Can Blood-Circulating Factors Unveil and Delay Your Biological Aging?


Abstract
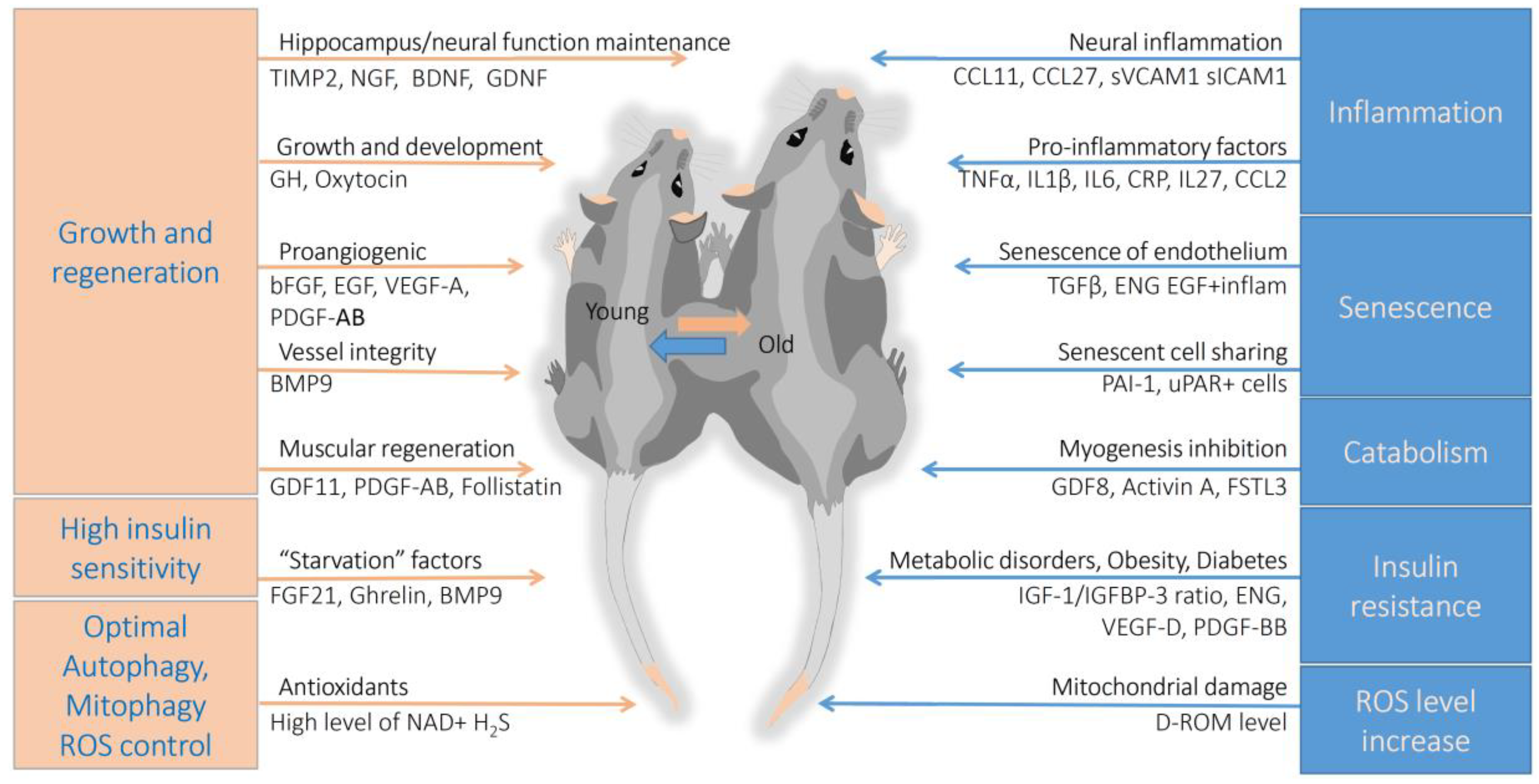
:1. Introduction
2. Age-Related Changes in Plasma Biochemistry and Vascular Dynamics
2.1. Lipids
2.2. NAD+/NADH Index
2.3. ROS
2.4. H2S
2.5. β2-Microglobulin
3. Circulating Hormones and Growth Factors Associated with Aging
3.1. TGF-β Superfamily
3.2. IGF-1
3.3. NGF and Neural Regeneration
3.4. PDGF/VEGF Vascular Remodeling
3.5. FGF21
3.6. Oxytocin
3.7. Growth Hormone
4. Age-Associated Inflammatory Factors
4.1. Well-Known Pro-Inflammatory Factors
4.2. CCL2
4.3. CCL11
4.4. CCL27
4.5. IL27, IL35, and TNF Interaction
4.6. Soluble VCAM1 and ICAM1
4.7. vWF
4.8. TIMP2-Anti-Inflammaging
4.9. Soluble uPAR and PAI-1
5. Conclusions
5.1. Vascular and Neural System Aging
5.2. Systemic Inflammaging
5.3. Regeneration and Metabolic Disorders
5.4. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Blüher, M.B.; Kahn, B.; Kahn, C.R. Extended longevity in mice lacking the insulin receptor in adipose tissue. Science 2003, 299, 572–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yegorov, Y.E.; Poznyak, A.V.; Nikiforov, N.G.; Sobenin, I.A.; Orekhov, A.N. The Link between Chronic Stress and Accelerated Aging. Biomedicines 2020, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Pyrkov, T.V.; Getmantsev, E.; Zhurov, B.; Avchaciov, K.; Pyatnitskiy, M.; Menshikov, L.; Khodova, K.; Gudkov, A.V.; Fedichev, P.O. Quantitative characterization of biological age and frailty based on locomotor activity records. Aging (Albany NY) 2018, 10, 2973–2990. [Google Scholar] [CrossRef] [PubMed]
- Berezina, T.N.; Rybtsova, N.N.; Rybtsov, S.A. Comparative Dynamics of Individual Ageing Among the Investigative Type of Professionals Living in Russia and Russian Migrants to the EU Countries. Eur. J. Investig. Health Psychol. Educ. 2020, 10, 749–762. [Google Scholar] [CrossRef]
- Bai, X. Biomarkers of Aging. Adv. Exp. Med. Biol. 2018, 1086, 217–234. [Google Scholar] [CrossRef] [PubMed]
- Hamczyk, M.R.; Nevado, R.M.; Barettino, A.; Fuster, V.; Andrés, V. Biological Versus Chronological Aging: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 919–930. [Google Scholar] [CrossRef]
- Gasmi, M.; Sellami, M.; Denham, J.; Padulo, J.; Kuvacic, G.; Selmi, W.; Khalifa, R. Time-restricted feeding influences immune responses without compromising muscle performance in older men. Nutrition 2018, 51–52, 29–37. [Google Scholar] [CrossRef]
- Hoshino, S.; Kobayashi, M.; Higami, Y. Mechanisms of the anti-aging and prolongevity effects of caloric restriction: Evidence from studies of genetically modified animals. Aging (Albany NY) 2018, 10, 2243–2251. [Google Scholar] [CrossRef]
- Berezina, T.N.; Buzanov, K.E.; Zinatullina, A.M.; Kalaeva, A.A.; Melnik, V.P. The expectation of retirement as a psychological stress that affects the biological age in the person of the Russian Federation. Relig. Rev. Cienc. Soc. Humanid. 2019, 4, 192–198. [Google Scholar]
- Bunning, B.J.; Contrepois, K.; Lee-McMullen, B.; Dhondalay, G.K.R.; Zhang, W.; Tupa, D.; Raeber, O.; Desai, M.; Nadeau, K.C.; Snyder, M.P.; et al. Global metabolic profiling to model biological processes of aging in twins. Aging Cell 2020, 19, e13073. [Google Scholar] [CrossRef] [Green Version]
- Piber, D.; Olmstead, R.; Cho, J.H.J.; Witarama, T.; Perez, C.; Dietz, N.; Seeman, T.E.; Breen, E.C.; Cole, S.W.; Irwin, M.R. Inflammaging: Age and Systemic, Cellular, and Nuclear Inflammatory Biology in Older Adults. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2019, 74, 1716–1724. [Google Scholar] [CrossRef] [PubMed]
- Torres, W.; Nava, M.; Galbán, N.; Gómez, Y.; Morillo, V.; Rojas, M.; Cano, C.; Chacín, M.; Luis, D.M.; Herazo, Y.; et al. Anti-Aging Effect of Metformin: A Molecular and Therapeutical Perspective. Curr. Pharm. Des. 2020, 26, 4496–4508. [Google Scholar] [CrossRef] [PubMed]
- Zhavoronkov, A. Geroprotective and senoremediative strategies to reduce the comorbidity, infection rates, severity, and lethality in gerophilic and gerolavic infections. Aging (Albany NY) 2020, 12, 6492–6510. [Google Scholar] [CrossRef] [PubMed]
- Fahy, G.M.; Brooke, R.T.; Watson, J.P.; Good, Z.; Vasanawala, S.S.; Maecker, H.; Leipold, M.D.; Lin, D.T.S.; Kobor, M.S.; Horvath, S. Reversal of epigenetic aging and immunosenescent trends in humans. Aging Cell 2019, 18, e13028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrzep-Poloczek, B.; Poloczek, J.; Chelmecka, E.; Dulska, A.; Romuk, E.; Idzik, M.; Kazura, W.; Nabrdalik, K.; Gumprecht, J.; Jochem, J.; et al. The Oxidative Stress Markers in the Erythrocytes and Heart Muscle of Obese Rats: Relate to a High-Fat Diet but Not to DJOS Bariatric Surgery. Antioxidants (Basel) 2020, 9, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, S.J.; Le Couteur, D.G.; Raubenheimer, D.; Solon-Biet, S.M.; Cooney, G.J.; Cogger, V.C.; Fontana, L. Dietary protein, aging and nutritional geometry. Ageing Res. Rev. 2017, 39, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Broskey, N.T.; Marlatt, K.L.; Most, J.; Erickson, M.L.; Irving, B.A.; Redman, L.M. The Panacea of Human Aging: Calorie Restriction Versus Exercise. Exerc. Sport Sci. Rev. 2019, 47, 169–175. [Google Scholar] [CrossRef]
- Coenen, P.; Huysmans, M.A.; Holtermann, A.; Krause, N.; van Mechelen, W.; Straker, L.M.; van der Beek, A.J. Do highly physically active workers die early? A systematic review with meta-analysis of data from 193 696 participants. Br. J. Sports Med. 2018, 52, 1320–1326. [Google Scholar] [CrossRef] [Green Version]
- Madore, C.; Yin, Z.; Leibowitz, J.; Butovsky, O. Microglia, Lifestyle Stress, and Neurodegeneration. Immunity 2020, 52, 222–240. [Google Scholar] [CrossRef]
- Barcena, C.; Valdes-Mas, R.; Mayoral, P.; Garabaya, C.; Durand, S.; Rodriguez, F.; Fernandez-Garcia, M.T.; Salazar, N.; Nogacka, A.M.; Garatachea, N.; et al. Healthspan and lifespan extension by fecal microbiota transplantation into progeroid mice. Nat. Med. 2019, 25, 1234–1242. [Google Scholar] [CrossRef]
- Biver, E.; Berenbaum, F.; Valdes, A.M.; de Carvalho, I.A.; Bindels, L.B.; Brandi, M.L.; Calder, P.C.; Castronovo, V.; Cavalier, E.; Cherubini, A.; et al. Gut microbiota and osteoarthritis management: An expert consensus of the European Society for Clinical and Economic Aspects of Osteoporosis, Osteoarthritis and Musculoskeletal Diseases (ESCEO). Ageing Res. Rev. 2019, 100946. [Google Scholar] [CrossRef] [PubMed]
- Shan, K.; Qu, H.; Zhou, K.; Wang, L.; Zhu, C.; Chen, H.; Gu, Z.; Cui, J.; Fu, G.; Li, J.; et al. Distinct Gut Microbiota Induced by Different Fat-to-Sugar-Ratio High-Energy Diets Share Similar Pro-obesity Genetic and Metabolite Profiles in Prediabetic Mice. mSystems 2019, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahadi, S.; Zhou, W.; Schüssler-Fiorenza Rose, S.M.; Sailani, M.R.; Contrepois, K.; Avina, M.; Ashland, M.; Brunet, A.; Snyder, M. Personal aging markers and ageotypes revealed by deep longitudinal profiling. Nat. Med. 2020, 26, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Hashemzehi, M.; Beheshti, F.; Hassanian, S.M.; Ferns, G.A.; Khazaei, M.; Avan, A. Therapeutic potential of renin angiotensin system inhibitors in cancer cells metastasis. Pathol. Res. Pr. 2020, 216, 153010. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Bang, E.; Arulkumar, R.; Ha, S.; Chung, K.W.; Park, M.H.; Choi, Y.J.; Yu, B.P.; Chung, H.Y. Senoinflammation: A major mediator underlying age-related metabolic dysregulation. Exp. Gerontol. 2020, 134, 110891. [Google Scholar] [CrossRef]
- Castellano, J.M. Blood-Based Therapies to Combat Aging. Gerontology 2019, 65, 84–89. [Google Scholar] [CrossRef]
- Vaiserman, A.; Koliada, A.; Lushchak, O. Developmental programming of aging trajectory. Ageing Res. Rev. 2018, 47, 105–122. [Google Scholar] [CrossRef]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging (Albany NY) 2018, 10, 573–591. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.J.; Sharma, A.; Mair, W.B. Metabolic Communication and Healthy Aging: Where Should We Focus Our Energy? Dev. Cell 2020, 54, 196–211. [Google Scholar] [CrossRef]
- Balistreri, C.R.; Garagnani, P.; Madonna, R.; Vaiserman, A.; Melino, G. Developmental programming of adult haematopoiesis system. Ageing Res. Rev. 2019, 54, 100918. [Google Scholar] [CrossRef]
- Childs, B.G.; Gluscevic, M.; Baker, D.J.; Laberge, R.M.; Marquess, D.; Dananberg, J.; Van Deursen, J.M. Senescent cells: An emerging target for diseases of ageing. Nat. Rev. Drug Discov. 2017, 16, 718–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alteber, Z.; Sharbi-Yunger, A.; Pevsner-Fischer, M.; Blat, D.; Roitman, L.; Tzehoval, E.; Elinav, E.; Eisenbach, L. The anti-inflammatory IFITM genes ameliorate colitis and partially protect from tumorigenesis by changing immunity and microbiota. Immunol. Cell Biol. 2018, 96, 284–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, H.; Chandel, N.S. To Claim Growth Turf, mTOR Says SOD Off. Mol. Cell 2018, 70, 383–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannini, N.; Campos, V.; Girotra, M.; Trachsel, V.; Rojas-Sutterlin, S.; Tratwal, J.; Ragusa, S.; Stefanidis, E.; Ryu, D.; Rainer, P.Y.; et al. The NAD-Booster Nicotinamide Riboside Potently Stimulates Hematopoiesis through Increased Mitochondrial Clearance. Cell Stem Cell 2019, 24, 405–418.e407. [Google Scholar] [CrossRef] [Green Version]
