The Biological Role of Apurinic/Apyrimidinic Endonuclease1/Redox Factor-1 as a Therapeutic Target for Vascular Inflammation and as a Serologic Biomarker



Abstract
:
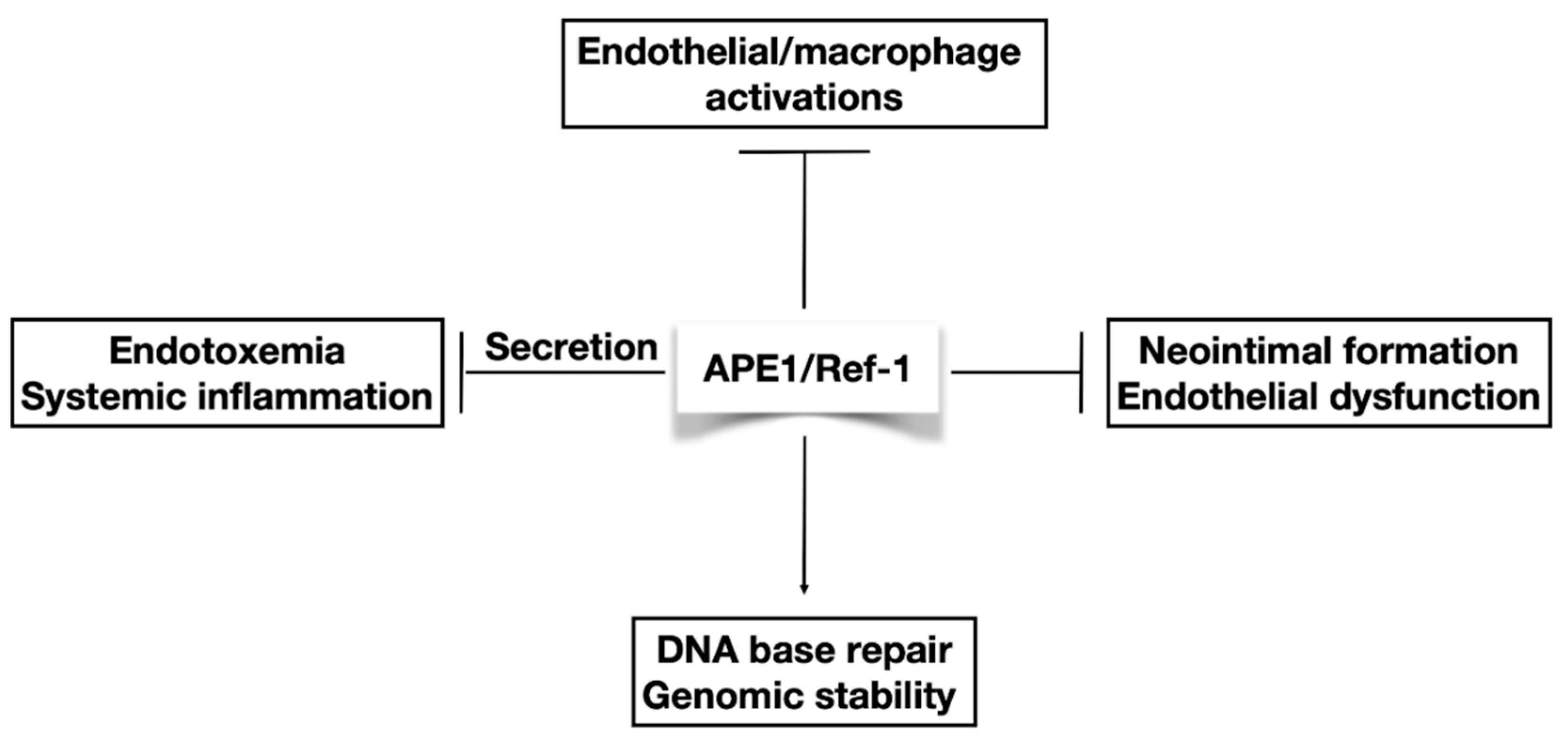
1. Endothelial Dysfunction and Vascular Inflammation
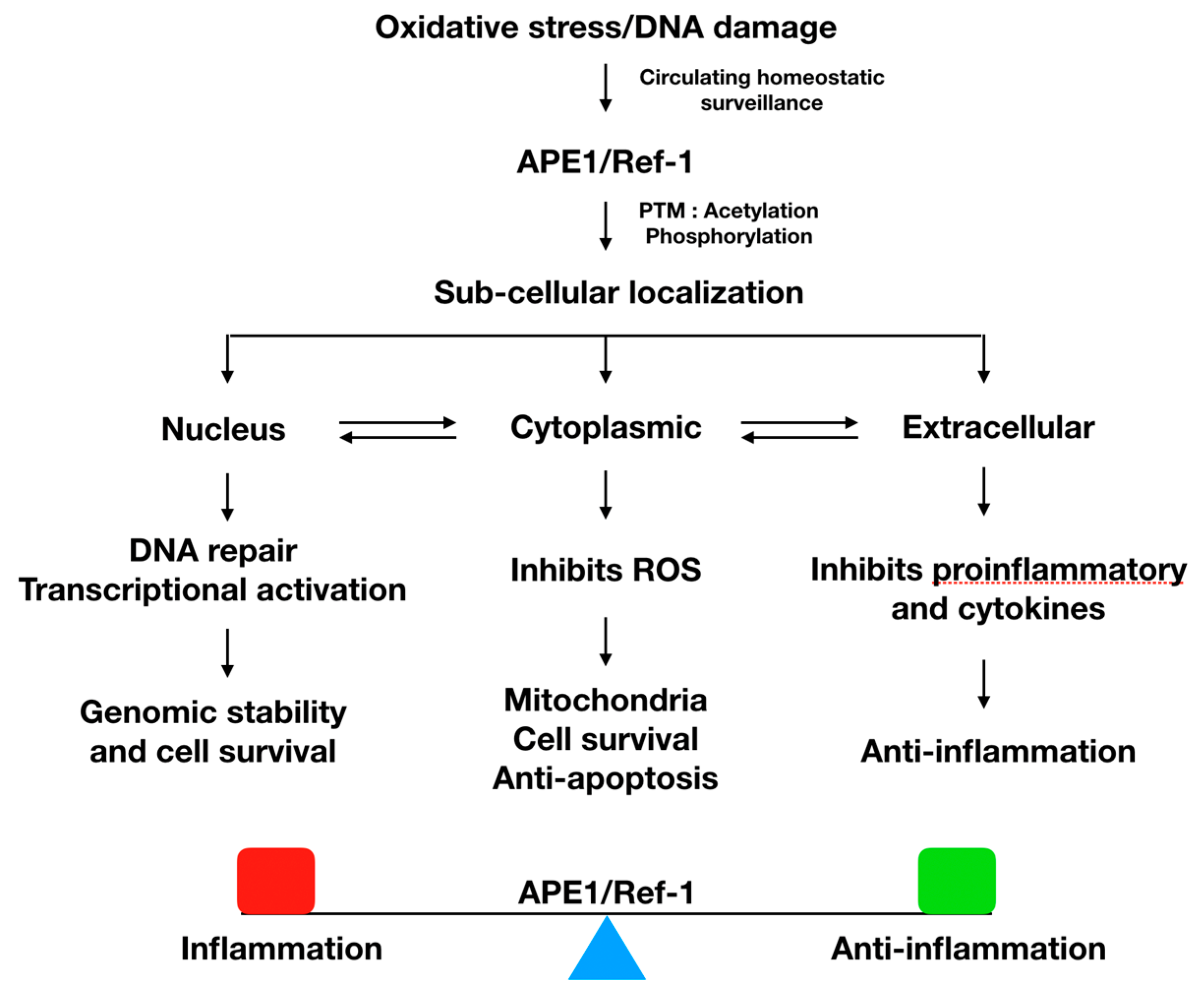
2. APE1/Ref-1 Protein Has Several Cellular Functions
2.1. Nuclear Function of APE1/Ref-1
2.2. Cytoplasmic Function of APE1/Ref-1
2.3. Extracellular Function of APE1/Ref-1
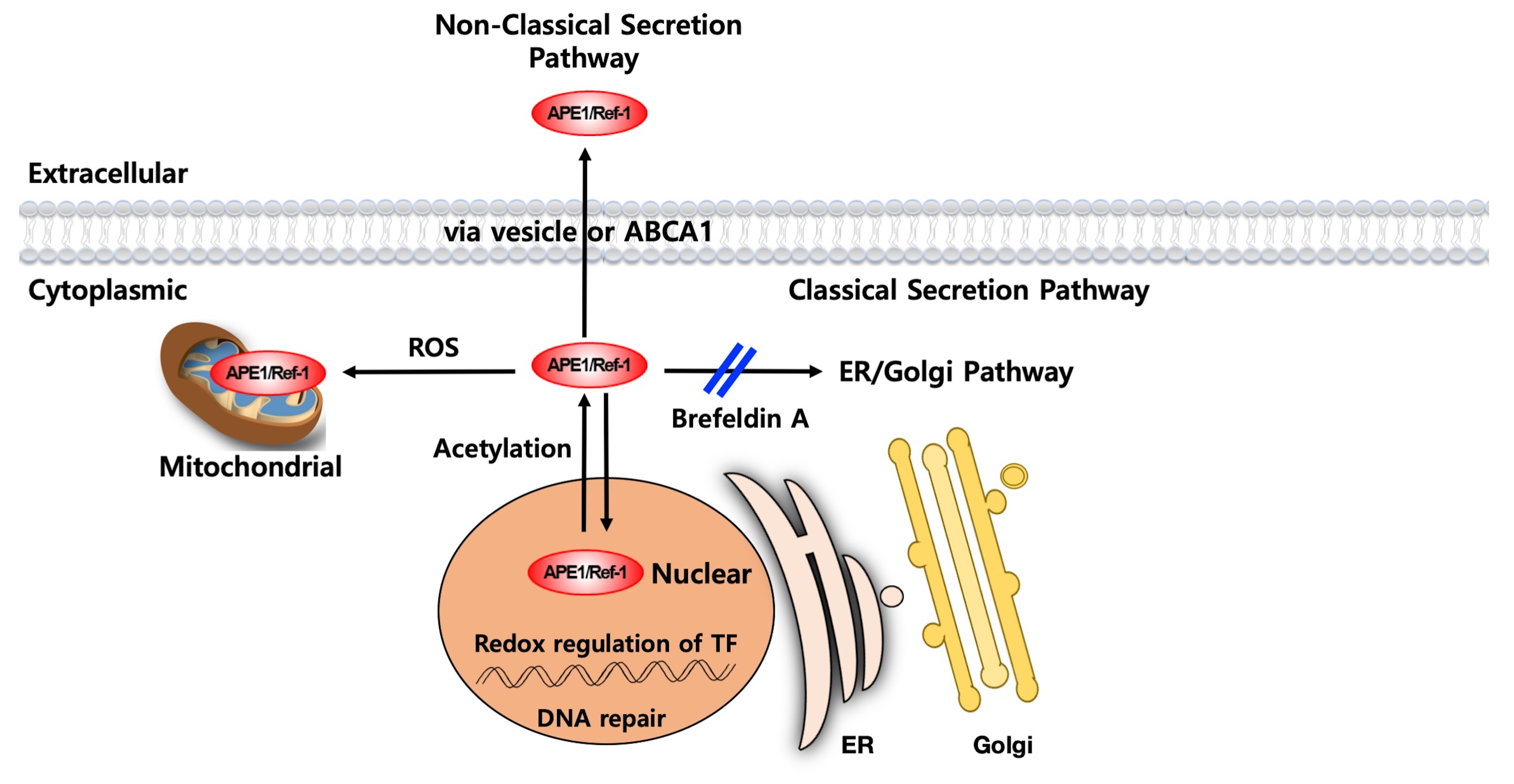
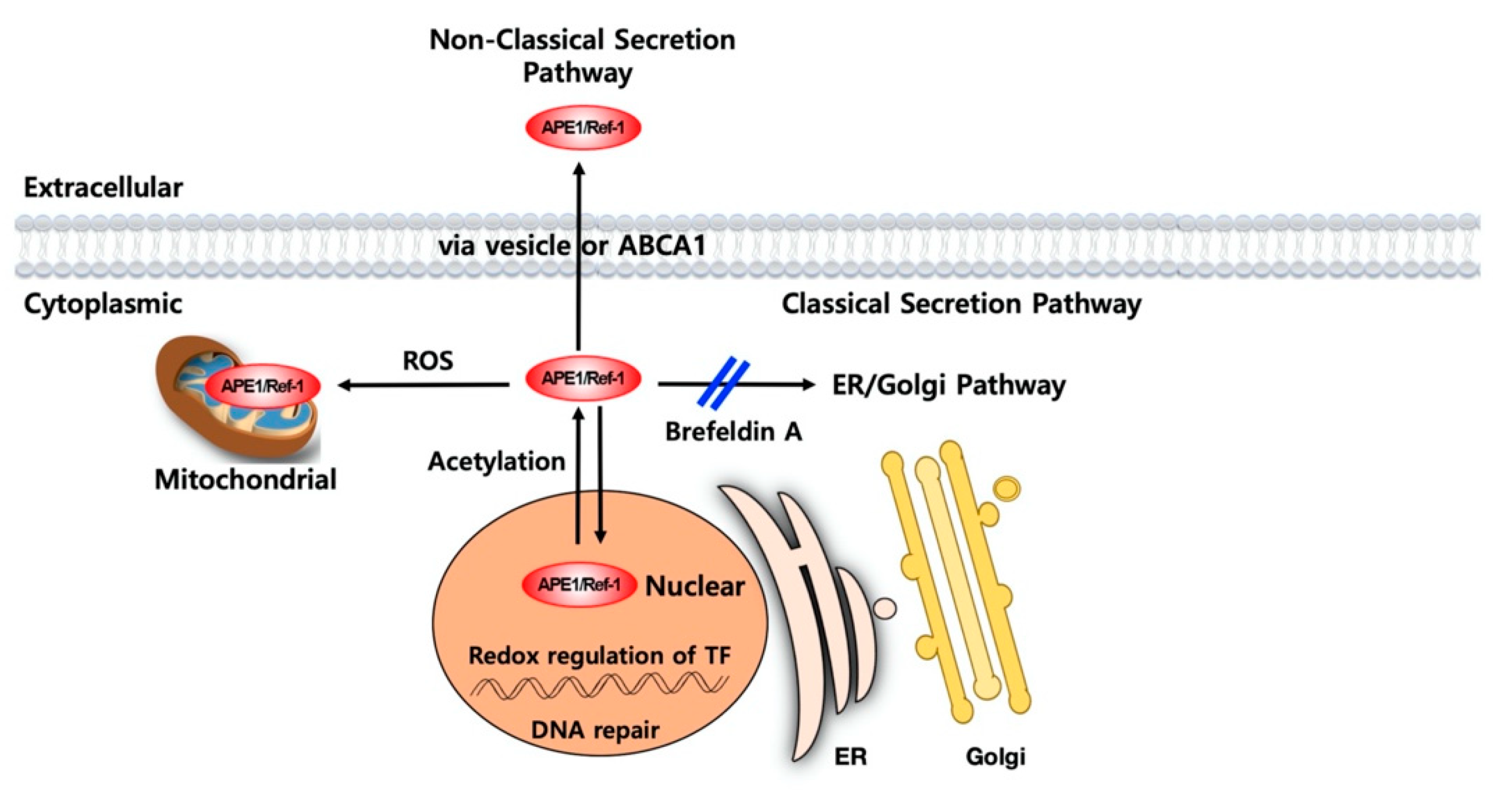
3. Mechanism of APE1/Ref-1 Secretion
4. Extracellular APE1/Ref-1 as a Serological Biomarker
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Liao, J.K. Linking endothelial dysfunction with endothelial cell activation. J. Clin. Investig. 2013, 123, 540–541. [Google Scholar] [CrossRef]
- Ross, R. Atherosclerosis--an inflammatory disease. N. Engl. J. Med. 1999, 340, 115–126. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Irani, K. Oxidant signaling in vascular cell growth, death, and survival: A review of the roles of reactive oxygen species in smooth muscle and endothelial cell mitogenic and apoptotic signaling. Circ. Res. 2000, 87, 179–183. [Google Scholar] [CrossRef]
- Gonzalez-Amaro, R.; Diaz-Gonzalez, F.; Sanchez-Madrid, F. Adhesion molecules in inflammatory diseases. Drugs 1998, 56, 977–988. [Google Scholar] [CrossRef]
- Zundler, S.; Becker, E.; Weidinger, C.; Siegmund, B. Anti-Adhesion Therapies in Inflammatory Bowel Disease-Molecular and Clinical Aspects. Front. Immunol. 2017, 8, 891. [Google Scholar] [CrossRef] [Green Version]
- Galkina, E.; Ley, K. Vascular adhesion molecules in atherosclerosis. Arter. Thromb. Vasc. Biol. 2007, 27, 2292–2301. [Google Scholar] [CrossRef] [PubMed]
- Galkina, E.; Ley, K. Immune and inflammatory mechanisms of atherosclerosis. Annu. Rev. Immunol. 2009, 27, 165–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.B.; Mengi, S.A.; Xu, Y.J.; Arneja, A.S.; Dhalla, N.S. Pathogenesis of atherosclerosis: A multifactorial process. Exp. Clin. Cardiol. 2002, 7, 40–53. [Google Scholar] [PubMed]
- Yu, X.H.; Fu, Y.C.; Zhang, D.W.; Yin, K.; Tang, C.K. Foam cells in atherosclerosis. Clin. Chim. Acta 2013, 424, 245–252. [Google Scholar] [CrossRef] [Green Version]
- Ohta, H.; Wada, H.; Niwa, T.; Kirii, H.; Iwamoto, N.; Fujii, H.; Saito, K.; Sekikawa, K.; Seishima, M. Disruption of tumor necrosis factor-alpha gene diminishes the development of atherosclerosis in ApoE-deficient mice. Atherosclerosis 2005, 180, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Bluml, S.; Scheinecker, C.; Smolen, J.S.; Redlich, K. Targeting TNF receptors in rheumatoid arthritis. Int. Immunol. 2012, 24, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridker, P.M. Anticytokine Agents: Targeting Interleukin Signaling Pathways for the Treatment of Atherothrombosis. Circ. Res. 2019, 124, 437–450. [Google Scholar] [CrossRef] [PubMed]
