | polymorphonuclear neutrophils |
| RGD | arginine-glycine-aspartate |
| shRNA | short hairpin ribonucleic acid |
| TGF-β | transforming growth factor-β |
| TLR | toll like receptor |
| TNF-α | tumor necrosis factor-α |
| TRAIL | tumor necrosis factor-related apoptosis-inducing ligand |
| TSP-1 | thrombospondin-1 |
| VEGF | vascular endothelial growth factor. |
References
- Nguyen, V.H.; Min, J.J. Salmonella-mediated cancer therapy: Roles and potentials. Nucl. Med. Mol. Imaging 2017, 51, 118–126. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Dagogo-Jack, I.; Shaw, A.T. Tumour heterogeneity and resistance to cancer therapies. Nat. Rev. Clin. Oncol. 2018, 15, 81–94. [Google Scholar] [CrossRef]
- Brown, J.M.; Wilson, W.R. Exploiting tumour hypoxia in cancer treatment. Nat. Rev. Cancer 2004, 4, 437–447. [Google Scholar] [CrossRef]
- Lee, C.H. Engineering bacteria toward tumor targeting for cancer treatment: Current state and perspectives. Appl. Microbiol. Biotechnol. 2012, 93, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Kasinskas, R.W.; Forbes, N.S. Salmonella typhimurium specifically chemotax and proliferate in heterogeneous tumor tissue in vitro. Biotechnol. Bioeng. 2006, 94, 710–721. [Google Scholar] [CrossRef]
- Chang, W.W.; Lee, C.H. Salmonella as an innovative therapeutic antitumor agent. Int. J. Mol. Sci. 2014, 15, 14546–14554. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Wu, C.L.; Chen, S.H.; Shiau, A.L. Humoral immune responses inhibit the antitumor activities mediated by Salmonella enterica serovar choleraesuis. J. Immunother. 2009, 32, 376–388. [Google Scholar] [CrossRef]
- Needham, B.D.; Carroll, S.M.; Giles, D.K.; Georgiou, G.; Whiteley, M.; Trent, M.S. Modulating the innate immune response by combinatorial engineering of endotoxin. Proc. Natl. Acad. Sci. USA 2013, 110, 1469–1499. [Google Scholar] [CrossRef]
- Lee, C.H.; Lin, Y.H.; Hsieh, J.L.; Chen, M.C.; Kuo, W.L. A polymer coating applied to Salmonella prevents the binding of Salmonella-specific antibodies. Int. J. Cancer 2013, 132, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Wei, M.Q.; Ellem, K.A.O.; Dunn, P.; West, M.J.; Bai, C.X.; Vogelstein, B. Facultative or obligate anaerobic bacteria have the potential for multimodality therapy of solid tumours. Eur. J. Cancer 2007, 43, 490–496. [Google Scholar] [CrossRef]
- St Jean, A.T.; Zhang, M.; Forbes, N.S. Bacterial therapies: Completing the cancer treatment toolbox. Curr. Opin. Biotechnol. 2008, 19, 511–517. [Google Scholar] [CrossRef]
- Yu, B.; Yang, M.; Shi, L.; Yao, Y.; Jiang, Q.; Li, X.; Tang, L.H.; Zheng, B.J.; Yuen, K.Y.; Smith, D.K.; et al. Explicit hypoxia targeting with tumor suppression by creating an “obligate” anaerobic Salmonella typhimurium strain. Sci. Rep. 2012, 2, 436. [Google Scholar] [CrossRef]
- Anderson, C.J.; Clark, D.E.; Adli, M.; Kendall, M.M. Ethanolamine signaling promotes Salmonella niche recognition and adaptation during infection. PLoS Pathog. 2015, 11, e1005278. [Google Scholar]
