
Plague Prevention and Therapy: Perspectives on Current and Future Strategies


Abstract
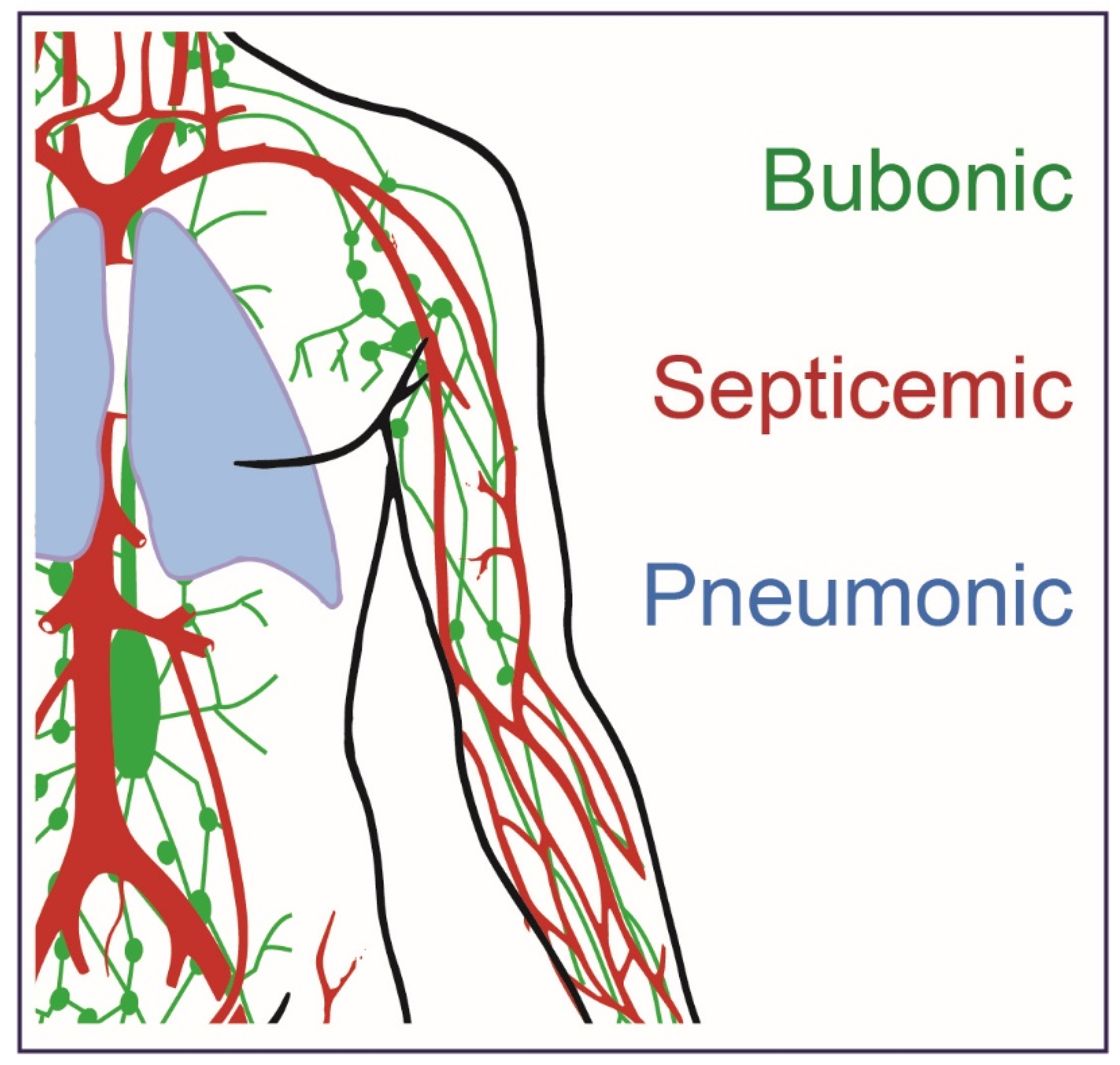
:1. Plague History and Disease Presentation
2. Yersinia pestis Virulence Plasmids
3. Outbreak Prevention
4. Current Antibiotic Therapies
5. Vaccines
6. Antibodies
7. Advantages of Monoclonal Antibodies (mAbs) and Final Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Pechous, R.D.; Sivaraman, V.; Stasulli, N.M.; Goldman, W.E. Pneumonic plague: The darker side of Yersinia pestis. Trends Microbiol. 2016, 24, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Demeure, C.E.; Dussurget, O.; Fiol, G.M.; Le Guern, A.S.; Savin, C.; Pizzarro-Cerdá, J. Yersinia pestis and plague: An updated view on evolution, virulence determinants, immune subversion, vaccination, and diagnostics. Genes Immun. 2019, 20, 357–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenseth, N.C.; Atshabar, B.B.; Begon, M.; Belmain, S.R.; Bertherat, E.; Carniel, E.; Gage, K.L.; Leirs, H.; Rahalison, L. Plague: Past, present, and future. PLoS Med. 2008, 5, e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achtman, M.; Zurth, K.; Morelli, G.; Torrea, G.; Guiyoule, A.; Carniel, E. Yersinia pestis, the cause of plague, is a recently emerged clone of Yersinia pseudotuberculosis. Proc. Natl. Acad. Sci. USA 1999, 96, 14043–14048. [Google Scholar] [CrossRef] [Green Version]
- Glatter, K.A.; Finkelman, P. History of the plague: An ancient pandemic for the age of COVID-19. Am. J. Med. 2021, 134, 176–181. [Google Scholar] [CrossRef]
- Brubaker, R.R. Yersinia pestis. Mol. Med. Microbiol. 2015, 3, 1845–1865. [Google Scholar]
- Frith, J. The history of plague—Part 1. The three great pandemics. J. Mil. Vet. Health 2012, 20, 11–16. [Google Scholar]
- Barbieri, R.; Signoli, M.; Chevé, D.; Costedoat, C.; Tzortzis, S.; Aboudharam, G.; Raoult, D.; Drancourt, M. Yersinia pestis: The natural history of plague. Clin. Microbiol. Rev. 2020, 34, e00044-19. [Google Scholar] [CrossRef]
- Drancourt, M.; Raoult, D. Molecular history of plague. Clin. Microbiol. Infect. 2016, 22, 911–915. [Google Scholar] [CrossRef] [Green Version]
- Bramanti, B.; Dean, K.R.; Walløe, L.; Stenseth, N.C. The third plague pandemic in Europe. Proc. Biol. Sci. 2019, 286, 20182429. [Google Scholar] [CrossRef]
- Eisen, R.J.; Gage, K.L. Adaptive strategies of Yersinia pestis to persist during inter-epizootic and epizootic periods. Vet. Res. 2009, 40, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raoult, D.; Mouffok, N.; Bitam, I.; Piarroux, R.; Drancourt, M. Plague: History and contemporary analysis. J. Infect. 2013, 66, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Melman, S.D.; Ettestad, P.E.; VinHatton, E.S.; Ragsdale, J.M.; Takacs, N.; Onischuk, L.M.; Leonard, P.M.; Master, S.S.; Lucero, V.S.; Kingry, L.C. Human case of bubonic plague resulting from the bite of a wild Gunnison’s prairie dog during translocation from a plague-endemic area. Zoonoses Public Health 2018, 65, e254–e258. [Google Scholar] [CrossRef] [Green Version]
- Dai, R.; Wei, B.; Xiong, H.; Yang, X.; Peng, Y.; He, J.; Jin, J.; Wang, Y.; Zha, X.; Zhang, Z.; et al. Human plague associated with Tibetan sheep originates in marmots. PLoS Negl. Trop. Dis. 2018, 12, e0006635. [Google Scholar] [CrossRef] [Green Version]
- Richgels, K.L.; Russell, R.E.; Bron, G.M.; Rocke, T.E. Evaluation of Yersinia pestis transmission pathways for sylvatic plague in prairie dog populations in the Western U.S. Ecohealth 2016, 13, 415–427. [Google Scholar] [CrossRef] [PubMed]
- CDC. Fatal laboratory-acquired infection with an attenuated Yersinia pestis Strain—Chicago, Illinois, 2009. MMWR Morb. Mortal. Wkly. Rep. 2011, 60, 201–205. [Google Scholar]
- Worsham, P.L.; McGovern, T.W.; Vietri, N.J.; Friedlander, A.M.; Bozue, J. Plague. In Textbook of Military Medicine: Medical Aspects of Biological Warfare; Bozue, J.A., Cote, C.K., Glass, P.J., Eds.; Borden Institute US Army Medical Department Center and School Health Readiness Center of Excellence: Houston, TX, USA, 2018; pp. 247–284. [Google Scholar]
- Yang, R. Plague: Recognition, treatment, and prevention. J. Clin. Microbiol. 2017, 56, e01519-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikiforov, V.V.; Gao, H.; Zhou, L.; Anisimov, A. Plague: Clinics, diagnosis and treatment. Adv. Exp. Med. Biol. 2016, 918, 293–312. [Google Scholar]
- Dennis, D.T.; Gage, K.L.; Gratz, N.G.; Poland, J.D.; Tikhomirov, E. Plague Manual: Epidemiology, Distribution, Surveillance and Control; No. WHO/CDS/CSR/EDC/99.2; World Health Organization: Geneva, Switzeralnd, 1999. [Google Scholar]
- Sebbane, F.; Lemaitre, N. Antibiotic therapy of plague: A review. Biomolecules 2021, 11, 724. [Google Scholar] [CrossRef]
- Bossi, P.; Tagnell, A.; Baka, A.; Loock, F.V.; Hendriks, J.; Werner, A.; Maidhof, H.; Gouvras, G. Bichat guidelines for the clinical management of plague and bioterrorism-related plague. Euro Surveill 2004, 9, E5–E6. [Google Scholar] [CrossRef]
- Lemaître, N.; Ricard, I.; Pradel, E.; Foligné, B.; Courcol, R.; Simonet, M.; Sebbane, F. Efficacy of ciprofloxacin-gentamicin combination therapy in murine bubonic plague. PLoS ONE 2012, 7, e52503. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, L.L.; Ettestad, P.; Fogarty, J.D.; Dennis, D.T.; Romig, D.; Mertz, G. Gentamicin and tetracyclines for the treatment of human plague: Review of 75 cases in New Mexico, 1985–1999. Clin. Infect. Dis. 2004, 38, 663–669. [Google Scholar] [CrossRef]
