Molecular and Lifestyle Factors Modulating Obesity Disease

 ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
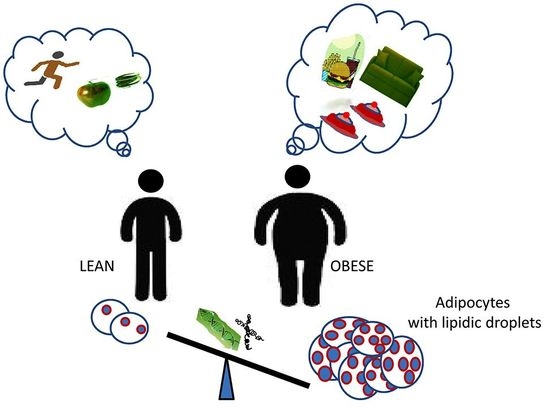
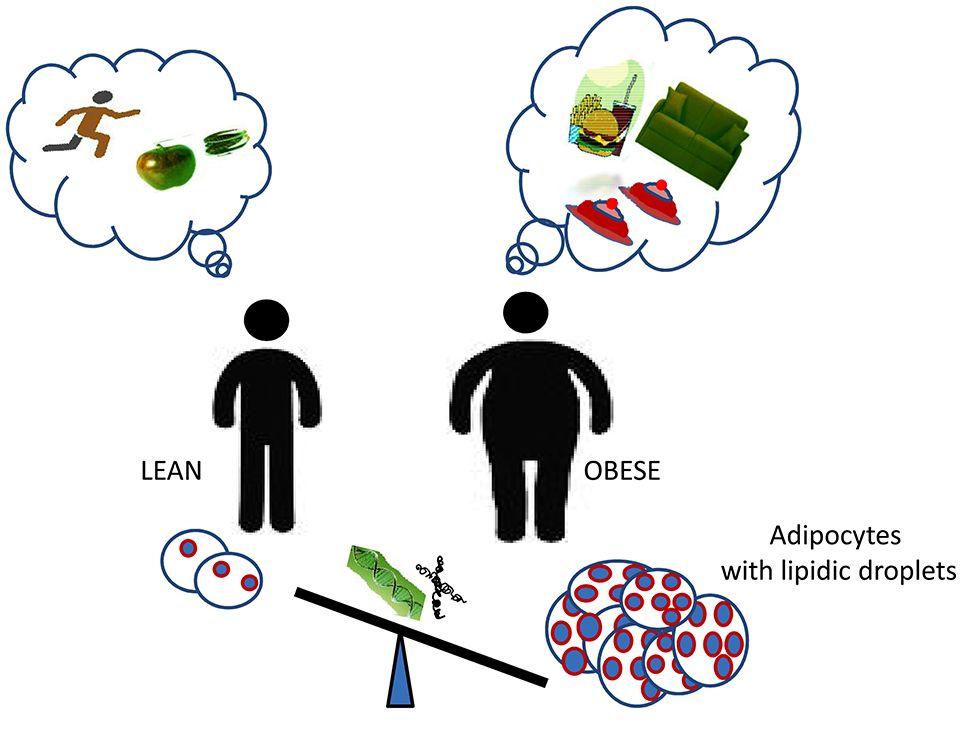
:
1. Introduction
2. Obesity and Mesenchymal Stem Cells
2.1. Stem Cell Lineage Differentiation towards Osteogenesis or Adipogenesis
2.2. Stem Cell Lineage Differentiation towards Chondrogenesis or Adipogenesis
2.3. Obesity and Osteoporosis Appear as Partenering Traits
3. Epigenetics and Adipogenesis
4. Vitamin D and Obesity
5. Physical Exercise and Changes in Gut Microbiota in Obese Individuals
6. Body Composition Alterations and Socio-economic Impact
6.1. Prenatal and Post-Natal Changes
6.2. Adulthood Changes
6.3. Senility Changes
6.4. Socioeconomic Impact of Obesity
7. Conclusions
Endnote
Author Contributions
Funding
Conflicts of Interest
Abbreviation
| ALP | Alkaline Phosphatase |
| AT | Adipose Tissue |
| BAT | Brown Adipose Tissue |
| BMI | Body Mass Index |
| BMPs | Bone Morphogenic Proteins |
| COL1A2 | Collagen type I isoform 2 |
| D [25(OH)D | 25-hydroxyvitamin DErk Extracellular signal-regulated kinase |
| FFM | Fat-free mass |
| FGFs | Fibroblast Growth Factors |
| GNPDA2 | Glucosamine-6-Phosphate Deaminase 2 |
| HFD | High-Fat Diet |
| HSPA12A | Heat Shock Protein Family A member 12A |
| IGF1 | Insulin like growth factor 1 |
| MSCs | Mesenchymal Stem Cells |
| OCN = | OsteocalcinPET-CT positron emission tomography |
| PI3K | PhosphatidylInositol 3-Kinase |
| PCNA | Proliferating Cell Nuclear Antigen |
| PPARγ | Peroxisome Proliferator-Activated Receptor gamma |
| PTH | Parathyroid Hormone |
| RBP4 | Retinol-Binding Protein |
| SEMA3G | Semaphorin-3G |
| SIRT | Sirtuin |
| SMAD | Small Mother Against Decapentaplegic |
| TGFβ | Transforming Growth Factor-β |
| UCP1 | Uncoupling Protein-1 |
| VDR | Vitamin D Receptor |
| ZFP423 | Zinc Finger Protein 423 |
| WAT | White Adipose Tissue |
| WC | Waist Circumference |
References
- Chamberlain, G.; Fox, J.; Ashton, B.; Middleton, J. Concise review: Mesenchymal stem cells: Their phenotype, differentiation capacity, immunological features, and potential for homing. Stem Cells 2007, 25, 2739–2749. [Google Scholar] [CrossRef] [Green Version]
- Rao, T.P.; Kuhl, M. An updated overview on Wnt signaling pathways: A prelude for more. Circ. Res. 2010, 106, 1798–1806. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARgamma and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vimalraj, S.; Arumugam, B.; Miranda, P.J.; Selvamurugan, N. Runx2: Structure, function, and phosphorylation in osteoblast differentiation. Int. J. Biol. Macromol. 2015, 78, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yang, S.; Shao, J.; Li, Y.P. Signaling and transcriptional regulation in osteoblast commitment and differentiation. Front. Biosci. 2007, 12, 3068–3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.Y.; Li, X.; Qian, S.W.; Guo, L.; Huang, H.Y.; He, Q.; Liu, Y.; Ma, C.G.; Tang, Q.Q. Down-regulation of type I Runx2 mediated by dexamethasone is required for 3T3-L1 adipogenesis. Mol. Endocrinol. 2012, 26, 798–808. [Google Scholar] [CrossRef] [Green Version]
- Valenti, M.T.; Garbin, U.; Pasini, A.; Zanatta, M.; Stranieri, C.; Manfro, S.; Zucal, C.; Dalle Carbonare, L. Role of Ox-PAPCs in the Differentiation of Mesenchymal Stem Cells (MSCs) and Runx2 and PPAR gamma 2 Expression in MSCs-Like of Osteoporotic Patients. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.M.; Zhao, H.; Wang, E.H. A high-fat diet induces obesity and impairs bone acquisition in young male mice. Mol. Med. Rep. 2013, 7, 1203–1208. [Google Scholar] [CrossRef] [Green Version]
- Chijimatsu, R.; Kobayashi, M.; Ebina, K.; Iwahashi, T.; Okuno, Y.; Hirao, M.; Fukuhara, A.; Nakamura, N.; Yoshikawa, H. Impact of dexamethasone concentration on cartilage tissue formation from human synovial derived stem cells in vitro. Cytotechnology 2018, 70, 819–829. [Google Scholar] [CrossRef]
