A Mini-Review: Clinical Development and Potential of Aptamers for Thrombotic Events Treatment and Monitoring



Abstract
:1. Introduction
1.1. A Brief History of Aptamers
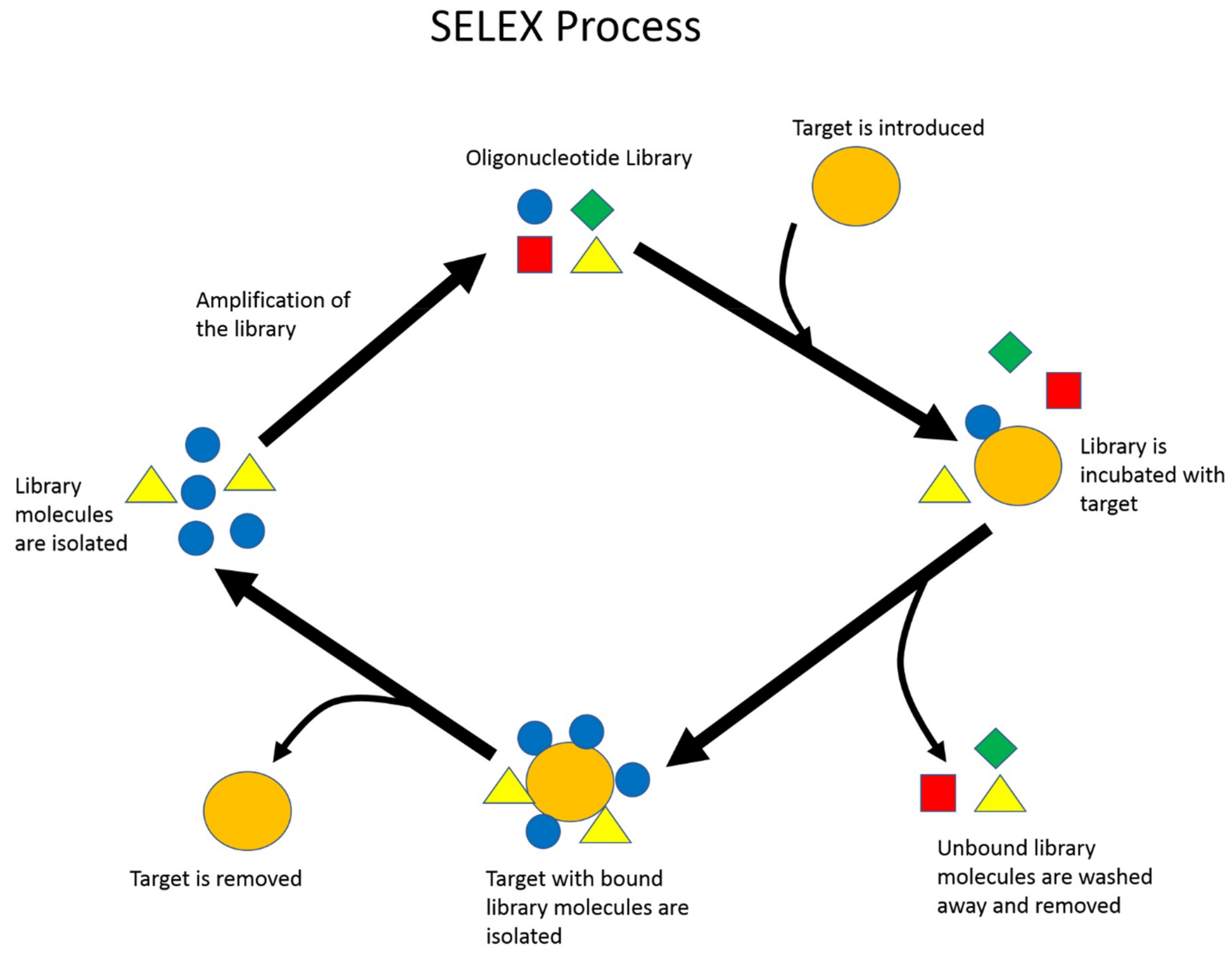
1.2. Aptamer Selection
1.3. Clinical Advantages and Disadvantages of Aptamers
2. Aptamer Clinical Research in Anticoagulation
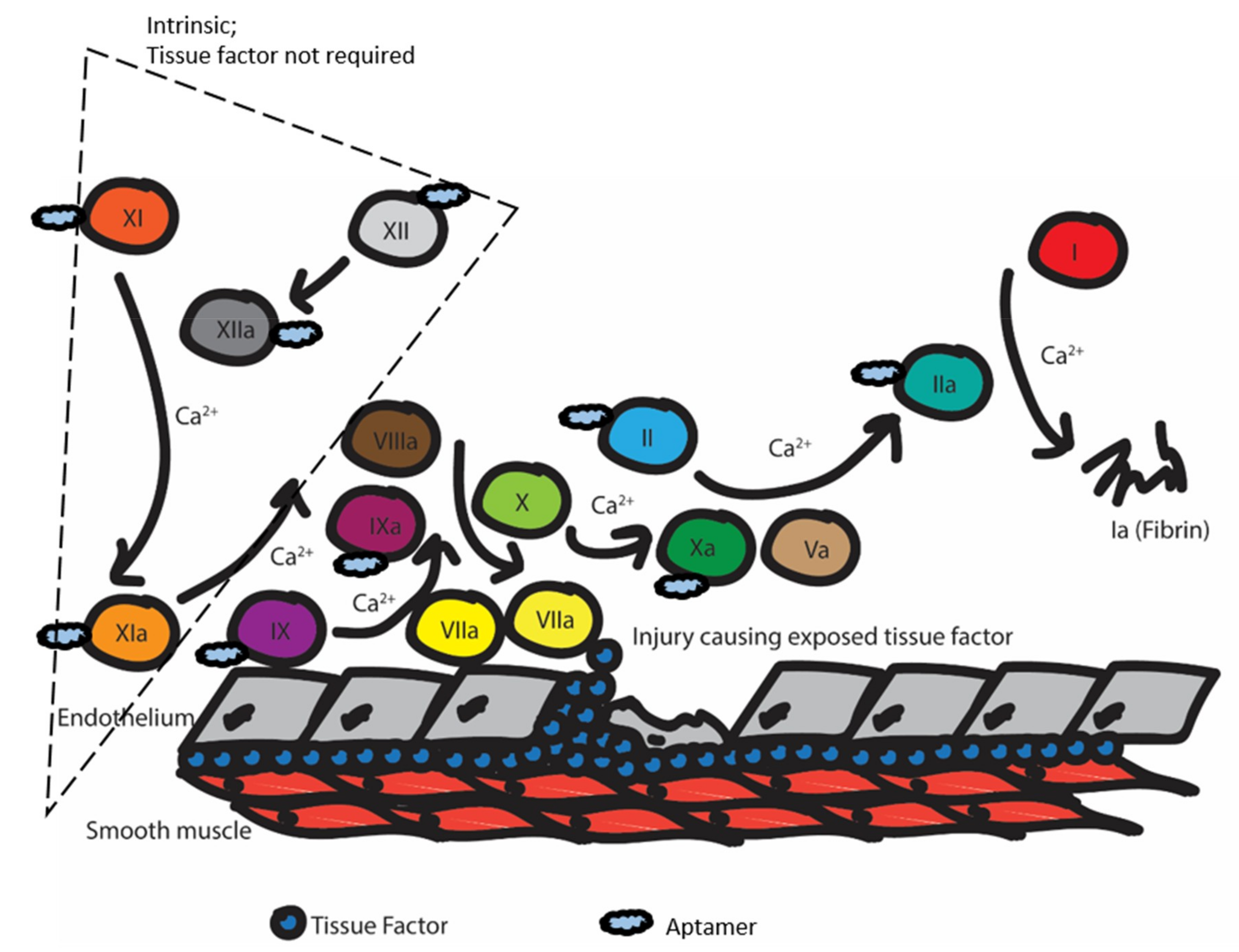
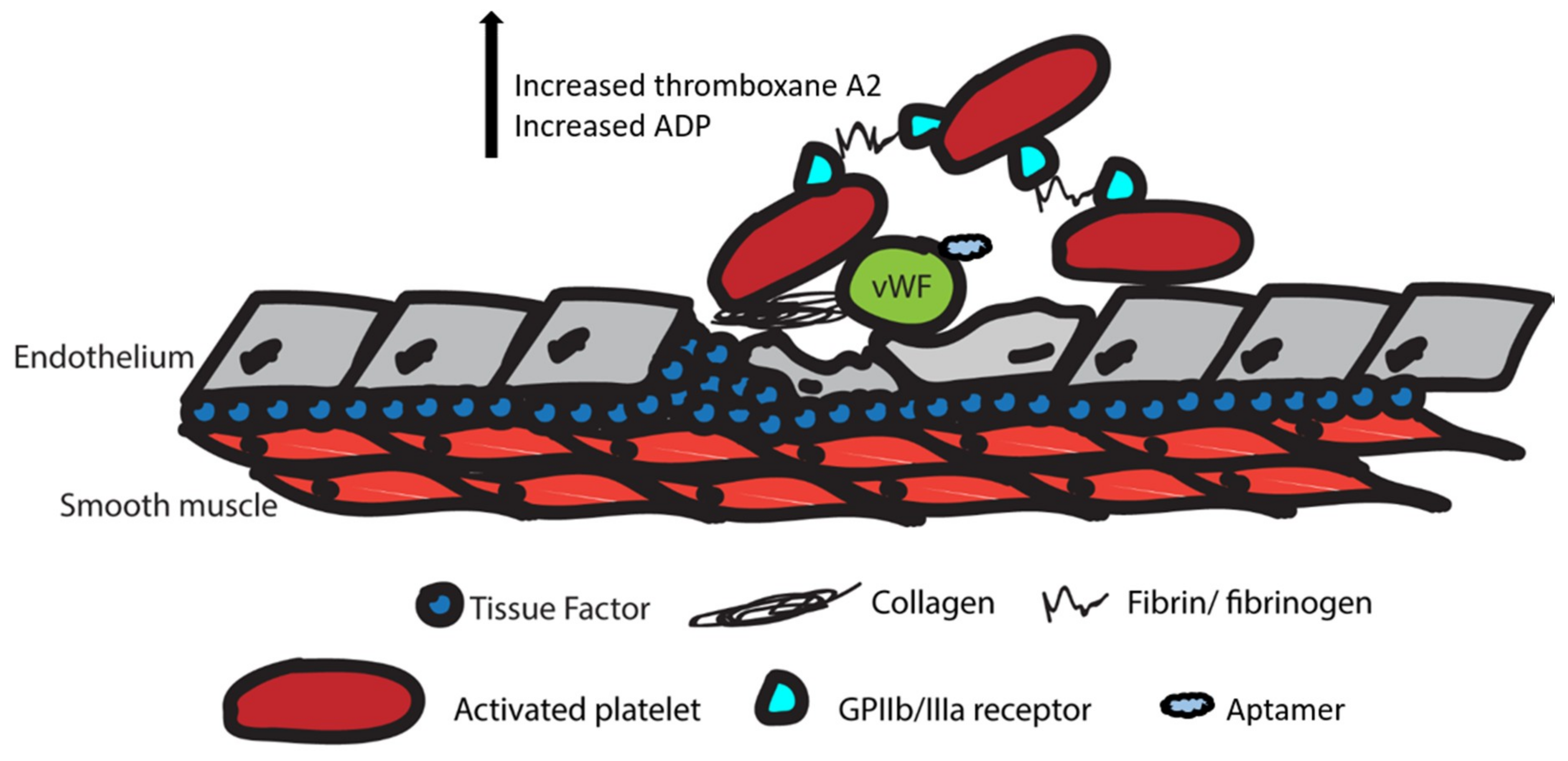
2.1. Summary of the Coagulation and Clotting Cascades
2.2. Anticoagulation Aptamers
2.2.1. Von Willebrand Factor Inhibitors
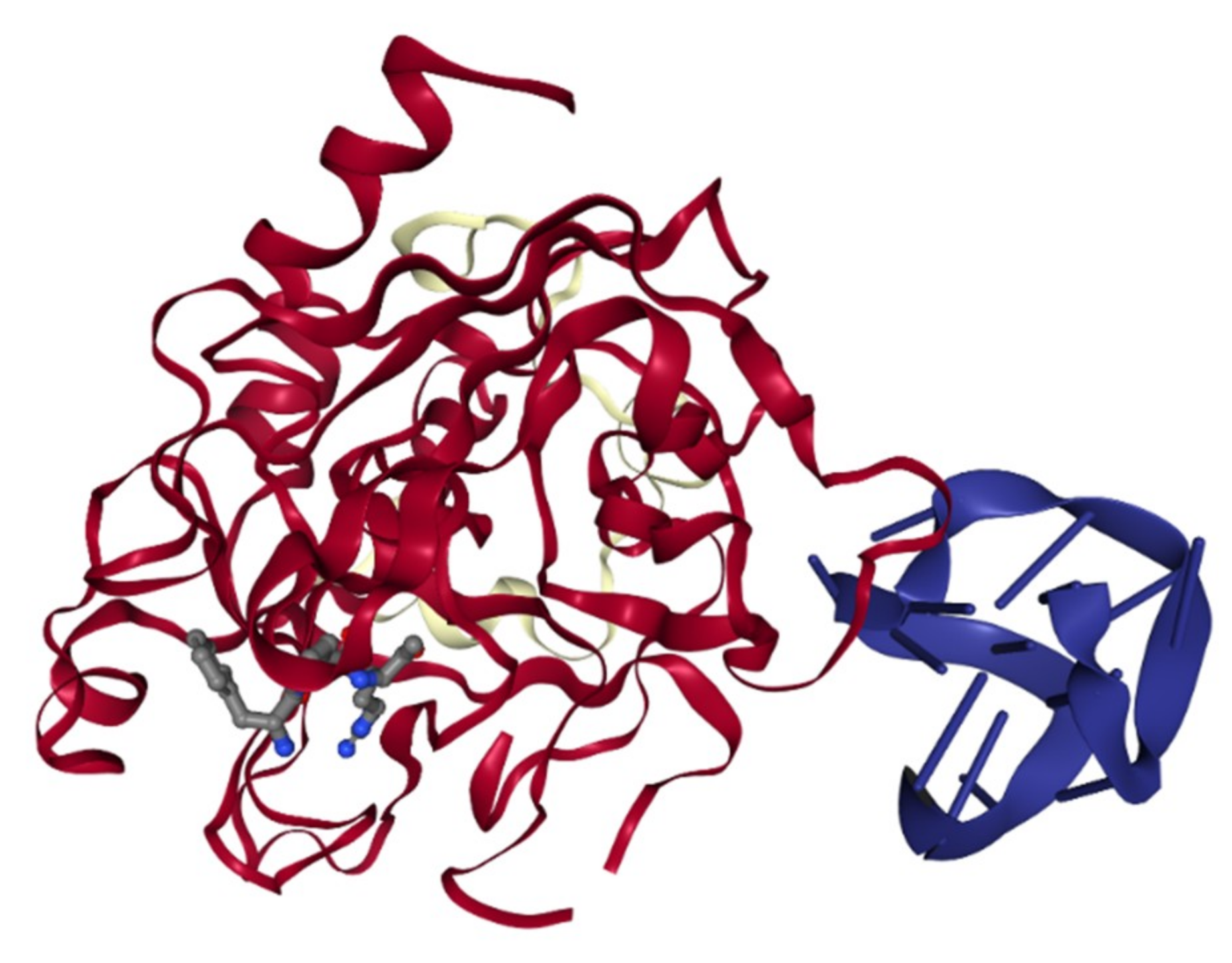
2.2.2. Factor II/ IIa Inhibitors
2.2.3. Factor IXa Inhibitors
2.2.4. Factor Xa Inhibitors
2.2.5. Factor XIa Inhibitors
2.2.6. Factor XII/ XIIa Inhibitors
2.2.7. Kallikrein Inhibitors
3. Aptamer in Anticoagulation Monitoring
4. Conclusions and Future Perspective
Author Contributions
Funding
Conflicts of Interest
References
- De la Torre, B.G.; Albericio, F. The Pharmaceutical Industry in 2017. An Analysis of FDA Drug Approvals from the Perspective of Molecules. Molecules 2018, 23, 533. [Google Scholar] [CrossRef] [PubMed]
- Lakhin, A.V.; Tarantul, V.Z.; Gening, L.V. Aptamers: problems, solutions and prospects. Acta Nat. 2013, 5, 34–43. [Google Scholar] [CrossRef]
- Tuerk, C.; Gold, L. Systematic evolution of ligands by exponential enrichment: RNA ligands to bacteriophage T4 DNA polymerase. Science 1990, 249, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Gold, L.; Janjic, N.; Jarvis, T.; Schneider, D.; Walker, J.J.; Wilcox, S.K.; Zichi, D. Aptamers and the RNA world, past and present. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Ismail, S.I.; Alshaer, W. Therapeutic aptamers in discovery, preclinical and clinical stages. Adv. Drug Deliv. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.L.; Sooter, L.J. Single-Stranded DNA Aptamers against Pathogens and Toxins: Identification and Biosensing Applications. BioMed Res. Int. 2015, 2015, 419318. [Google Scholar] [CrossRef]
- Harbaugh, S.V.; Martin, J.A.; Weinstein, J.; Ingram, G.; Kelley-Loughnane, N. Screening and selection of artificial riboswitches. Methods 2018, 143, 77–89. [Google Scholar] [CrossRef]
- Ni, X.; Castanares, M.; Mukherjee, A.; Lupold, S.E. Nucleic acid aptamers: Clinical applications and promising new horizons. Curr. Med. Chem. 2011, 18, 4206–4214. [Google Scholar] [CrossRef]
