Zebra-Fishing for Regenerative Awakening in Mammals


Abstract
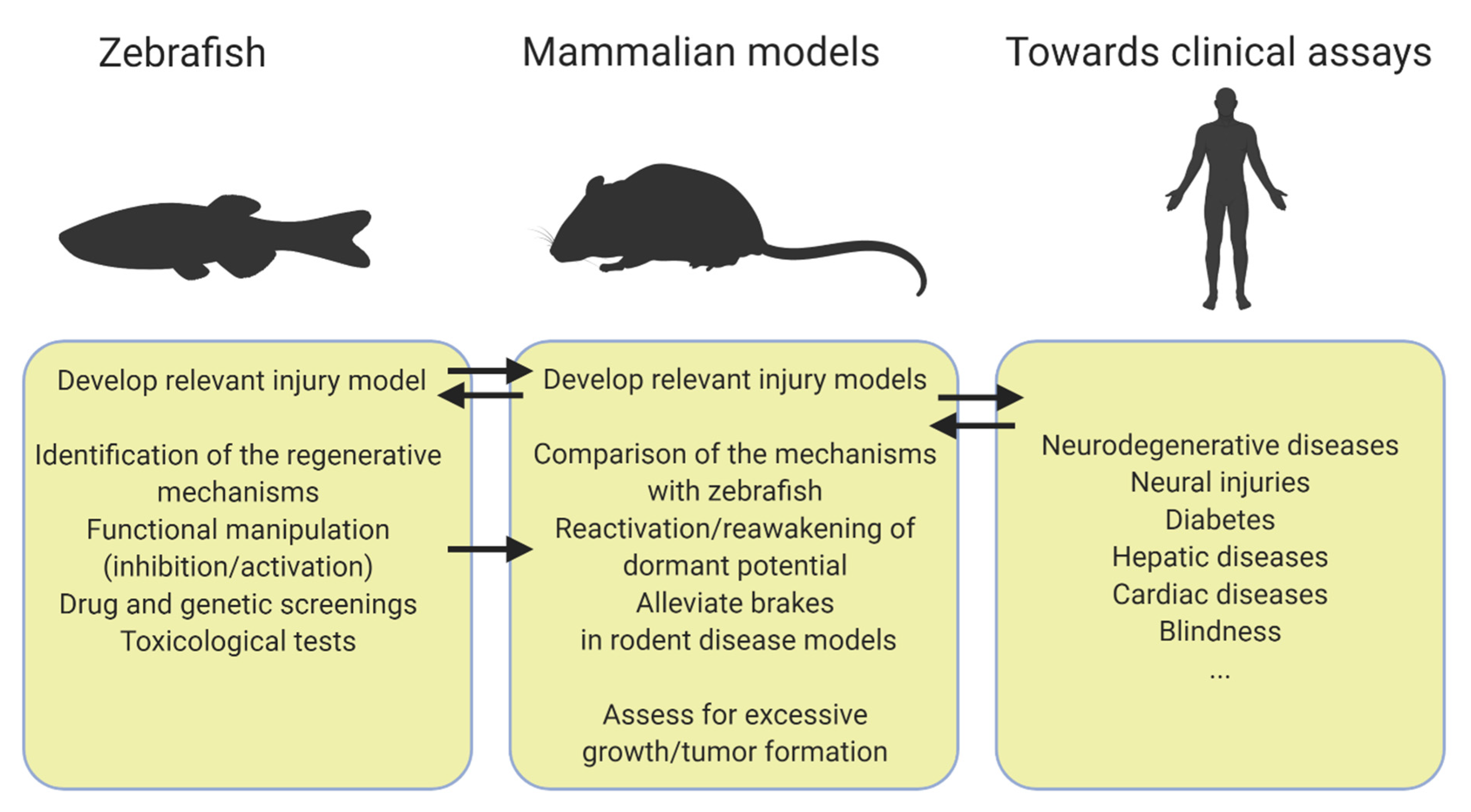
:1. Introduction
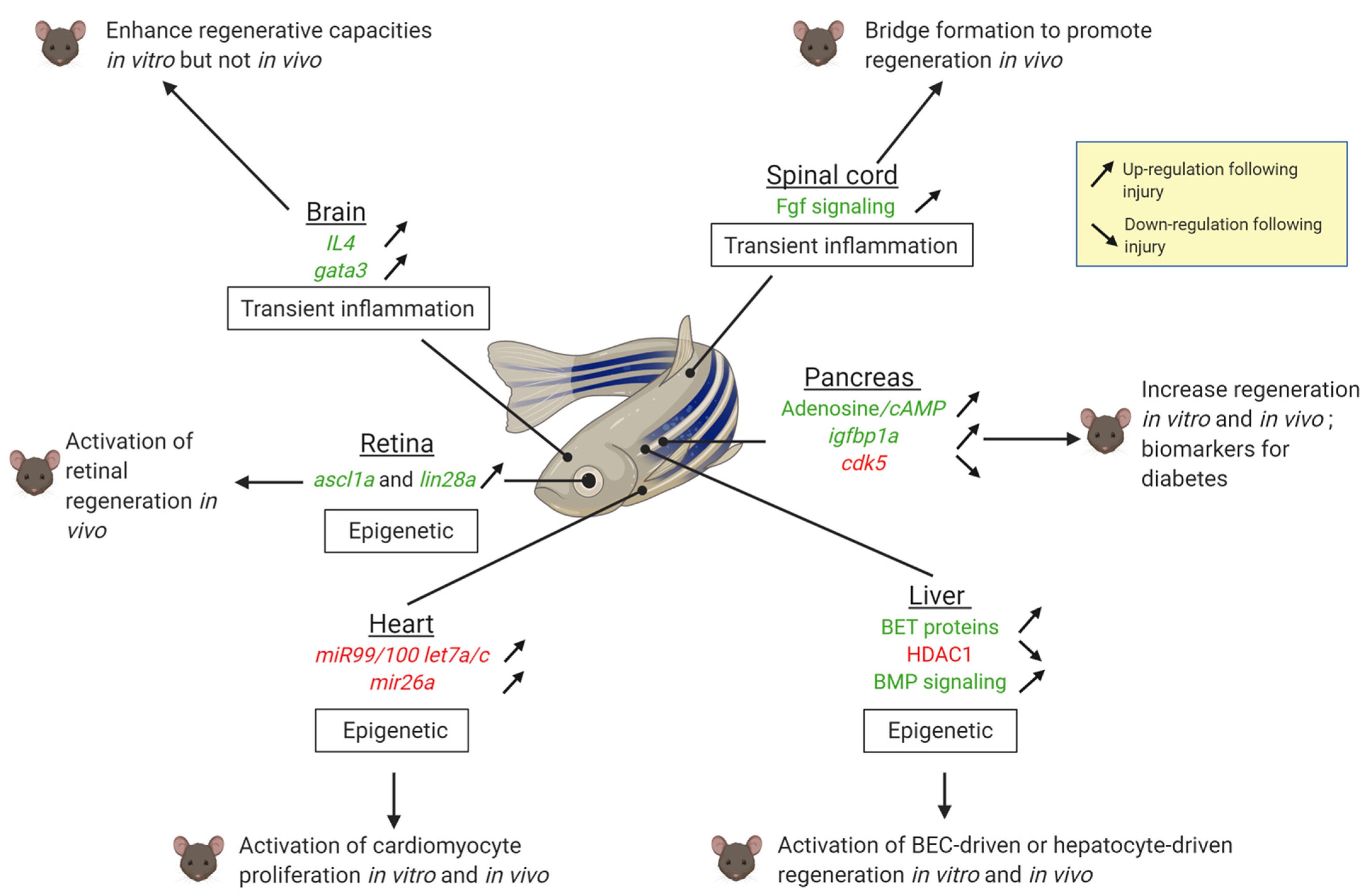
2. Awakening the Regenerative Capacity in Different Organs
2.1. The Heart

{kind=link}
{kind=link}
| Organ | Model Organism | Injury Model | Type of Injury | Mechanism of Regeneration | Characteristics | References |
|---|---|---|---|---|---|---|
| Heart | Zebrafish | Ventricular resection | Surgical | Proliferation from pre-existing myocytes | - | [11] |
| Cryoinjury | Surgical | Proliferation from pre-existing myocytes | Clinically relevant to mammalian infarcts with massive cell death | [13] | ||
| Mouse | Ventricular resection | Surgical | Proliferation from pre-existing myocytes | Fully regenerates a functional myocardium in 1–2 months without scarring | [11,12] | |
| Myocardial infarction | Surgical | Proliferation from pre-existing myocytes | Left anterior descending coronary artery occluded with a nylon suture | [25] | ||
| Liver | Zebrafish | Partial hepatectomy | Surgical | Hepatocyte-driven | Clinically relevant | [26] |
| APAP overdose | Chemical | BEC-driven regeneration | Paracetamol overdose | [26] | ||
| Nitroreductase (NTR)-mediated ablation | Genetic/Chemical | Hepatocyte-driven BEC-driven regeneration | Tg(fabp10a:CFP-NTR) | [27,28,29] | ||
| Mouse | CDE diet | Chemical | BEC-driven regeneration | Ethionine, a toxic analog of methionine, in association with choline deficiency, leads to hepatocyte death and liver inflammation | [30] | |
| Ctnnb1 hepatocyte KO | Genetic | BEC-driven regeneration | Represses hepatocyte proliferation—in combination with an injury model | [29] | ||
| Mdm2 deletion (hepatocyte-specific) | Genetic | BEC-driven regeneration | AhCreMdm2flox/floxInducible, repress hepatocyte proliferation | [30] | ||
| Beta cell/Pancreas | Zebrafish | NTR-mediated ablation | Genetic/Chemical | Beta cell proliferation;alpha cell transdifferentiation; Neogenesis from ductal progenitors | Tg(ins:CFP-NTR) In cells expressing NTR, reduces non-toxic pro-drug into cytotoxic products causing targeted cell apoptosis | [31,32,33,34] |
| Mouse | Pancreatic Duct Ligation (PDL) | Surgical | Neogenesis from ductal progenitors | Induces acinar cell death and acute inflammation without destruction of beta cells | [35,36,37] | |
| Streptozotocin (STZ) | Chemical | Beta cell proliferation; Neogenesis from ductal progenitors | Toxic glucose analogue that enters into beta cells via the GLUT2 transporter causing their death | [31,38] | ||
| Diphtheria Toxin Analogue (DTA) | Genetic/Chemical | Alpha cell transdifferentiation (adult only) Delta cell transdifferentiation (neonatal only) | Tg(RIP:DTR) The toxin enters in cells expressing the DTR and inhibits protein synthesis, leading to cell apoptosis. Here targeted in beta cells with the Rat Insulin Promoter (RIP). | [39,40] | ||
| Spinal Cord | Zebrafish | Spinal cord transection | Surgical | Glial bridge | Complete cutting of the vertebral column | [41] |
| Mouse | Laminectomy and spinal cord hemisection | Surgical | - | Hemisection leading to complete paralysis of the ipsilateral limb | [42] | |
| Brain | Zebrafish | Stab-lesion assay | Surgical | Regeneration from radial cells | Injury in the telencephalon parenchyma | [43] |
| B42 mediated injury | Surgical/Chemical | Regeneration from radial cells | Alzheimer’s-disease-like | [44] | ||
| Mouse | AD-like model | Genetic | No regeneration | APP/PS1dE9 transgenic | [45] | |
| Retina | Zebrafish | Needle poke | Surgical | From Muller cells | - | [46] |
| Optic nerve lesion | Surgical | From Muller cells | - | [47] | ||
| Mouse | NMDA | Chemical | From Muller cells | - | [46,48] | |
| Excessive light | Surgical | From Muller cells | - | [46] | ||
| AD-like model | Genetic | No regeneration | APP/PS1dE9 transgenic | [45] |
2.2. The Liver
2.3. The Pancreas
2.4. The Central Nervous System (CNS)