- Kirton, J.P.; Crofts, N.J.; George, S.J.; Brennan, K.; Canfield, A.E. Wnt/beta-catenin signaling stimulates chondrogenic and inhibits adipogenic differentiation of pericytes: Potential relevance to vascular disease? Circ. Res. 2007, 101, 581–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornstein, I.; Alcover, A.; Katzav, S. Vav proteins, masters of the world of cytoskeleton organization. Cell Signal. 2004, 16, 1–11. [Google Scholar] [CrossRef]
- Qu, P.; Wang, L.; Min, Y.; McKennett, L.; Keller, J.R.; Lin, P.C. Vav1 Regulates Mesenchymal Stem Cell Differentiation Decision Between Adipocyte and Chondrocyte via Sirt1. Stem Cells 2016, 34, 1934–1946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Sul, H.S. Pref-1 regulates mesenchymal cell commitment and differentiation through Sox9. Cell Metab. 2009, 9, 287–302. [Google Scholar] [CrossRef] [Green Version]
- Stockl, S.; Bauer, R.J.; Bosserhoff, A.K.; Gottl, C.; Grifka, J.; Grassel, S. Sox9 modulates cell survival and adipogenic differentiation of multipotent adult rat mesenchymal stem cells. J. Cell Sci. 2013, 126, 2890–2902. [Google Scholar] [CrossRef] [Green Version]
- Ushita, M.; Saito, T.; Ikeda, T.; Yano, F.; Higashikawa, A.; Ogata, N.; Chung, U.; Nakamura, K.; Kawaguchi, H. Transcriptional induction of SOX9 by NF-kappaB family member RelA in chondrogenic cells. Osteoarthr. Cartil. 2009, 17, 1065–1075. [Google Scholar] [CrossRef] [Green Version]
- Correa, D.; Somoza, R.A.; Lin, P.; Greenberg, S.; Rom, E.; Duesler, L.; Welter, J.F.; Yayon, A.; Caplan, A.I. Sequential exposure to fibroblast growth factors (FGF) 2, 9 and 18 enhances hMSC chondrogenic differentiation. Osteoarthr. Cartil. 2015, 23, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Kalomoiris, S.; Cicchetto, A.C.; Lakatos, K.; Nolta, J.A.; Fierro, F.A. Fibroblast Growth Factor 2 Regulates High Mobility Group A2 Expression in Human Bone Marrow-Derived Mesenchymal Stem Cells. J. Cell Biochem. 2016, 117, 2128–2137. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Hutley, L.J.; Webster, J.A.; Kim, Y.H.; Liu, D.F.; Newell, F.S.; Widberg, C.H.; Bachmann, A.; Turner, N.; Schmitz-Peiffer, C.; et al. Identification of BMP and activin membrane-bound inhibitor (BAMBI) as a potent negative regulator of adipogenesis and modulator of autocrine/paracrine adipogenic factors. Diabetes 2012, 61, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Boney, C.M.; Gruppuso, P.A.; Faris, R.A.; Frackelton, A.R., Jr. The critical role of Shc in insulin-like growth factor-I-mediated mitogenesis and differentiation in 3T3-L1 preadipocytes. Mol. Endocrinol. 2000, 14, 805–813. [Google Scholar] [CrossRef]
- Starkman, B.G.; Cravero, J.D.; Delcarlo, M.; Loeser, R.F. IGF-I stimulation of proteoglycan synthesis by chondrocytes requires activation of the PI 3-kinase pathway but not ERK MAPK. Biochem. J. 2005, 389, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Chen, S.; Jiang, Q.; Pei, M. Determinants of stem cell lineage differentiation toward chondrogenesis versus adipogenesis. Cell Mol. Life Sci. 2019, 76, 1653–1680. [Google Scholar] [CrossRef] [PubMed]
- Riedt, C.S.; Cifuentes, M.; Stahl, T.; Chowdhury, H.A.; Schlussel, Y.; Shapses, S.A. Overweight postmenopausal women lose bone with moderate weight reduction and 1 g/day calcium intake. J. Bone Miner. Res. 2005, 20, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Cifuentes, M.; Riedt, C.S.; Brolin, R.E.; Field, M.P.; Sherrell, R.M.; Shapses, S.A. Weight loss and calcium intake influence calcium absorption in overweight postmenopausal women. Am. J. Clin. Nutr. 2004, 80, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Migliaccio, S.; Greco, E.A.; Aversa, A.; Lenzi, A. Age-associated (cardio)metabolic diseases and cross-talk between adipose tissue and skeleton: Endocrine aspects. Horm. Mol. Biol. Clin. Investig. 2014, 20, 25–38. [Google Scholar] [CrossRef]