- Kanwar, J.R.; Roy, K.; Maremanda, N.G.; Subramanian, K.; Veedu, R.N.; Bawa, R.; Kanwar, R.K. Nucleic acid-based aptamers: Applications, development and clinical trials. Curr. Med. Chem. 2015, 22, 2539–2557. [Google Scholar] [CrossRef]
- Park, J.W.; Tatavarty, R.; Kim, D.W.; Jung, H.T.; Gu, M.B. Immobilization-free screening of aptamers assisted by graphene oxide. Chem. Commun. 2012, 48, 2071–2073. [Google Scholar] [CrossRef] [PubMed]
- Ruscito, A.; DeRosa, M.C. Small-Molecule Binding Aptamers: Selection Strategies, Characterization, and Applications. Front. Chem. 2016, 4, 14. [Google Scholar] [CrossRef] [PubMed]
- Catuogno, S.; Esposito, C.L. Aptamer Cell-Based Selection: Overview and Advances. Biomedicines 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Huang, R.; Deng, Y.; He, N. Progress in selection and biomedical applications of aptamers. J. Biomed. Nanotechnol. 2014, 10, 3043–3062. [Google Scholar] [CrossRef] [PubMed]
- White, R.R.; Sullenger, B.A.; Rusconi, C.P. Developing aptamers into therapeutics. J. Clin. Investig. 2000, 106, 929–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, S.; Yao, H.; Wang, L.; Lu, J.; Jiang, F.; Lu, A.; Zhang, G. Chemical Modifications of Nucleic Acid Aptamers for Therapeutic Purposes. Int. J. Mol. Sci. 2017, 18, 1683. [Google Scholar] [CrossRef]
- Wang, R.E.; Wu, H.; Niu, Y.; Cai, J. Improving the stability of aptamers by chemical modification. Curr. Med. Chem. 2011, 18, 4126–4138. [Google Scholar] [CrossRef]
- Matsunaga, K.I.; Kimoto, M.; Hirao, I. High-Affinity DNA Aptamer Generation Targeting von Willebrand Factor A1-Domain by Genetic Alphabet Expansion for Systematic Evolution of Ligands by Exponential Enrichment Using Two Types of Libraries Composed of Five Different Bases. J. Am. Chem. Soc. 2017, 139, 324–334. [Google Scholar] [CrossRef]
- Dua, P.; Kim, S.; Lee, D.K. Nucleic acid aptamers targeting cell-surface proteins. Methods 2011, 54, 215–225. [Google Scholar] [CrossRef]
- Takahashi, M. Aptamers targeting cell surface proteins. Biochimie 2018, 145, 63–72. [Google Scholar] [CrossRef]
- Yoon, S.; Rossi, J.J. Targeted Molecular Imaging Using Aptamers in Cancer. Pharmaceuticals 2018, 11, 71. [Google Scholar] [CrossRef] [PubMed]