2.4.1. The Spinal Cord
2.4.2. The Brain
2.4.3. The Retina
3. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
List of Acronyms
| AD | Alzheimer’s disease |
| APAP | N-acetyl-p-aminophenol |
| ASCL1 | Achaete Scute-Like family bHLH transcription factor 1 |
| BECs | Biliary Epithelial Cells |
| BET | Bromodomain and Extra-Terminal proteins |
| BMP | Bone Morphogenetic Protein |
| CDE | Choline-Deficient Ethionine-supplemented |
| CNS | Central Nervous System |
| DTA | Diphtheria Toxin Subunit A |
| DTR | Diphtheria Toxin Receptor |
| FGF | Fibroblast Growth Factor |
| HDAC1 | Histone Deacetylase 1 |
| Igf | insulin-like growth factor |
| igfbp1 | Igf binding-protein 1 |
| IL4 | interleukin-4 |
| LEN | lepb-linked enhancer |
| LPCs | Liver Progenitors Cells |
| MCs | Müller cells |
| miRNAs | micro RNAs |
| NECA | 50-N-ethylcarboxamidoadenosine |
| NMDA | N-methyl D-Aspartate |
| NTR | Nitroreductase |
| PDL | Pancreatic Duct Ligation |
| PIAA | (E)-3-(3-phenylbenzo[c]isoxazol-5-yl) acrylic acid |
| STZ | StrepToZotocin |
| T1D | Type 1 Diabetes |
| T2D | Type 2 Diabetes |
References
- Elliott, S.A.; Sánchez Alvarado, A. Planarians and the History of Animal Regeneration: Paradigm Shifts and Key Concepts in Biology. In Planarian Regeneration. Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; Volume 1774, ISBN 9781493978021. [Google Scholar]
- Maden, M. The evolution of regeneration—Where does that leave mammals? Int. J. Dev. Biol. 2018, 62, 369–372. [Google Scholar] [PubMed] [Green Version]
- Gemberling, M.; Bailey, T.J.; Hyde, D.R.; Poss, K.D. The zebrafish as a model for complex tissue regeneration. Trends Genet. 2013, 29, 611–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinth, P.; Mahesh, G.; Panwar, Y. Mapping of zebrafish research: A global outlook. Zebrafish 2013, 10, 510–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehta, A.S.; Singh, A. Insights into regeneration tool box: An animal model approach. Dev. Biol. 2019, 453, 111–129. [Google Scholar] [CrossRef]
- Marques, I.J.; Lupi, E.; Mercader, N. Model systems for regeneration: Zebrafish. Development 2019, 146, dev167692. [Google Scholar] [CrossRef] [Green Version]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Muffato, M.; Collins, J.E.; Humphray, S.; Mclaren, K.; Matthews, L.; Mclaren, S.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2014, 496, 498–503. [Google Scholar] [CrossRef] [Green Version]
- Roth, G.A.; Johnson, C.; Abajobir, A.; Abd-Allah, F.; Abera, S.F.; Abyu, G.; Ahmed, M.; Aksut, B.; Alam, T.; Alam, K.; et al. Global, Regional, and National Burden of Cardiovascular Diseases for 10 Causes, 1990 to 2015. J. Am. Coll. Cardiol. 2017, 70, 1–25. [Google Scholar] [CrossRef]
- Bergmann, O.; Bhardwaj, R.D.; Bernard, S.; Zdunek, S.; Barnabé-Heider, F.; Walsh, S.; Zupicich, J.; Alkass, K.; Buchholz, B.A.; Druid, H.; et al. Evidence for cardiomyocyte renewal in humans. Science 2009, 324, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, H.; Olson, E.N.; Bassel-Duby, R. Therapeutic approaches for cardiac regeneration and repair. Nat. Rev. Cardiol. 2018, 15, 585–600. [Google Scholar] [CrossRef]
- Poss, K.D.; Wilson, L.G.; Keating, M.T. Heart Regeneration in Zebrafish. Science 2002, 298, 2188–2190. [Google Scholar] [CrossRef]
- Raya, A.; Koth, C.M.; Büscher, D.; Kawakami, Y.; Itoh, T.; Raya, R.M.; Sternik, G.; Tsai, H.-J.; Rodríguez-Esteban, C.; Izpisúa-Belmonte, J.C. Activation of Notch signaling pathway precedes heart regeneration in zebrafish. Proc. Natl. Acad. Sci. USA 2003, 100 (Suppl. 1), 11889–11895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chablais, F.; Veit, J.; Rainer, G.; Jaźwińska, A. The zebrafish heart regenerates after cryoinjury-induced myocardial infarction. BMC Dev. Biol. 2011, 11, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Rosa, J.M.; Martín, V.; Peralta, M.; Torres, M.; Mercader, N. Extensive scar formation and regression during heart regeneration after cryoinjury in zebrafish. Development 2011, 138, 1663–1674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, N.; Harrison, M.R.; Krainock, M.; Kim, R.; Lien, C.-L. Recent advancements in understanding endogenous heart regeneration-insights from adult zebrafish and neonatal mice. Semin. Cell Dev. Biol. 2016, 58, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Hill, J.A.; Richardson, J.A.; Olson, E.N.; Sadek, H.A. Transient regenerative potential of the neonatal mouse heart. Science 2011, 331, 1078–1080. [Google Scholar] [CrossRef] [Green Version]