- Silva-Valenzuela, C.A.; Desai, P.T.; Molina-Quiroz, R.C.; Pezoa, D.; Zhang, Y.; Porwollik, S.; Zhao, M.; Hoffman, R.M.; Contreras, I.; Santiviago, C.A.; et al. Solid tumors provide niche-specific conditions that lead to preferential growth of Salmonella. Oncotarget 2016, 7, 35169–35180. [Google Scholar] [CrossRef] [PubMed]
- Toley, B.J.; Forbes, N.S. Motility is critical for effective distribution and accumulation of bacteria in tumor tissue. Integr. Biol. 2012, 4, 165–176. [Google Scholar] [CrossRef]
- Ganai, S.; Arenas, R.B.; Sauer, J.P.; Bentley, B.; Forbes, N.S. In tumors Salmonella migrate away from vasculature toward the transition zone and induce apoptosis. Cancer Gene Ther. 2011, 18, 457–466. [Google Scholar] [CrossRef]
- Broadway, K.M.; Denson, E.A.P.; Jensen, R.V.; Scharf, B.E. Rescuing chemotaxis of the anticancer agent Salmonella enterica serovar Typhimurium VNP20009. J. Biotechnol. 2015, 211, 117–120. [Google Scholar] [CrossRef]
- Clairmont, C.; Lee, K.C.; Pike, J.; Ittensohn, M.; Low, K.B.; Pawelek, J.; Bermudes, D.; Brecher, S.M.; Margitich, D.; Turnier, J.; et al. Biodistribution and Genetic Stability of the Novel Antitumor Agent VNP20009, a Genetically Modified Strain of Salmonella Typhimurium. J. Infect. Dis. 2000, 181, 1996–2002. [Google Scholar] [CrossRef]
- Broadway, K.M.; Suh, S.; Behkam, B.; Scharf, B.E. Optimizing the restored chemotactic behavior of anticancer agent Salmonella enterica serovar Typhimurium VNP20009. J. Biotechnol. 2017, 251, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Coutermarsh-Ott, S.L.; Broadway, K.M.; Scharf, B.E.; Allen, I.C. Effect of Salmonella enterica serovar Typhimurium VNP20009 and VNP20009 with restored chemotaxis on 4T1 mouse mammary carcinoma progression. Oncotarget 2017, 8, 33601–33613. [Google Scholar] [CrossRef]
- Arrach, N.; Cheng, P.; Zhao, M.; Santiviago, C.A.; Hoffman, R.M.; McClelland, M. High-throughput screening for Salmonella avirulent mutants that retain targeting of solid tumors. Cancer Res. 2010, 70, 2165–2170. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Forbes, N.S. Trg-deficient Salmonella colonize quiescent tumor regions by exclusively penetrating or proliferating. J. Control Release 2015, 199, 180–189. [Google Scholar] [CrossRef]
- Palffy, R.; Gardlik, R.; Hodosy, J.; Behuliak, M.; Resko, P.; Radvansky, J.; Celec, P. Bacteria in gene therapy: Bactofection versus alternative gene therapy. Gene Ther. 2006, 13, 101–105. [Google Scholar] [CrossRef]
- Felgner, S.; Kocijancic, D.; Frahm, M.; Heise, U.; Rohde, M.; Zimmermann, K.; Falk, C.; Erhardt, M.; Weiss, S. Engineered Salmonella enterica serovar Typhimurium overcomes limitationsof anti-bacterial immunity in bacteria-mediated tumor therapy. Oncoimmunology 2018, 7, e1382791. [Google Scholar] [CrossRef]
- Vinay, D.S.; Ryan, E.P.; Pawelec, G.; Talib, W.H.; Stagg, J.; Elkord, E.; Lichtor, T.; Decker, W.K.; Whelan, R.L.; Kumarai, H.M.C.S.; et al. Immune evasion in cancer: Mechanistic basis and therapeutic strategies. Semin. Cancer Biol. 2015, 35, S185–S198. [Google Scholar] [CrossRef] [Green Version]