- Ho, T.T.; Warr, M.R.; Adelman, E.R.; Lansinger, O.M.; Flach, J.; Verovskaya, E.V.; Figueroa, M.E.; Passegue, E. Autophagy maintains the metabolism and function of young and old stem cells. Nature 2017, 543, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Shang, D.; Sun, D.; Shi, C.; Xu, J.; Shen, M.; Hu, X.; Liu, H.; Tu, Z. Activation of epidermal growth factor receptor signaling mediates cellular senescence induced by certain pro-inflammatory cytokines. Aging Cell 2020, 19, e13145. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. Aging: ROS or TOR. Cell Cycle 2008, 7, 3344–3354. [Google Scholar] [CrossRef]
- Chung, H.Y.; Kim, D.H.; Lee, E.K.; Chung, K.W.; Chung, S.; Lee, B.; Seo, A.Y.; Chung, J.H.; Jung, Y.S.; Im, E.; et al. Redefining Chronic Inflammation in Aging and Age-Related Diseases: Proposal of the Senoinflammation Concept. Aging Dis. 2019, 10, 367–382. [Google Scholar] [CrossRef] [Green Version]
- Hickson, L.J.; Langhi Prata, L.G.P.; Bobart, S.A.; Evans, T.K.; Giorgadze, N.; Hashmi, S.K.; Herrmann, S.M.; Jensen, M.D.; Jia, Q.; Jordan, K.L.; et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine 2019, 47, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Sikora, E.; Bielak-Zmijewska, A.; Mosieniak, G. Targeting normal and cancer senescent cells as a strategy of senotherapy. Ageing Res. Rev. 2019, 100941. [Google Scholar] [CrossRef]
- Xu, M.; Pirtskhalava, T.; Farr, J.N.; Weigand, B.M.; Palmer, A.K.; Weivoda, M.M.; Inman, C.L.; Ogrodnik, M.B.; Hachfeld, C.M.; Fraser, D.G.; et al. Senolytics improve physical function and increase lifespan in old age. Nat. Med. 2018, 24, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Kirkland, J.L.; Tchkonia, T. Senolytic drugs: From discovery to translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef] [PubMed]
- Bharath, L.P.; Agrawal, M.; McCambridge, G.; Nicholas, D.A.; Hasturk, H.; Liu, J.; Jiang, K.; Liu, R.; Guo, Z.; Deeney, J.; et al. Metformin Enhances Autophagy and Normalizes Mitochondrial Function to Alleviate Aging-Associated Inflammation. Cell Metab. 2020. [Google Scholar] [CrossRef] [PubMed]
- Fielder, E.; Tweedy, C.; Wilson, C.; Oakley, F.; LeBeau, F.E.N.; Passos, J.F.; Mann, D.A.; von Zglinicki, T.; Jurk, D. Anti-inflammatory treatment rescues memory deficits during aging in nfkb1(-/-) mice. Aging Cell 2020, 19, e13188. [Google Scholar] [CrossRef]
- Young, K.; Eudy, E.; Bell, R.; Loberg, M.; Stearns, T.; Velten, L.; Haas, S.; Trowbridge, J.J. Hematopoietic Stem and Progenitor Cell Aging is Initiated at Middle Age Through Decline in Local Insulin-Like Growth Factor 1 (IGF1). bioRxiv 2020. [Google Scholar] [CrossRef]
- Xu, W.; Lau, Z.W.X.; Fulop, T.; Larbi, A. The Aging of γδ T Cells. Cells 2020, 9, 1181. [Google Scholar] [CrossRef]
- Yousef, H.; Czupalla, C.J.; Lee, D.; Chen, M.B.; Burke, A.N.; Zera, K.A.; Zandstra, J.; Berber, E.; Lehallier, B.; Mathur, V.; et al. Aged blood impairs hippocampal neural precursor activity and activates microglia via brain endothelial cell VCAM1. Nat. Med. 2019, 25, 988–1000. [Google Scholar] [CrossRef]
- Fang, Y.; Zhu, L.; An, N.; Jiang, G.; Qian, J.; Zhao, R.; Yuan, N.; Zhang, S.; Wang, J. Blood autophagy defect causes accelerated non-hematopoietic organ aging. Aging (Albany NY) 2019, 11, 4910–4922. [Google Scholar] [CrossRef]
- Oh, S.J.; Lee, J.K.; Shin, O.S. Aging and the Immune System: The Impact of Immunosenescence on Viral Infection, Immunity and Vaccine Immunogenicity. Immune Netw. 2019, 19, e37. [Google Scholar] [CrossRef]
- Ovadya, Y.; Landsberger, T.; Leins, H.; Vadai, E.; Gal, H.; Biran, A.; Yosef, R.; Sagiv, A.; Agrawal, A.; Shapira, A.; et al. Impaired immune surveillance accelerates accumulation of senescent cells and aging. Nat. Commun. 2018, 9, 5435. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Liu, X.; Le, W.; Xie, L.; Li, H.; Wen, W.; Wang, S.; Ma, S.; Huang, Z.; Ye, J.; et al. A human circulating immune cell landscape in aging and COVID-19. Protein Cell 2020, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Kiss, T.; Tarantini, S.; Csipo, T.; Balasubramanian, P.; Nyúl-Tóth, Á.; Yabluchanskiy, A.; Wren, J.D.; Garman, L.; Huffman, D.M.; Csiszar, A.; et al. Circulating anti-geronic factors from heterochonic parabionts promote vascular rejuvenation in aged mice: Transcriptional footprint of mitochondrial protection, attenuation of oxidative stress, and rescue of endothelial function by young blood. Geroscience 2020, 42, 727–748. [Google Scholar] [CrossRef] [PubMed]
- Boerman, E.M.; Segal, S.S. Depressed perivascular sensory innervation of mouse mesenteric arteries with advanced age. J. Physiol. 2016, 594, 2323–2338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrose, C.T. Pro-Angiogenesis Therapy and Aging: A Mini-Review. Gerontology 2017, 63, 393–400. [Google Scholar] [CrossRef] [Green Version]
- Carallo, C.; Irace, C.; De Franceschi, M.S.; Coppoletta, F.; Tiriolo, R.; Scicchitano, C.; Scavelli, F.; Gnasso, A. The effect of aging on blood and plasma viscosity. An 11.6 years follow-up study. Clin. Hemorheol. Microcirc. 2011, 47, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Coppola, L.; Caserta, F.; De Lucia, D.; Guastafierro, S.; Grassia, A.; Coppola, A.; Marfella, R.; Varricchio, M. Blood viscosity and aging. Arch. Gerontol. Geriatr. 2000, 31, 35–42. [Google Scholar] [CrossRef]
- Moeller, M.; Pink, C.; Endlich, N.; Endlich, K.; Grabe, H.J.; Völzke, H.; Dörr, M.; Nauck, M.; Lerch, M.M.; Köhling, R.; et al. Mortality is associated with inflammation, anemia, specific diseases and treatments, and molecular markers. PLoS ONE 2017, 12, e0175909. [Google Scholar] [CrossRef]
- Matsue, Y.; van der Meer, P.; Damman, K.; Metra, M.; O’Connor, C.M.; Ponikowski, P.; Teerlink, J.R.; Cotter, G.; Davison, B.; Cleland, J.G.; et al. Blood urea nitrogen-to-creatinine ratio in the general population and in patients with acute heart failure. Heart 2017, 103, 407–413. [Google Scholar] [CrossRef]
- Kooman, J.P.; Kotanko, P.; Schols, A.M.; Shiels, P.G.; Stenvinkel, P. Chronic kidney disease and premature ageing. Nat. Rev. Nephrol. 2014, 10, 732–742. [Google Scholar] [CrossRef]
- Dzaye, O.; Al Rifai, M.; Dardari, Z.; Shaw, L.J.; Al-Mallah, M.H.; Handy Marshall, C.; Rozanski, A.; Mortensen, M.B.; Duebgen, M.; Matsushita, K.; et al. Coronary Artery Calcium as a Synergistic Tool for the Age- and Sex-Specific Risk of Cardiovascular and Cancer Mortality: The Coronary Artery Calcium Consortium. J. Am. Heart Assoc. 2020, 9, e015306. [Google Scholar] [CrossRef]
- Mahlknecht, U.; Kaiser, S. Age-related changes in peripheral blood counts in humans. Exp. Med. 2010, 1, 1019–1025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.W.K.; Braidy, N.; Pickford, R.; Vafaee, F.; Crawford, J.; Muenchhoff, J.; Schofield, P.; Attia, J.; Brodaty, H.; Sachdev, P.; et al. Plasma lipidome variation during the second half of the human lifespan is associated with age and sex but minimally with BMI. PLoS ONE 2019, 14, e0214141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laks, R.; Araújo, L.M.; Almada Filho Cde, M.; Cendoroglo, M.S. The importance of HDL-C and CRP in cardiovascular risk evaluation in longevous elderly individuals. Einstein (Sao Paulo) 2011, 9, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Shi, L.; Zou, Y.; Tang, J.; Cai, J.; Wei, Y.; Qin, J.; Zhang, Z. Positive association of familial longevity with the moderate-high HDL-C concentration in Bama Aging Study. Aging (Albany NY) 2018, 10, 3528–3540. [Google Scholar] [CrossRef]
- Stone, N.J.; Robinson, J.G.; Lichtenstein, A.H.; Bairey Merz, C.N.; Blum, C.B.; Eckel, R.H.; Goldberg, A.C.; Gordon, D.; Levy, D.; Lloyd-Jones, D.M.; et al. 2013 ACC/AHA guideline on the treatment of blood cholesterol to reduce atherosclerotic cardiovascular risk in adults: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. Circulation 2014, 129, S1–S45. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Covarrubias, V.; Beekman, M.; Uh, H.W.; Dane, A.; Troost, J.; Paliukhovich, I.; van der Kloet, F.M.; Houwing-Duistermaat, J.; Vreeken, R.J.; Hankemeier, T.; et al. Lipidomics of familial longevity. Aging Cell 2013, 12, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Deelen, J.; Kettunen, J.; Fischer, K.; van der Spek, A.; Trompet, S.; Kastenmüller, G.; Boyd, A.; Zierer, J.; van den Akker, E.B.; Ala-Korpela, M.; et al. A metabolic profile of all-cause mortality risk identified in an observational study of 44,168 individuals. Nat. Commun. 2019, 10, 3346. [Google Scholar] [CrossRef] [Green Version]
- Tabassum, R.; Rämö, J.T.; Ripatti, P.; Koskela, J.T.; Kurki, M.; Karjalainen, J.; Palta, P.; Hassan, S.; Nunez-Fontarnau, J.; Kiiskinen, T.T.J.; et al. Genetic architecture of human plasma lipidome and its link to cardiovascular disease. Nat. Commun. 2019, 10, 4329. [Google Scholar] [CrossRef] [Green Version]