- Jopling, C.; Sleep, E.; Raya, M.; Martí, M.; Raya, A.; Izpisúa Belmonte, J.C. Zebrafish heart regeneration occurs by cardiomyocyte dedifferentiation and proliferation. Nature 2010, 464, 606–609. [Google Scholar] [CrossRef]
- González-Rosa, J.M.; Sharpe, M.; Field, D.; Soonpaa, M.H.; Field, L.J.; Burns, C.E.; Burns, C.G. Myocardial Polyploidization Creates a Barrier to Heart Regeneration in Zebrafish. Dev. Cell 2018, 44, 433–446.e7. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wang, X.; Capasso, J.M.; Gerdes, A.M. Rapid Transition of Cardiac Myocytes from Hyperplasia to Hypertrophy During Postnatal Development. J. Mol. Cell. Cardiol. 1996, 28, 1737–1746. [Google Scholar] [CrossRef]
- Soonpaa, M.H.; Kim, K.K.; Pajak, L.; Franklin, M.; Field, L.J. Cardiomyocyte DNA synthesis and binucleation during murine development. Am. J. Physiol. 1996, 271, H2183–H2189. [Google Scholar] [CrossRef]
- Mollova, M.; Bersell, K.; Walsh, S.; Savla, J.; Das, L.T.; Park, S.-Y.; Silberstein, L.E.; Dos Remedios, C.G.; Graham, D.; Colan, S.; et al. Cardiomyocyte proliferation contributes to heart growth in young humans. Proc. Natl. Acad. Sci. USA 2013, 110, 1446–1451. [Google Scholar] [CrossRef] [Green Version]
- Vivien, C.J.; Hudson, J.E.; Porrello, E.R. Evolution, comparative biology and ontogeny of vertebrate heart regeneration. NPJ Regen. Med. 2016, 1, 16012. [Google Scholar] [CrossRef] [Green Version]
- Green, R.A.; Paluch, E.; Oegema, K. Cytokinesis in Animal Cells. Annu. Rev. Cell Dev. Biol. 2012, 28, 29–58. [Google Scholar] [CrossRef] [Green Version]
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Johnson, B.A.; Grinsfelder, D.; Canseco, D.; Mammen, P.P.; Rothermel, B.A.; Olson, E.N.; Sadek, H.A. Regulation of neonatal and adult mammalian heart regeneration by the miR-15 family. Proc. Natl. Acad. Sci. USA 2013, 110, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, A.; Montserrat, N.; Zacchigna, S.; Nivet, E.; Hishida, T.; Krause, M.N.; Kurian, L.; Ocampo, A.; Vazquez-Ferrer, E.; Rodriguez-Esteban, C.; et al. In vivo activation of a conserved microRNA program induces robust mammalian heart regeneration. Cell Stem Cell 2014, 15, 589–604. [Google Scholar] [CrossRef] [Green Version]
- Russell, J.O.; Ko, S.; Saggi, H.S.; Singh, S.; Poddar, M.; Shin, D.; Monga, S.P. Bromodomain and Extraterminal (BET) Proteins Regulate Hepatocyte Proliferation in Hepatocyte-Driven Liver Regeneration. Am. J. Pathol. 2018, 188, 1389–1405. [Google Scholar] [CrossRef] [Green Version]
- Choi, T.; Khaliq, M.; Tsurusaki, S.; Ninov, N.; Stainier, D.; Tanaka, M.; Shin, D. Bmp Signaling Governs Biliary-Driven Liver Regeneration in Zebrafish via Tbx2b and Id2a. Hepatology 2017, 66, 1616–1630. [Google Scholar] [CrossRef]
- Ko, S.; Choi, T.; Russell, J.O.; So, J.; Shin, D. Bromodomain and extraterminal (BET) proteins regulate biliary-driven liver regeneration. J. Hepatol. 2016, 64, 316–325. [Google Scholar] [CrossRef] [Green Version]
- Ko, S.O.; Russell, J.; Tian, J.; Gao, C.; Kobayashi, M.; Feng, R.; Yuan, X.; Shao, C.; Ding, H.; Poddar, M.; et al. Hdac1 Regulates Differentiation of Bipotent Liver Progenitor Cells During Regeneration via Sox9b and Cdk8. Gastroenterology 2019, 156, 187–202.e4. [Google Scholar] [CrossRef]