- Lawrenz, M.B. Model systems to study plague pathogenesis and develop new therapeutics. Front. Microbiol. 2010, 1, 119. [Google Scholar] [CrossRef] [Green Version]
- Dillard, R.L.; Juergens, A.L. Plague; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Cleri, D.J.; Marton, R.; Rabbat, M.; Vernaleo, J. Pneumonia Caused by Yersinia pestis: Plague Pneumonia, in Community-Acquired Pneumonia; Marrie, T.J., Ed.; Springer: Boston, MA, USA, 2001; pp. 777–799. [Google Scholar]
- Kool, J. Risk of person-to-person transmission of pneumonic plague. Clin. Infect. Dis. 2005, 40, 1166–1172. [Google Scholar] [CrossRef] [PubMed]
- Dennis, D.T.; Mead, P.S. Plague. Trop. Infect. Dis. 2006, 1, 471–481. [Google Scholar]
- Gage, K.L.; Beard, C.B. 126—Plague. In Infectious Diseases, 4th ed.; Cohen, J., Powderly, W.G., Opal, S.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1078–1084.e1. [Google Scholar]
- Wagar, E. Bioterrorism and the role of the clinical microbiology laboratory. Clin. Microbiol. Rev. 2016, 29, 175–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, I.; Grier, G.; Byers, M. Deliberate release: Plague—A review. J. Biosaf. Biosecur. 2020, 2, 10–22. [Google Scholar] [CrossRef]
- Du, Z.; Wang, X. Pathology and pathogenesis of Yersinia pestis. Adv. Exp. Med. Biol. 2016, 918, 193–222. [Google Scholar]
- Hewitt, J.A.; Lanning, L.L.; Campbell, J.L. The African green monkey model of pneumonic plague and US Food and Drug Administration approval of antimicrobials under the animal rule. Clin. Infect. Dis. 2020, 70 (Suppl. S1), S51–S59. [Google Scholar] [CrossRef]
- Warren, R.; Lockman, H.; Barnewall, R.; Krile, R.; Bermeo Blanco, O.; Vasconcelos, D.; Price, J.; House, R.V.; Bolanowksi, M.A.; Fellows, P. Cynomolgus macaque model for pneumonic plague. Microb. Pathog. 2011, 50, 12–22. [Google Scholar] [CrossRef]
- Bubeck, S.S.; Cantwell, A.M.; Dube, P.H. Delayed inflammatory response to primary pneumonic plague occurs in both outbred and inbred mice. Infect. Immun. 2007, 75, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Lathem, W.W.; Crosby, S.D.; Miller, V.L.; Goldman, W.E. Progression of primary pneumonic plague: A mouse model of infection, pathology, and bacterial transcriptional activity. Proc. Natl. Acad. Sci. USA 2005, 102, 17786–17791. [Google Scholar] [CrossRef] [Green Version]
- Pechous, R.D.; Sivaraman, V.; Price, P.A.; Stasulli, N.M.; Goldman, W.E. Early host cell targets of Yersinia pestis during primary pneumonic plague. PLoS Pathog. 2013, 9, e1003679. [Google Scholar] [CrossRef]
- Banerjee, S.K.; Crane, S.D.; Pechous, R.D. A dual role for the plasminogen activator protease during the preinflammatory phase of primary pneumonic plague. J. Infect. Dis. 2020, 222, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Pechous, R.D.; Broberg, C.A.; Stasulli, N.M.; Miller, V.L.; Goldman, W.E. In vivo transcriptional profiling of Yersinia pestis reveals a novel bacterial mediator of pulmonary inflammation. mBio 2015, 6, e02302-14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, D.; Yang, R. Molecular Darwinian evolution of virulence in Yersina pestis. Infect. Immun. 2020, 77, 2242–2250. [Google Scholar] [CrossRef] [Green Version]
- Skurnik, M.; Peippo, A.; Ervelä, E. Characterization of the O-antigen gene clusters of Yersinia pseudotuberculosis and the cryptic O-antigen gene cluster of Yersinia pestis shows that the plague bacillus is most closely related to and has evolved from Y. pseudotuberculosis serotype O:1b. Mol. Microbiol. 2000, 37, 316–330. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, A.E.; Stuckey, J.A.; Zhao, Y.; Matthews, H.R.; Patton, W.A.; Moss, I.; Dixon, J.E. Expression, characterization, and mutagenesis of the Yersinia pestis murine toxin, a phospholipase D superfamily member. J. Biol. Chem. 1999, 274, 11824–11831. [Google Scholar] [CrossRef] [Green Version]
- Schar, M.; Meyer, K.F. Studies on immunization against plague. XV. The pathophysiologic action of the toxin of Pasteurella pestis in experimental animals. Schweiz Z Pathol. Bakteriol. 1956, 19, 51–70. [Google Scholar]
- Bergdoll, M.S.; Enterotoxins, T.C.; Montie, S.J.A. Microbial Toxins; Academic Press: New York, NY, USA, 1970; Volume 3, pp. 265–326. [Google Scholar]
- Hinnebusch, B.J.; Rudolph, A.E.; Cherepanov, P.; Dixon, J.E.; Schwan, T.G. Role of Yersinia murine toxin in survival of Yersinia pestis in the midgut of the flea vector. Science 2002, 296, 733–735. [Google Scholar] [CrossRef]
- Chen, T.H.; Crocker, T.T.; Meyer, K.F. Electron microscopic study of the extracellular materials of Pasteurella pestis. J. Bacteriol. 1956, 72, 851–857. [Google Scholar]
- Davis, K.J.; Fritz, D.L.; Pitt, M.L.; Welkos, S.L.; Worsham, P.L.; Friedlander, A.M. Pathology of experimental pneumonic plague produced by fraction 1-positive and fraction 1-negative Yersinia pestis in African green monkeys (Cercopithecus aethiops). Arch. Pathol. Lab. Med. 1996, 120, 156–163. [Google Scholar]
- Engelsberg, E.; Levy, J.B. Studies on immunization against plague. VI. Growth of Pasteurella pestis and the production of the envelope and other soluble antigens in a casein hydrolyzate mineral glucose medium. J. Bacteriol. 1954, 67, 438–449. [Google Scholar] [CrossRef] [Green Version]