- De Caterina, R.; Husted, S.; Wallentin, L.; Andreotti, F.; Arnesen, H.; Bachmann, F.; Baigent, C.; Huber, K.; Jespersen, J.; Kristensen, S.D.; et al. General mechanisms of coagulation and targets of anticoagulants (Section I). Position Paper of the ESC Working Group on Thrombosis—Task Force on Anticoagulants in Heart Disease. Thromb. Haemost. 2013, 109, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Diener, J.L.; Daniel Lagasse, H.A.; Duerschmied, D.; Merhi, Y.; Tanguay, J.F.; Hutabarat, R.; Gilbert, J.; Wagner, D.D.; Schaub, R. Inhibition of von Willebrand factor-mediated platelet activation and thrombosis by the anti-von Willebrand factor A1-domain aptamer ARC1779. J. Thromb. Haemost. 2009, 7, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Nimjee, S.M.; Dornbos, D.; Pitoc, G.A.; Wheeler, D.G.; Layzer, J.M.; Venetos, N.; Huttinger, A.; Talentino, S.E.; Musgrave, N.J.; Moody, H.; et al. Preclinical Development of a vWF Aptamer to Limit Thrombosis and Engender Arterial Recanalization of Occluded Vessels. Mol. Ther. 2019, 27, 1228–1241. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Outes, A.; Suarez-Gea, M.L.; Lecumberri, R.; Rocha, E.; Pozo-Hernandez, C.; Vargas-Castrillon, E. New parenteral anticoagulants in development. Ther. Adv. Cardiovasc. Dis. 2011, 5, 33–59. [Google Scholar] [CrossRef] [PubMed]
- Trapaidze, A.; Hérault, J.P.; Herbert, J.M.; Bancaud, A.; Gué, A.M. Investigation of the selectivity of thrombin-binding aptamers for thrombin titration in murine plasma. Biosens. Bioelectron. 2016, 78, 58–66. [Google Scholar] [CrossRef]
- MÜLler, J.; Freitag, D.; Mayer, G.; PÖTzsch, B. Anticoagulant characteristics of HD1-22, a bivalent aptamer that specifically inhibits thrombin and prothrombinase. J. Thromb. Haemost. 2008, 6, 2105–2112. [Google Scholar] [CrossRef]
- Kotkowiak, W.; Wengel, J.; Scotton, C.J.; Pasternak, A. Improved RE31 Analogues Containing Modified Nucleic Acid Monomers: Thermodynamic, Structural, and Biological Effects. J. Med. Chem. 2019, 62, 2499–2507. [Google Scholar] [CrossRef]
- Wakui, K.; Yoshitomi, T.; Yamaguchi, A.; Tsuchida, M.; Saito, S.; Shibukawa, M.; Furusho, H.; Yoshimoto, K. Rapidly Neutralizable and Highly Anticoagulant Thrombin-Binding DNA Aptamer Discovered by MACE SELEX. Mol. Ther. Nucleic Acids 2019, 16, 348–359. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Qi, X.; Liu, Y.; Zhang, F.; Yan, H. DNA Nanoscaffold-Assisted Selection of Femtomolar Bivalent Aptamers for Human α-Thrombin with Potent Anticoagulant Activity. ChemBioChem 2019. [Google Scholar] [CrossRef]