- Migliaccio, S.; Greco, E.A.; Wannenes, F.; Donini, L.M.; Lenzi, A. Adipose, bone and muscle tissues as new endocrine organs: Role of reciprocal regulation for osteoporosis and obesity development. Horm. Mol. Biol. Clin. Investig. 2014, 17, 39–51. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, D.; Baek, M.Y.; Tchah, H.; Kim, Y.S.; Ryoo, E.; Kim, Y.M. Abdominal Subcutaneous Fat Thickness Measured by Ultrasonography Correlates with Hyperlipidemia and Steatohepatitis in Obese Children. Pediatr. Gastroenterol. Hepatol. Nutr. 2015, 18, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.G.; Kim, B.; Soung do, Y.; Vance, T.; Lee, J.S.; Lee, J.Y.; Koo, S.I.; Kim, D.O.; Drissi, H.; Chun, O.K. Relationship between oxidative stress and bone mass in obesity and effects of berry supplementation on bone remodeling in obese male mice: An exploratory study. J. Med. Food 2015, 18, 476–482. [Google Scholar] [CrossRef]
- Ozcivici, E.; Luu, Y.K.; Adler, B.; Qin, Y.X.; Rubin, J.; Judex, S.; Rubin, C.T. Mechanical signals as anabolic agents in bone. Nat. Rev. Rheumatol. 2010, 6, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Gimble, J.M.; Bunnell, B.A.; Chiu, E.S.; Guilak, F. Concise review: Adipose-derived stromal vascular fraction cells and stem cells: Let’s not get lost in translation. Stem Cells 2011, 29, 749–754. [Google Scholar] [CrossRef]
- MacKellar, J.; Cushman, S.W.; Periwal, V. Waves of adipose tissue growth in the genetically obese Zucker fatty rat. PLoS ONE 2010, 5, e8197. [Google Scholar] [CrossRef]
- Kozak, L.P.; Anunciado-Koza, R. UCP1: Its involvement and utility in obesity. Int. J. Obes. 2008, 32, S32–S38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.J.; Enerback, S.; et al. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Deans, C.; Maggert, K.A. What do you mean, “epigenetic”? Genetics 2015, 199, 887–896. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.D.; Le, T.; Fan, G. DNA methylation and its basic function. Neuropsychopharmacology 2013, 38, 23–38. [Google Scholar] [CrossRef] [Green Version]
- Deaton, A.M.; Bird, A. CpG islands and the regulation of transcription. Genes Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef] [Green Version]
- Niculescu, M.D. Nutritional epigenetics. Ilar J. 2012, 53, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Liang, X.; Yang, Q.; Fu, X.; Rogers, C.J.; Wang, B.; Pan, H.; Zhu, M.J.; Nathanielsz, P.W.; Du, M. Maternal obesity epigenetically alters visceral fat progenitor cell properties in male offspring mice. J. Physiol. 2016, 594, 4453–4466. [Google Scholar] [CrossRef] [Green Version]
- Palou, M.; Pico, C.; McKay, J.A.; Sanchez, J.; Priego, T.; Mathers, J.C.; Palou, A. Protective effects of leptin during the suckling period against later obesity may be associated with changes in promoter methylation of the hypothalamic pro-opiomelanocortin gene. Br. J. Nutr. 2011, 106, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Arreguin, A.; Ribot, J.; Musinovic, H.; von Lintig, J.; Palou, A.; Bonet, M.L. Dietary vitamin A impacts DNA methylation patterns of adipogenesis-related genes in suckling rats. Arch. Biochem. Biophys. 2018, 650, 75–84. [Google Scholar] [CrossRef]
- Jacobsen, S.C.; Brons, C.; Bork-Jensen, J.; Ribel-Madsen, R.; Yang, B.; Lara, E.; Hall, E.; Calvanese, V.; Nilsson, E.; Jorgensen, S.W.; et al. Effects of short-term high-fat overfeeding on genome-wide DNA methylation in the skeletal muscle of healthy young men. Diabetologia 2012, 55, 3341–3349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malodobra-Mazur, M.; Cierzniak, A.; Dobosz, T. Oleic acid influences the adipogenesis of 3T3-L1 cells via DNA Methylation and may predispose to obesity and obesity-related disorders. Lipids Health Dis. 2019, 18, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanimura, K.; Suzuki, T.; Vargas, D.; Shibata, H.; Inagaki, T. Epigenetic regulation of beige adipocyte fate by histone methylation. Endocr. J. 2019, 66, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.; Lim, B.; Lodish, H.F. MicroRNAs induced during adipogenesis that accelerate fat cell development are downregulated in obesity. Diabetes 2009, 58, 1050–1057. [Google Scholar] [CrossRef] [Green Version]