- Runco, L.M.; Myrczek, S.; Bliska, J.B.; Thanassi, D.G. Biogenesis of the fraction 1 capsule and analysis of the ultrastructure of Yersinia pestis. J. Bacteriol. 2008, 190, 3381–3385. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Rosqvist, R.; Forsberg, Å. Role of fraction 1 antigen of Yersinia pestis in inhibition of phagocytosis. Infect. Immun. 2002, 70, 1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welkos, S.L.; Davis, K.M.; Pitt, L.M.; Worsham, P.L.; Friedlander, A.M. Studies on the contribution of the F1 capsule-associated plasmid pFra to the virulence of Yersinia pestis. Contrib. Microbiol. Immunol. 1995, 13, 299–305. [Google Scholar] [PubMed]
- Winter, C.C.; Cherry, W.B.; Moody, M.D. An unusual strain of Pasteurella pestis isolated from a fatal human case of plague. Bull. World Health Organ. 1960, 23, 408–409. [Google Scholar]
- Sebbane, F.; Uversky, V.N.; Anisimov, A.P. Yersinia pestis plasminogen activator. Biomolecules 2020, 10, 1554. [Google Scholar] [CrossRef] [PubMed]
- Caulfield, A.J.; Lathem, W.W. Substrates of the plasminogen activator protease of Yersinia pestis. Adv. Exp. Med. Biol. 2012, 954, 253–260. [Google Scholar]
- Degen, J.L.; Bugge, T.H.; Goguen, J.D. Fibrin and fibrinolysis in infection and host defense. J. Thromb. Haemost. 2007, 5 (Suppl. S1), 24–31. [Google Scholar] [CrossRef]
- Sodeinde, O.A.; Subrahmanyam, Y.V.; Stark, K.; Quan, T.; Bao, Y.; Goguen, J.D. A surface protease and the invasive character of plague. Science 1992, 258, 1004–1007. [Google Scholar] [CrossRef]
- Caulfield, A.J.; Walker, M.E.; Gielda, L.M.; Lathem, W.W. The Pla protease of Yersinia pestis degrades fas ligand to manipulate host cell death and inflammation. Cell Host Microbe 2014, 15, 424–434. [Google Scholar] [CrossRef] [Green Version]
- Lathem, W.W.; Price, P.A.; Miller, V.L.; Goldman, W.E. A plasminogen-activating protease specifically controls the development of primary pneumonic plague. Science 2007, 315, 509–513. [Google Scholar] [CrossRef]
- Kienle, Z.; Emödy, L.; Svanborg, C.; O’Toole, P.W. Adhesive properties conferred by the plasminogen activator of Yersinia pestis. J. Gen. Microbiol. 1992, 138, 1679–1687. [Google Scholar] [CrossRef] [Green Version]
- Lähteenmäki, K.; Virkola, R.; Sarén, A.; Emödy, L.; Korhonen, T.K. Expression of plasminogen activator Pla of Yersinia pestis enhances bacterial attachment to the mammalian extracellular matrix. Infect. Immun. 1998, 66, 5755–5762. [Google Scholar] [CrossRef] [Green Version]
- Lobo, L.A. Adhesive properties of the purified plasminogen activator Pla of Yersinia pestis. FEMS Microbiol. Lett. 2006, 262, 158–162. [Google Scholar] [CrossRef] [Green Version]
- Cowan, C.; Jones, H.A.; Kaya, Y.H.; Perry, R.D.; Straley, S.C. Invasion of epithelial cells by Yersinia pestis: Evidence for a Y. pestis-specific invasin. Infect. Immun. 2000, 68, 4523–4530. [Google Scholar] [CrossRef] [Green Version]
- Lähteenmäki, K.; Kukkonen, M.; Korhonen, T.K. The Pla surface protease/adhesin of Yersinia pestis mediates bacterial invasion into human endothelial cells. FEBS Lett. 2001, 504, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Fields, K.A.; Straley, S.C. LcrV of Yersinia pestis enters infected eukaryotic cells by a virulence plasmid-independent mechanism. Infect. Immun. 1999, 67, 4801–4813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derewenda, U.; Mateja, A.; Devedjiev, Y.; Routzahn, K.M.; Evdokimov, A.G.; Derewenda, Z.S.; Waugh, D.S. The structure of Yersinia pestis V-antigen, an essential virulence factor and mediator of immunity against plague. Structure 2004, 12, 301–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, S.B.; Cowan, C.; Perry, R.D.; Straley, S.C. The Yersinia pestis V antigen is a regulatory protein necessary for Ca2(+)-dependent growth and maximal expression of low-Ca2+ response virulence genes. J. Bacteriol. 1991, 173, 2649–2657. [Google Scholar] [CrossRef] [Green Version]
- Hamad, M.A.; Nilles, M.L. Structure-function analysis of the C-terminal domain of LcrV from Yersinia pestis. J. Bacteriol. 2007, 189, 6734–6739. [Google Scholar] [CrossRef] [Green Version]
- DiMezzo, T.L.; Ruthel, G.; Brueggemann, E.E.; Hines, H.B.; Ribot, W.J.; Chapman, C.E.; Powell, B.S.; Welkos, S.L. In vitro intracellular trafficking of virulence antigen during infection by Yersinia pestis. PLoS ONE 2009, 4, e6281. [Google Scholar] [CrossRef] [PubMed]
- Sing, A.; Roggenkamp, A.; Geiger, A.M.; Heesemann, J. Yersinia enterocolitica evasion of the host innate immune response by V antigen-induced IL-10 production of macrophages is abrogated in IL-10-deficient mice. J. Immunol. 2002, 168, 1315–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.D.; Haddix, P.; Atkins, E.B.; Soughers, T.K.; Straley, S.C. Regulation of expression of V antigen and outer membrane proteins in Yersinia pestis. Contrib. Microbiol. Immunol. 1987, 9, 173–178. [Google Scholar] [PubMed]
- Perry, R.D.; Harmon, P.A.; Bowmer, W.S.; Straley, S.C. A low-Ca2+ response operon encodes the V antigen of Yersinia pestis. Infect. Immun. 1986, 54, 428–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilles, M.L.; Williams, A.W.; Skrzypek, E.; Straley, S.C. Yersinia pestis LcrV forms a stable complex with LcrG and may have a secretion-related regulatory role in the low-Ca2+ response. J. Bacteriol. 1997, 179, 1307–1316. [Google Scholar] [CrossRef] [Green Version]
- Fields, K.A.; Nilles, M.L.; Cowan, C.; Straley, S.C. Virulence role of V antigen of Yersinia pestis at the bacterial surface. Infect. Immun. 1999, 67, 5395–5408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philipovskiy, A.V.; Cowan, C.; Wulff-Strobel, C.R.; Burnett, S.H.; Kerschen, E.J.; Cohen, D.A.; Kaplan, A.M.; Straley, S.C. Antibody against V antigen prevents Yop-dependent growth of Yersinia pestis. Infect. Immun. 2005, 73, 1532–1542. [Google Scholar] [CrossRef] [Green Version]