- Spranger, S.; Gajewski, T.F. Mechanisms of tumor cell–intrinsic immune evasion. Ann. Rev. Cancer Biol. 2018, 2, 213–228. [Google Scholar] [CrossRef]
- Kim, J.E.; Phan, T.X.; Nguyen, V.H.; Dinh-Vu, H.V.; Zheng, J.H.; Yun, M.; Park, S.G.; Hong, Y.; Choy, H.E.; Szardenings, M.; et al. Salmonella typhimurium suppresses tumor growth via the pro-inflammatory cytokine interleukin-1β. Theranostics 2015, 5, 1328–1342. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.X.; Nguyen, V.H.; Duong, M.T.; Hong, Y.; Choy, H.E.; Min, J.J. Activation of inflammasome by attenuated Salmonella typhimurium in bacteria-mediated cancer therapy. Microbiol. Immunol. 2015, 59, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.W.; Kullas, A.L.; Mena, P.; Bliska, J.B.; van der Velden, A.W.M. CD11b+ Ly6Chi Ly6G− immature myeloid cells recruited in response to Salmonella enterica serovar Typhimurium infection exhibit protective and immunosuppressive properties. Infect. Immunity 2014, 82, 2606–2614. [Google Scholar] [CrossRef]
- Kaimala, S.; Mohamed, Y.A.; Nader, N.; Issac, J.; Elkord, E.; Chouaib, S.; Fernandez-Cabezudo, M.J.; al-Ramadi, B.K. Salmonella-mediated tumor regression involves targeting of tumor myeloid suppressor cells causing a shift to M1-like phenotype and reduction in suppressive capacity. Cancer Immunol. Immunother. 2014, 63, 587–599. [Google Scholar] [CrossRef]
- Lee, C.H.; Wu, C.L.; Shiau, A.L. Toll-like receptor 4 mediates an antitumor host response induced by Salmonella choleraesuis. Clin. Cancer Res. 2008, 14, 1905–1912. [Google Scholar] [CrossRef]
- Grille, S.; Moreno, M.; Bascua, T.; Marqués, J.M.; Muñoz, N.; Lens, D.; Chabalgoity, J.A. Salmonella enterica serovar Typhimurium immunotherapy for B-cell lymphoma induces broad anti-tumour immunity with therapeutic effect. Immunology 2014, 143, 428–437. [Google Scholar] [CrossRef]
- Kocijancic, D.; Leschner, S.; Felgner, S.; Komoll, R.M.; Frahm, M.; Pawar, V.; Weiss, S. Therapeutic benefit of Salmonella attributed to LPS and TNF-α is exhaustible and dictated by tumor susceptibility. Oncotarget 2017, 8, 36492–36508. [Google Scholar] [CrossRef]
- Zheng, J.H.; Nguyen, V.H.; Jiang, S.N.; Park, S.H.; Tan, W.; Hong, S.H.; Shin, M.G.; Chung, I.J.; Hong, Y.; Bom, H.S.; et al. Two-step enhanced cancer immunotherapy with engineered Salmonella typhimurium secreting heterologous flagellin. Sci. Transl. Med. 2017, 9, eaak9537. [Google Scholar] [CrossRef]
- De Melo, F.M.; Braga, C.J.; Pereira, F.V.; Maricato, J.T.; Origassa, C.S.; Souza, M.F.; Melo, A.C.; Silva, P.; Tomaz, S.L.; Gimenes, K.P.; et al. Anti-metastatic immunotherapy based on mucosal administration of flagellin and immunomodulatory P10. Immunol. Cell Biol. 2015, 93, 86–98. [Google Scholar] [CrossRef]
- Lee, C.H.; Hsieh, J.L.; Wu, C.L.; Hsu, P.Y.; Shiau, A.L. T cell augments the antitumor activity of tumor-targeting Salmonella. Appl. Microbiol. Biotechnol. 2011, 90, 1381–1388. [Google Scholar] [CrossRef]
- Stern, C.; Kasnitz, N.; Kocijancic, D.; Trittel, S.; Riese, P.; Guzman, C.A.; Leschner, S.; Weiss, S. Induction of CD4(+) and CD8(+) anti-tumor effector T cell responses by bacteria mediated tumor therapy. Int. J. Cancer 2015, 137, 2019–2028. [Google Scholar] [CrossRef]