- Landrier, J.F.; Derghal, A.; Mounien, L. MicroRNAs in Obesity and Related Metabolic Disorders. Cells 2019, 8, 859. [Google Scholar] [CrossRef] [Green Version]
- Zhong, H.; Ma, M.; Liang, T.; Guo, L. Role of MicroRNAs in Obesity-Induced Metabolic Disorder and Immune Response. J. Immunol. Res. 2018, 2018, 2835761. [Google Scholar] [CrossRef] [Green Version]
- Ortega, F.J.; Mercader, J.M.; Catalan, V.; Moreno-Navarrete, J.M.; Pueyo, N.; Sabater, M.; Gomez-Ambrosi, J.; Anglada, R.; Fernandez-Formoso, J.A.; Ricart, W.; et al. Targeting the circulating microRNA signature of obesity. Clin. Chem. 2013, 59, 781–792. [Google Scholar] [CrossRef] [Green Version]
- Arner, E.; Mejhert, N.; Kulyte, A.; Balwierz, P.J.; Pachkov, M.; Cormont, M.; Lorente-Cebrian, S.; Ehrlund, A.; Laurencikiene, J.; Heden, P.; et al. Adipose tissue microRNAs as regulators of CCL2 production in human obesity. Diabetes 2012, 61, 1986–1993. [Google Scholar] [CrossRef] [Green Version]
- Hulsmans, M.; Holvoet, P. MicroRNAs as early biomarkers in obesity and related metabolic and cardiovascular diseases. Curr. Pharm. Des. 2013, 19, 5704–5717. [Google Scholar] [CrossRef]
- Chien, H.Y.; Lee, T.P.; Chen, C.Y.; Chiu, Y.H.; Lin, Y.C.; Lee, L.S.; Li, W.C. Circulating microRNA as a diagnostic marker in populations with type 2 diabetes mellitus and diabetic complications. J. Chin. Med. Assoc. 2015, 78, 204–211. [Google Scholar] [CrossRef] [Green Version]
- Mittelbrunn, M.; Sanchez-Madrid, F. Intercellular communication: Diverse structures for exchange of genetic information. Nat. Rev. Mol. Cell Biol. 2012, 13, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Castano, C.; Kalko, S.; Novials, A.; Parrizas, M. Obesity-associated exosomal miRNAs modulate glucose and lipid metabolism in mice. Proc. Natl. Acad Sci. USA 2018, 115, 12158–12163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez-Dominguez, J.R.; Bai, Z.; Xu, D.; Yuan, B.; Lo, K.A.; Yoon, M.J.; Lim, Y.C.; Knoll, M.; Slavov, N.; Chen, S.; et al. De Novo Reconstruction of Adipose Tissue Transcriptomes Reveals Long Non-coding RNA Regulators of Brown Adipocyte Development. Cell Metab. 2015, 21, 764–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, C.; Guo, X. The clinical potential of circulating microRNAs in obesity. Nat. Rev. Endocrinol. 2019, 15, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, Z.; Sun, W.; Yang, G.; Xing, C.; Yuan, L. Delivery Efficacy Differences of Intravenous and Intraperitoneal Injection of Exosomes:Perspectives from Tracking Dye Labeled and MiRNA Encapsulated Exosomes. Curr. Drug Deliv. 2020. [Google Scholar] [CrossRef] [PubMed]
- DeLuca, H.F. Overview of general physiologic features and functions of vitamin D. Am. J. Clin. Nutr. 2004, 80, 1689S–1696S. [Google Scholar] [CrossRef] [Green Version]
- Wacker, M.; Holick, M.F. Vitamin D - effects on skeletal and extraskeletal health and the need for supplementation. Nutrients 2013, 5, 111–148. [Google Scholar] [CrossRef] [Green Version]
- Migliaccio, S.; Di Nisio, A.; Mele, C.; Scappaticcio, L.; Savastano, S.; Colao, A. Obesity and hypovitaminosis D: Causality or casualty? Int. J. Obes. Suppl. 2019, 9, 20–31. [Google Scholar] [CrossRef]
- Pereira-Santos, M.; Costa, P.R.; Assis, A.M.; Santos, C.A.; Santos, D.B. Obesity and vitamin D deficiency: A systematic review and meta-analysis. Obes. Rev. 2015, 16, 341–349. [Google Scholar] [CrossRef]
- Need, A.G.; Morris, H.A.; Horowitz, M.; Nordin, C. Effects of skin thickness, age, body fat, and sunlight on serum 25-hydroxyvitamin D. Am. J. Clin. Nutr. 1993, 58, 882–885. [Google Scholar] [CrossRef]
- Wortsman, J.; Matsuoka, L.Y.; Chen, T.C.; Lu, Z.; Holick, M.F. Decreased bioavailability of vitamin D in obesity. Am. J. Clin. Nutr. 2000, 72, 690–693. [Google Scholar] [CrossRef]