- Mills, K.F.; Yoshida, S.; Stein, L.R.; Grozio, A.; Kubota, S.; Sasaki, Y.; Redpath, P.; Migaud, M.E.; Apte, R.S.; Uchida, K.; et al. Long-Term Administration of Nicotinamide Mononucleotide Mitigates Age-Associated Physiological Decline in Mice. Cell Metab. 2016, 24, 795–806. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, J.; Baur, J.A.; Imai, S.I. NAD(+) Intermediates: The Biology and Therapeutic Potential of NMN and NR. Cell Metab. 2018, 27, 513–528. [Google Scholar] [CrossRef] [Green Version]
- Seyedsadjadi, N.; Berg, J.; Bilgin, A.A.; Braidy, N.; Salonikas, C.; Grant, R. High protein intake is associated with low plasma NAD+ levels in a healthy human cohort. PLoS ONE 2018, 13, e0201968. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Pereira, J.; Tarragó, M.G.; Chini, C.C.S.; Nin, V.; Escande, C.; Warner, G.M.; Puranik, A.S.; Schoon, R.A.; Reid, J.M.; Galina, A.; et al. CD38 Dictates Age-Related NAD Decline and Mitochondrial Dysfunction through an SIRT3-Dependent Mechanism. Cell Metab. 2016, 23, 1127–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarragó, M.G.; Chini, C.C.S.; Kanamori, K.S.; Warner, G.M.; Caride, A.; de Oliveira, G.C.; Rud, M.; Samani, A.; Hein, K.Z.; Huang, R.; et al. A Potent and Specific CD38 Inhibitor Ameliorates Age-Related Metabolic Dysfunction by Reversing Tissue NAD(+) Decline. Cell Metab. 2018, 27, 1081–1095.e1010. [Google Scholar] [CrossRef] [Green Version]
- Schottker, B.; Brenner, H.; Jansen, E.H.; Gardiner, J.; Peasey, A.; Kubinova, R.; Pajak, A.; Topor-Madry, R.; Tamosiunas, A.; Saum, K.U.; et al. Evidence for the free radical/oxidative stress theory of ageing from the CHANCES consortium: A meta-analysis of individual participant data. BMC Med. 2015, 13, 300. [Google Scholar] [CrossRef] [Green Version]
- Xuan, Y.; Gao, X.; Anusruti, A.; Holleczek, B.; Jansen, E.; Muhlack, D.C.; Brenner, H.; Schottker, B. Association of Serum Markers of Oxidative Stress With Incident Major Cardiovascular Events, Cancer Incidence, and All-Cause Mortality in Type 2 Diabetes Patients: Pooled Results From Two Cohort Studies. Diabetes Care 2019, 42, 1436–1445. [Google Scholar] [CrossRef]
- Wei, B.; Willems, P.; Huang, J.; Tian, C.; Yang, J.; Messens, J.; Van Breusegem, F. Identification of Sulfenylated Cysteines in Arabidopsis thaliana Proteins Using a Disulfide-Linked Peptide Reporter. Front. Plant Sci. 2020, 11, 777. [Google Scholar] [CrossRef]
- Heppner, D.E.; Hristova, M.; Ida, T.; Mijuskovic, A.; Dustin, C.M.; Bogdándi, V.; Fukuto, J.M.; Dick, T.P.; Nagy, P.; Li, J.; et al. Cysteine perthiosulfenic acid (Cys-SSOH): A novel intermediate in thiol-based redox signaling? Redox Biol. 2018, 14, 379–385. [Google Scholar] [CrossRef]
- Silva-Palacios, A.; Ostolga-Chavarria, M.; Zazueta, C.; Konigsberg, M. Nrf2: Molecular and epigenetic regulation during aging. Ageing Res. Rev. 2018, 47, 31–40. [Google Scholar] [CrossRef]
- Rangasamy, T.; Cho, C.Y.; Thimmulappa, R.K.; Zhen, L.; Srisuma, S.S.; Kensler, T.W.; Yamamoto, M.; Petrache, I.; Tuder, R.M.; Biswal, S. Genetic ablation of Nrf2 enhances susceptibility to cigarette smoke-induced emphysema in mice. J. Clin. Investig. 2004, 114, 1248–1259. [Google Scholar] [CrossRef]
- Fao, L.; Mota, S.I.; Rego, A.C. Shaping the Nrf2-ARE-related pathways in Alzheimer’s and Parkinson’s diseases. Ageing Res. Rev. 2019, 54, 100942. [Google Scholar] [CrossRef]
- Huang, K.; Gao, X.; Wei, W. The crosstalk between Sirt1 and Keap1/Nrf2/ARE anti-oxidative pathway forms a positive feedback loop to inhibit FN and TGF-β1 expressions in rat glomerular mesangial cells. Exp. Cell Res. 2017, 361, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Zivanovic, J.; Kouroussis, E.; Kohl, J.B.; Adhikari, B.; Bursac, B.; Schott-Roux, S.; Petrovic, D.; Miljkovic, J.L.; Thomas-Lopez, D.; Jung, Y.; et al. Selective Persulfide Detection Reveals Evolutionarily Conserved Antiaging Effects of S-Sulfhydration. Cell Metab. 2019, 30, 1152–1170.e1113. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qabazard, B.; Ahmed, S.; Li, L.; Arlt, V.M.; Moore, P.K.; Sturzenbaum, S.R.C. elegans aging is modulated by hydrogen sulfide and the sulfhydrylase/cysteine synthase cysl-2. PLoS ONE 2013, 8, e80135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perridon, B.W.; Leuvenink, H.G.; Hillebrands, J.L.; van Goor, H.; Bos, E.M. The role of hydrogen sulfide in aging and age-related pathologies. Aging (Albany NY) 2016, 8, 2264–2289. [Google Scholar] [CrossRef] [Green Version]
- Karunya, R.; Jayaprakash, K.S.; Gaikwad, R.; Sajeesh, P.; Ramshad, K.; Muraleedharan, K.M.; Dixit, M.; Thangaraj, P.R.; Sen, A.K. Rapid measurement of hydrogen sulphide in human blood plasma using a microfluidic method. Sci. Rep. 2019, 9, 3258. [Google Scholar] [CrossRef]
- Shilova, V.; Zatsepina, O.; Zakluta, A.; Karpov, D.; Chuvakova, L.; Garbuz, D.; Evgen’ev, M. Age-dependent expression profiles of two adaptogenic systems and thermotolerance in Drosophila melanogaster. Cell Stress Chaperones 2020, 25, 305–315. [Google Scholar] [CrossRef]
- Zatsepina, O.; Karpov, D.; Chuvakova, L.; Rezvykh, A.; Funikov, S.; Sorokina, S.; Zakluta, A.; Garbuz, D.; Shilova, V.; Evgen’ev, M. Genome-wide transcriptional effects of deletions of sulphur metabolism genes in Drosophila melanogaster. Redox Biol. 2020, 36, 101654. [Google Scholar] [CrossRef]
- Grossi, V.; Forte, G.; Sanese, P.; Peserico, A.; Tezil, T.; Lepore Signorile, M.; Fasano, C.; Lovaglio, R.; Bagnulo, R.; Loconte, D.C.; et al. The longevity SNP rs2802292 uncovered: HSF1 activates stress-dependent expression of FOXO3 through an intronic enhancer. Nucleic Acids Res. 2018, 46, 5587–5600. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Polhemus, D.J.; Lefer, D.J. Evolution of Hydrogen Sulfide Therapeutics to Treat Cardiovascular Disease. Circ. Res. 2018, 123, 590–600. [Google Scholar] [CrossRef]
- Nasi, S.; Ehirchiou, D.; Chatzianastasiou, A.; Nagahara, N.; Papapetropoulos, A.; Bertrand, J.; Cirino, G.; So, A.; Busso, N. The protective role of the 3-mercaptopyruvate sulfurtransferase (3-MST)-hydrogen sulfide (H2S) pathway against experimental osteoarthritis. Arthritis Res. Ther. 2020, 22, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hine, C.; Harputlugil, E.; Zhang, Y.; Ruckenstuhl, C.; Lee, B.C.; Brace, L.; Longchamp, A.; Trevino-Villarreal, J.H.; Mejia, P.; Ozaki, C.K.; et al. Endogenous hydrogen sulfide production is essential for dietary restriction benefits. Cell 2015, 160, 132–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qabazard, B.; Sturzenbaum, S.R. H2S: A New Approach to Lifespan Enhancement and Healthy Ageing? Handb Exp Pharm. 2015, 230, 269–287. [Google Scholar] [CrossRef]
- Cohen, G.F.; Vyas, N.S. Sodium thiosulfate in the treatment of calciphylaxis. J. Clin. Aesthetic Derm. 2013, 6, 41–44. [Google Scholar]
- Li, L.; Dong, M.; Wang, X.G. The Implication and Significance of Beta 2 Microglobulin: A Conservative Multifunctional Regulator. Chin. Med. J. (Engl.) 2016, 129, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.K.; He, Y.; Park, J.-S.; Bieri, G.; Snethlage, C.E.; Lin, K.; Gontier, G.; Wabl, R.; Plambeck, K.E.; Udeochu, J.; et al. β2-microglobulin is a systemic pro-aging factor that impairs cognitive function and neurogenesis. Nat. Med. 2015, 21, 932–937. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Fu, S.; Zhao, L.; Zhen, B.; Ye, L.; Niu, X.; Li, X.; Zhang, P.; Bai, J. Quantitation of circulating GDF-11 and β2-MG in aged patients with age-related impairment in cognitive function. Clin. Sci. (Lond.) 2017, 131, 1895–1904. [Google Scholar] [CrossRef] [Green Version]
- Iadanza, M.G.; Jackson, M.P.; Hewitt, E.W.; Ranson, N.A.; Radford, S.E. A new era for understanding amyloid structures and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 755–773. [Google Scholar] [CrossRef]
- Filler, G.; Priem, F.; Lepage, N.; Sinha, P.; Vollmer, I.; Clark, H.; Keely, E.; Matzinger, M.; Akbari, A.; Althaus, H.; et al. Beta-trace protein, cystatin C, beta(2)-microglobulin, and creatinine compared for detecting impaired glomerular filtration rates in children. Clin. Chem. 2002, 48, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Foster, M.C.; Inker, L.A.; Levey, A.S.; Selvin, E.; Eckfeldt, J.; Juraschek, S.P.; Coresh, J. Novel filtration markers as predictors of all-cause and cardiovascular mortality in US adults. Am. J. Kidney Dis. 2013, 62, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.Y.; Shen, Y.Y.; Ji, L.J.; Jiang, X.Y.; Wang, X.F.; Shi, Y. Association between serum β2-microglobulin levels and frailty in an elderly Chinese population: Results from RuLAS. Clin. Interv. Aging 2017, 12, 1725–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, R.; Peel, N.M.; Krosch, M.; Hubbard, R.E. Frailty and chronic kidney disease: A systematic review. Arch. Gerontol. Geriatr. 2017, 68, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludwig, F.C.; Elashoff, R.M. Mortality in syngeneic rat parabionts of different chronological age. Trans. N. Y. Acad. Sci. 1972, 34, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Kamran, P.; Sereti, K.I.; Zhao, P.; Ali, S.R.; Weissman, I.L.; Ardehali, R. Parabiosis in mice: A detailed protocol. J. Vis. Exp. 2013. [Google Scholar] [CrossRef] [Green Version]
- Kheifets, V.; Braithwaite, S.P. Plasma-Based Strategies for Therapeutic Modulation of Brain Aging. Neurotherapeutics 2019, 16, 675–684. [Google Scholar] [CrossRef]