- Bompiani, K.M.; Monroe, D.M.; Church, F.C.; Sullenger, B.A. A high affinity, antidote-controllable prothrombin and thrombin-binding RNA aptamer inhibits thrombin generation and thrombin activity. J. Thromb. Haemost. 2012, 10, 870–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyke, C.K.; Steinhubl, S.R.; Kleiman, N.S.; Cannon, R.O.; Aberle, L.G.; Lin, M.; Myles, S.K.; Melloni, C.; Harrington, R.A.; Alexander, J.H.; et al. First-in-human experience of an antidote-controlled anticoagulant using RNA aptamer technology: A phase 1a pharmacodynamic evaluation of a drug-antidote pair for the controlled regulation of factor IXa activity. Circulation 2006, 114, 2490–2497. [Google Scholar] [CrossRef] [PubMed]
- Lincoff, A.M.; Mehran, R.; Povsic, T.J.; Zelenkofske, S.L.; Huang, Z.; Armstrong, P.W.; Steg, P.G.; Bode, C.; Cohen, M.G.; Buller, C.; et al. Effect of the REG1 anticoagulation system versus bivalirudin on outcomes after percutaneous coronary intervention (REGULATE-PCI): A randomised clinical trial. Lancet 2016, 387, 349–356. [Google Scholar] [CrossRef]
- Vavalle, J.P.; Cohen, M.G. The REG1 anticoagulation system: A novel actively controlled factor IX inhibitor using RNA aptamer technology for treatment of acute coronary syndrome. Future Cardiol. 2012, 8, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Vavalle, J.P.; Rusconi, C.P.; Zelenkofske, S.; Wargin, W.A.; Alexander, J.H.; Becker, R.C. A phase 1 ascending dose study of a subcutaneously administered factor IXa inhibitor and its active control agent. J. Thromb. Haemost. 2012, 10, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Buddai, S.K.; Layzer, J.M.; Lu, G.; Rusconi, C.P.; Sullenger, B.A.; Monroe, D.M.; Krishnaswamy, S. An Anticoagulant RNA Aptamer That Inhibits Proteinase-Cofactor Interactions within Prothrombinase. J. Biol. Chem. 2010, 285, 5212–5223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donkor, D.A.; Bhakta, V.; Eltringham-Smith, L.J.; Stafford, A.R.; Weitz, J.I.; Sheffield, W.P. Selection and characterization of a DNA aptamer inhibiting coagulation factor XIa. Sci. Rep. 2017, 7, 2102. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, R.S.; Ivanov, I.; Verhamme, I.M.; Sun, M.F.; Gailani, D.; Sullenger, B.A. Generation and characterization of aptamers targeting factor XIa. Thromb. Res. 2017, 156, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Woodruff, R.S.; Xu, Y.; Layzer, J.; Wu, W.; Ogletree, M.L.; Sullenger, B.A. Inhibiting the intrinsic pathway of coagulation with a factor XII–targeting RNA aptamer. J. Thromb. Haemost. 2013, 11, 1364–1373. [Google Scholar] [CrossRef]
- Steen Burrell, K.A.; Layzer, J.; Sullenger, B.A. A kallikrein-targeting RNA aptamer inhibits the intrinsic pathway of coagulation and reduces bradykinin release. J. Thromb. Haemost. 2017, 15, 1807–1817. [Google Scholar] [CrossRef]
- Jilma-Stohlawetz, P.; Gilbert, J.C.; Gorczyca, M.E.; Knobl, P.; Jilma, B. A dose ranging phase I/II trial of the von Willebrand factor inhibiting aptamer ARC1779 in patients with congenital thrombotic thrombocytopenic purpura. Thromb. Haemost. 2011, 106, 539–547. [Google Scholar] [CrossRef] [PubMed]
- Mazurov, A.V.; Titaeva, E.V.; Khaspekova, S.G.; Storojilova, A.N.; Spiridonova, V.A.; Kopylov, A.M.; Dobrovolsky, A.B. Characteristics of a New DNA Aptamer, Direct Inhibitor of Thrombin. Bull. Exp. Biol. Med. 2011, 150, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Derszniak, K.; Przyborowski, K.; Matyjaszczyk, K.; Moorlag, M.; de Laat, B.; Nowakowska, M.; Chlopicki, S. Comparison of Effects of Anti-thrombin Aptamers HD1 and HD22 on Aggregation of Human Platelets, Thrombin Generation, Fibrin Formation, and Thrombus Formation Under Flow Conditions. Front. Pharmacol. 2019, 10, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, T.X.; Lai, P.X.; Mao, J.Y.; Chu, H.W.; Unnikrishnan, B.; Anand, A.; Huang, C.C. Supramolecular Aptamers on Graphene Oxide for Efficient Inhibition of Thrombin Activity. Front. Chem. 2019, 7, 280. [Google Scholar] [CrossRef] [PubMed]
- Krissanaprasit, A.; Key, C.; Fergione, M.; Froehlich, K.; Pontula, S.; Hart, M.; Carriel, P.; Kjems, J.; Andersen, E.S.; LaBean, T.H. Genetically Encoded, Functional Single-Strand RNA Origami: Anticoagulant. Adv. Mater. 2019, 31, 1808262. [Google Scholar] [CrossRef] [PubMed]