- Avogadri, F.; Martinoli, C.; Petrovska, L.; Chiodoni, C.; Transidico, P.; Bronte, V.; Longhi, R.; Colombo, M.P.; Dougan, G.; Rescigno, M. Cancer immunotherapy based on killing of Salmonella-infected tumor cells. Cancer Res. 2005, 65, 3920–3927. [Google Scholar] [CrossRef]
- Saccheri, F.; Pozzi, C.; Avogadri, F.; Barozzi, S.; Faretta, M.; Fusi, P.; Rescigno, M. Bacteria-induced gap junctions in tumors favor antigen cross-presentation and antitumor immunity. Sci. Transl. Med. 2010, 2, 44–57. [Google Scholar] [CrossRef]
- Chang, W.W.; Lai, C.H.; Chen, M.C.; Liu, C.F.; Kuan, Y.D.; Lin, S.T.; Lee, C.H. Salmonella enhance chemosensitivity in tumor through connexin 43 upregulation. Int. J. Cancer 2013, 133, 1926–1935. [Google Scholar] [CrossRef] [Green Version]
- Brandacher, G.; Perathoner, A.; Ladurner, R.; Schneeberger, S.; Obrist, P.; Winkler, C.; Werner, E.R.; Werner-Felmayer, G.; Weiss, H.G.; Gobel, G.; et al. Prognostic value of indoleamine 2,3-dioxygenase expression in colorectal cancer: Effect on tumor-infiltrating T cells. Clin. Cancer Res. 2006, 12, 1144–1151. [Google Scholar] [CrossRef]
- Kuan, Y.D.; Lee, C.H. Salmonella overcomes tumor immune tolerance by inhibition of tumor indoleamine 2, 3-dioxygenase 1 expression. Oncotarget 2015, 7, 374–385. [Google Scholar] [CrossRef]
- Sun, J.; Yu, J.; Li, H.; Yang, L.; Wei, F.; Yu, W.; Liu, J.; Ren, X. Upregulated expression of indoleamine 2, 3-dioxygenase in CHO cells induces apoptosis of competent T cells and increases proportion of Treg cells. J. Exp. Clin. Cancer Res. 2011, 30, 82. [Google Scholar] [CrossRef]
- Blache, C.A.; Manuel, E.R.; Kaltcheva, T.I.; Wong, A.N.; Ellenhorn, J.D.I.; Blazar, B.R.; Diamond, D.J. Systemic delivery of Salmonella typhimurium transformed with IDO shRNA enhances intratumoral vector colonization and suppresses tumor growth. Cancer Res. 2012, 72, 6447–6456. [Google Scholar] [CrossRef]
- Lee, C.H.; Lin, S.T.; Liu, J.J.; Chang, W.W.; Hsieh, J.L.; Wang, W.K. Salmonella induce autophagy in melanoma by the downregulation of AKT/mTOR pathway. Gene Ther. 2014, 21, 309–316. [Google Scholar] [CrossRef]
- Fu, W.; Chu, L.; Han, X.; Liu, X.; Ren, D. Synergistic antitumoral effects of human telomerase reverse transcriptase-mediated dual-apoptosis-related gene vector delivered by orally attenuated Salmonella enterica Serovar Typhimurium in murine tumor models. J. Gene Med. 2008, 10, 690–701. [Google Scholar] [CrossRef]
- Chen, J.; Yang, B.; Cheng, X.; Qiao, Y.; Tang, B.; Chen, G.; Wei, J.; Liu, X.; Cheng, W.; Du, P.; et al. Salmonella-mediated tumor-targeting TRAIL gene therapy significantly suppresses melanoma growth in mouse model. Cancer Sci. 2012, 103, 325–333. [Google Scholar] [CrossRef]
- Tu, D.G.; Chang, W.W.; Lin, S.T.; Kuo, C.Y.; Tsao, Y.T.; Lee, C.H. Salmonella inhibits tumor angiogenesis by downregulation of vascular endothelial growth factor. Oncotarget 2016, 7, 37513–37523. [Google Scholar] [CrossRef]