- Nilles, M.L.; Fields, K.A.; Straley, S.C. The V antigen of Yersinia pestis regulates Yop vectorial targeting as well as Yop secretion through effects on YopB and LcrG. J. Bacteriol. 1998, 180, 3410–3420. [Google Scholar] [CrossRef] [Green Version]
- Rosqvist, R.; Forsberg, A.; Rimpiläinen, M.; Bergman, T.; Wolf-Watz, H. The cytotoxic protein YopE of Yersinia obstructs the primary host defence. Mol. Microbiol. 1990, 4, 657–667. [Google Scholar] [CrossRef]
- Bliska, J.B.; Black, D.S. Inhibition of the Fc receptor-mediated oxidative burst in macrophages by the Yersinia pseudotuberculosis tyrosine phosphatase. Infect. Immun. 1995, 63, 681–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fällman, M.; Andersson, K.; Håkansson, S.; Magnusson, K.E.; Stendahl, O.; Wolf-Watz, H. Yersinia pseudotuberculosis inhibits Fc receptor-mediated phagocytosis in J774 cells. Infect. Immun. 1995, 63, 3117–3124. [Google Scholar] [CrossRef] [Green Version]
- Grosdent, N.; Maridonneau-Parini, I.; Paule Sory, M.; Cornelis, G.R. Role of Yops and adhesins in resistance of Yersinia enterocolitica to phagocytosis. Infect. Immun. 2002, 70, 4165–4176. [Google Scholar] [CrossRef] [Green Version]
- Visser, L.G.; Annema, A.; van Furth, R. Role of Yops in inhibition of phagocytosis and killing of opsonized Yersinia enterocolitica by human granulocytes. Infect. Immun. 1995, 63, 2570–2575. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Schneewind, O. Yersinia enterocolitica type III secretion: An mRNA signal that couples translation and secretion of YopQ. Mol. Microbiol. 1999, 31, 1139–1148. [Google Scholar] [CrossRef]
- Heitzinger, K.; Impouma, B.; Farham, B.L.; Hambion, E.L.; Lukoya, C.; Machingaidze, C.; Rakotonjanabelo, L.A.; Yao, M.; Diallo, B.; Djingarey, M.H.; et al. Using evidence to inform response to the 2017 plague outbreak in Madagascar: A view from the WHO African Regional Office. Epidemiol. Infect. 2018, 147, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.K.; Parra-Rojas, C.; Hernandez-Vargas, E.A. The 2017 plague outbreak in Madagascar: Data descriptions and epidemic modelling. Epidemics 2018, 25, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Vallès, X.; Stenseth, N.C.; Demeure, C.; Horby, P.; Mead, P.S.; Cabanillas, O.; Ratsitorahina, M.; Rajerison, M.; Andrianaivoarimanana, V.; Ramasindrazana, B.; et al. Human plague: An old scourge that needs new answers. PLoS Negl. Trop. Dis. 2020, 14, e0008251. [Google Scholar] [CrossRef]
- Carniel, E. Plague today. Med. Hist. Suppl. 2008, 52, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Bertherat, E. Plague around the world, 2010–2015. Wkly. Epidemiol. Rec. 2016, 91, 89–93. [Google Scholar]
- Bevins, S.N.; Baroch, J.A.; Nolte, D.L.; Zhang, M.; He, H. Yersinia pestis: Examining wildlife plague surveillance in China and the USA. Integr. Zool. 2012, 7, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Alderson, J.; Quastel, M.; Wilson, E.; Bellamy, D. Factors influencing the re-emergence of plague in Madagascar. Emerg. Top. Life Sci. 2020, 4, 411–421. [Google Scholar] [PubMed]
- Randremanana, R.; Andrianaivoarimanana, V.; Nikolay, B.; Ramasindrazana, B.; Paireau, J.; Ten Bosch, Q.A.; Rakotondramanga, J.M.; Rahajandraibe, S.; Rahelinirina, S.; Rakotomanana, F.; et al. Epidemiological characteristics of an urban plague epidemic in Madagascar, August–November, 2017: An outbreak report. Lancet Infect. Dis. 2019, 19, 537–545. [Google Scholar] [CrossRef] [Green Version]
- Andrianaivoarimanana, V.; Piola, P.; Wagner, D.M.; Rakotomanana, F.; Maheriniaina, V.; Andrianalimanana, S.; Chanteau, S.; Rahalison, L.; Ratsitorahina, M.; Rajerison, M. Trends of human plague, Madagascar, 1998–2016. Emerg. Infect. Dis. 2019, 25, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Rabaan, A.A.; Al-Ahmed, S.H.; Alsuliman, S.A.; Aldrazi, F.A.; Alfouzan, W.A.; Haque, S. The rise of pneumonic plague in Madagascar: Current plague outbreak breaks usual seasonal mould. J. Med. Microbiol. 2019, 68, 292–302. [Google Scholar] [CrossRef]
- Barbieri, R.; Drancourt, M.; Raoult, D. The role of louse-transmitted diseases in historical plague pandemics. Lancet Infect. Dis. 2021, 21, e17–e25. [Google Scholar] [CrossRef]
- WHO. Madagascar Plague Outbreak: External Situation; Report #14; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- Bragazzi, N.L.; Mahroum, N. Google trends predicts present and future plague cases during the plague outbreak in Madagascar: Infodemiological study. JMIR Public Health Surveill 2019, 5, e13142. [Google Scholar] [CrossRef] [Green Version]
- Kilonzo, B.S.; Mvena, Z.S.; Machangu, R.S.; Mbise, T.J. Preliminary observations on factors responsible for long persistence and continued outbreaks of plague in Lushoto district, Tanzania. Acta Trop. 1997, 68, 215–227. [Google Scholar] [CrossRef]
- Galimand, M.; Guiyoule, A.; Gerbaud, G.; Rasoamanana, B.; Chanteau, S.; Carniel, E.; Courvalin, P. Multidrug resistance in Yersinia pestis mediated by a transferable plasmid. N. Engl. J. Med. 1997, 337, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Godfred-Cato, S.; Cooley, K.M.; Fleck-Derderian, S.; Becksted, H.A.; Russell, Z.; Meaney-Delman, D.; Mead, P.S.; Nelson, C.A. Treatment of human plague: A systematic review of published aggregate data on antimicrobial efficacy, 1939–2019. Clin. Infect. Dis. 2020, 70 (Suppl. S1), S11–S19. [Google Scholar] [CrossRef] [PubMed]