- Amato, T.; Virgilio, A.; Pirone, L.; Vellecco, V.; Bucci, M.; Pedone, E.; Esposito, V.; Galeone, A. Investigating the properties of TBA variants with twin thrombin binding domains. Sci. Rep. 2019, 9, 9184. [Google Scholar] [CrossRef]
- Ganson, N.J.; Povsic, T.J.; Sullenger, B.A.; Alexander, J.H.; Zelenkofske, S.L.; Sailstad, J.M.; Rusconi, C.P.; Hershfield, M.S. Pre-existing anti-polyethylene glycol antibody linked to first-exposure allergic reactions to pegnivacogin, a PEGylated RNA aptamer. J. Allergy Clin. Immunol. 2016, 137, 1610–1613. [Google Scholar] [CrossRef]
- Povsic, T.J.; Lawrence, M.G.; Lincoff, A.M.; Mehran, R.; Rusconi, C.P.; Zelenkofske, S.L.; Huang, Z.; Sailstad, J.; Armstrong, P.W.; Steg, P.G.; et al. Pre-existing anti-PEG antibodies are associated with severe immediate allergic reactions to pegnivacogin, a PEGylated aptamer. J. Allergy Clin. Immunol. 2016, 138, 1712–1715. [Google Scholar] [CrossRef] [Green Version]
- Staudacher, D.L.; Putz, V.; Heger, L.; Reinöhl, J.; Hortmann, M.; Zelenkofske, S.L.; Becker, R.C.; Rusconi, C.P.; Bode, C.; Ahrens, I. Direct factor IXa inhibition with the RNA-aptamer pegnivacogin reduces platelet reactivity in vitro and residual platelet aggregation in patients with acute coronary syndromes. Eur. Heart J. Acute Cardiovasc. Care 2017. [Google Scholar] [CrossRef]
- Moreno, A.; Pitoc, G.A.; Ganson, N.J.; Layzer, J.M.; Hershfield, M.S.; Tarantal, A.F.; Sullenger, B.A. Anti-PEG Antibodies Inhibit the Anticoagulant Activity of PEGylated Aptamers. Cell Chem. Biol. 2019, 26, 634–644. [Google Scholar] [CrossRef]
- Gunaratne, R.; Kumar, S.; Frederiksen, J.W.; Stayrook, S.; Lohrmann, J.L.; Perry, K.; Bompiani, K.M.; Chabata, C.V.; Thalji, N.K.; Ho, M.D.; et al. Combination of aptamer and drug for reversible anticoagulation in cardiopulmonary bypass. Nat. Biotechnol. 2018, 36, 606. [Google Scholar] [CrossRef] [PubMed]
- Bock, L.C.; Griffin, L.C.; Latham, J.A.; Vermaas, E.H.; Toole, J.J. Selection of single-stranded DNA molecules that bind and inhibit human thrombin. Nature 1992, 355, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, K.; Padmanabhan, K.P.; Ferrara, J.D.; Sadler, J.E.; Tulinsky, A. The structure of alpha-thrombin inhibited by a 15-mer single-stranded DNA aptamer. J. Biol. Chem. 1993, 268, 17651–17654. [Google Scholar] [PubMed]