- Tsao, Y.T.; Kuo, C.Y.; Cheng, S.P.; Lee, C.H. Downregulations of AKT/mTOR Signaling Pathway for Salmonella-Mediated Suppression of Matrix Metalloproteinases-9 Expression in Mouse Tumor Models. Int. J. Mol. Sci. 2018, 19, 1630. [Google Scholar] [CrossRef]
- Yang, C.J.; Chang, W.W.; Lin, S.T.; Chen, M.C.; Lee, C.H. Salmonella Overcomes Drug resistance in tumor through P-glycoprotein downregulation. Int. J. Med. Sci. 2018, 15, 574–579. [Google Scholar] [CrossRef]
- Gupta, G.P.; Massagué, J. Cancer metastasis: Building a framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef]
- Nguyen, D.X.; Bos, P.D.; Massagué, J. Metastasis: From dissemination to organ-specific colonization. Nat. Rev. Cancer 2009, 9, 274–284. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Zetter, B.R. Angiogenesis and tumor metastasis. Annu. Rev. Med. 1998, 49, 407–424. [Google Scholar] [CrossRef]
- Baeriswyl, V.; Christofori, G. The angiogenic switch in carcinogenesis. Semin. Cancer Biol. 2009, 19, 329–337. [Google Scholar] [CrossRef]
- Ferrara, N. Vascular endothelial growth factor. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 789–791. [Google Scholar] [CrossRef]
- Carmeliet, P. VEGF as a key mediator of angiogenesis in cancer. Oncology 2005, 69, 4–10. [Google Scholar] [CrossRef]
- Semenza, G.L. Targeting HIF-1 for cancer therapy. Nat. Rev. Cancer 2003, 3, 721–732. [Google Scholar] [CrossRef]
- Chen, M.C.; Hsu, W.L.; Hwang, P.A.; Chou, T.C. Low molecular weight fucoidan inhibits tumor angiogenesis through downregulation of HIF-1/VEGF signaling under Hypoxia. Mar. Drugs 2015, 13, 4436–4451. [Google Scholar] [CrossRef]
- Ataie-Kachoie, P.; Pourgholami, M.H.; Bahrami, B.F.; Badar, S.; Morris, D.L. Minocycline attenuates hypoxia-inducible factor-1α expression correlated with modulation of p53 andAKT/mTOR/p70S6K/4E-BP1 pathway in ovarian cancer: In vitro and in vivo studies. Am. J. Cancer Res. 2015, 5, 575–588. [Google Scholar] [PubMed]
- Yilmaz, M.; Christofori, G.; Lehembre, F. Distinct mechanisms of tumor invasion and metastasis. Trends Mol. Med. 2007, 13, 535–541. [Google Scholar] [CrossRef]
- Stamenkovic, I. Extracellular matrix remodelling: The role of matrix metalloproteinases. J. Pathol. 2003, 200, 448–464. [Google Scholar] [CrossRef]
- Parks, W.C.; Wilson, C.L.; Lopez-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221. [Google Scholar] [CrossRef]
- Chen, P.; Parks, W.C. Role of matrix metalloproteinases in epithelial migration. J. Cell. Biochem. 2009, 108, 1233–1243. [Google Scholar] [CrossRef]
- Klein, G.; Vellenga, E.; Fraaije, M.; Kamps, W.; De Bont, E. The possible role of matrix metalloproteinase (MMP)-2 and MMP-9 in cancer, e.g., acute leukemia. Crit. Rev. Oncol. Hematol. 2004, 50, 87–100. [Google Scholar] [CrossRef]
- Cotignola, J.; Reva, B.; Mitra, N.; Ishill, N.; Chuai, S.; Patel, A.; Shah, S.; Vanderbeek, G.; Coit, D.; Busam, K.; et al. Matrix Metalloproteinase-9 (MMP-9) polymorphisms in patients with cutaneous malignant melanoma. BMC Med. Genet. 2007, 8, 10. [Google Scholar] [CrossRef]