- Wyss-Coray, T. Ageing, neurodegeneration and brain rejuvenation. Nature 2016, 539, 180–186. [Google Scholar] [CrossRef]
- Chen, M.B.; Yang, A.C.; Yousef, H.; Lee, D.; Chen, W.; Schaum, N.; Lehallier, B.; Quake, S.R.; Wyss-Coray, T. Brain Endothelial Cells Are Exquisite Sensors of Age-Related Circulatory Cues. Cell Rep. 2020, 30, 4418–4432.e4414. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Xu, L.; Brunet, A. Turning back time with emerging rejuvenation strategies. Nat. Cell Biol. 2019, 21, 32–43. [Google Scholar] [CrossRef]
- Villeda, S.A.; Plambeck, K.E.; Middeldorp, J.; Castellano, J.M.; Mosher, K.I.; Luo, J.; Smith, L.K.; Bieri, G.; Lin, K.; Berdnik, D.; et al. Young blood reverses age-related impairments in cognitive function and synaptic plasticity in mice. Nat. Med. 2014, 20, 659–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehdipour, M.; Etienne, J.; Chen, C.C.; Gathwala, R.; Rehman, M.; Kato, C.; Liu, C.; Liu, Y.; Zuo, Y.; Conboy, M.J.; et al. Rejuvenation of brain, liver and muscle by simultaneous pharmacological modulation of two signaling determinants, that change in opposite directions with age. Aging (Albany NY) 2019, 11, 5628–5645. [Google Scholar] [CrossRef] [PubMed]
- Rybtsov, S.A.; Dumacheva, E.V.; Dumachev, D.V.; Mustafin, I.G.; Kagansky, A.M.; Rybtsova, N.N.; Zhdanov, R.I. Ethical and scientific aspects of human embryonic material research: The Great Britain regulations. Genes Cells 2016, 11, 82–89. [Google Scholar]
- Rebo, J.; Mehdipour, M.; Gathwala, R.; Causey, K.; Liu, Y.; Conboy, M.J.; Conboy, I.M. A single heterochronic blood exchange reveals rapid inhibition of multiple tissues by old blood. Nat. Commun. 2016, 7, 13363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannatà, A.; Marcon, G.; Cimmino, G.; Camparini, L.; Ciucci, G.; Sinagra, G.; Loffredo, F.S. Role of circulating factors in cardiac aging. J. Thorac. Dis. 2017, 9, S17–S29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiepen, C.; Mendez, P.L.; Knaus, P. It Takes Two to Tango: Endothelial TGFβ/BMP Signaling Crosstalk with Mechanobiology. Cells 2020, 9, 1965. [Google Scholar] [CrossRef]
- Wang, Y.; Ma, C.; Sun, T.; Ren, L. Potential roles of bone morphogenetic protein-9 in glucose and lipid homeostasis. J. Physiol. Biochem. 2020. [Google Scholar] [CrossRef]
- Chen, W.; Ten Dijke, P. Immunoregulation by members of the TGFβ superfamily. Nat. Rev. Immunol. 2016, 16, 723–740. [Google Scholar] [CrossRef]
- Yousef, H.; Conboy, M.J.; Morgenthaler, A.; Schlesinger, C.; Bugaj, L.; Paliwal, P.; Greer, C.; Conboy, I.M.; Schaffer, D. Systemic attenuation of the TGF-β pathway by a single drug simultaneously rejuvenates hippocampal neurogenesis and myogenesis in the same old mammal. Oncotarget 2015, 6, 11959–11978. [Google Scholar] [CrossRef] [Green Version]
- Roh, J.D.; Hobson, R.; Chaudhari, V.; Quintero, P.; Yeri, A.; Benson, M.; Xiao, C.; Zlotoff, D.; Bezzerides, V.; Houstis, N.; et al. Activin type II receptor signaling in cardiac aging and heart failure. Sci. Transl. Med. 2019, 11. [Google Scholar] [CrossRef]
- Verhamme, F.M.; Bracke, K.R.; Amatngalim, G.D.; Verleden, G.M.; Van Pottelberge, G.R.; Hiemstra, P.S.; Joos, G.F.; Brusselle, G.G. Role of activin-A in cigarette smoke-induced inflammation and COPD. Eur. Respir. J. 2014, 43, 1028–1041. [Google Scholar] [CrossRef] [Green Version]
- Rochette, L.; Zeller, M.; Cottin, Y.; Vergely, C. Growth and differentiation factor 11 (GDF11): Functions in the regulation of erythropoiesis and cardiac regeneration. Pharmacol. Ther. 2015, 156, 26–33. [Google Scholar] [CrossRef]
- Tang, N.; Rao, S.; Ying, Y.; Huang, Y. New insights into BMP9 signaling in organ fibrosis. Eur. J. Pharm. 2020, 882, 173291. [Google Scholar] [CrossRef] [PubMed]
- Schafer, M.J.; Atkinson, E.J.; Vanderboom, P.M.; Kotajarvi, B.; White, T.A.; Moore, M.M.; Bruce, C.J.; Greason, K.L.; Suri, R.M.; Khosla, S.; et al. Quantification of GDF11 and Myostatin in Human Aging and Cardiovascular Disease. Cell Metab. 2016, 23, 1207–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morissette, M.R.; Stricker, J.C.; Rosenberg, M.A.; Buranasombati, C.; Levitan, E.B.; Mittleman, M.A.; Rosenzweig, A. Effects of myostatin deletion in aging mice. Aging Cell 2009, 8, 573–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jackson, M.F.; Luong, D.; Vang, D.D.; Garikipati, D.K.; Stanton, J.B.; Nelson, O.L.; Rodgers, B.D. The aging myostatin null phenotype: Reduced adiposity, cardiac hypertrophy, enhanced cardiac stress response, and sexual dimorphism. J. Endocrinol. 2012, 213, 263–275. [Google Scholar] [CrossRef] [Green Version]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- Rose, F.F., Jr.; Mattis, V.B.; Rindt, H.; Lorson, C.L. Delivery of recombinant follistatin lessens disease severity in a mouse model of spinal muscular atrophy. Hum. Mol. Genet. 2009, 18, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Ozek, C.; Krolewski, R.C.; Buchanan, S.M.; Rubin, L.L. Growth Differentiation Factor 11 treatment leads to neuronal and vascular improvements in the hippocampus of aged mice. Sci. Rep. 2018, 8, 17293. [Google Scholar] [CrossRef] [Green Version]
- Mei, W.; Xiang, G.; Li, Y.; Li, H.; Xiang, L.; Lu, J.; Dong, J.; Liu, M. GDF11 Protects against Endothelial Injury and Reduces Atherosclerotic Lesion Formation in Apolipoprotein E-Null Mice. Mol. Ther. 2016, 24, 1926–1938. [Google Scholar] [CrossRef] [Green Version]
- Idkowiak-Baldys, J.; Santhanam, U.; Buchanan, S.M.; Pfaff, K.L.; Rubin, L.L.; Lyga, J. Growth differentiation factor 11 (GDF11) has pronounced effects on skin biology. PLoS ONE 2019, 14, e0218035. [Google Scholar] [CrossRef] [Green Version]
- Harper, S.C.; Brack, A.; MacDonnell, S.; Franti, M.; Olwin, B.B.; Bailey, B.A.; Rudnicki, M.A.; Houser, S.R. Is Growth Differentiation Factor 11 a Realistic Therapeutic for Aging-Dependent Muscle Defects? Circ. Res. 2016, 118, 1143–1150, discussion 1150. [Google Scholar] [CrossRef] [Green Version]
- Semba, R.D.; Zhang, P.; Zhu, M.; Fabbri, E.; Gonzalez-Freire, M.; Moaddel, R.; Geng-Spyropoulos, M.; Ferrucci, L. A targeted proteomic assay for the measurement of plasma proteoforms related to human aging phenotypes. Proteomics 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Loffredo, F.S.; Steinhauser, M.L.; Jay, S.M.; Gannon, J.; Pancoast, J.R.; Yalamanchi, P.; Sinha, M.; Dall’Osso, C.; Khong, D.; Shadrach, J.L.; et al. Growth differentiation factor 11 is a circulating factor that reverses age-related cardiac hypertrophy. Cell 2013, 153, 828–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.E.; Cadena, S.M.; Gong, C.; Wang, X.; Chen, Z.; Wang, S.X.; Vickers, C.; Chen, H.; Lach-Trifilieff, E.; Hadcock, J.R.; et al. Supraphysiologic Administration of GDF11 Induces Cachexia in Part by Upregulating GDF15. Cell Rep. 2018, 22, 1522–1530. [Google Scholar] [CrossRef] [Green Version]
- Suragani, R.N.V.S.; Cadena, S.M.; Cawley, S.M.; Sako, D.; Mitchell, D.; Li, R.; Davies, M.V.; Alexander, M.J.; Devine, M.; Loveday, K.S.; et al. Transforming growth factor-β superfamily ligand trap ACE-536 corrects anemia by promoting late-stage erythropoiesis. Nat. Med. 2014, 20, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Latres, E.; Mastaitis, J.; Fury, W.; Miloscio, L.; Trejos, J.; Pangilinan, J.; Okamoto, H.; Cavino, K.; Na, E.; Papatheodorou, A.; et al. Activin A more prominently regulates muscle mass in primates than does GDF8. Nat. Commun. 2017, 8, 15153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.J.; Reed, L.A.; Davies, M.V.; Girgenrath, S.; Goad, M.E.; Tomkinson, K.N.; Wright, J.F.; Barker, C.; Ehrmantraut, G.; Holmstrom, J.; et al. Regulation of muscle growth by multiple ligands signaling through activin type II receptors. Proc. Natl. Acad. Sci. USA 2005, 102, 18117–18122. [Google Scholar] [CrossRef] [Green Version]
- Walton, K.L.; Makanji, Y.; Harrison, C.A. New insights into the mechanisms of activin action and inhibition. Mol. Cell Endocrinol. 2012, 359, 2–12. [Google Scholar] [CrossRef]
- Walker, R.G.; Czepnik, M.; Goebel, E.J.; McCoy, J.C.; Vujic, A.; Cho, M.; Oh, J.; Aykul, S.; Walton, K.L.; Schang, G.; et al. Structural basis for potency differences between GDF8 and GDF11. BMC Biol. 2017, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Magga, J.; Vainio, L.; Kilpio, T.; Hulmi, J.J.; Taponen, S.; Lin, R.; Rasanen, M.; Szabo, Z.; Gao, E.; Rahtu-Korpela, L.; et al. Systemic Blockade of ACVR2B Ligands Protects Myocardium from Acute Ischemia-Reperfusion Injury. Mol. Ther. 2019, 27, 600–610. [Google Scholar] [CrossRef] [Green Version]
- Maguer-Satta, V.; Rimokh, R. FLRG, member of the follistatin family, a new player in hematopoiesis. Mol Cell Endocrinol. 2004, 225, 109–118. [Google Scholar] [CrossRef]
- Maguer-Satta, V.; Forissier, S.; Bartholin, L.; Martel, S.; Jeanpierre, S.; Bachelard, E.; Rimokh, R. A novel role for fibronectin type I domain in the regulation of human hematopoietic cell adhesiveness through binding to follistatin domains of FLRG and follistatin. Exp. Cell Res. 2006, 312, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Garber, K. No longer going to waste. Nat. Biotechnol. 2016, 34, 458–461. [Google Scholar] [CrossRef] [PubMed]