- Tucker, W.O.; Shum, K.T.; Tanner, J.A. G-quadruplex DNA Aptamers and their Ligands: Structure, Function and Application. Curr. Pharm. Des. 2012, 18, 2014–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platella, C.; Riccardi, C.; Montesarchio, D.; Roviello, G.N.; Musumeci, D. G-quadruplex-based aptamers against protein targets in therapy and diagnostics. Biochim. Biophys. Acta (BBA) Gen. Subj. 2017, 1861, 1429–1447. [Google Scholar] [CrossRef] [PubMed]
- Anna, A.; Carme, F.; Maria, T.; Ramon, E. Thrombin Binding Aptamer, More than a Simple Aptamer: Chemically Modified Derivatives and Biomedical Applications. Curr. Pharm. Des. 2012, 18, 2036–2047. [Google Scholar] [CrossRef] [Green Version]
- Deng, B.; Lin, Y.; Wang, C.; Li, F.; Wang, Z.; Zhang, H.; Li, X.F.; Le, X.C. Aptamer binding assays for proteins: The thrombin example—A review. Anal. Chim. Acta 2014, 837, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.C.; Gopinath, S.C.B.; Rebecca, L.W.X.; Perumal, V.; Lakshmipriya, T.; Saheed, M.S.M. Diagnosing human blood clotting deficiency. Int. J. Biol. Macromol. 2018, 116, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Djulbegovic, M.; Lee, A.I. An Update on the “Novel” and Direct Oral Anticoagulants, and Long-Term Anticoagulant Therapy. Clin. Chest Med. 2018, 39, 583–593. [Google Scholar] [CrossRef]
- Artang, R.; Anderson, M.; Riley, P.; Nielsen, J.D. Assessment of the effect of direct oral anticoagulants dabigatran, rivaroxaban, and apixaban in healthy male volunteers using a thrombin generation assay. Res. Pract. Thromb. Haemost. 2017, 1, 194–201. [Google Scholar] [CrossRef]
- Pursley, J.; Shen, J.X.; Schuster, A.; Dang, O.T.; Lehman, J.; Buonarati, M.H.; Song, Y.; Aubry, A.F.; Arnold, M.E. LC-MS/MS determination of apixaban (BMS-562247) and its major metabolite in human plasma: An application of polarity switching and monolithic HPLC column. Bioanalysis 2014, 6, 2071–2082. [Google Scholar] [CrossRef] [PubMed]
- Aljohani, M.M.; Chinnappan, R.; Eissa, S.; Alsager, O.A.; Weber, K.; Cialla-May, D.; Popp, J.; Zourob, M. In Vitro Selection of Specific DNA Aptamers Against the Anti-Coagulant Dabigatran Etexilate. Sci. Rep. 2018, 8, 13290. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aptamer | Type | Target | Developer | Evaluation Stages | Reference |
|---|---|---|---|---|---|