- Lu, W.Y.; Bird, T.G.; Boulter, L.; Tsuchiya, A.; Cole, A.M.; Hay, T.; Guest, R.V.; Wojtacha, D.; Man, T.Y.; Mackinnon, A.; et al. Hepatic progenitor cells of biliary origin with liver repopulation capacity. Nat. Cell Biol. 2015, 17, 973–983. [Google Scholar] [CrossRef]
- Andersson, O.; Adams, B.A.; Yoo, D.; Ellis, G.C.; Gut, P.; Anderson, R.M.; German, M.S.; Stainier, D.Y.R. Adenosine Signaling Promotes Regeneration of Pancreatic b Cells In Vivo. Cell Metab. 2012, 885–894. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Jia, Y.F.; Tapadar, S.; Weaver, J.D.; Raji, I.O.; Pithadia, D.J.; Javeed, N.; García, A.J.; Choi, D.S.; Matveyenko, A.V.; et al. Inhibition of TBK1/IKKε Promotes Regeneration of Pancreatic β-cells. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Liu, K.; Schulz, N.; Karampelias, C.; Charbord, J.; Hilding, A.; Rautio, L.; Bertolino, P.; Östenson, C.; Brismar, K.; et al. IGFBP 1 increases β-cell regeneration by promoting α- to β-cell transdifferentiation. EMBO J. 2016, 35, 2026–2044. [Google Scholar] [CrossRef]
- Ye, L.; Robertson, M.A.; Hesselson, D.; Stainier, D.Y.R.; Anderson, R.M. Glucagon is essential for alpha cell transdifferentiation and beta cell neogenesis. Development 2015, 142, 1407–1417. [Google Scholar] [CrossRef] [Green Version]
- Kopp, J.L.; Dubois, C.L.; Schaffer, A.E.; Hao, E.; Shih, H.P.; Seymour, P.A.; Ma, J.; Sander, M. Sox9+ ductal cells are multipotent progenitors throughout development but do not produce new endocrine cells in the normal or injured adult pancreas. Development 2011, 138, 653–665. [Google Scholar] [CrossRef] [Green Version]
- Solar, M.; Cardalda, C.; Houbracken, I.; Martín, M.; Maestro, M.A.; De Medts, N.; Xu, X.; Grau, V.; Heimberg, H.; Bouwens, L.; et al. Pancreatic Exocrine Duct Cells Give Rise to Insulin-Producing β Cells during Embryogenesis but Not after Birth. Dev. Cell 2009, 17, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; D’Hoker, J.; Stangé, G.; Bonné, S.; De Leu, N.; Xiao, X.; Van De Casteele, M.; Mellitzer, G.; Ling, Z.; Pipeleers, D.; et al. β Cells Can Be Generated from Endogenous Progenitors in Injured Adult Mouse Pancreas. Cell 2008, 132, 197–207. [Google Scholar] [CrossRef] [Green Version]
- Al-Hasani, K.; Pfeifer, A.; Courtney, M.; Ben-Othman, N.; Gjernes, E.; Vieira, A.; Druelle, N.; Avolio, F.; Ravassard, P.; Leuckx, G.; et al. Adult duct-lining cells can reprogram into β-like cells able to counter repeated cycles of toxin-induced diabetes. Dev. Cell 2013, 26, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Chera, S. Diabetes Recovery By Age-Dependent Conversion of Pancreatic δ-Cells. Nature 2014, 514, 503–507. [Google Scholar] [CrossRef]
- Thorel, F.; Nepote, V.; Avril, I.; Kohno, K.; Desgraz, R.; Chera, S.; Herrera, P.L.; Népote, V.; Kohno, K.; Desgraz, R. Conversion of Adult Pancreatic alpha-cells to Beta-cells after Extreme Beta Cell Loss. Nature 2010, 464, 1149–1154. [Google Scholar] [CrossRef] [Green Version]
- Becker, T.; Wullimann, M.F.; Becker, C.G.; Bernhardt, R.R.; Schachner, M. Axonal regrowth after spinal cord transection in adult zebrafish. J. Comp. Neurol. 1997, 377, 577–595. [Google Scholar] [CrossRef]
- Goldshmit, Y.; Frisca, F.; Pinto, A.R.; Pébay, A.; Tang, J.-K.K.Y.; Siegel, A.L.; Kaslin, J.; Currie, P.D. Fgf2 improves functional recovery-decreasing gliosis and increasing radial glia and neural progenitor cells after spinal cord injury. Brain Behav. 2014, 4, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Kizil, C.; Kyritsis, N.; Dudczig, S.; Kroehne, V.; Freudenreich, D.; Kaslin, J.; Brand, M. Regenerative Neurogenesis from Neural Progenitor Cells Requires Injury-Induced Expression of Gata3. Dev. Cell 2012, 23, 1230–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Froc, C.; Reinhardt, S.; Kurth, T.; Dahl, A.; Zhang, Y.; et al. IL4/STAT6 Signaling Activates Neural Stem Cell Proliferation and Neurogenesis upon Amyloid-β42 Aggregation in Adult Zebrafish Brain. Cell Rep. 2016, 17, 941–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mashkaryan, V.; Siddiqui, T.; Popova, S.; Cosacak, M.I.; Bhattarai, P.; Brandt, K.; Govindarajan, N.; Petzold, A.; Reinhardt, S.; Dahl, A.; et al. Type 1 Interleukin-4 Signaling Obliterates Mouse Astroglia in vivo but Not in vitro. Front. Cell Dev. Biol. 2020, 8, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsaeidi, F.; Macpherson, P.; Mills, E.A.; Jui, J.; Flannery, J.G.; Goldman, D. Notch suppression collaborates with Ascl1 and Lin28 to unleash a regenerative response in fish retina, but not in mice. J. Neurosci. 