- Yvan-Charvet, L.; Wang, N.; Tall, A.R. Role of HDL, ABCA1, and ABCG1 transporters in cholesterol efflux and immune responses. Arter. Thromb. Vasc. Biol. 2010, 30, 139–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yvan-Charvet, L.; Ranalletta, M.; Wang, N.; Han, S.; Terasaka, N.; Li, R.; Welch, C.; Tall, A.R. Combined deficiency of ABCA1 and ABCG1 promotes foam cell accumulation and accelerates atherosclerosis in mice. J. Clin. Investig. 2007, 117, 3900–3908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerterp, M.; Tsuchiya, K.; Tattersall, I.W.; Fotakis, P.; Bochem, A.E.; Molusky, M.M.; Ntonga, V.; Abramowicz, S.; Parks, J.S.; Welch, C.L.; et al. Deficiency of ATP-Binding Cassette Transporters A1 and G1 in Endothelial Cells Accelerates Atherosclerosis in Mice. Arter. Thromb. Vasc. Biol. 2016, 36, 1328–1337. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.; Joo, H.K.; Jeon, B.H. Dynamic Regulation of APE1/Ref-1 as a Therapeutic Target Protein. Chonnam Med. J. 2016, 52, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.S.; Son, S.J.; Kim, E.K.; Kim, S.N.; Yoo, D.G.; Kim, H.S.; Ryoo, S.W.; Lee, S.D.; Irani, K.; Jeon, B.H. Apurinic/apyrimidinic endonuclease1/redox factor-1 inhibits monocyte adhesion in endothelial cells. Cardiovasc. Res. 2006, 69, 520–526. [Google Scholar] [CrossRef]
- Jeon, B.H.; Gupta, G.; Park, Y.C.; Qi, B.; Haile, A.; Khanday, F.A.; Liu, Y.X.; Kim, J.M.; Ozaki, M.; White, A.R.; et al. Apurinic/apyrimidinic endonuclease 1 regulates endothelial NO production and vascular tone. Circ. Res. 2004, 95, 902–910. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, D.L.; MacInnes, M.A.; Takiguchi, Y.; Purtymun, P.E.; Henrie, M.; Flannery, M.; Meneses, J.; Pedersen, R.A.; Chen, D.J. A murine AP-endonuclease gene-targeted deficiency with post-implantation embryonic progression and ionizing radiation sensitivity. Mutat. Res. 1998, 409, 17–29. [Google Scholar] [CrossRef]
- Xanthoudakis, S.; Smeyne, R.J.; Wallace, J.D.; Curran, T. The redox/DNA repair protein, Ref-1, is essential for early embryonic development in mice. Proc. Natl. Acad. Sci. USA 1996, 93, 8919–8923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, H.K.; Lee, Y.R.; Lee, E.O.; Park, M.S.; Choi, S.; Kim, C.S.; Park, J.B.; Jeon, B.H. The extracellular role of Ref-1 as anti-inflammatory function in lipopolysaccharide-induced septic mice. Free Radic. Biol. Med. 2019, 139, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.M.; Jeon, B.H.; Won, K.J.; Lee, C.K.; Park, T.K.; Choi, W.S.; Bae, Y.M.; Kim, H.S.; Lee, S.K.; Park, S.H.; et al. Gene transfer of redox factor-1 inhibits neointimal formation: Involvement of platelet-derived growth factor-beta receptor signaling via the inhibition of the reactive oxygen species-mediated Syk pathway. Circ. Res. 2009, 104, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakolyris, S.; Kaklamanis, L.; Giatromanolaki, A.; Koukourakis, M.; Hickson, I.D.; Barzilay, G.; Turley, H.; Leek, R.D.; Kanavaros, P.; Georgoulias, V.; et