- Kugeler, K.J.; Mead, P.S.; Campbell, S.B.; Nelson, C.A. Antimicrobial treatment patterns and illness outcome among United States patients with plague, 1942–2018. Clin. Infect. Dis. 2020, 70 (Suppl. S1), S20–S26. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.A.; Fleck-Derderian, S.; Cooley, K.M.; Meaney-Delman, D.; Becksted, H.A.; Russell, Z.; Renaud Bertherat, E.; Mead, P.S. Antimicrobial treatment of human plague: A systematic review of the literature on individual cases, 1937–2019. Clin. Infect. Dis. 2020, 70 (Suppl. S1), S3–S10. [Google Scholar] [CrossRef]
- Oyston, P.C.; Williamson, E.D. Prophylaxis and therapy of plague. Expert Rev. Anti-Infect. Ther. 2013, 11, 817–829. [Google Scholar] [CrossRef]
- Dennis, D.T. Plague as a Biological Weapon. In Bioterrorism and Infectious Agents: A New Dilemma for the 21st Century; Fong, I.W., Alibek, K., Eds.; Springer New York: New York, NY, USA, 2009; pp. 37–70. [Google Scholar]
- Wendte, J.M.; Ponnusamy, D.; Reiber, D.; Blair, J.L.; Clinkenbeard, K.D. In vitro efficacy of antibiotics commonly used to treat human plague against intracellular Yersinia pestis. Antimicrob. Agents Chemother. 2011, 55, 3752–3757. [Google Scholar] [CrossRef] [Green Version]
- Apangu, T.; Griffith, K.; Abaru, J.; Candini, G.; Apio, H.; Okoth, F.; Okello, R.; Kaggwa, J.; Acayo, S.; Ezama, G.; et al. Successful treatment of human plague with oral ciprofloxacin. Emerg. Infect. Dis. 2017, 23, 553–555. [Google Scholar] [CrossRef]
- Welch, T.J.; Fricke, W.F.; McDermott, P.F.; White, D.G.; Rosso, M.L.; Rasko, D.A.; Mammel, M.K.; Eppinger, M.; Rosovitz, M.L.; Wagner, D.; et al. Multiple antimicrobial resistance in plague: An emerging public health risk. PLoS ONE 2007, 2, e309. [Google Scholar] [CrossRef] [Green Version]
- Galimand, M.; Carniel, E.; Courvalin, P. Resistance of Yersinia pestis to antimicrobial agents. Antimicrob. Agents Chemother. 2006, 50, 3233–3236. [Google Scholar] [CrossRef] [Green Version]
- Guiyoule, A.; Gerbaud, G.; Buchrieser, C.; Galimand, M.; Rahalison, L.; Chanteau, S.; Couvalin, P.; Carniel, E. Transferable plasmid-mediated resistance to streptomycin in a clinical isolate of Yersinia pestis. Emerg. Infect. Dis. 2001, 7, 43–48. [Google Scholar] [CrossRef] [Green Version]
- Popowska, M.; Krawczyk-Balska, A. Broad-host-range IncP-1 plasmids and their resistance potential. Front. Microbiol. 2013, 4, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabanel, N.; Bouchier, C.; Rajerison, M.; Carniel, E. Plasmid-mediated doxycycline resistance in a Yersinia pestis strain isolated from a rat. Int. J. Antimicrob. Agents 2018, 51, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Dai, R.; He, J.; Zha, X.; Wang, Y.; Zhang, X.; Gao, H.; Yang, X.; Li, J.; Xin, Y.; Wang, Y.; et al. A novel mechanism of streptomycin resistance in Yersinia pestis: Mutation in the rpsL gene. PLoS Negl. Trop. Dis. 2021, 15, e0009324. [Google Scholar] [CrossRef] [PubMed]
- Taitt, C.R.; Leski, T.A.; Chen, A.; Berk, K.L.; Dorsey, R.W.; Gregory, M.J.; Sozhamannan, S.; Frey, K.G.; Dutt, D.L.; Vora, G.J. A survey of antimicrobial resistance determinants in category a select agents, exempt strains, and near-neighbor species. Int. J. Mol. Sci. 2020, 21, 1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smiley, S.T. Current challenges in the development of vaccines for pneumonic plague. Expert Rev. Vaccinines 2008, 7, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Titball, R.W.; Williamson, E.D. Vaccination against bubonic and pneumonic plague. Vaccine 2001, 19, 4175–4184. [Google Scholar] [CrossRef]
- Sun, W.; Singh, A.K. Plague vaccine: Recent progress and prospects. NPJ Vaccines 2019, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavanaugh, D.C. K.F. Meyer’s work on plague. J. Infect. Dis. 1974, 129 (Suppl. S1), S10–S12. [Google Scholar] [CrossRef]
- Meyer, K.F.; Smith, G.; Foster, L.E.; Marshall, J.D., Jr.; Cavanaugh, D.C. Plague immunization. IV. Clinical reactions and serologic response to inoculations of Haffkine and freeze-dried plague vaccine. J. Infect. Dis. 1974, 129, S30–S36. [Google Scholar] [CrossRef]
- Bartelloni, P.J.; Marshall, J.D., Jr.; Cavanaugh, D.C. Clinical and serological responses to plague vaccine U.S.P. Mil. Med. 1973, 138, 720–722. [Google Scholar] [CrossRef]
- Cavanaugh, D.C.; Elisberg, B.L.; Llewellyn, C.H.; Marshall, J.D., Jr.; Rust, J.H., Jr.; Williams, J.E.; Meyer, K.F. Plague immunization. V. Indirect evidence for the efficacy of plague vaccine. J. Infect. Dis. 1974, 129 (Suppl. S1), S37–S40. [Google Scholar] [CrossRef]
- Meyer, K.F.; Smith, G.; Foster, L.; Brookman, M.; Sung, M. Live, attenuated Yersinia pestis vaccine: Virulent in nonhuman primates, harmless to guinea pigs. J. Infect. Dis. 1974, 129, S85–S112. [Google Scholar] [CrossRef]
- Titball, R.W.; Williamson, E.D. Yersinia pestis (plague) vaccines. Expert Opin. Biol. Ther. 2004, 4, 965–973. [Google Scholar] [CrossRef]
- Marshall, J.D., Jr.; Bartelloni, P.J.; Cavanaugh, D.C.; Kadull, P.J.; Meyer, K.F. Plague immunization. II. Relation of adverse clinical reactions to multiple immunizations with killed vaccine. J. Infect. Dis. 1974, 129, S19–S25. [Google Scholar] [CrossRef]
- Russell, P.; Eley, S.M.; Hibbs, S.E.; Manchee, R.J.; Stagg, A.J.; Titball, R.W. A comparison of Plague vaccine, USP and EV76 vaccine induced protection against Yersinia pestis in a murine model. Vaccine 1995, 13, 1551–1556. [Google Scholar] [CrossRef]