2018, 38, 2246–2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fausett, B.V.; Goldman, D. A role for alpha1 tubulin-expressing Müller glia in regeneration of the injured zebrafish retina. J. Neurosci. 2006, 26, 6303–6313. [Google Scholar] [CrossRef] [Green Version]
- Jorstad, N.L.; Wilken, M.S.; Grimes, W.N.; Wohl, S.G.; Vandenbosch, L.S.; Yoshimatsu, T.; Wong, R.O.; Rieke, F.; Reh, T.A. Stimulation of functional neuronal regeneration from Müller glia in adult mice. Nature 2017, 548, 103–107. [Google Scholar] [CrossRef]
- Hodgkinson, C.P.; Dzau, V.J. Conserved MicroRNA Program as Key to Mammalian Cardiac Regeneration. Circ. Res. 2015, 116, 1109–1111. [Google Scholar] [CrossRef] [Green Version]
- Crippa, S.; Nemir, M.; Ounzain, S.; Ibberson, M.; Berthonneche, C.; Sarre, A.; Boisset, G.; Maison, D.; Harshman, K.; Xenarios, I.; et al. Comparative transcriptome profiling of the injured zebrafish and mouse hearts identifies miRNA-dependent repair pathways. Cardiovasc. Res. 2016, 110, 73–84. [Google Scholar] [CrossRef]
- Kang, J.; Hu, J.; Karra, R.; Dickson, A.L.; Tornini, V.A.; Nachtrab, G.; Gemberling, M.; Goldman, J.A.; Black, B.L.; Poss, K.D. Modulation of tissue repair by regeneration enhancer elements. Nature 2016, 532, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Bersell, K.; Arab, S.; Haring, B.; Kühn, B. Neuregulin1/ErbB4 signaling induces cardiomyocyte proliferation and repair of heart injury. Cell 2009, 138, 257–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Begeman, I.J.; Kang, J. Transcriptional Programs and Regeneration Enhancers Underlying Heart Regeneration. J. Cardiovasc. Dev. Dis. 2018, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J.C.; Ruck, P.; Adam, A.; Wang, T.X.; Kaiserling, E. Small epithelial cells in human liver cirrhosis exhibit features of hepatic stem-like cells: Immunohistochemical, electron microscopic and immunoelectron microscopic findings. Histopathology 2003, 42, 141–149. [Google Scholar] [CrossRef] [PubMed]
- So, J.; Kim, A.; Lee, S.H.; Shin, D. Liver progenitor cell-driven liver regeneration. Exp. Mol. Med. 2020, 52, 1230–1238. [Google Scholar] [CrossRef]
- Ko, S.; Russell, J.O.; Molina, L.M.; Monga, S.P. Liver Progenitors and Adult Cell Plasticity in Hepatic Injury and Repair: Knowns and Unknowns. Annu. Rev. Pathol. Mech. Dis. 2020, 15, 23–50. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Zhou, Y.; Ji, H.; Wang, Y.; Wu, Q.; Bao, J.; Ye, F.; Shi, Y.; Bu, H. Loss of histone deacetylases 1 and 2 in hepatocytes impairs murine liver regeneration through Ki67 depletion. Hepatology 2013, 58, 2089–2098. [Google Scholar] [CrossRef]
- Dor, Y.; Brown, J.; Martinez, O.I.; Melton, D.A. Adult pancreatic beta-cells are formed by self-duplication rather than stem-cell differentiation. Nature 2004, 429, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Pan, F.C.; Bankaitis, E.D.; Boyer, D.; Xu, X.; Van de Casteele, M.; Magnuson, M.A.; Heimberg, H.; Wright, C.V.E. Spatiotemporal patterns of multipotentiality in Ptf1aexpressing cells during pancreas organogenesis and injuryinduced facultative restoration. Development 2013, 140, 751–764. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, N.; Ninov, N.; Delawary, M.; Osman, S.; Roh, A.S.; Gut, P.; Stainier, D.Y.R. Whole organism high content screening identifies stimulators of pancreatic beta-cell proliferation. PLoS ONE 2014, 9, e104112. [Google Scholar] [CrossRef] [Green Version]