al. Expression and subcellular localization of human AP endonuclease 1 (HAP1/Ref-1) protein: A basis for its role in human disease. Histopathology 1998, 33, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, A.M.; Freudenthal, B.D. APE1: A skilled nucleic acid surgeon. DNA Repair 2018, 71, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Luo, M.; Kelley, M.R. Human apurinic endonuclease 1 (APE1) expression and prognostic significance in osteosarcoma: Enhanced sensitivity of osteosarcoma to DNA damaging agents using silencing RNA APE1 expression inhibition. Mol. Cancer 2004, 3, 679–686. [Google Scholar]
- Luo, M.; Kelley, M.R. Inhibition of the human apurinic/apyrimidinic endonuclease (APE1) repair activity and sensitization of breast cancer cells to DNA alkylating agents with lucanthone. Anticancer Res. 2004, 24, 2127–2134. [Google Scholar]
- Luo, M.; Zhang, J.; He, H.; Su, D.; Chen, Q.; Gross, M.L.; Kelley, M.R.; Georgiadis, M.M. Characterization of the redox activity and disulfide bond formation in apurinic/apyrimidinic endonuclease. Biochemistry 2012, 51, 695–705. [Google Scholar] [CrossRef] [Green Version]
- Park, M.S.; Kim, C.S.; Joo, H.K.; Lee, Y.R.; Kang, G.; Kim, S.J.; Choi, S.; Lee, S.D.; Park, J.B.; Jeon, B.H. Cytoplasmic localization and redox cysteine residue of APE1/Ref-1 are associated with its anti-inflammatory activity in cultured endothelial cells. Mol. Cells 2013, 36, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Proudfoot, D.; Skepper, J.N.; Hegyi, L.; Bennett, M.R.; Shanahan, C.M.; Weissberg, P.L. Apoptosis regulates human vascular calcification in vitro: Evidence for initiation of vascular calcification by apoptotic bodies. Circ. Res. 2000, 87, 1055–1062. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; Lee, E.O.; Lee, Y.R.; Joo, H.K.; Park, M.S.; Kim, C.S.; Choi, S.; Jeong, J.O.; Jeon, B.H. APE1/Ref-1 Inhibits Phosphate-Induced Calcification and Osteoblastic Phenotype Changes in Vascular Smooth Muscle Cells. Int. J. Mol. Sci. 2017, 18, 2053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, J.L.; Wang, X.; Van, A.; Zhao, Y.; Gibbons, G.H. Overexpression of Ref-1 Inhibits Hypoxia and Tumor Necrosis Factor–Induced Endothelial Cell Apoptosis Through Nuclear Factor-kappab-independent and -dependent pathway. Circ. Res. 2001, 88, 1247–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tell, G.; Damante, G.; Caldwell, D.; Kelley, M.R. The intracellular localization of APE1/Ref-1: More than a passive phenomenon? Antioxid. Redox Signal. 2005, 7, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Tell, G.; Crivellato, E.; Pines, A.; Paron, I.; Pucillo, C.; Manzini, G.; Bandiera, A.; Kelley, M.R.; Di Loreto, C.; Damante, G. Mitochondrial localization of APE/Ref-1 in thyroid cells. Mutat. Res. 2001, 485, 143–152. [Google Scholar] [CrossRef]