- Mehner, C.; Hockla, A.; Miller, E.; Ran, S.; Radisky, D.C.; Radisky, E.S. Tumor cell-produced matrix metalloproteinase 9 (MMP-9) drives malignant progression and metastasis of basal-like triple negative breast cancer. Oncotarget 2014, 5, 2736–2749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Yan, Y.; Cheng, Z.; Hu, Y.; Liu, T. Sotetsuflavone suppresses invasion and metastasis in non-small-cell lung cancer A549 cells by reversing EMT via the TNF-α/NF-κB and PI3K/AKT signaling pathway. Cell Death Discov. 2018, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zheng, L.; Liu, J.; Zhou, Z.; Cao, X.; Lv, X.; Chen, F. Shikonin inhibits prostate cancer cells metastasis by reducing matrix metalloproteinase-2/-9 expression via AKT/mTOR and ROS/ERK1/2 pathways. Int. Immunopharmacol. 2014, 21, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Kou, B.; Liu, W.; He, W.; Zhang, Y.Y.; Zheng, J.J.; Yan, Y.; Zhang, Y.J.; Xu, S.C.; Wang, H. Tetrandrine suppresses metastatic phenotype of prostate cancer cells by regulating Akt/mTOR/MMP-9 signaling pathway. Oncol. Rep. 2016, 35, 2880–2886. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, Q.; Bao, J.; Du, C.; Wang, J.; Tong, Q.; Liu, C. Schisandrin B suppresses glioma cell metastasis mediated by inhibition of mTOR/MMP-9 signal pathway. Biomed. Pharmacother. 2015, 74, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, W.; Wang, K.; Fan, Y.; Chen, J.; Ma, J.; Wang, X.; He, D.; Zeng, J.; Li, L. Tetrandrine inhibits migration and invasion of human renal cell carcinoma by regulating Akt/NF-κB/MMP-9 signaling. PLoS ONE 2017, 12, e0173725. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, A.; Dayan, G.; Conseil, G.; Steinfels, E.; Krell, T.; Trompier, D.; Baubichon-Cortay, H.; Jault, J.M. P-glycoprotein-mediated resistance to chemotherapy in cancer cells: Using recombinant cytosolic domains to establish structure-function relationships. Braz. J. Med. Biol. Res. 1999, 32, 925–939. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, M.; Pavlopoulou, A.; Georgakilas, A.G.; Kyrodimos, E. The challenge of drug resistance in cancer treatment: A current overview. Clin. Exp. Metastasis 2018, 35, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Mohammadi, A.; Davudian, S.; Shirjang, S.; Baradaran, B. The different mechanisms of cancer drug resistance: A brief review. Adv. Pharm. Bull. 2017, 7, 339–348. [Google Scholar] [CrossRef]
- Siccardi, D.; Mumy, K.L.; Wall, D.M.; Bien, J.D.; McCormick, B.A. Salmonella enterica serovar Typhimurium modulates P-glycoprotein in the intestinal epithelium. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1392–G1400. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.Y.; Hu, M.E.; Yang, T.Y.; Hsin, I.L.; Ko, J.L.; Tsai, K.J.; Sheu, G.T. Immunomodulatory protein from Ganoderma microsporum induces pro-death autophagy through Akt-mTOR-p70S6K pathway inhibition in multidrug resistant lung cancer cells. PLoS ONE 2015, 10, e0125774. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, C.; Jia, Y.; Liu, Z.; Shu, X.; Liu, K. Resveratrol increases anti-proliferative activity of Bestatin through downregulating P-Glycoprotein expression via inhibiting PI3K/Akt/mTOR pathway in K562/ADR cells. J. Cell Biochem. 2016, 117, 1233–1239. [Google Scholar] [CrossRef]