- Nolan-Stevaux, O.; Zhong, W.; Culp, S.; Shaffer, K.; Hoover, J.; Wickramasinghe, D.; Ruefli-Brasse, A. Endoglin requirement for BMP9 signaling in endothelial cells reveals new mechanism of action for selective anti-endoglin antibodies. PLoS ONE 2012, 7, e50920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Li, X.; Yang, G.; Li, L.; Hu, W.; Zhang, L.; Liu, H.; Zheng, H.; Tan, M.; Zhu, D. Circulating bone morphogenetic protein-9 in relation to metabolic syndrome and insulin resistance. Sci. Rep. 2017, 7, 17529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owen, N.E.; Alexander, G.J.; Sen, S.; Bunclark, K.; Polwarth, G.; Pepke-Zaba, J.; Davenport, A.P.; Morrell, N.W.; Upton, P.D. Reduced circulating BMP10 and BMP9 and elevated endoglin are associated with disease severity, decompensation and pulmonary vascular syndromes in patients with cirrhosis. EBioMedicine 2020, 56, 102794. [Google Scholar] [CrossRef]
- Junnila, R.K.; List, E.O.; Berryman, D.E.; Murrey, J.W.; Kopchick, J.J. The GH/IGF-1 axis in ageing and longevity. Nat. Rev. Endocrinol. 2013, 9, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Forbes, B.E.; Blyth, A.J.; Wit, J.M. Disorders of IGFs and IGF-1R signaling pathways. Mol. Cell Endocrinol. 2020, 518, 111035. [Google Scholar] [CrossRef]
- Stuard, W.L.; Titone, R.; Robertson, D.M. The IGF/Insulin-IGFBP Axis in Corneal Development, Wound Healing, and Disease. Front Endocrinol. (Lausanne) 2020, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Juul, A.; Bang, P.; Hertel, N.T.; Main, K.; Dalgaard, P.; Jørgensen, K.; Müller, J.; Hall, K.; Skakkebaek, N.E. Serum insulin-like growth factor-I in 1030 healthy children, adolescents, and adults: Relation to age, sex, stage of puberty, testicular size, and body mass index. J. Clin. Endocrinol. Metab. 1994, 78, 744–752. [Google Scholar] [CrossRef]
- Zhang, W.B.; Aleksic, S.; Gao, T.; Weiss, E.F.; Demetriou, E.; Verghese, J.; Holtzer, R.; Barzilai, N.; Milman, S. Insulin-like Growth Factor-1 and IGF Binding Proteins Predict All-Cause Mortality and Morbidity in Older Adults. Cells 2020, 9, 1368. [Google Scholar] [CrossRef]
- Tsai, S.; Clemente-Casares, X.; Zhou, A.C.; Lei, H.; Ahn, J.J.; Chan, Y.T.; Choi, O.; Luck, H.; Woo, M.; Dunn, S.E.; et al. Insulin Receptor-Mediated Stimulation Boosts T Cell Immunity during Inflammation and Infection. Cell Metab. 2018, 28, 922–934.e924. [Google Scholar] [CrossRef] [Green Version]
- Omarjee, L.; Janin, A.; Perrot, F.; Laviolle, B.; Meilhac, O.; Mahe, G. Targeting T-cell senescence and cytokine storm with rapamycin to prevent severe progression in COVID-19. Clin. Immunol. 2020, 108464. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Kirkwood, T.B.L.; Bohr, V.A. Mitochondria in the signaling pathways that control longevity and health span. Ageing Res. Rev. 2019, 54, 100940. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Hsin, H.; Libina, N.; Kenyon, C. Regulation of the Caenorhabditis elegans longevity protein DAF-16 by insulin/IGF-1 and germline signaling. Nat. Genet. 2001, 28, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Y. Searching for the real function of mTOR signaling in the regulation of PD-L1 expression. Transl. Oncol. 2020, 13, 100847. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, Q.; Tan, L.; Xu, Y.; Xie, X.; Zhao, Y. The Regulatory Effects of mTOR Complexes in the Differentiation and Function of CD4(+) T Cell Subsets. J. Immunol. Res. 2020, 2020, 3406032. [Google Scholar] [CrossRef] [Green Version]
- Ieronymaki, E.; Daskalaki, M.G.; Lyroni, K.; Tsatsanis, C. Insulin Signaling and Insulin Resistance Facilitate Trained Immunity in Macrophages Through Metabolic and Epigenetic Changes. Front. Immunol. 2019, 10, 1330. [Google Scholar] [CrossRef]
- Daenthanasanmak, A.; Iamsawat, S.; Chakraborty, P.; Nguyen, H.D.; Bastian, D.; Liu, C.; Mehrotra, S.; Yu, X.Z. Targeting Sirt-1 controls GVHD by inhibiting T-cell allo-response and promoting Treg stability in mice. Blood 2019, 133, 266–279. [Google Scholar] [CrossRef] [Green Version]
- Barzilai, N.; Huffman, D.M.; Muzumdar, R.H.; Bartke, A. The critical role of metabolic pathways in aging. Diabetes 2012, 61, 1315–1322. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Finkel, T. Key proteins and pathways that regulate lifespan. J. Biol. Chem. 2017, 292, 6452–6460. [Google Scholar] [CrossRef] [Green Version]
- Rahmani, J.; Kord Varkaneh, H.; Clark, C.; Zand, H.; Bawadi, H.; Ryan, P.M.; Fatahi, S.; Zhang, Y. The influence of fasting and energy restricting diets on IGF-1 levels in humans: A systematic review and meta-analysis. Ageing Res. Rev. 2019, 53, 100910. [Google Scholar] [CrossRef] [PubMed]
- Sanese, P.; Forte, G.; Disciglio, V.; Grossi, V.; Simone, C. FOXO3 on the Road to Longevity: Lessons From SNPs and Chromatin Hubs. Comput. Struct. Biotechnol. J. 2019, 17, 737–745. [Google Scholar] [CrossRef] [PubMed]
- Kano, Y.; Hiragami, F.; Motoda, H.; Akiyama, J.; Koike, Y.; Gomita, Y.; Inoue, S.; Kawaura, A.; Furuta, T.; Kawamura, K. C-SH2 point mutation converts p85β regulatory subunit of phosphoinositide 3-kinase to an anti-aging gene. Sci. Rep. 2019, 9, 12683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flachsbart, F.; Caliebe, A.; Kleindorp, R.; Blanche, H.; von Eller-Eberstein, H.; Nikolaus, S.; Schreiber, S.; Nebel, A. Association of FOXO3A variation with human longevity confirmed in German centenarians. Proc. Natl. Acad. Sci. USA 2009, 106, 2700–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bever, K.M.; Borazanci, E.H.; Thompson, E.A.; Durham, J.N.; Pinero, K.; Jameson, G.S.; Vrana, A.; Liu, M.; Wilt, C.; Wu, A.A.; et al. An exploratory study of metformin with or without rapamycin as maintenance therapy after induction chemotherapy in patients with metastatic pancreatic adenocarcinoma. Oncotarget 2020, 11, 1929–1941. [Google Scholar] [CrossRef]
- Zaidi, S.; Gandhi, J.; Joshi, G.; Smith, N.L.; Khan, S.A. The anticancer potential of metformin on prostate cancer. Prostate Cancer Prostatic Dis. 2019, 22, 351–361. [Google Scholar] [CrossRef]
- Geagea, A.G.; Rizzo, M.; Jurjus, A.; Cappello, F.; Leone, A.; Tomasello, G.; Gracia, C.; Kattar, S.A.; Massaad-Massade, L.; Eid, A. Correction: A novel therapeutic approach to colorectal cancer in diabetes: Role of metformin and rapamycin. Oncotarget 2019, 10, 6538–6539. [Google Scholar] [CrossRef]
- Ebadi, P.; Fazeli, M. Anti-photoaging potential of propolis extract in UVB-irradiated human dermal fibroblasts through increasing the expression of FOXO3A and NGF genes. Biomed Pharm. 2017, 95, 47–54. [Google Scholar] [CrossRef]
- Shahinozzaman, M.; Obanda, D.N.; Tawata, S. Chemical composition and pharmacological properties of Macaranga-type Pacific propolis: A review. Phytother. Res. 2020. [Google Scholar] [CrossRef]
- Morgentaler, A. Nerve growth factor as a new treatment for testosterone deficiency? EBioMedicine 2018, 36, 10–11. [Google Scholar] [CrossRef] [Green Version]
- Kalra, S.; Baruah, M.P.; Unnikrishnan, A.G. Celebrating life with Rita Levi-Montalcini: A hundred years and more. Indian J. Endocrinol. Metab. 2012, 16, 327–328. [Google Scholar] [CrossRef] [PubMed]
- Kassem, D.H.; Kamal, M.M. Therapeutic efficacy of umbilical cord-derived stem cells for diabetes mellitus: A meta-analysis study. Stem Cell Res. 2020, 11, 484. [Google Scholar] [CrossRef] [PubMed]
- Dorling, J.L.; Martin, C.K.; Redman, L.M. Calorie restriction for enhanced longevity: The role of novel dietary strategies in the present obesogenic environment. Ageing Res. Rev. 2020, 101038. [Google Scholar] [CrossRef] [PubMed]
- Viña, J.; Rodriguez-Mañas, L.; Salvador-Pascual, A.; Tarazona-Santabalbina, F.J.; Gomez-Cabrera, M.C. Exercise: The lifelong supplement for healthy ageing and slowing down the onset of frailty. J. Physiol. 2016, 594, 1989–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Yang, Y.; Zhang, T.; Su, Z.; Yu, D.; Lin, Q.; Chen, H.; Zhang, Q.; Xiang, Q.; Xue, W.; et al. Nasal delivery of nerve growth factor rescue hypogonadism by up-regulating GnRH and testosterone in aging male mice. EBioMedicine 2018, 35, 295–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, G.; Zhang, Q.; Xiao, F.; Xiang, Q.; Su, Z.; Huang, Y. Intranasal TAT-haFGF Improves Cognition and Amyloid-β Pathology in an AβPP/PS1 Mouse Model of Alzheimer’s Disease. J. Alzheimer’s Dis. 2016, 51, 985–990. [Google Scholar] [CrossRef]
- Mitra, S.; Behbahani, H.; Eriksdotter, M. Innovative Therapy for Alzheimer’s Disease-With Focus on Biodelivery of NGF. Front. Neurosci. 2019, 13, 38. [Google Scholar] [CrossRef] [Green Version]
- Forlenza, O.V.; Miranda, A.S.; Guimar, I.; Talib, L.L.; Diniz, B.S.; Gattaz, W.F.; Teixeira, A.L. Decreased Neurotrophic Support is Associated with Cognitive Decline in Non-Demented Subjects. J. Alzheimer’s Dis. 2015, 46, 423–429. [Google Scholar] [CrossRef]
- Chen, J.Y.; Lin, J.R.; Cimprich, K.A.; Meyer, T. A two-dimensional ERK-AKT signaling code for an NGF-triggered cell-fate decision. Mol. Cell 2012, 45, 196–209. [Google Scholar] [CrossRef] [Green Version]