- Andrews, G.P.; Health, D.G.; Anderson, G.W.; Welkos, S.L.; Friedlander, A.M. Fraction 1 capsular antigen (F1) purification from Yersinia pestis CO92 and from an Escherichia coli recombinant strain and efficacy against lethal plague challenge. Infect. Immun. 1996, 64, 2180–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, H.; Packman, L.P. A filtered non-toxic plague vaccine which protects guinea-pigs and mice. Br. J. Exp. Pathol. 1966, 47, 25–34. [Google Scholar] [PubMed]
- Pitt, L.M. Non-human primates as a model for pneumonic plague: Animal models and correlates of protection for plague. In Proceedings of the Plague Vaccines Workshop, Gaithersburg, MD, USA, 13–14 October 2004. [Google Scholar]
- Branger, C.G.; Torres-Escobar, A.; Sun, W.; Perry, R.; Fetherston, J.D.; Roland, K.; Curtiss, R. Oral vaccination with LcrV from Yersinia pestis KIM delivered by live attenuated Salmonella enterica serovar Typhimurium elicits a protective immune response against challenge with Yersinia pseudotuberculosis and Yersinia enterocolitica. Vaccine 2009, 27, 5363–5370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branger, C.G.; Sun, W.; Torres-Escobar, A.; Perry, R.; Roland, K.L.; Fetherston, J.; Curtiss, R. Evaluation of Psn, HmuR and a modified LcrV protein delivered to mice by live attenuated Salmonella as a vaccine against bubonic and pneumonic Yersinia pestis challenge. Vaccine 2010, 29, 274–282. [Google Scholar] [CrossRef] [Green Version]
- Branger, C.G.; Fetherston, J.D.; Perry, R.D.; Curtiss, R. Oral vaccination with different antigens from Yersinia pestis KIM delivered by live attenuated Salmonella typhimurium elicits a protective immune response against plague. Adv. Exp. Med. Biol. 2007, 603, 387–399. [Google Scholar]
- Girard, G.; Robic, J. Current status of the plague in Madagascar and vaccinal prophylaxis with the aid of the EV virus-vaccine. Bull. Soc. Path. Exot. 1942, 35, 43–49. [Google Scholar]
- Feodorov, V.A.; Lyapina, A.M.; Ulianova, O.V.; Lyapina, E.P.; Sayapina, L.V.; Lyapin, M.N.; Shcherbakov, A.A.; Telepnev, M.V.; Motin, V.L. Serologic markers for long-term immunity in humans vaccinated with live Yersinia pestis EV NIIEG. Procedia Vaccinol. 2012, 6, 10–13. [Google Scholar] [CrossRef] [Green Version]
- Meyer, K.F.; Cavanaugh, D.C.; Bartelloni, P.J.; Marshall, J.D., Jr. Plague immunization. I. Past and present trends. J. Infect. Dis. 1974, 129, S13–S18. [Google Scholar] [CrossRef]
- Feodorova, V.A.; Sayapina, L.V.; Corbel, M.J.; Motin, V.L. Russian vaccines against especially dangerous bacterial pathogens. Emerg. Microbes Infect. 2014, 3, e86. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, X.; Zhou, D.; Yang, R. Live-attenuated Yersinia pestis vaccines. Expert Rev. Vaccines 2013, 12, 677–686. [Google Scholar] [CrossRef]
- Sun, W.; Roland, K.L.; Curtiss, R. Developing live vaccines against plague. J. Infect. Dev. Ctries. 2011, 5, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Curtiss, R. Rational considerations about development of live attenuated Yersinia pestis vaccines. Curr. Pharm. Biotechnol. 2013, 14, 878–886. [Google Scholar] [CrossRef] [PubMed]
- Bubeck, S.S.; Dube, P.H. Yersinia pestis CO92 delta yopH is a potent live, attenuated plague vaccine. Clin. Vaccine Immunol. 2007, 14, 1235–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozue, J.; Cote, C.K.; Webster, W.; Bassett, A.; Tobery, S.; Little, S.; Swietnicki, W. A Yersinia pestis YscN ATPase mutant functions as a live attenuated vaccine against bubonic plague in mice. FEMS Microbiol. Lett. 2012, 332, 113–121. [Google Scholar] [CrossRef]
- Culbreth, M.J.; Biryukov, S.S.; Shoe, J.L.; Dankmeyer, J.L.; Hunter, M.; Klimko, C.P.; Rosario-Acevedo, R.; Fetterer, D.P.; Moreau, A.M.; Welkos, S.L.; et al. The use of analgesics during vaccination with a live attenuated Yersinia pestis vaccine alters the resulting immune response in mice. Vaccines 2019, 7, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cote, C.K.; Biryukov, S.S.; Klimko, C.P.; Shoe, J.L.; Hunter, M.; Rosario-Acevedo, R.; Fetterer, D.P.; Moody, K.L.; Meyer, J.R.; Rill, N.O.; et al. Protection elicited by attenuated Live Yersinia pestis vaccine strains against lethal infection with virulent, Y. pestis. Vaccines 2021, 9, 161. [Google Scholar] [CrossRef]
- Jenkins, A.L.; Worsham, P.L.; Welkos, S.L. A strategy to verify the absence of the pgm locus in Yersinia pestis strain candidates for select agent exemption. J. Microbiol. Methods 2009, 77, 316–319. [Google Scholar] [CrossRef]
- Welkos, S.; Pitt, M.L.; Martinez, M.; Friedlander, A.; Vogel, P.; Tammariello, R. Determination of the virulence of the pigmentation-deficient and pigmentation-/plasminogen activator-deficient strains of Yersinia pestis in non-human primate and mouse models of pneumonic plague. Vaccine 2002, 20, 2206–2214. [Google Scholar] [CrossRef]
- Demeure, C.E.; Derbise, A.; Carniel, E. Oral vaccination against plague using Yersinia pseudotuberculosis. Chem. Biol. Interact. 2017, 267, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Curtiss, R., 3rd; Sun, W. A recombinant attenuated Yersinia pseudotuberculosis vaccine delivering a Y. pestis YopE(Nt138)-LcrV fusion elicits broad protection against plague and yersiniosis in mice. Infect. Immun. 2019, 87, e00296-19. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Sanapala, S.; Rahav, H.; Curtiss, R., 3rd. Oral administration of a recombinant attenuated Yersinia pseudotuberculosis strain elicits protective immunity against plague. Vaccine 2015, 33, 6727–6735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Sanapala, S.; Henderson, J.C.; Sam, S.; Olinzock, J.; Trent, M.S.; Curtiss, R., 3rd. LcrV delivered via type III secretion system of live attenuated Yersinia pseudotuberculosis enhances immunogenicity against pneumonic plague. Infect. Immun. 2014, 82, 4390–4404. [Google Scholar] [CrossRef] [Green Version]