- Yuk, J.M.; Yang, C.S.; Shin, D.M.; Kim, K.K.; Lee, S.K.; Song, Y.J.; Lee, H.M.; Cho, C.H.; Jeon, B.H.; Jo, E.K. A dual regulatory role of apurinic/apyrimidinic endonuclease 1/redox factor-1 in HMGB1-induced inflammatory responses. Antioxid. Redox Signal. 2009, 11, 575–588. [Google Scholar] [CrossRef]
- Qu, J.; Liu, G.H.; Huang, B.; Chen, C. Nitric oxide controls nuclear export of APE1/Ref-1 through S-nitrosation of cysteines 93 and 310. Nucleic Acids Res. 2007, 35, 2522–2532. [Google Scholar] [CrossRef]
- Baek, H.; Lim, C.S.; Byun, H.S.; Cho, H.S.; Lee, Y.R.; Shin, Y.S.; Kim, H.W.; Jeon, B.H.; Kim, D.W.; Hong, J.; et al. The anti-inflammatory role of extranuclear apurinic/apyrimidinic endonuclease 1/redox effector factor-1 in reactive astrocytes. Mol. Brain 2016, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Hao, J.; Du, H.; Liu, F.; Lu, J.C.; Yang, X.C.; Cui, W. Apurinic/apyrimidinic endonuclease/redox factor 1 (APE1) alleviates myocardial hypoxia-reoxygenation injury by inhibiting oxidative stress and ameliorating mitochondrial dysfunction. Exp. Med. 2019, 17, 2143–2151. [Google Scholar] [CrossRef] [Green Version]
- Angkeow, P.; Deshpande, S.S.; Qi, B.; Liu, Y.X.; Park, Y.C.; Jeon, B.H.; Ozaki, M.; Irani, K. Redox factor-1: An extra-nuclear role in the regulation of endothelial oxidative stress and apoptosis. Cell Death Differ. 2002, 9, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Davidson, S.M.; Duchen, M.R. Endothelial mitochondria: Contributing to vascular function and disease. Circ. Res. 2007, 100, 1128–1141. [Google Scholar] [CrossRef] [Green Version]
- Joo, H.K.; Lee, Y.R.; Park, M.S.; Choi, S.; Park, K.; Lee, S.K.; Kim, C.S.; Park, J.B.; Jeon, B.H. Mitochondrial APE1/Ref-1 suppressed protein kinase C-induced mitochondrial dysfunction in mouse endothelial cells. Mitochondrion 2014, 17, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Barchiesi, A.; Wasilewski, M.; Chacinska, A.; Tell, G.; Vascotto, C. Mitochondrial translocation of APE1 relies on the MIA pathway. Nucleic Acids Res. 2015, 43, 5451–5464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballista-Hernandez, J.; Martinez-Ferrer, M.; Velez, R.; Climent, C.; Sanchez-Vazquez, M.M.; Torres, C.; Rodriguez-Munoz, A.; Ayala-Pena, S.; Torres-Ramos, C.A. Mitochondrial DNA Integrity Is Maintained by APE1 in Carcinogen-Induced Colorectal Cancer. Mol. Cancer Res. 2017, 15, 831–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Lee, Y.R.; Park, M.S.; Joo, H.K.; Cho, E.J.; Kim, H.S.; Kim, C.S.; Park, J.B.; Irani, K.; Jeon, B.H. Histone deacetylases inhibitor trichostatin A modulates the extracellular release of APE1/Ref-1. Biochem. Biophys. Res. Commun. 2013, 435, 403–407. [Google Scholar] [CrossRef]
- Park, M.S.; Lee, Y.R.; Choi, S.; Joo, H.K.; Cho, E.J.; Kim, C.S.; Park, J.B.; Jo, E.K.; Jeon, B.H. Identification of plasma APE1/Ref-1 in lipopolysaccharide-induced endotoxemic rats: Implication of serological biomarker for an endotoxemia. Biochem. Biophys. Res. Commun. 2013, 435, 621–626. [Google Scholar] [CrossRef]