- Sampattavanich, S.; Steiert, B.; Kramer, B.A.; Gyori, B.M.; Albeck, J.G.; Sorger, P.K. Encoding Growth Factor Identity in the Temporal Dynamics of FOXO3 under the Combinatorial Control of ERK and AKT Kinases. Cell Syst. 2018, 6, 664–678.e669. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Peng, X.; Wu, T.; Yang, W.; Liu, W.; Zhang, J.; Su, Y.; Kong, F.; Dou, X.; Li, J. Restricting the induction of NGF in ovarian stroma engenders selective follicular activation through the mTOR signaling pathway. Cell Death Dis. 2017, 8, e2817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelberg, J.M.; Cai, D.; Xaymardan, M. Translation of PDGF cardioprotective pathways. Cardiovasc. Toxicol. 2003, 3, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.; Zheng, J.; Duignan, I.J.; Edelberg, J.M. PDGF-AB-based functional cardioprotection of the aging rat heart. Exp. Gerontol. 2006, 41, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Keppler, L.; Rutkowski, J. A review of platelet derived growth factor playing pivotal role in bone regeneration. J. Oral Implant. 2014, 40, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Pola, R.; Aprahamian, T.R.; Bosch-Marcé, M.; Curry, C.; Gaetani, E.; Flex, A.; Smith, R.C.; Isner, J.M.; Losordo, D.W. Age-dependent VEGF expression and intraneural neovascularization during regeneration of peripheral nerves. Neurobiol. Aging 2004, 25, 1361–1368. [Google Scholar] [CrossRef] [PubMed]
- Stacker, S.A.; Achen, M.G. Emerging Roles for VEGF-D in Human Disease. Biomolecules 2018, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.; Björndahl, M.A.; Religa, P.; Clasper, S.; Garvin, S.; Galter, D.; Meister, B.; Ikomi, F.; Tritsaris, K.; Dissing, S.; et al. PDGF-BB induces intratumoral lymphangiogenesis and promotes lymphatic metastasis. Cancer Cell 2004, 6, 333–345. [Google Scholar] [CrossRef] [Green Version]
- Rivera, P.; Martos-Moreno, G.; Barrios, V.; Suárez, J.; Pavón, F.J.; Chowen, J.A.; Rodríguez de Fonseca, F.; Argente, J. A combination of circulating chemokines as biomarkers of obesity-induced insulin resistance at puberty. Pediatric Obes. 2020, e12711. [Google Scholar] [CrossRef]
- Karaman, S.; Hollmén, M.; Robciuc, M.R.; Alitalo, A.; Nurmi, H.; Morf, B.; Buschle, D.; Alkan, H.F.; Ochsenbein, A.M.; Alitalo, K.; et al. Blockade of VEGF-C and VEGF-D modulates adipose tissue inflammation and improves metabolic parameters under high-fat diet. Mol. Metab. 2015, 4, 93–105. [Google Scholar] [CrossRef]
- Arum, O.; Saleh, J.K.; Boparai, R.K.; Kopchick, J.J.; Khardori, R.K.; Bartke, A. Preservation of blood glucose homeostasis in slow-senescing somatotrophism-deficient mice subjected to intermittent fasting begun at middle or old age. Age (Dordr) 2014, 36, 9651. [Google Scholar] [CrossRef] [Green Version]
- Panowski, S.H.; Wolff, S.; Aguilaniu, H.; Durieux, J.; Dillin, A. PHA-4/Foxa mediates diet-restriction-induced longevity of C. elegans. Nature 2007, 447, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xie, Y.; Berglund, E.D.; Coate, K.C.; He, T.T.; Katafuchi, T.; Xiao, G.; Potthoff, M.J.; Wei, W.; Wan, Y.; et al. The starvation hormone, fibroblast growth factor-21, extends lifespan in mice. eLife 2012, 1, e00065. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Integrated stress response stimulates FGF21 expression: Systemic enhancer of longevity. Cell Signal 2017, 40, 10–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, S.A.; Sargeant, J.A.; Yates, T.; Takamura, T.; Takayama, H.; Gupta, V.; Brittain, E.; Crawford, J.; Parry, S.A.; Thackray, A.E.; et al. Acute Hyperenergetic, High-Fat Feeding Increases Circulating FGF21, LECT2, and Fetuin-A in Healthy Men. J. Nutr. 2020, 150, 1076–1085. [Google Scholar] [CrossRef]
- Reitman, M.L. FGF21 mimetic shows therapeutic promise. Cell Metab. 2013, 18, 307–309. [Google Scholar] [CrossRef] [Green Version]
- Søberg, S.; Sandholt, C.H.; Jespersen, N.Z.; Toft, U.; Madsen, A.L.; von Holstein-Rathlou, S.; Grevengoed, T.J.; Christensen, K.B.; Bredie, W.L.P.; Potthoff, M.J.; et al. FGF21 Is a Sugar-Induced Hormone Associated with Sweet Intake and Preference in Humans. Cell Metab. 2017, 25, 1045–1053.e1046. [Google Scholar] [CrossRef] [Green Version]
- Elabd, C.; Cousin, W.; Upadhyayula, P.; Chen, R.Y.; Chooljian, M.S.; Li, J.; Kung, S.; Jiang, K.P.; Conboy, I.M. Oxytocin is an age-specific circulating hormone that is necessary for muscle maintenance and regeneration. Nat. Commun. 2014, 5, 4082. [Google Scholar] [CrossRef] [Green Version]
- Maheshwari, H.G.; Silverman, B.L.; Dupuis, J.; Baumann, G. Phenotype and genetic analysis of a syndrome caused by an inactivating mutation in the growth hormone-releasing hormone receptor: Dwarfism of Sindh. J. Clin. Endocrinol. Metab. 1998, 83, 4065–4074. [Google Scholar] [CrossRef] [Green Version]
- Veldhuis, J.D. Aging and hormones of the hypothalamo-pituitary axis: Gonadotropic axis in men and somatotropic axes in men and women. Ageing Res. Rev. 2008, 7, 189–208. [Google Scholar] [CrossRef] [Green Version]
- Bartke, A.; Sun, L.Y.; Longo, V. Somatotropic signaling: Trade-offs between growth, reproductive development, and longevity. Physiol. Rev. 2013, 93, 571–598. [Google Scholar] [CrossRef]
- Brown-Borg, H.M.; Borg, K.E.; Meliska, C.J.; Bartke, A. Dwarf mice and the ageing process. Nature 1996, 384, 33. [Google Scholar] [CrossRef] [PubMed]
- Dominici, F.P.; Hauck, S.; Argentino, D.P.; Bartke, A.; Turyn, D. Increased insulin sensitivity and upregulation of insulin receptor, insulin receptor substrate (IRS)-1 and IRS-2 in liver of Ames dwarf mice. J. Endocrinol. 2002, 173, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Icyuz, M.; Liu, Z.; Fitch, M.; Sun, L.Y. Insulin sensitivity in long-lived growth hormone-releasing hormone knockout mice. Aging (Albany NY) 2020, 12, 18033–18051. [Google Scholar] [CrossRef] [PubMed]
- Van der Spoel, E.; Jansen, S.W.; Akintola, A.A.; Ballieux, B.E.; Cobbaert, C.M.; Slagboom, P.E.; Blauw, G.J.; Westendorp, R.G.J.; Pijl, H.; Roelfsema, F.; et al. Growth hormone secretion is diminished and tightly controlled in humans enriched for familial longevity. Aging Cell 2016, 15, 1126–1131. [Google Scholar] [CrossRef] [PubMed]
- Perls, T.T.; Reisman, N.R.; Olshansky, S.J. Provision or distribution of growth hormone for “antiaging”: Clinical and legal issues. JAMA 2005, 294, 2086–2090. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Chang, J.; Lartigue, L.; Bolen, C.R.; Haddad, F.; Gaudilliere, B.; Ganio, E.A.; Fragiadakis, G.K.; Spitzer, M.H.; Douchet, I.; et al. Expression of specific inflammasome gene modules stratifies older individuals into two extreme clinical and immunological states. Nat. Med. 2017, 23, 174–184. [Google Scholar] [CrossRef] [PubMed]
- Mauer, J.; Chaurasia, B.; Goldau, J.; Vogt, M.C.; Ruud, J.; Nguyen, K.D.; Theurich, S.; Hausen, A.C.; Schmitz, J.; Brönneke, H.S.; et al. Signaling by IL-6 promotes alternative activation of macrophages to limit endotoxemia and obesity-associated resistance to insulin. Nat. Immunol. 2014, 15, 423–430. [Google Scholar] [CrossRef]
- Yu, B.; Chang, J.; Liu, Y.; Li, J.; Kevork, K.; Al-Hezaimi, K.; Graves, D.T.; Park, N.H.; Wang, C.Y. Wnt4 signaling prevents skeletal aging and inflammation by inhibiting nuclear factor-κB. Nat. Med. 2014, 20, 1009–1017. [Google Scholar] [CrossRef] [Green Version]
- Perry, V.H.; Cunningham, C.; Holmes, C. Systemic infections and inflammation affect chronic neurodegeneration. Nat. Rev. Immunol. 2007, 7, 161–167. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Burska, A.N.; Sakthiswary, R.; Sattar, N. Effects of Tumour Necrosis Factor Antagonists on Insulin Sensitivity/Resistance in Rheumatoid Arthritis: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0128889. [Google Scholar] [CrossRef]
- Lu, L.; Zhong, W.; Bian, Z.; Li, Z.; Zhang, K.; Liang, B.; Zhong, Y.; Hu, M.; Lin, L.; Liu, J.; et al. A comparison of mortality-related risk factors of COVID-19, SARS, and MERS: A systematic review and meta-analysis. J. Infect. 2020. [Google Scholar] [CrossRef] [PubMed]
- Brooke, R.T.; Fahy, G.M. Reversing immunosenescence for prevention of COVID-19. Aging (Albany NY) 2020, 12, 11161–11162. [Google Scholar] [CrossRef] [PubMed]
- Wilk, A.J.; Rustagi, A.; Zhao, N.Q.; Roque, J.; Martínez-Colón, G.J.; McKechnie, J.L.; Ivison, G.T.; Ranganath, T.; Vergara, R.; Hollis, T.; et al. A single-cell atlas of the peripheral immune response in patients with severe COVID-19. Nat. Med. 2020, 26, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Spencer, M.E.; Jain, A.; Matteini, A.; Beamer, B.A.; Wang, N.Y.; Leng, S.X.; Punjabi, N.M.; Walston, J.D.; Fedarko, N.S. Serum levels of the immune activation marker neopterin change with age and gender and are modified by race, BMI, and percentage of body fat. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65, 858–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, H.Y.; Cesari, M.; Anton, S.; Marzetti, E.; Giovannini, S.; Seo, A.Y.; Carter, C.; Yu, B.P.; Leeuwenburgh, C. Molecular inflammation: Underpinnings of aging and age-related diseases. Ageing Res. Rev. 2009, 8, 18–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queck, A.; Bode, H.; Uschner, F.E.; Brol, M.J.; Graf, C.; Schulz, M.; Jansen, C.; Praktiknjo, M.; Schierwagen, R.; Klein, S.; et al. Systemic MCP-1 Levels Derive Mainly From Injured Liver and Are Associated With Complications in Cirrhosis. Front. Immunol. 2020, 11, 354. [Google Scholar] [CrossRef] [Green Version]