- Hervé, C.; Laupèze, B.; Del Giudice, G.; Didierlaurent, A.M.; Tavares Da Silva, F. The how’s and what’s of vaccine reactogenicity. NPJ Vaccines 2019, 4, 39. [Google Scholar] [CrossRef] [Green Version]
- Williamson, E.D.; Eley, S.M.; Stagg, A.J.; Green, M.; Russell, P.; Titball, R.W. A sub-unit vaccine elicits IgG in serum, spleen cell cultures and bronchial washings and protects immunized animals against pneumonic plague. Vaccine 1997, 15, 1079–1084. [Google Scholar] [CrossRef]
- Jones, S.M.; Day, F.; Stagg, A.J.; Williamson, E.D. Protection conferred by a fully recombinant sub-unit vaccine against Yersinia pestis in male and female mice of four inbred strains. Vaccine 2000, 19, 358–366. [Google Scholar] [CrossRef]
- Al-Jawdah, A.D.; Ivanova, I.G.; Waller, H.; Perkins, N.D.; Lakey, J.H.; Peters, D.T. Induction of the immunoprotective coat of Yersinia pestis at body temperature is mediated by the Caf1R transcription factor. BMC Microbiol. 2019, 19, 68. [Google Scholar] [CrossRef]
- Galyov, E.E.; Smirnov, O.Y.; Karlishev, A.V.; Volkovoy, K.I.; Denesyuk, A.I.; Nazimov, I.V.; Rubtsov, K.S.; Abramov, V.M.; Dalvadyanz, S.M.; Zavyalov, V.P. Nucleotide sequence of the Yersinia pestis gene encoding F1 antigen and the primary structure of the protein. FEBS Lett. 1990, 277, 230–232. [Google Scholar] [CrossRef] [Green Version]
- Karlyshev, A.V.; Galyov, E.E.; Smirnov, O.Y.; Guzayev, A.P.; Abramov, V.M.; Zavyalov, V.P. A new gene of the ƒ1 operon of Y. pestis involved in the capsule biogenesis. FEBS Lett. 1992, 297, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Donavan, J.E.; Ham, D.; Fukui, G.M.; Surgalla, M.J. Role of the capsule of Pasteurella Pestis in bubonic plague in the guinea pig. J. Infect. Dis. 1961, 109, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Vorontsov, E.D.; Dubichev, A.G.; Serdobintsev, L.N.; Naumov, A.V. Association-dissociation processes and supermolecular organisation of the capsule antigen (protein F1) of Yersinia pestis. Biomed. Sci. 1990, 1, 391–396. [Google Scholar] [PubMed]
- Burrows, T.W.; Bacon, G.A. The effects of loss of different virulence determinants on the virulence and immunogenicity of strains of Pasteurella pestis. Br. J. Exp. Pathol. 1958, 39, 278–291. [Google Scholar] [PubMed]
- Anderson, G.W., Jr.; Leary, S.E.; Williamson, E.D.; Titball, R.; Welkos, S.L.; Wosham, P.L.; Friendlander, A.M. Recombinant V antigen protects mice against pneumonic and bubonic plague caused by F1-capsule-positive and -negative strains of Yersinia pestis. Infect. Immun. 1996, 64, 4580–4585. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.; Leary, S.E.; Griffin, K.F.; Williamson, E.D.; Titball, R.W. Regions of Yersinia pestis V antigen that contribute to protection against plague identified by passive and active immunization. Infect. Immun. 1997, 65, 4476–4482. [Google Scholar] [CrossRef] [Green Version]
- Leary, S.E.; Williamson, E.D.; Griffin, K.F.; Russell, P.; Eley, R.W.; Titball, R.W. Active immunization with recombinant V antigen from Yersinia pestis protects mice against plague. Infect. Immun. 1995, 63, 2854–2858. [Google Scholar] [CrossRef] [Green Version]
- Weeks, S.; Hill, J.; Friendlander, A.; Welkos, S. Anti-V antigen antibody protects macrophages from Yersinia pestis -induced cell death and promotes phagocytosis. Microb. Pathog. 2002, 32, 227–237. [Google Scholar] [CrossRef]
- Pettersson, J.; Holmström, A.; Hill, J.; Leary, S.; Frithz-Lindsten, E.; von Euler-Matell, A.; Carlsson, E.; Titball, R.; Forsberg, A.; Wolf-Watz, H. The V-antigen of Yersinia is surface exposed before target cell contact and involved in virulence protein translocation. Mol. Microbiol. 1999, 32, 961–976. [Google Scholar] [CrossRef]
- Sawa, T.; Yahr, T.L.; Ohara, M.; Kurahashi, K.; Gropper, M.A.; Wiener-Kronish, J.P.; Frank, D.W. Active and passive immunization with the Pseudomonas V antigen protects against type III intoxication and lung injury. Nat. Med. 1999, 5, 392–398. [Google Scholar] [CrossRef]
- Anisimov, A.P.; Dentovskaya, A.V.; Panfertsev, E.A.; Svetoch, T.E.; Kopylov, P.K.; Segelke, B.W.; Zemla, A.; Telepnev, M.V.; Motin, V.L. Amino acid and structural variability of Yersinia pestis LcrV protein. Infect. Genet. Evol. 2010, 10, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, W.J.; Thomas, R.E.; Schwan, T.G. Recombinant capsular antigen (fraction 1) from Yersinia pestis induces a protective antibody response in BALB/c mice. Am. J. Trop. Med. Hyg. 1990, 43, 389–396. [Google Scholar] [CrossRef] [PubMed]
- Carr, S.; Miller, J.; Leary, S.E.; Bennett, A.M.; Ho, A.; Williamson, E.D. Expression of a recombinant form of the V antigen of Yersinia pestis, using three different expression systems. Vaccine 1999, 18, 153–159. [Google Scholar] [CrossRef]
- Chichester, J.A.; Musiychuk, K.; Farrance, C.E.; Mett, V.; Lyons, J.; Mett, V.; Yusibov, V. A single component two-valent LcrV-F1 vaccine protects non-human primates against pneumonic plague. Vaccine 2009, 27, 3471–3474. [Google Scholar] [CrossRef] [PubMed]
- Williamson, E.D.; Vesey, P.M.; Gillhespy, K.J.; Eley, S.M.; Green, M.; Titball, R.W. An IgG1 titre to the F1 and V antigens correlates with protection against plague in the mouse model. Clin. Exp. Immunol. 1999, 116, 107–114. [Google Scholar] [CrossRef]