- Carrelli, A.; Bucovsky, M.; Horst, R.; Cremers, S.; Zhang, C.; Bessler, M.; Schrope, B.; Evanko, J.; Blanco, J.; Silverberg, S.J.; et al. Vitamin D Storage in Adipose Tissue of Obese and Normal Weight Women. J. Bone Miner. Res. 2017, 32, 237–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, M.A. Physiological functions of Vitamin D in adipose tissue. J. Steroid. Biochem. Mol. Biol. 2017, 165, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Lou, Y.; Kong, J. VDR regulates energy metabolism by modulating remodeling in adipose tissue. Eur. J. Pharmacol. 2019, 865, 172761. [Google Scholar] [CrossRef] [PubMed]
- Honek, J.; Lim, S.; Fischer, C.; Iwamoto, H.; Seki, T.; Cao, Y. Brown adipose tissue, thermogenesis, angiogenesis: Pathophysiological aspects. Horm. Mol. Biol. Clin. Investig. 2014, 19, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Dalle Carbonare, L.; Valenti, M.T.; Del Forno, F.; Piacentini, G.; Pietrobelli, A. Vitamin D Daily versus Monthly Administration: Bone Turnover and Adipose Tissue Influences. Nutrients 2018, 10, 1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cembranel, F.; d’Orsi, E.; Jakovljevic Pudla Wagner, K.; Weber Corseuil Giehl, M.; Moreno, Y.M.F.; Gonzalez-Chica, D.A. Obesity and 25(OH)D Serum Concentration Are More Important than Vitamin D Intake for Changes in Nutritional Status Indicators: A Population-Based Longitudinal Study in a State Capital City in Southern Brazil. Nutrients 2019, 11, 2366. [Google Scholar] [CrossRef] [Green Version]
- Moschonis, G.; Androutsos, O.; Hulshof, T.; Dracopoulou, M.; Chrousos, G.P.; Manios, Y. Vitamin D insufficiency is associated with insulin resistance independently of obesity in primary schoolchildren. The healthy growth study. Pediatr Diabetes 2018, 19, 866–873. [Google Scholar] [CrossRef]
- Valenti, M.T.; Gianluigi, D.; Mottes, M. Effects of physical exercise on the prevention of stem cells senescence. Stem Cell Rev. Rep. 2019, 16, 33–40. [Google Scholar] [CrossRef]
- Dalle Carbonare, L.; Manfredi, M.; Caviglia, G.; Conte, E.; Robotti, E.; Marengo, E.; Cheri, S.; Zamboni, F.; Gabbiani, D.; Deiana, M.; et al. Can half-marathon affect overall health? The yin-yang of sport. J. Prot. 2018, 170, 80–87. [Google Scholar] [CrossRef]
- Maredziak, M.; Smieszek, A.; Chrzastek, K.; Basinska, K.; Marycz, K. Physical Activity Increases the Total Number of Bone-Marrow-Derived Mesenchymal Stem Cells, Enhances Their Osteogenic Potential, and Inhibits Their Adipogenic Properties. Stem Cells Int. 2015, 2015, 379093. [Google Scholar] [CrossRef] [PubMed]
- Sen, B.; Xie, Z.; Case, N.; Ma, M.; Rubin, C.; Rubin, J. Mechanical strain inhibits adipogenesis in mesenchymal stem cells by stimulating a durable beta-catenin signal. Endocrinology 2008, 149, 6065–6075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Recommandation of Physical Activity for Health 2010, 1-58. Available online: http://www.who.int/dietphysicalactivity/publications/9789241599979/en/. (accessed on 2 February 2020).
- Grazioli, E.; Dimauro, I.; Mercatelli, N.; Wang, G.; Pitsiladis, Y.; Di Luigi, L.; Caporossi, D. Physical activity in the prevention of human diseases: Role of epigenetic modifications. BMC Genom. 2017, 18, 802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moholdt, T.; Wisloff, U.; Lydersen, S.; Nauman, J. Current physical activity guidelines for health are insufficient to mitigate long-term weight gain: More data in the fitness versus fatness debate (The HUNT study, Norway). Br. J. Sports Med. 2014, 48, 1489–1496. [Google Scholar] [CrossRef] [Green Version]
- Jakicic, J.M. The effect of physical activity on body weight. Obesity 2009, 17, S34–S38. [Google Scholar] [CrossRef]
- Donnelly, J.E.; Blair, S.N.; Jakicic, J.M.; Manore, M.M.; Rankin, J.W.; Smith, B.K.; American College of Sports M: American College of Sports Medicine Position Stand. Appropriate physical activity intervention strategies for weight loss and prevention of weight regain for adults. Med. Sci. Sports Exerc. 2009, 41, 459–471. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [Green Version]
- Campbell, S.C.; Wisniewski, P.J. 2nd: Exercise is a Novel Promoter of Intestinal Health and Microbial Diversity. Exerc. Sport Sci. Rev. 2017, 45, 41–47. [Google Scholar] [CrossRef]