- Gschwandtner, M.; Derler, R.; Midwood, K.S. More Than Just Attractive: How CCL2 Influences Myeloid Cell Behavior Beyond Chemotaxis. Front. Immunol. 2019, 10, 2759. [Google Scholar] [CrossRef] [Green Version]
- Tsaur, I.; Noack, A.; Makarevic, J.; Oppermann, E.; Waaga-Gasser, A.M.; Gasser, M.; Borgmann, H.; Huesch, T.; Gust, K.M.; Reiter, M.; et al. CCL2 Chemokine as a Potential Biomarker for Prostate Cancer: A Pilot Study. Cancer Res. Treat. 2015, 47, 306–312. [Google Scholar] [CrossRef]
- Lubowicka, E.; Przylipiak, A.; Zajkowska, M.; Piskór, B.M.; Malinowski, P.; Fiedorowicz, W.; Ławicki, S. Plasma Chemokine CCL2 and Its Receptor CCR2 Concentrations as Diagnostic Biomarkers for Breast Cancer Patients. Biomed. Res. Int. 2018, 2018, 2124390. [Google Scholar] [CrossRef]
- Mulholland, B.S.; Forwood, M.R.; Morrison, N.A. Monocyte Chemoattractant Protein-1 (MCP-1/CCL2) Drives Activation of Bone Remodelling and Skeletal Metastasis. Curr. Osteoporos. Rep. 2019, 17, 538–547. [Google Scholar] [CrossRef] [Green Version]
- Joly-Amado, A.; Hunter, J.; Quadri, Z.; Zamudio, F.; Rocha-Rangel, P.V.; Chan, D.; Kesarwani, A.; Nash, K.; Lee, D.C.; Morgan, D.; et al. CCL2 Overexpression in the Brain Promotes Glial Activation and Accelerates Tau Pathology in a Mouse Model of Tauopathy. Front. Immunol. 2020, 11, 997. [Google Scholar] [CrossRef] [PubMed]
- Zhu, E.; Liu, Z.; He, W.; Deng, B.; Shu, X.; He, Z.; Wu, X.; Ke, X.; Nie, R. CC chemokine receptor 2 functions in osteoblastic transformation of valvular interstitial cells. Life Sci. 2019, 228, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Sádaba, J.R.; Martínez-Martínez, E.; Arrieta, V.; Álvarez, V.; Fernández-Celis, A.; Ibarrola, J.; Melero, A.; Rossignol, P.; Cachofeiro, V.; López-Andrés, N. Role for Galectin-3 in Calcific Aortic Valve Stenosis. J. Am. Heart Assoc. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Yousefzadeh, M.J.; Schafer, M.J.; Noren Hooten, N.; Atkinson, E.J.; Evans, M.K.; Baker, D.J.; Quarles, E.K.; Robbins, P.D.; Ladiges, W.C.; LeBrasseur, N.K.; et al. Circulating levels of monocyte chemoattractant protein-1 as a potential measure of biological age in mice and frailty in humans. Aging Cell 2018, 17. [Google Scholar] [CrossRef]
- Villeda, S.A.; Luo, J.; Mosher, K.I.; Zou, B.; Britschgi, M.; Bieri, G.; Stan, T.M.; Fainberg, N.; Ding, Z.; Eggel, A.; et al. The ageing systemic milieu negatively regulates neurogenesis and cognitive function. Nature 2011, 477, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, A.L.; Gama, C.S.; Rocha, N.P.; Teixeira, M.M. Revisiting the Role of Eotaxin-1/CCL11 in Psychiatric Disorders. Front. Psychiatry 2018, 9, 241. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.K.; Giles, D.A.; Segal, B.M.; Irani, D.N. An emerging role for eotaxins in neurodegenerative disease. Clin. Immunol. 2018, 189, 29–33. [Google Scholar] [CrossRef]
- Kawamoto, D.; Amado, P.P.L.; Albuquerque-Souza, E.; Bueno, M.R.; Vale, G.C.; Saraiva, L.; Mayer, M.P.A. Chemokines and cytokines profile in whole saliva of patients with periodontitis. Cytokine 2020, 135, 155197. [Google Scholar] [CrossRef]
- Korbecki, J.; Kojder, K.; Barczak, K.; Simińska, D.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Hypoxia Alters the Expression of CC Chemokines and CC Chemokine Receptors in a Tumor-A Literature Review. Int. J. Mol. Sci. 2020, 21, 5647. [Google Scholar] [CrossRef]
- Thijs, J.; Krastev, T.; Weidinger, S.; Buckens, C.F.; de Bruin-Weller, M.; Bruijnzeel-Koomen, C.; Flohr, C.; Hijnen, D. Biomarkers for atopic dermatitis: A systematic review and meta-analysis. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 453–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; Xu, P.; Zhang, X.; Liao, M.; Dong, Q.; Cong, T.; Tang, B.; Yang, X.; Ye, M.; Chang, Y.J.; et al. Aging-induced IL27Ra Signaling Impairs Hematopoietic Stem Cells. Blood 2020. [Google Scholar] [CrossRef]
- Peshkova, I.O.; Aghayev, T.; Fatkhullina, A.R.; Makhov, P.; Titerina, E.K.; Eguchi, S.; Tan, Y.F.; Kossenkov, A.V.; Khoreva, M.V.; Gankovskaya, L.V.; et al. IL-27 receptor-regulated stress myelopoiesis drives abdominal aortic aneurysm development. Nat. Commun. 2019, 10, 5046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, C.U.; Birkholz, J.; Weins, N.; Doganci, A.; Gehring, S.; Zepp, F.; Knuf, M. Dendritic cells change IL-27 production pattern during childhood. BMC Res. Notes 2015, 8, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; He, C.; Nair, L.; Yeung, J.; Egwuagu, C.E. Interleukin 12 (IL-12) family cytokines: Role in immune pathogenesis and treatment of CNS autoimmune disease. Cytokine 2015, 75, 249–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, W.J.; Chen, L.K.; Liang, C.K.; Peng, L.N.; Chiou, S.T.; Chou, P. Soluble ICAM-1, Independent of IL-6, Is Associated with Prevalent Frailty in Community-Dwelling Elderly Taiwanese People. PLoS ONE 2016, 11, e0157877. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wu, Y.-L.; Zhong, J.; Jiang, F.-X.; Tian, X.-l.; Yu, L.-F. Angiotensin II Type 1 Receptor Antagonist Suppress Angiogenesis and Growth of Gastric Cancer Xenografts. Dig. Dis. Sci. 2008, 53, 1206–1210. [Google Scholar] [CrossRef]
- Cosarderelioglu, C.; Nidadavolu, L.S.; George, C.J.; Oh, E.S.; Bennett, D.A.; Walston, J.D.; Abadir, P.M. Brain Renin-Angiotensin System at the Intersect of Physical and Cognitive Frailty. Front. Neurosci. 2020, 14, 586314. [Google Scholar] [CrossRef]
- Sadler, J.E.; Budde, U.; Eikenboom, J.C.; Favaloro, E.J.; Hill, F.G.; Holmberg, L.; Ingerslev, J.; Lee, C.A.; Lillicrap, D.; Mannucci, P.M.; et al. Update on the pathophysiology and classification of von Willebrand disease: A report of the Subcommittee on von Willebrand Factor. J. Thromb. Haemost. 2006, 4, 2103–2114. [Google Scholar] [CrossRef]
- Gragnano, F.; Sperlongano, S.; Golia, E.; Natale, F.; Bianchi, R.; Crisci, M.; Fimiani, F.; Pariggiano, I.; Diana, V.; Carbone, A.; et al. The Role of von Willebrand Factor in Vascular Inflammation: From Pathogenesis to Targeted Therapy. Mediat. Inflamm. 2017, 2017, 5620314. [Google Scholar] [CrossRef]
- Seaman, C.D.; Ragni, M.V. The Association of Aging With Von Willebrand Factor Levels and Bleeding Risk in Type 1 Von Willebrand Disease. Clin. Appl. Thromb. Hemost. 2018, 24, 434–438. [Google Scholar] [CrossRef] [PubMed]
- Bertuglia, S.; Veronese, F.M.; Pasut, G. Polyethylene glycol and a novel developed polyethylene glycol-nitric oxide normalize arteriolar response and oxidative stress in ischemia-reperfusion. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1536–H1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, M.D.; Atkinson, T.M.; Lindner, J.R. Platelets and von Willebrand factor in atherogenesis. Blood 2017, 129, 1415–1419. [Google Scholar] [CrossRef] [PubMed]
- Castellano, J.M.; Mosher, K.I.; Abbey, R.J.; McBride, A.A.; James, M.L.; Berdnik, D.; Shen, J.C.; Zou, B.; Xie, X.S.; Tingle, M.; et al. Human umbilical cord plasma proteins revitalize hippocampal function in aged mice. Nature 2017, 544, 488–492. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Kim, H.S. The anti-inflammatory role of tissue inhibitor of metalloproteinase-2 in lipopolysaccharide-stimulated microglia. J. Neuroinflamm. 2014, 11, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, M.Y.; Kim, B.J.; Kim, H.J.; Jin, J.M.; Yoon, H.J.; Hwang, J.S.; Park, K.K. Anti-cancer effect of dung beetle glycosaminoglycans on melanoma. BMC Cancer 2019, 19, 9. [Google Scholar] [CrossRef]
- Escalona, R.M.; Bilandzic, M.; Western, P.; Kadife, E.; Kannourakis, G.; Findlay, J.K.; Ahmed, N. TIMP-2 regulates proliferation, invasion and STAT3-mediated cancer stem cell-dependent chemoresistance in ovarian cancer cells. BMC Cancer 2020, 20, 960. [Google Scholar] [CrossRef]
- Bussian, T.J.; Aziz, A.; Meyer, C.F.; Swenson, B.L.; van Deursen, J.M.; Baker, D.J. Clearance of senescent glial cells prevents tau-dependent pathology and cognitive decline. Nature 2018, 562, 578–582. [Google Scholar] [CrossRef]
- Baker, D.J.; Childs, B.G.; Durik, M.; Wijers, M.E.; Sieben, C.J.; Zhong, J.; Saltness, R.A.; Jeganathan, K.B.; Verzosa, G.C.; Pezeshki, A.; et al. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature 2016, 530, 184–189. [Google Scholar] [CrossRef] [Green Version]
- Baar, M.P.; Brandt, R.M.C.; Putavet, D.A.; Klein, J.D.D.; Derks, K.W.J.; Bourgeois, B.R.M.; Stryeck, S.; Rijksen, Y.; van Willigenburg, H.; Feijtel, D.A.; et al. Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell 2017, 169, 132–147.e116. [Google Scholar] [CrossRef] [Green Version]
- Hoare, M.; Narita, M. Transmitting senescence to the cell neighbourhood. Nat. Cell Biol. 2013, 15, 887–889. [Google Scholar] [CrossRef]
- Eren, M.; Boe, A.E.; Klyachko, E.A.; Vaughan, D.E. Role of plasminogen activator inhibitor-1 in senescence and aging. Semin. Thromb. Hemost. 2014, 40, 645–651. [Google Scholar] [CrossRef]