- Ordway, J.M.; Eberhart, D.; Curran, T. Cysteine 64 of Ref-1 is not essential for redox regulation of AP-1 DNA binding. Mol. Cell Biol. 2003, 23, 4257–4266. [Google Scholar] [CrossRef] [Green Version]
- Nath, S.; Roychoudhury, S.; Kling, M.J.; Song, H.; Biswas, P.; Shukla, A.; Band, H.; Joshi, S.; Bhakat, K.K. The extracellular role of DNA damage repair protein APE1 in regulation of IL-6 expression. Cell. Signal. 2017, 39, 18–31. [Google Scholar] [CrossRef]
- Park, M.S.; Choi, S.; Lee, Y.R.; Joo, H.K.; Kang, G.; Kim, C.S.; Kim, S.J.; Lee, S.D.; Jeon, B.H. Secreted APE1/Ref-1 inhibits TNF-alpha-stimulated endothelial inflammation via thiol-disulfide exchange in TNF receptor. Sci. Rep. 2016, 6, 23015. [Google Scholar] [CrossRef] [Green Version]
- Cinel, I.; Opal, S.M. Molecular biology of inflammation and sepsis: A primer. Crit. Care Med. 2009, 37, 291–304. [Google Scholar] [CrossRef]
- Lee, K.H.; Lee, S.K.; Kim, H.S.; Cho, E.J.; Joo, H.K.; Lee, E.J.; Lee, J.Y.; Park, M.S.; Chang, S.J.; Cho, C.H.; et al. Overexpression of Ref-1 Inhibits Lead-induced Endothelial Cell Death via the Upregulation of Catalase. Korean J. Physiol. Pharm. 2009, 13, 431–436. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.B.; Kim, C.S.; Kim, Y.R.; Naqvi, A.; Yamamori, T.; Kumar, A.; Irani, K. Redox factor-1 anctivates endothelial SIRTUIN1 though reduction of conserved cystein sulfhydryls in its deacetylase domain. PLoS ONE 2013, 8, e65415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.K.; Chung, J.I.; Park, M.S.; Joo, H.K.; Lee, E.J.; Cho, E.J.; Park, J.B.; Ryoo, S.; Irani, K.; Jeon, B.H. Apurinic/apyrimidinic endonuclease 1 inhibits protein kinase C-mediated p66shc phosphorylation and vasoconstriction. Cardiovasc. Res. 2011, 91, 502–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, D.G.; Kim, C.S.; Lee, S.K.; Kim, H.S.; Cho, E.J.; Park, M.S.; Lee, S.D.; Park, J.B.; Jeon, B.H. Redox Factor-1 Inhibits Cyclooxygenase-2 Expression via Inhibiting of p38 MAPK in the A549 Cells. Korean J. Physiol. Pharm. 2010, 14, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.R.; Kim, K.M.; Jeon, B.H.; Choi, S. Extracellularly secreted APE1/Ref-1 triggers apoptosis in triple-negative breast cancer cells via RAGE binding, which is mediated through acetylation. Oncotarget 2015, 6, 23383–23398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.A.; Lim, B.K.; Seo, H.J.; Kim, S.K.; Ahn, K.T.; Jeon, B.H.; Jeong, J.O. Elevation of Serum APE1/Ref-1 in Experimental Murine Myocarditis. Int. J. Mol. Sci. 2017, 18, 2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.R.; Lim, J.S.; Shin, J.H.; Choi, S.; Joo, H.K.; Jeon, B.H. Altered Secretory Activity of APE1/Ref-1 D148E Variants Identified in Human Patients With Bladder Cancer. Int. Neurourol. J. 2016, 20, S30–S37. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.R.; Joo, H.K.; Lee, E.O.; Cho, H.S.; Choi, S.; Kim, C.S.; Jeon, B.H. ATP Binding Cassette Transporter A1 is Involved in Extracellular Secretion of Acetylated APE1/Ref-1. Int. J. Mol. Sci. 2019, 20, 3178. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.R.; Park, M.S.; Joo, H.K.; Kim, K.M.; Kim, J.; Jeon, B.H.; Choi, S. Therapeutic positioning of secretory acetylated APE1/Ref-1 requirement for suppression of tumor growth in triple-negative breast cancer in vivo. Sci. Rep. 2018, 8, 8701. [Google Scholar] [CrossRef] [Green Version]
- Bhakat, K.K.; Izumi, T.; Yang, S.H.; Hazra, T.K.; Mitra, S. Role of acetylated human AP-endonuclease (APE1/Ref-1) in regulation of the parathyroid hormone gene. EMBO J. 2003, 22, 6299–6309. [Google Scholar] [CrossRef] [Green Version]
- Scaffidi, P.; Misteli, T.; Bianchi, M.E. Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 2002, 418, 191–195. [Google Scholar] [CrossRef]
- Jin, S.A.; Seo, H.J.; Kim, S.K.; Lee, Y.R.; Choi, S.; Ahn, K.T.; Kim, J.H.; Park, J.H.; Lee, J.H.; Choi, S.W.; et al. Elevation of the Serum Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 in Coronary Artery Disease. Korean Circ. J. 2015, 45, 364–371. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.T., Jr. Myocarditis. N. Engl. J. Med. 2009, 360, 1526–1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magnani, J.W.; Dec, G.W. Myocarditis: Current trends in diagnosis and treatment. Circulation 2006, 113, 876–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDowell, S.A.C.; Quail, D.F. Immunological Regulation of Vascular Inflammation During Cancer Metastasis. Front. Immunol. 2019, 10, 1984. [Google Scholar] [CrossRef]
- Yuan, C.L.; He, F.; Ye, J.Z.; Wu, H.N.; Zhang, J.Y.; Liu, Z.H.; Li, Y.-Q.; Luo, X.-L.; Lin, Y.; Liang, R. APE1 overexpression is associated with poor survival in patients with solid tumors: A meta-analysis. Oncotarget 2017, 8, 59720–59728. [Google Scholar] [CrossRef] [Green Version]
- Fishel, M.L.; Kelley, M.R. The DNA base excision repair protein Ape1/Ref-1 as a therapeutic and chemopreventive target. Mol. Asp. Med. 2007, 28, 375–395. [Google Scholar] [CrossRef]
- Choi, S.; Shin, J.H.; Lee, Y.R.; Joo, H.K.; Song, K.H.; Na, Y.G.; Chang, S.J.; Lim, J.S.; Jeon, B.H. Urinary APE1/Ref-1: A Potential Bladder Cancer Biomarker. Dis. Markers 2016, 2016, 7276502. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.H.; Choi, S.; Lee, Y.R.; Park, M.S.; Na, Y.G.; Irani, K.; Lee, S.D.; Park, J.B.; Kim, J.M.; Lim, J.S.; et al. APE1/Ref-1 as a Serological Biomarker for the Detection of Bladder Cancer. Cancer Res. Treat. 2015, 47, 823–833. [Google Scholar] [CrossRef] [Green Version]
- Pascut, D.; Sukowati, C.H.C.; Antoniali, G.; Mangiapane, G.; Burra, S.; Mascaretti, L.G.; Buonocore, M.R.; Crocè, L.S.; Tiribelli, C.; Tell, G. Serum AP-endonuclease 1 (sAPE1) as novel biomarker for hepatocellular carcinoma. Oncotarget 2019, 10, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.M.; Yeo, M.K.; Lim, J.S.; Song, I.S.; Chun, K.; Kim, K.H. APEX1 Expression as a Potential Diagnostic Biomarker of Clear Cell Renal Cell Carcinoma and Hepatobiliary Carcinomas. J. Clin. Med. 2019, 8, 1151. [Google Scholar] [CrossRef] [Green Version]