- Codella, R.; Luzi, L.; Terruzzi, I. Exercise has the guts: How physical activity may positively modulate gut microbiota in chronic and immune-based diseases. Dig. Liver Dis. 2018, 50, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.C.; LePard, K.J.; Kwak, J.W.; Stancukas, M.C.; Laskowski, S.; Dougherty, J.; Moulton, L.; Glawe, A.; Wang, Y.; Leone, V.; et al. Exercise prevents weight gain and alters the gut microbiota in a mouse model of high fat diet-induced obesity. PLoS ONE 2014, 9, e92193. [Google Scholar] [CrossRef]
- Remely, M.; Tesar, I.; Hippe, B.; Gnauer, S.; Rust, P.; Haslberger, A.G. Gut microbiota composition correlates with changes in body fat content due to weight loss. Benef. Microbes. 2015, 6, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I. Host energy regulation via SCFAs receptors, as dietary nutrition sensors, by gut microbiota. Yakugaku Zasshi 2014, 134, 1037–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, B.S.; Shaito, A.; Motoike, T.; Rey, F.E.; Backhed, F.; Manchester, J.K.; Hammer, R.E.; Williams, S.C.; Crowley, J.; Yanagisawa, M.; et al. Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor, Gpr41. Proc. Natl. Acad Sci. USA 2008, 105, 16767–16772. [Google Scholar] [CrossRef] [Green Version]
- Deiana, M.; Malerba, G.; Dalle Carbonare, L.; Cheri, S.; Patuzzo, C.; Tsenov, G.; Moron Dalla Tor, L.; Mori, A.; Saviola, G.; Zipeto, D.; et al. Physical Activity Prevents Cartilage Degradation: A Metabolomics Study Pinpoints the Involvement of Vitamin B6. Cells 2019, 8, 1374. [Google Scholar] [CrossRef] [Green Version]
- Said, Z.M.; Subramanian, V.S.; Vaziri, N.D.; Said, H.M. Pyridoxine uptake by colonocytes: A specific and regulated carrier-mediated process. Am. J. Physiol. Cell Physiol. 2008, 294, C1192–C1197. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Navarrete, J.M.; Jove, M.; Ortega, F.; Xifra, G.; Ricart, W.; Obis, E.; Pamplona, R.; Portero-Otin, M.; Fernandez-Real, J.M. Metabolomics uncovers the role of adipose tissue PDXK in adipogenesis and systemic insulin sensitivity. Diabetologia 2016, 59, 822–832. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, P.; Zhao, Z.H.; Zhang, Y.; Ma, Z.M.; Wang, S.X. Vitamin B6 Prevents Endothelial Dysfunction, Insulin Resistance, and Hepatic Lipid Accumulation in Apoe (-/-) Mice Fed with High-Fat Diet. J. Diabetes Res. 2016, 2016, 1748065. [Google Scholar] [CrossRef] [Green Version]
- Riebe, D. ACSM’s Guidelines for Exercise Testing and Prescription, 10th ed.; Wolters Kluwer/Lippincott Williams & Wilkins Health: Philadelphia, PA, USA, 2017; pp. 117–118. [Google Scholar]
- Miller, W.C.; Wallace, J.P.; Eggert, K.E. Predicting max HR and the HR-VO2 relationship for exercise prescription in obesity. Med. Sci. Sports Exerc. 1993, 25, 1077–1081. [Google Scholar] [CrossRef]
- Zhu, N.; Suarez-Lopez, J.R.; Sidney, S.; Sternfeld, B.; Schreiner, P.J.; Carnethon, M.R.; Lewis, C.E.; Crow, R.S.; Bouchard, C.; Haskell, W.L.; et al. Longitudinal examination of age-predicted symptom-limited exercise maximum HR. Med. Sci. Sports Exerc. 2010, 42, 1519–1527. [Google Scholar] [CrossRef] [Green Version]
- Ratter, J.; Radlinger, L.; Lucas, C. Several submaximal exercise tests are reliable, valid and acceptable in people with chronic pain, fibromyalgia or chronic fatigue: A systematic review. J. Physiother. 2014, 60, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Lifetime Physical Fitness and Wellness Di Wener Hoeger, Sharon Hoeger 2007. Available online: http://college.cengage.com/health/course360/fitness_and_wellness_1111764867/ebook/hoeger_1111990018_ch03.pdf (accessed on 2 February 2020).
- Wen, C.P.; Wai, J.P.; Tsai, M.K.; Yang, Y.C.; Cheng, T.Y.; Lee, M.C.; Chan, H.T.; Tsao, C.K.; Tsai, S.P.; Wu, X. Minimum amount of physical activity for reduced mortality and extended life expectancy: A prospective cohort study. Lancet 2011, 378, 1244–1253. [Google Scholar] [CrossRef]