- Basile, G.; Kulkarni, R.N.; Morgan, N.G. How, When, and Where Do Human β-Cells Regenerate? Curr. Diabetes Rep. 2019, 19, 48. [Google Scholar] [CrossRef]
- Sharma, A.; Purohit, S.; Sharma, S.; Bai, S.; Zhi, W.; Ponny, S.R.; Hopkins, D.; Steed, L.; Bode, B.; Anderson, S.W.; et al. IGF-binding proteins in type-1 diabetes are more severely altered in the presence of complications. Front. Endocrinol. (Lausanne) 2016, 7, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inada, A.; Nienaber, C.; Katsuta, H.; Fujitani, Y.; Levine, J.; Morita, R.; Sharma, A.; Bonner-Weir, S. Carbonic anhydrase II-positive pancreatic cells are progenitors for both endocrine and exocrine pancreas after birth. Proc. Natl. Acad. Sci. USA 2008, 105, 19915–19919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninov, N.; Hesselson, D.; Gut, P.; Zhou, A.; Fidelin, K.; Stainier, D.Y.R. Metabolic regulation of cellular plasticity in the pancreas. Curr. Biol. 2013, 23, 1242–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghaye, A.P.; Bergemann, D.; Tarifeño-Saldivia, E.; Flasse, L.C.; Von Berg, V.; Peers, B.; Voz, M.L.; Manfroid, I. Progenitor potential of nkx6.1-expressing cells throughout zebrafish life and during beta cell regeneration. BMC Biol. 2015, 13, 70. [Google Scholar] [CrossRef] [Green Version]
- Delaspre, F.; Beer, R.L.; Rovira, M.; Huang, W.; Wang, G.; Gee, S.; Del Carmen Vitery, M.; Wheelan, S.J.; Parsons, M.J. Centroacinar cells are progenitors that contribute to endocrine pancreas regeneration. Diabetes 2015, 64, 3499–3509. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.C.; Leuckx, G.; Sakano, D.; Seymour, P.A.; Mattsson, C.L.; Rautio, L.; Staels, W.; Verdonck, Y.; Serup, P.; Kume, S.; et al. Inhibition of Cdk5 promotes β-cell differentiation from ductal progenitors. Diabetes 2018, 67, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Jurisch-Yaksi, N.; Yaksi, E.; Kizil, C. Radial glia in the zebrafish brain: Functional, structural, and physiological comparison with the mammalian glia. Glia 2020, 68, 2451–2470. [Google Scholar] [CrossRef]
- Asher, R.A.; Morgenstern, D.A.; Fidler, P.S.; Adcock, K.H.; Oohira, A.; Braistead, J.E.; Levine, J.M.; Margolis, R.U.; Rogers, J.H.; Fawcett, J.W. Neurocan is upregulated in injured brain and in cytokine-treated astrocytes. J. Neurosci. 2000, 20, 2427–2438. [Google Scholar] [CrossRef] [Green Version]
- Stichel, C.C.; Hermanns, S.; Luhmann, H.J.; Lausberg, F.; Niermann, H.; D’Urso, D.; Servos, G.; Hartwig, H.G.; Müller, H.W. Inhibition of collagen IV deposition promotes regeneration of injured CNS axons. Eur. J. Neurosci. 1999, 11, 632–646. [Google Scholar] [CrossRef]
- McKeon, R.J.; Schreiber, R.C.; Rudge, J.S.; Silver, J. Reduction of neurite outgrowth in a model of glial scarring following CNS injury is correlated with the expression of inhibitory molecules on reactive astrocytes. J. Neurosci. 1991, 11, 3398–3411. [Google Scholar] [CrossRef] [Green Version]
- Goldshmit, Y.; Sztal, T.E.; Jusuf, P.R.; Hall, T.E.; Nguyen-Chi, M.; Currie, P.D. Fgf-dependent glial cell bridges facilitate spinal cord regeneration in zebrafish. J. Neurosci. 2012, 32, 7477–7492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cigliola, V.; Becker, C.J.; Poss, K.D. Building bridges, not walls: Spinal cord regeneration in zebrafish. Dis. Models Mech. 2020, 13, 5. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Hui, S.P. Axonal regeneration in zebrafish spinal cord. Regeneration 2018, 5, 43–60. [Google Scholar] [CrossRef] [PubMed]
- Reimer, M.M.; Kuscha, V.; Wyatt, C.; Sörensen, I.; Frank, R.E.; Knüwer, M.; Becker, T.; Becker, C.G. Sonic hedgehog is a polarized signal for motor neuron regeneration in adult zebrafish. J. Neurosci. 2009, 29, 15073–15082. [Google Scholar] [CrossRef]