- Tummanatsakun, D.; Proungvitaya, T.; Roytrakul, S.; Limpaiboon, T.; Wongkham, S.; Wongkham, C.; Silsirivanit, A.; Somintara, O.; Sangkhamanon, S.; Proungvitaya, S. Serum Apurinic/Apyrimidinic Endodeoxyribonuclease 1 (APEX1) Level as a Potential Biomarker of Cholangiocarcinoma. Biomolecules 2019, 9, 413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; He, L.; Dai, N.; Guan, W.; Shan, J.; Yang, X.; Zhong, Z.; Qing, Y.; Jin, F.; Chen, C.; et al. Serum APE1 as a predictive marker for platinum-based chemotherapy of non-small cell lung cancer patients. Oncotarget 2016, 7, 77482–77494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Li, Y.B.; Li, Y.; Lin, B.C.; Shen, X.M.; Cui, R.L.; Gu, Y.J.; Gao, M.; Li, Y.G.; Zhang, S. Prediction of Lymph Node Metastases in Gastric Cancer by Serum APE1 Expression. J. Cancer 2017, 8, 1492–1497. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Li, Y.; Kong, J.; Li, C. Apurinic/Apyrimidinic Endonuclease 1/Redox Factor-1 Could Serve as a Potential Serological Biomarker for the Diagnosis and Prognosis of Oral Squamous Cell Carcinoma. J. Oral Maxillofac. Surg. 2019, 77, 859–866. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| APE1/Ref-1 | Tissue/Cells | Functions | Reference |
|---|---|---|---|
| Intracellular | Endothelial cells | Inhibits endothelial dysfunction Inhibits cellular ROS and increases NO production Inhibits NF-kB and apoptosis Inhibits VCAM-1 expression | [18,19,23,29,50,51] |
| Endothelial mitochondria | Inhibit mitochondrial dysfunctions Inhibits mitochondrial ROS Inhibits p66shc activation Mitochondrial DNA repair | [41,52] [42] | |
| A549 cells | Inhibits COX-2 expression Inhibits p38 MAPK | [53] | |
| Vascular smooth muscle cells | Inhibits Pi-induced calcification Inhibits osteoblastic phenotype changes | [31] | |
| Cytoplasmic | Endothelial cells | Inhibits rac1 or NADPH oxidase | [29,39] |
| Glial cells | Inhibits neuroinflammatory response | [37] | |
| THP-1 cells | Inhibits HMGB1-mediated ROS and cytokines | [35] | |
| Extracellular | HEK293 cells | Trichostatin A induced APE1/Ref-1 secretion | [44] |
| MDA-MD-231 cells | Ac-APE1/Ref-1 induces apoptosis | [54] | |
| Endothelial cells | Inhibits VCAM-1 expression | [48] | |
| Inhibits COX-2 expression | [22] |
| Diseases | Clinical Significance | Patients (n) | Control (n) | Sensitivity (%) | Specificity (%) | AUC or 95% CI | Reference |
|---|---|---|---|---|---|---|---|
| Liposaccharide-induced endotoxemia (Preclinical study) | APE1/Ref-1 is elevated in plasma of lipopolysaccharide (LPS)-treated mice and reached a maximum at 12 h after injection of LPS. | - | - | NA | NA | [45] | |