- Warden, S.J.; Hurst, J.A.; Sanders, M.S.; Turner, C.H.; Burr, D.B.; Li, J. Bone adaptation to a mechanical loading program significantly increases skeletal fatigue resistance. J Bone Min. Res 2005, 20, 809–816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, F.O.; Gomes-Gatto, C.D.V.; Oishi, J.C.; Lino, A.D.S.; Stotzer, U.S.; Rodrigues, M.F.C.; Gatti da Silva, G.H.; Selistre-de-Araujo, H.S. Physical training improves visceral adipose tissue health by remodelling extracellular matrix in rats with estrogen absence: A gene expression analysis. Int. J. Exp. Pathol. 2017, 98, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Pagnotti, G.M.; Styner, M.; Uzer, G.; Patel, V.S.; Wright, L.E.; Ness, K.K.; Guise, T.A.; Rubin, J.; Rubin, C.T. Combating osteoporosis and obesity with exercise: Leveraging cell mechanosensitivity. Nat Rev Endocrinol 2019, 15, 339–355. [Google Scholar] [CrossRef]
- Dalle Carbonare, L.; Mottes, M.; Cheri, M.; Deiana, F.; Zamboni, D.; Gabbiani, F.; Schena, G.L.; Salvagno, G.; Lippi, M.; Valenti, T. Increased Gene Expression of RUNX2 and SOX9 in MesenchymalCirculating Progenitors Is Associated with Autophagy duringPhysical Activity. Oxidative Med. Cell. Oxid Med. Cell. Longev. 2019. [Google Scholar]
- Liu, M.; Xie, S.; Liu, W.; Li, J.; Li, C.; Huang, W.; Li, H.; Song, J.; Zhang, H. Mechanism of SEMA3G knockdown-mediated attenuation of high-fat diet-induced obesity. J. Endocrinol. 2019. [Google Scholar] [CrossRef]
- Bostrom, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Bostrom, E.A.; Choi, J.H.; Long, J.Z. A PGC1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Y.; Wang, F.; Donelan, W.; Zona, M.C.; Li, S.; Reeves, W.; Ding, Y.; Tang, D.; Yang, L. Effects of irisin on the differentiation and browning of human visceral white adipocytes. Am. J. Transl. Res. 2019, 11, 7410–7421. [Google Scholar]
- Valenti, M.T.; Dalle Carbonare, L.; Mottes, M. Role of microRNAs in progenitor cell commitment and osteogenic differentiation in health and disease (Review). Int. J. Mol. Med. 2018, 41, 2441–2449. [Google Scholar] [CrossRef] [Green Version]
- Valenti, M.T.; Deiana, M.; Cheri, S.; Dotta, M.; Zamboni, F.; Gabbiani, D.; Schena, F.; Dalle Carbonare, L.; Mottes, M. Physical Exercise Modulates miR-21-5p, miR-129-5p, miR-378-5p, and miR-188-5p Expression in Progenitor Cells Promoting Osteogenesis. Cells 2019, 8. [Google Scholar] [CrossRef] [Green Version]
- Li, C.J.; Cheng, P.; Liang, M.K.; Chen, Y.S.; Lu, Q.; Wang, J.Y.; Xia, Z.Y.; Zhou, H.D.; Cao, X.; Xie, H.; et al. MicroRNA-188 regulates age-related switch between osteoblast and adipocyte differentiation. J. Clin. Investig. 2015, 125, 1509–1522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Loan, M.D. Body fat distribution from subcutaneous to intraabdominal: A perspective. Am. J. Clin. Nutr. 1996, 64, 787–788. [Google Scholar] [CrossRef] [PubMed]
- Piche, M.E.; Poirier, P.; Lemieux, I.; Despres, J.P. Overview of Epidemiology and Contribution of Obesity and Body Fat Distribution to Cardiovascular Disease: An Update. Prog. Cardiovasc. Dis. 2018, 61, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.J.; Braunschweig, C.A. Prevalence of Sarcopenia and Associated Outcomes in the Clinical Setting. Nutr. Clin. Pr. 2016, 31, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Pietrobelli, A.; Agosti, M.; Zuccotti, G. Putting the Barker Theory into the Future: Time to Act on Preventing Pediatric Obesity. Int. J. Env. Res. Public Health 2016, 13. [Google Scholar] [CrossRef] [Green Version]
- Lemos, T.; Gallagher, D. Current body composition measurement techniques. Curr. Opin. Endocrinol. Diabetes Obes. 2017, 24, 310–314. [Google Scholar] [CrossRef]
- Haslam, D.W.; James, W.P. Obesity. Lancet 2005, 366, 1197–1209. [Google Scholar] [CrossRef]
- Starling, A.P.; Brinton, J.T.; Glueck, D.H.; Shapiro, A.L.; Harrod, C.S.; Lynch, A.M.; Siega-Riz, A.M.; Dabelea, D. Associations of maternal BMI and gestational weight gain with neonatal adiposity in the Healthy Start study. Am. J. Clin. Nutr. 2015, 101, 302–309. [Google Scholar] [CrossRef] [Green Version]