- Goldshmit, Y.; Frisca, F.; Kaslin, J.; Pinto, A.R.; Tang, J.-K.K.Y.; Pébay, A.; Pinkas-Kramarski, R.; Currie, P.D. Decreased anti-regenerative effects after spinal cord injury in spry4-/- mice. Neuroscience 2015, 287, 104–112. [Google Scholar] [CrossRef]
- Tsai, M.-C.; Shen, L.-F.; Kuo, H.-S.; Cheng, H.; Chak, K.-F. Involvement of acidic fibroblast growth factor in spinal cord injury repair processes revealed by a proteomics approach. Mol. Cell. Proteom. 2008, 7, 1668–1687. [Google Scholar] [CrossRef] [Green Version]
- Kasai, M.; Jikoh, T.; Fukumitsu, H.; Furukawa, S. FGF-2-responsive and spinal cord-resident cells improve locomotor function after spinal cord injury. J. Neurotrauma 2014, 31, 1584–1598. [Google Scholar] [CrossRef] [Green Version]
- Sakai, K.; Yamamoto, A.; Matsubara, K.; Nakamura, S.; Naruse, M.; Yamagata, M.; Sakamoto, K.; Tauchi, R.; Wakao, N.; Imagama, S.; et al. Human dental pulp-derived stem cells promote locomotor recovery after complete transection of the rat spinal cord by multiple neuro-regenerative mechanisms. J. Clin. Investig. 2012, 122, 80–90. [Google Scholar] [CrossRef]
- Nagashima, K.; Miwa, T.; Soumiya, H.; Ushiro, D.; Takeda-Kawaguchi, T.; Tamaoki, N.; Ishiguro, S.; Sato, Y.; Miyamoto, K.; Ohno, T.; et al. Priming with FGF2 stimulates human dental pulp cells to promote axonal regeneration and locomotor function recovery after spinal cord injury. Sci. Rep. 2017, 7, 13500. [Google Scholar] [CrossRef] [Green Version]
- Zambusi, A.; Ninkovic, J. Regeneration of the central nervous system-principles from brain regeneration in adult zebrafish. World J. Stem Cells 2020, 12, 8–24. [Google Scholar] [CrossRef]
- Lange, C.; Brand, M. Vertebrate brain regeneration—A community effort of fate-restricted precursor cell types. Curr. Opin. Genet. Dev. 2020, 64, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Kroehne, V.; Freudenreich, D.; Hans, S.; Kaslin, J.; Brand, M. Regeneration of the adult zebrafish brain from neurogenic radial glia-type progenitors. Development 2011, 138, 4831–4841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celikkaya, H.; Cosacak, M.I.; Papadimitriou, C.; Popova, S.; Bhattarai, P.; Biswas, S.N.; Siddiqui, T.; Wistorf, S.; Nevado-Alcalde, I.; Naumann, L.; et al. GATA3 promotes the neural progenitor state but not neurogenesis in 3D traumatic injury model of primary human cortical astrocytes. Front. Cell. Neurosci. 2019, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, C.; Celikkaya, H.; Cosacak, M.I.; Mashkaryan, V.; Bray, L.; Bhattarai, P.; Brandt, K.; Hollak, H.; Chen, X.; He, S.; et al. 3D Culture Method for Alzheimer’s Disease Modeling Reveals Interleukin-4 Rescues Aβ42-Induced Loss of Human Neural Stem Cell Plasticity. Dev. Cell 2018, 46, 85–101.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langhe, R.; Pearson, R.A. Rebuilding the Retina: Prospects for Müller Glial-mediated Self-repair. Curr. Eye Res. 2020, 45, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Roesch, K.; Jadhav, A.P.; Trimarchi, J.M.; Stadler, M.B.; Roska, B.; Sun, B.B.; Cepko, C.L. The transcriptome of retinal Müller glial cells. J. Comp. Neurol. 2008, 509, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Cepko, C.; Dyer, M.A. Control of Müller glial cell proliferation and activation following retinal injury. Nat. Neurosci. 2000, 3, 873–880. [Google Scholar]
- Hamon, A.; Roger, J.E.; Yang, X.J.; Perron, M. Müller glial cell-dependent regeneration of the neural retina: An overview across vertebrate model systems. Dev. Dyn. 2016, 245, 727–738. [Google Scholar] [CrossRef]
- Fausett, B.V.; Gumerson, J.D.; Goldman, D. The proneural basic helix-loop-helix gene Ascl1a is required for retina regeneration. J. Neurosci. 2008, 28, 1109–1117. [Google Scholar] [CrossRef]
- Pollak, J.; Wilken, M.S.; Ueki, Y.; Cox, K.E.; Sullivan, J.M.; Taylor, R.J.; Levine, E.M.; Reh, T.A. ASCL1 reprograms mouse Müller glia into neurogenic retinal progenitors. Development 2013, 140, 2619–2631. [Google Scholar] [CrossRef] [Green Version]