| ARC-1779 | DNA | Von Willebrand Factor | Archemix | Phase II | [23] |
| Rn-DsDsDs-44 | Unnatural DNA | Von Willebrand Factor | TagCyx Biotechnologies, RIKEN Center for Life Science Technologies | Pre-clinical | [18] |
| DTRI-031 | RNA * | Von Willebrand Factor | Duke Medical Center | Pre-clinical | [24] |
| NU172 | DNA | IIa | ARCA Biopharm | Phase II | [25,26] |
| HD22 | DNA | IIa | University of Bonn | Pre-clinical | [27] |
| HD1-22 | DNA | IIa | University of Bonn | Pre-clinical | [27] |
| RE31 | Modified DNA | IIa | Polish Academy of Sciences | Pre-clinical | [28] |
| M08 | DNA | IIa | The University of Tokyo | Pre-clinical | [29] |
| ThAD | DNA | IIa | Arizona State University | Pre-clinical | [30] |
| R9d14t | RNA * | II/ IIa | Duke Medical Center | Pre-clinical | [31] |
| REG1 | RNA * | IXa | Regado Biosciences | Phase III | [32,33,34] |
| REG2 | RNA * | IXa | Regado Biosciences | Phase I | [35] |
| 11F7t | RNA * | Xa | Duke Medical Center | Pre-clinical | [36] |
| FELIAP | DNA | XIa | McMaster University, Canadian Blood Services Thrombosis and Atherosclerosis Research | Pre-clinical | [37] |
| 11.16 | RNA * | XIa | Duke Medical Center | Pre-clinical | [38] |
| 12.7 | RNA * | XIa | Duke Medical Center | Pre-clinical | [38] |
| R4cXII-1 | RNA * | XII/XIIa | Duke Medica Center | Pre-clinical | [39] |
| Kall1-T4 | RNA * | Kallikrein | Duke Medical Center | Pre-clinical | [40] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ponce, A.T.; Hong, K.L. A Mini-Review: Clinical Development and Potential of Aptamers for Thrombotic Events Treatment and Monitoring. Biomedicines 2019, 7, 55. https://doi.org/10.3390/biomedicines7030055
Ponce AT, Hong KL. A Mini-Review: Clinical Development and Potential of Aptamers for Thrombotic Events Treatment and Monitoring. Biomedicines. 2019; 7(3):55. https://doi.org/10.3390/biomedicines7030055
Chicago/Turabian StylePonce, Alex T., and Ka Lok Hong. 2019. "A Mini-Review: Clinical Development and Potential of Aptamers for Thrombotic Events Treatment and Monitoring" Biomedicines 7, no. 3: 55. https://doi.org/10.3390/biomedicines7030055
APA StylePonce, A. T., & Hong, K. L. (2019). A Mini-Review: Clinical Development and Potential of Aptamers for Thrombotic Events Treatment and Monitoring. Biomedicines, 7(3), 55. https://doi.org/10.3390/biomedicines7030055