- Lawlor, D.A.; Lichtenstein, P.; Fraser, A.; Langstrom, N. Does maternal weight gain in pregnancy have long-term effects on offspring adiposity? A sibling study in a prospective cohort of 146,894 men from 136,050 families. Am. J. Clin. Nutr. 2011, 94, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Larque, E.; Labayen, I.; Flodmark, C.E.; Lissau, I.; Czernin, S.; Moreno, L.A.; Pietrobelli, A.; Widhalm, K. From conception to infancy - early risk factors for childhood obesity. Nat. Rev. Endocrinol. 2019, 15, 456–478. [Google Scholar] [CrossRef]
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980-2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Pietrobelli, A.; Agosti, M.; MeNu, G. Nutrition in the First 1000 Days: Ten Practices to Minimize Obesity Emerging from Published Science. Int. J. Env. Res. Public Health 2017, 14, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slawik, M.; Vidal-Puig, A.J. Lipotoxicity, overnutrition and energy metabolism in aging. Ageing Res. Rev. 2006, 5, 144–164. [Google Scholar] [CrossRef] [PubMed]
- Wilmot, E.G.; Edwardson, C.L.; Achana, F.A.; Davies, M.J.; Gorely, T.; Gray, L.J.; Khunti, K.; Yates, T.; Biddle, S.J. Sedentary time in adults and the association with diabetes, cardiovascular disease and death: Systematic review and meta-analysis. Diabetologia 2012, 55, 2895–2905. [Google Scholar] [CrossRef] [PubMed]
- Jura, M.; Kozak, L.P. Obesity and related consequences to ageing. Age (Dordr) 2016, 38, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharifi, M.; Franz, C.; Horan, C.M.; Giles, C.M.; Long, M.W.; Ward, Z.J.; Resch, S.C.; Marshall, R.; Gortmaker, S.L.; Taveras, E.M. Cost-Effectiveness of a Clinical Childhood Obesity Intervention. Pediatrics 2017, 140, 5. [Google Scholar] [CrossRef] [Green Version]
- Konnopka, A.; Dobroschke, A.; Lehnert, T.; Konig, H.H. The Costs of Overweight and Obesity: A Systematic Review. Gesundheitswesen 2018, 80, 471–481. [Google Scholar]
- Seidell, J.C.; Halberstadt, J. The global burden of obesity and the challenges of prevention. Ann. Nutr. Metab. 2015, 66, 7–12. [Google Scholar] [CrossRef]
- Liang, X.; Yang, Q.; Zhang, L.; Maricelli, J.W.; Rodgers, B.D.; Zhu, M.J.; Du, M. Maternal high-fat diet during lactation impairs thermogenic function of brown adipose tissue in offspring mice. Sci. Rep. 2016, 6, 34345. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
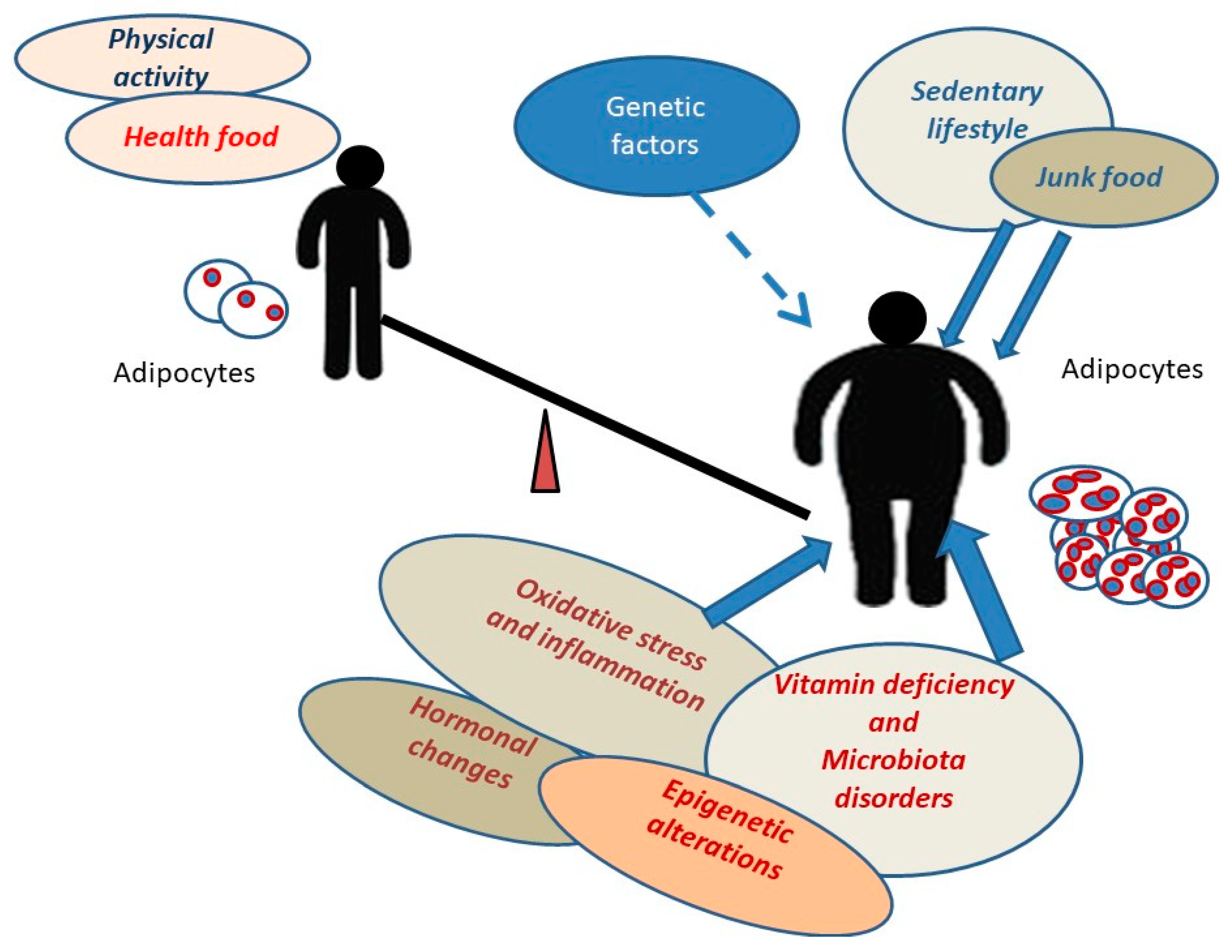{kind=link}
| Author | Topic | Reference |
|---|---|---|
| Lefterova et al. | PPARgamma and adipogenesis (2014) | [4] |
| Valenti et al. | Ox-PAPCs and Differentiation of MSCs (2011) | [8] |
| Qu et al. | Vav1 and MSCs (2016) | [13] |
| Stockl et al. | SOX 9 and MSC (2013) | [15] |
| Boney et al. | Shc and preadipocytes (2000) | [20] |
| Zhou et al. | Chondrogenesis versus adipogenesis (2019) | [22] |
| Liang et al. | Maternal HFD and Brown tissue (2016) | [122] |
| Maredziak et al. | Physical activity and adipogenesis (2015) | [71] |
| Liang et al. | Maternal obesity and epigenetics (2016) | [38] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valenti, M.T.; Pietrobelli, A.; Romanelli, M.G.; Franzolin, E.; Malerba, G.; Zipeto, D.; Mottes, M.; Dalle Carbonare, L. Molecular and Lifestyle Factors Modulating Obesity Disease. Biomedicines 2020, 8, 46. https://doi.org/10.3390/biomedicines8030046
Valenti MT, Pietrobelli A, Romanelli MG, Franzolin E, Malerba G, Zipeto D, Mottes M, Dalle Carbonare L. Molecular and Lifestyle Factors Modulating Obesity Disease. Biomedicines. 2020; 8(3):46. https://doi.org/10.3390/biomedicines8030046
Chicago/Turabian StyleValenti, Maria Teresa, Angelo Pietrobelli, Maria Grazia Romanelli, Elia Franzolin, Giovanni Malerba, Donato Zipeto, Monica Mottes, and Luca Dalle Carbonare. 2020. "Molecular and Lifestyle Factors Modulating Obesity Disease" Biomedicines 8, no. 3: 46. https://doi.org/10.3390/biomedicines8030046
APA StyleValenti, M. T., Pietrobelli, A., Romanelli, M. G., Franzolin, E., Malerba, G., Zipeto, D., Mottes, M., & Dalle Carbonare, L. (2020). Molecular and Lifestyle Factors Modulating Obesity Disease. Biomedicines, 8(3), 46. https://doi.org/10.3390/biomedicines8030046