- Ueki, Y.; Wilken, M.S.; Cox, K.E.; Chipman, L.; Jorstad, N.; Sternhagen, K.; Simic, M.; Ullom, K.; Nakafuku, M.; Reh, T.A. Transgenic expression of the proneural transcription factor Ascl1 in Müller glia stimulates retinal regeneration in young mice. Proc. Natl. Acad. Sci. USA 2015, 112, 13717–13722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramachandran, R.; Fausett, B.V.; Goldman, D. Ascl1A regulates Muller glia dedifferentiation and retina regeneration via a Lin-28 dependent, let7 miRNA signaling pathway. Nat. Cell Biol. 2010, 7, 1959–1967. [Google Scholar]
- Pisharath, H.; Rhee, J.M.; Swanson, M.A.; Leach, S.D.; Parsons, M.J. Targeted ablation of beta cells in the embryonic zebrafish pancreas using E.coli nitroreductase. Mech. Dev. 2007, 124, 218–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curado, S.; Anderson, R.M.; Jungblut, B.; Mumm, J.; Schroeter, E.; Stainier, D.Y.R. Conditional targeted cell ablation in zebrafish: A new tool for regeneration studies. Dev. Dyn. 2007, 236, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Bergemann, D.; Massoz, L.; Bourdouxhe, J.; Carril Pardo, C.A.; Voz, M.L.; Peers, B.; Manfroid, I. Nifurpirinol: A more potent and reliable substrate compared to metronidazole for nitroreductase-mediated cell ablations. Wound Repair Regen. 2018, 26, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Sharrock, A.; Mulligan, T.; Hall, K.; Williams, E.; White, D.; Zhang, L.; Matthews, F.; Nimmagadda, S.; Washington, S.; Le, K.; et al. NTR 2.0: A rationally-engineered prodrug converting enzyme with substantially enhanced efficacy for targeted cell ablation. bioRxiv 2020, 1–26. [Google Scholar] [CrossRef]
- Wilkinson, P.D.; Alencastro, F.; Delgado, E.R.; Leek, M.P.; Weirich, M.P.; Otero, P.A.; Roy, N.; Brown, W.K.; Oertel, M.; Duncan, A.W. Polyploid Hepatocytes Facilitate Adaptation and Regeneration to Chronic Liver Injury. Am. J. Pathol. 2019, 189, 1241–1255. [Google Scholar] [CrossRef]
- Donne, R.; Saroul-Aïnama, M.; Cordier, P.; Celton-Morizur, S.; Desdouets, C. Polyploidy in liver development, homeostasis and disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 391–405. [Google Scholar] [CrossRef]
- Julier, Z.; Park, A.J.; Briquez, P.S.; Martino, M.M. Promoting tissue regeneration by modulating the immune system. Acta Biomater. 2017, 53, 13–28. [Google Scholar] [CrossRef]
- Lai, S.L.; Marín-Juez, R.; Moura, P.L.; Kuenne, C.; Lai, J.K.H.; Tsedeke, A.T.; Guenther, S.; Looso, M.; Stainier, D.Y.R. Reciprocal analyses in zebrafish and medaka reveal that harnessing the immune response promotes cardiac regeneration. Elife 2017, 6, 1–20. [Google Scholar] [CrossRef]
- Ryan, R.; Moyse, B.R.; Richardson, R.J. Zebrafish cardiac regeneration-looking beyond cardiomyocytes to a complex microenvironment. Histochem. Cell Biol. 2020, 154, 533–548. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Ochi, H. Regeneration enhancers: A clue to reactivation of developmental genes. Dev. Growth Regen. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zafar, H.; Lin, C.; Bar-Joseph, Z. Single-cell lineage tracing by integrating CRISPR-Cas9 mutations with transcriptomic data. Nat. Commun. 2020, 11, 305. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massoz, L.; Dupont, M.A.; Manfroid, I. Zebra-Fishing for Regenerative Awakening in Mammals. Biomedicines 2021, 9, 65. https://doi.org/10.3390/biomedicines9010065
Massoz L, Dupont MA, Manfroid I. Zebra-Fishing for Regenerative Awakening in Mammals. Biomedicines. 2021; 9(1):65. https://doi.org/10.3390/biomedicines9010065
Chicago/Turabian StyleMassoz, Laura, Marie Alice Dupont, and Isabelle Manfroid. 2021. "Zebra-Fishing for Regenerative Awakening in Mammals" Biomedicines 9, no. 1: 65. https://doi.org/10.3390/biomedicines9010065
APA StyleMassoz, L., Dupont, M. A., & Manfroid, I. (2021). Zebra-Fishing for Regenerative Awakening in Mammals. Biomedicines, 9(1), 65. https://doi.org/10.3390/biomedicines9010065