| Viral myocarditis (Preclinical study) | Serum APE1/Ref-1 is increased in coxsackievirus-induced myocarditis and is well-correlated with the degree of myocardial inflammation. Serum APE1/Ref-1 is useful for myocardial injury in viral myocarditis without endomyocardial biopsy | - | - | NA | NA | [55] | |
| Coronary arterial diseases | Serum APE1/Ref-1 level was higher in coronary arterial diseases, which higher in myocardial infarction than angina | 303 | 57 | 36 | 95 | 0.66 | [61] |
| Bladder cancer | Urinary APE1/Ref-1 is increased in bladder cancer and it correlated with tumor grade and stage | 169 | 108 | 82 | 80 | 0.83 | [67] |
| Oral squamous cell carcinoma | Serum APE1/Ref-1 is a novel potential diagnostic biomarker of oral cancer and can reflect the treatment responses | 58 | 40 | 67 | 87 | 0.80 | [74] |
| Cholangiocarcinoma | Serum APE1/Ref-1 level is a potential diagnostic marker of cholangiocarcinoma and cytoplasmic expression in cancer cells predicts relapses | 46 | 39 | 73.9 | 97.4 | 0.709–0.886 | [71] |
| Hepatocellular carcinoma | Serum APE1/Ref-1 may be considered as a promising diagnostic biomarker for hepatocellular carcinoma | 99 | 100 | 98 | 83 | 0.98 | [69] |
| Renal cell carcinoma | Serum APE1/Ref-1 level may be a diagnostic markers of renal cell carcinoma | 40 | 39 | 82.5 | 97.4 | 0.862–0.981 | [70] |
| Non-small cell lung cancer | Serum APE1/Ref-1 is a biomarker for predicting prognosis and therapeutic efficacy in nonsmall cell lung cancer and post-treatment high serum APE1/Ref-1 level was associated with poor survival. | 200 | 200 | 55.6 | 70.8 | 0.653 | [72] |
| Gastric cancer (lymph node positive and negative | Serum APE1/Ref-1 is a valuable marker for prediction of lymph node metastasis in patients with gastric cancer | 52 | 35 | 49 | 85.7 | 0.666 | [73] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.R.; Joo, H.K.; Jeon, B.H. The Biological Role of Apurinic/Apyrimidinic Endonuclease1/Redox Factor-1 as a Therapeutic Target for Vascular Inflammation and as a Serologic Biomarker. Biomedicines 2020, 8, 57. https://doi.org/10.3390/biomedicines8030057
Lee YR, Joo HK, Jeon BH. The Biological Role of Apurinic/Apyrimidinic Endonuclease1/Redox Factor-1 as a Therapeutic Target for Vascular Inflammation and as a Serologic Biomarker. Biomedicines. 2020; 8(3):57. https://doi.org/10.3390/biomedicines8030057
Chicago/Turabian StyleLee, Yu Ran, Hee Kyoung Joo, and Byeong Hwa Jeon. 2020. "The Biological Role of Apurinic/Apyrimidinic Endonuclease1/Redox Factor-1 as a Therapeutic Target for Vascular Inflammation and as a Serologic Biomarker" Biomedicines 8, no. 3: 57. https://doi.org/10.3390/biomedicines8030057
APA StyleLee, Y. R., Joo, H. K., & Jeon, B. H. (2020). The Biological Role of Apurinic/Apyrimidinic Endonuclease1/Redox Factor-1 as a Therapeutic Target for Vascular Inflammation and as a Serologic Biomarker. Biomedicines, 8(3), 57. https://doi.org/10.3390/biomedicines8030057