- Mercado-Lubo, R.; Zhang, Y.; Zhao, L.; Rossi, K.; Wu, X.; Zou, Y.; Castillo, A.; Leonard, J.; Bortell, R.; Greiner, D.L.; et al. A Salmonella nanoparticle mimic overcomes multidrug resistance in tumours. Nat. Commun. 2016, 7, 12225. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.J.; Wei, D.P.; Sun, Q.M.; Jin, G.H.; Li, S.F.; Huang, Y.; Hua, Z.C. Tumor-targeting Salmonella typhimurium improves cyclophosphamide chemotherapy at maximum tolerated dose and low-dose metronomic regimens in a murine melanoma model. Int. J. Cancer 2007, 121, 666–674. [Google Scholar] [CrossRef]
- Bascuas, T.; Moreno, M.; Grille, S.; Chabalgoity, J.A. Salmonella immunotherapy improves the outcome of CHOP chemotherapy in non-Hodgkin lymphoma-bearing mice. Front. Immunol. 2018, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Binder, D.C.; Arina, A.; Wen, F.; Tu, T.; Zhao, M.; Hoffman, R.M.; Wainwright, D.A.; Schreiber, H. Tumor relapse prevented by combining adoptive T cell therapy with Salmonella typhimurium. Oncoimmunology 2016, 5, e1130207. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, Y.; Qin, M.; Zhang, X.; Zhang, Z.; Sun, X.; Gu, Z. Bacteria-driven hypoxia targeting for combined biotherapy and photothermal therapy. ACS Nano 2018, 12, 5995–6005. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Cheng, P.; Chen, P.; Pu, K. Recent progress in the development of near-infrared organic photothermal and photodynamic nanotherapeutics. Biomater. Sci. 2018, 6, 746–765. [Google Scholar] [CrossRef] [PubMed]
- Toso, J.F.; Gill, V.J.; Hwu, P.; Marincola, F.M.; Restifo, N.P.; Schwartzentruber, D.J.; Sherry, R.M.; Topalian, S.L.; Yang, J.C.; Stock, F.; et al. Phase I study of the intravenous administration of attenuated Salmonella typhimurium to patients with metastatic melanoma. J. Clin. Oncol. 2002, 20, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Kazmierczak, R.A.; Eisenstark, A. Strains, Mechanism, and Perspective: Salmonella-Based Cancer Therapy. Int. J. Microbiol. 2016, 5678702. [Google Scholar] [CrossRef]
- Park, S.H.; Zheng, J.H.; Nguyen, V.H.; Jiang, S.N.; Kim, D.Y.; Szardenings, M.; Min, J.H.; Hong, Y.; Choy, H.E.; Min, J.J. RGD Peptide cell-surface display enhances the targeting and therapeutic efficacy of attenuated salmonella-mediated cancer therapy. Theranostics 2016, 6, 1672–1682. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.H.; Min, J.J. Targeted Cancer Therapy Using Engineered Salmonella typhimurium. Chonnam Med. J. 2016, 52, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Toley, B.J.; Swofford, C.A.; Forbes, N.S. construction of an inducible cell-communication system that amplifies Salmonella gene expression in tumor tissue. Biotechnol. Bioeng. 2013, 110, 1769–1781. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S.; Ernst, R.K.; McCoy, A.J.; Miller, S.I. Constitutive Mutations of the Salmonella enterica Serovar Typhimurium Transcriptional Virulence Regulator phoP. Infect. Immun. 2000, 68, 3758–3762. [Google Scholar] [CrossRef]