- Ghosh, A.K.; Rai, R.; Park, K.E.; Eren, M.; Miyata, T.; Wilsbacher, L.D.; Vaughan, D.E. A small molecule inhibitor of PAI-1 protects against doxorubicin-induced cellular senescence. Oncotarget 2016, 7, 72443–72457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Li, P.; Gao, L.; Zhang, J.; Yang, Z.; Bledsoe, G.; Chang, E.; Chao, L.; Chao, J. Kallistatin reduces vascular senescence and aging by regulating microRNA-34a-SIRT1 pathway. Aging Cell 2017, 16, 837–846. [Google Scholar] [CrossRef]
- Amor, C.; Feucht, J.; Leibold, J.; Ho, Y.J.; Zhu, C.; Alonso-Curbelo, D.; Mansilla-Soto, J.; Boyer, J.A.; Li, X.; Giavridis, T.; et al. Senolytic CAR T cells reverse senescence-associated pathologies. Nature 2020, 583, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, L.J.H.; Caspi, A.; Ambler, A.; Danese, A.; Elliott, M.; Eugen-Olsen, J.; Hariri, A.R.; Harrington, H.; Houts, R.; Poulton, R.; et al. Association Between Elevated suPAR, a New Biomarker of Inflammation, and Accelerated Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Berezina, T.N.; Stelmakh, S.A.; Dergacheva, E.V. The effect of retirement stress on the biopsychological age in Russia and the Republic of Kazakhstan: A cross-cultural study. Psychologist 2019, 5. [Google Scholar] [CrossRef]
- Galkin, F.; Zhang, B.; Dmitriev, S.E.; Gladyshev, V.N. Reversibility of irreversible aging. Ageing Res. Rev. 2019, 49, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Mattina, G.F.; Van Lieshout, R.J.; Steiner, M. Inflammation, depression and cardiovascular disease in women: The role of the immune system across critical reproductive events. Adv. Cardiovasc. Dis. 2019, 13. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.B.; Lee-McMullen, B.; Phelan, D.; Gilkes, J.; Carter, C.S.; Buford, T.W. The renin-angiotensin system and prevention of age-related functional decline: Where are we now? Age 2015, 37, 9753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, N.; Dey, S. Protective response of Sestrin under stressful conditions in aging. Ageing Res. Rev. 2020, 64, 101186. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Jedrychowski, M.P.; Schweppe, D.K.; Huttlin, E.L.; Yu, Q.; Heppner, D.E.; Li, J.; Long, J.; Mills, E.L.; Szpyt, J.; et al. A Quantitative Tissue-Specific Landscape of Protein Redox Regulation during Aging. Cell 2020, 180, 968–983.e924. [Google Scholar] [CrossRef] [PubMed]
- Marschallinger, J.; Schäffner, I.; Klein, B.; Gelfert, R.; Rivera, F.J.; Illes, S.; Grassner, L.; Janssen, M.; Rotheneichner, P.; Schmuckermair, C.; et al. Structural and functional rejuvenation of the aged brain by an approved anti-asthmatic drug. Nat. Commun. 2015, 6, 8466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, R.; Padwad, Y. In search of nutritional anti-aging targets: TOR inhibitors, SASP modulators, and BCL-2 family suppressors. Nutrition 2019, 65, 33–38. [Google Scholar] [CrossRef]
- Zhang, D.Y.; Gao, T.; Xu, R.J.; Sun, L.; Zhang, C.F.; Bai, L.; Chen, W.; Liu, K.Y.; Zhou, Y.; Jiao, X.; et al. SIRT3 Transfection of Aged Human Bone Marrow-Derived Mesenchymal Stem Cells Improves Cell Therapy-Mediated Myocardial Repair. Rejuvenation Res. 2020. [Google Scholar] [CrossRef]
- Bradshaw, P.C. Cytoplasmic and Mitochondrial NADPH-Coupled Redox Systems in the Regulation of Aging. Nutrients 2019, 11, 504. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Circulating Indicators of Aging | Dynamics during Aging | Function/Risk Factor in the Elderly | Reasons of the Condition | Lifespan Influence |
|---|---|---|---|---|
| Age-dependent vascular disorders | ||||
| Capillary network | Decrease | Reduction of the peripheral tissue oxygen level | Reduction of VEGF-A and PDGF-AB level | Negative |
| Maximum capillary diameter | Decrease | Thrombosis, high blood pressure | Angiotensin system disorder, neural atrophy | Negative |
| Number of sensor neurons | Decrease | Stress-dependent blood pressure | Neural function decline, VEGF level reduction | Negative |
| Inflammation indicators | ||||
| Fibrinogen | Increase | Tissue inflammation | Immunity misbalance, autoimmunity | Negative |
| PUFA | Increase | Tissue inflammation | Immunity misbalance, autoimmunity | Negative |
| β2-Microglobulin | Increase | Inflammation | Renal function decline | Negative |
| Neopterin | Increase | Inflammation, autoimmunity | Intracellular infections | Unknown |
| Anti-inflammatory indicator | ||||
| Sphingomyelin | Decrease | Cell protection, intestinal infection protection | Age-related decline of sphingomyelin synthase | Positive |
| Kidney disorder indicators | ||||
| BUN/creatinine ratio | Increase | Risk of cardiovascular mortality | Renal function decline | Negative |
| Albumin/creatinine ratio | Increase | Risk of cardiovascular mortality | Renal function decline | Negative |
| Calcium | Increase and level variation | Risk of cardiovascular and cancer mortality | Renal function decline | Negative |
| Lipids | ||||
| Abundancy of different classes of lipids | Decrease | Vascular protection/nutrition | Lipid metabolism disorders | Unknown |
| Reactive oxygen species/antioxidants | ||||
| NAD+ | Decrease | Supports energy processes, antioxidant | Inflammation, metabolic misbalance | Positive |
| ROS (D-ROM/TTL ratio) | Increase | Risk of cardiovascular, diabetes, and cancer mortality | Aging process, Nrf2 level decline | Negative |
| H2S | Decrease | Regulation of 1nflammation, antioxidant | CGL oxidation stress response decline | Positive |
| Circulating Molecule | Dynamics during Aging | Function/Risk Factor in Elderly | Molecule Longevity Influence |
|---|---|---|---|
| Growth factors and hormones variation during aging | |||
| TGF-β | Increase | Niche and nerve cells senescence induction | Negative |
| GDF8 | Increase | Muscular regeneration block, obesity, heart failure risk | Negative |
| GDF11 | Decrease | Muscular regeneration, improved angiogenesis in the brain | Positive |
| Activin A | Increase | Cardiovascular and heart dysfunction risk | Negative |
| FSTL3 | Increase | Cardiovascular and heart dysfunction indicator | Unknown |
| BMP9 | Decrease | Endothelial maturation and integrity, new vessel formation inhibition, insulin resistance inhibition. A high level of ENG is a sign of diabetes or liver disease. | Unknown |
| IGF-1/IGFBP-3 | Increase | All-cause mortality risk, reducing autophagy, insulin resistance | Conditional |
| NGF, BDNF, GDNF | Decrease | Neurons survival, prevention of cognitive decline | Positive |
| PDGF-AB | Decrease | Cardiovascular and heart dysfunction risk reduction | Positive |
| VEGF | Decrease | Vessel growth and remodeling | Conditional |
| VEGF-D and PDGF-BB | Increase | Lymphatic vessel formation in fat tissue, insulin resistance | Unknown |
| FGF21 | Decrease | Starvation factor, IGF/GHR signaling suppression, Cell survival under stress support | Positive |
| Oxytocin | Decrease | Muscle and tissue regeneration | Positive |
| Growth hormone (GH) | Decrease | Muscle and tissue regeneration, insulin resistance rise | Conditional |
| Ghrelin “hunger hormone” | Decrease | Secreted in response to diet restriction, regulates GH secretion and energy metabolism. Impact on obesity and insulin resistance. | Conditional |
| Pro-inflammatory factors | |||
| TNFα, IL1β, IL6, CRP | Increase | Chronic increase indicates tissue inflammation and age-dependent immunity dysfunction | Negative |
| CCL2 | Increase | Aortic inflammation, Aortic stenosis | Negative |
| CCL11 | Increase | Recruitment of immune cells to inflamed tissues, cognitive function decline | Negative |
| CCL27 | Increase | Recruitment of T cells to inflamed tissues, cognitive function decline | Negative |
| IL27 | Increase | Blood stem cell aging, tissue inflammation, infection, stress response, vascular damage | Negative |
| Soluble VCAM1 ICAM1 | Increase | Indicator of immune cells recruitment to inflamed tissues | Negative |
| vWF | Increase | Vascular inflammation, risk of diabetes, stroke, myocardial infarction | Negative |
| Anti-inflammatory factors | |||
| TIMP2 | Decrease | Hippocampus function maintenance | Positive |
| Senescence indicators | |||
| PAI-1 | Increase | Immune cells senescence indicator | Negative |
| Soluble uPAR | Increase | Immune cells senescence indicator | Negative |
| Kallistatin | Unknown | Oxidative stress inhibition, senescence prevention | Positive |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rybtsova, N.; Berezina, T.; Kagansky, A.; Rybtsov, S. Can Blood-Circulating Factors Unveil and Delay Your Biological Aging? Biomedicines 2020, 8, 615. https://doi.org/10.3390/biomedicines8120615
Rybtsova N, Berezina T, Kagansky A, Rybtsov S. Can Blood-Circulating Factors Unveil and Delay Your Biological Aging? Biomedicines. 2020; 8(12):615. https://doi.org/10.3390/biomedicines8120615
Chicago/Turabian StyleRybtsova, Natalia, Tatiana Berezina, Alexander Kagansky, and Stanislav Rybtsov. 2020. "Can Blood-Circulating Factors Unveil and Delay Your Biological Aging?" Biomedicines 8, no. 12: 615. https://doi.org/10.3390/biomedicines8120615
APA StyleRybtsova, N., Berezina, T., Kagansky, A., & Rybtsov, S. (2020). Can Blood-Circulating Factors Unveil and Delay Your Biological Aging? Biomedicines, 8(12), 615. https://doi.org/10.3390/biomedicines8120615