- Niethammer, A.G.; Lubenau, H.; Mikus, G.; Knebel, P.; Hohmann, N.; Leowardi, C.; Beckhove, P.; Akhisaroglu, M.; Ge, Y.; Springer, M.; et al. Double-blind, placebo-controlled first in human study to investigate an oral vaccine aimed to elicit an immune reaction against the VEGF-Receptor 2 in patients with stage IV and locally advanced pancreatic cancer. BMC Cancer 2012, 12, 361. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Winnenthal, F.H.; Hohmann, N.; Niethammer, A.G.; Friedrich, T.; Lubenau, H.; Springer, M.; Breiner, K.M.; Mikus, G.; Weitz, J.; Ulrich, A.; et al. Anti-angiogenic activity of VXM01, an oral T-cell vaccine against VEGF receptor 2, in patients with advanced pancreatic cancer: A randomized, placebo-controlled, phase 1 trial. Oncoimmunology 2015, 4, e1001217. [Google Scholar] [CrossRef] [Green Version]
- Schmitz-Winnenthal, F.H.; Hohmann, N.; Schmidt, T.; Podola, L.; Friedrich, T.; Lubenau, H.; Springer, M.; Wieckowski, S.; Breiner, K.M.; Mikus, G.; et al. A phase 1 trial extension to assess immunologic efficacy and safety of prime-boost vaccination with VXM01, an oral T cell vaccine against VEGFR2, in patients with advanced pancreatic cancer. Oncoimmunology 2018, 7, e1303584. [Google Scholar] [CrossRef] [PubMed]
- U.S. National Institutes of Health; Clinicaltrials.gov. Multiple Myeloma Trial of Orally Administered Salmonella Based Survivin Vaccine (MAPSS). Identifier: NCT03762291. Available online: https://clinicaltrials.gov/ct2/show/NCT03762291 (accessed on 23 April 2019).



{kind=link}
{kind=link}
{kind=link}
| Protein Targets | Control | Mechanisms | Ref. |
|---|---|---|---|
| Connexin 43 | ▲ | Increases tumor cell chemosensitivity | [41] |
| IDO | ▼ | Suppresses tumor immune tolerance | [43,45] |
| Beclin-1 and LC3 | ▲ | Induce autophagy | [46] |
| Cleaved Caspase-3 | ▲ | Activates apoptosis | [46] |
| HIF-1/VEGF | ▼ | Inhibits angiogenesis | [49] |
| Matrix MMP-9 | ▼ | Delays cell migration and metastasis | [50] |
| P-glycoprotein | ▼ | Increases tumor cell chemosensitivity | [51] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pangilinan, C.R.; Lee, C.-H. Salmonella-Based Targeted Cancer Therapy: Updates on A Promising and Innovative Tumor Immunotherapeutic Strategy. Biomedicines 2019, 7, 36. https://doi.org/10.3390/biomedicines7020036
Pangilinan CR, Lee C-H. Salmonella-Based Targeted Cancer Therapy: Updates on A Promising and Innovative Tumor Immunotherapeutic Strategy. Biomedicines. 2019; 7(2):36. https://doi.org/10.3390/biomedicines7020036
Chicago/Turabian StylePangilinan, Christian Ronquillo, and Che-Hsin Lee. 2019. "Salmonella-Based Targeted Cancer Therapy: Updates on A Promising and Innovative Tumor Immunotherapeutic Strategy" Biomedicines 7, no. 2: 36. https://doi.org/10.3390/biomedicines7020036
APA StylePangilinan, C. R., & Lee, C. -H. (2019). Salmonella-Based Targeted Cancer Therapy: Updates on A Promising and Innovative Tumor Immunotherapeutic Strategy. Biomedicines, 7(2), 36. https://doi.org/10.3390/biomedicines7020036