- Powell, B.S.; Andrews, G.P.; Enama, J.T.; Jenderk, S.; Bolt, C.; Worsham, P.; Pullen, J.K.; Ribot, W.; Hines, H.; Smith, L.; et al. Design and testing for a nontagged F1-V fusion protein as vaccine antigen against bubonic and pneumonic plague. Biotechnol. Prog. 2005, 21, 1490–1510. [Google Scholar] [CrossRef] [Green Version]
- Goodin, J.L.; Nellis, D.F.; Powell, B.S.; Vyas, V.V.; Enama, J.T.; Wang, L.C.; Clark, P.K.; Giardina, S.L.; Adamovicz, J.J.; Michiel, D.F. Purification and protective efficacy of monomeric and modified Yersinia pestis capsular F1-V antigen fusion proteins for vaccination against plague. Protein Exp. Purif. 2007, 53, 63–79. [Google Scholar] [CrossRef] [Green Version]
- Fellows, P.; Adamovicz, J.; Hartings, J.; Sherwood, R.; Mega, W.; Brasel, T.; Barr, E.; Holland, L.; Lin, W.; Rom, A.; et al. Protection in mice passively immunized with serum from cynomolgus macaques and humans vaccinated with recombinant plague vaccine (rF1V). Vaccine 2010, 28, 7748–7756. [Google Scholar] [CrossRef]
- Glynn, A.; Roy, C.J.; Powell, B.S.; Adamovicz, J.J.; Freytag, L.C.; Clements, J.D. Protection against aerosolized Yersinia pestis challenge following homologous and heterologous prime-boost with recombinant plague antigens. Infect. Immun. 2005, 73, 5256–5261. [Google Scholar] [CrossRef] [Green Version]
- Heath, D.G.; Anderson, G.W.; Mauro, J.M.; Welkos, S.L.; Andrews, G.P.; Adamovicz, J.; Friedlander, A.M. Protection against experimental bubonic and pneumonic plague by a recombinant capsular F1-V antigen fusion protein vaccine. Vaccine 1998, 16, 1131–1137. [Google Scholar] [CrossRef]
- Williamson, E.D.; Eley, S.M.; Stagg, A.J.; Green, M.; Russell, P.; Titball, R.W. A single dose sub-unit vaccine protects against pneumonic plague. Vaccine 2000, 19, 566–571. [Google Scholar] [CrossRef]
- Williamson, E.D.; Eley, S.M.; Griffin, K.F.; Green, M.; Russell, P.; Leary, S.E.; Oyston, P.C.; Easterbrokk, T.; Reddin, K.M.; Robinson, A. A new improved sub-unit vaccine for plague: The basis of protection. FEMS Immunol. Med. Microbiol. 1995, 12, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Elvin, S.J.; Williamson, E.D. The F1 and V subunit vaccine protects against plague in the absence of IL-4 driven immune responses. Microb. Pathog. 2000, 29, 223–230. [Google Scholar] [CrossRef]
- Sun, W. Plague Vaccines: Status and Future. Adv. Exp. Med. Biol. 2016, 918, 313–360. [Google Scholar]
- Quenee, L.E.; Schneewind, O. Plague vaccines and the molecular basis of immunity against Yersinia pestis. Hum. Vaccine 2009, 5, 817–823. [Google Scholar] [CrossRef]
- Quenee, L.E.; Ciletti, N.A.; Elli, D.; Hermanas, T.M.; Schneewind, O. Prevention of pneumonic plague in mice, rats, guinea pigs and non-human primates with clinical grade rV10, rV10-2 or F1-V vaccines. Vaccine 2011, 29, 6572–6583. [Google Scholar] [CrossRef] [Green Version]
- Williamson, E.D.; Packer, P.J.; Waters, E.L.; Simpson, D.D.; Hartings, J.; Twenhafel, N.; Pitt, M.L. Recombinant (F1+V) vaccine protects cynomolgus macaques against pneumonic plague. Vaccine 2011, 29, 4771–4777. [Google Scholar] [CrossRef]
- Amemiya, K.; Meyers, J.L.; Rogers, T.E.; Fast, R.L.; Bassett, A.D.; Worsham, P.L.; Powell, B.S.; Norris, S.L.; Krieg, A.M.; Adamovicz, J.J. CpG oligodeoxynucleotides augment the murine immune response to the Yersinia pestis F1-V vaccine in bubonic and pneumonic models of plague. Vaccine 2009, 27, 2220–2229. [Google Scholar] [CrossRef]
- D’Arco, C.; McCormick, A.A.; Arnaboldi, P.M. Single-dose intranasal subunit vaccine rapidly clears secondary sepsis in a high-dose pneumonic plague infection. Vaccine 2021, 39, 1435–1444. [Google Scholar] [CrossRef]
- Kilgore, P.B.; Sha, J.; Anderson, J.A.; Motin, V.L.; Chopra, A.K. A new generation needle- and adjuvant-free trivalent plague vaccine utilizing adenovirus-5 nanoparticle platform. NPJ Vaccines 2021, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Hamzabegovic, F.; Goll, J.B.; Hooper, W.F.; Frey, S.; Gelber, C.E.; Abate, G. Flagellin adjuvanted F1/V subunit plague vaccine induces T cell and functional antibody responses with unique gene signatures. NPJ Vaccines 2020, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Prete, G.; Santi, L.; Andrianaivoarimanana, V.; Amedei, A.; Domarle, O.; D’Elios, M.M.; Arntzen, C.J.; Rahalison, L.; Mason, H.S. Plant-derived recombinant F1, V, and F1-V fusion antigens of Yersinia pestis activate human cells of the innate and adaptive immune system. Int. J. Immunopathol. Pharmacol. 2009, 22, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.; Adamovicz, J.J.; Cyr, S.L.; Bolt, C.R.; Bellerose, N.; Pitt, L.M.; Lowell, G.H.; Burt, D.S. Intranasal Protollin/F1-V vaccine elicits respiratory and serum antibody responses and protects mice against lethal aerosolized plague infection. Vaccine 2006, 24, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Goodin, J.L.; Powell, B.S.; Enama, J.T.; Raab, R.W.; McKown, R.L.; Coffman, G.L.; Andrews, G.P. Purification and characterization of a recombinant Yersinia pestis V-F1 “Reversed” fusion protein for use as a new subunit vaccine against plague. Protein Exp. Purif. 2011, 76, 136–144. [Google Scholar] [CrossRef]
- Yamanaka, H.; Hoyt, T.; Yang, X.; Golden, S.; Bosio, C.M.; Crist, K.; Becker, T.; Maddaloni, M.; Pascual, D.W. A nasal interleukin-12 DNA vaccine coexpressing Yersinia pestis F1-V fusion protein confers protection against pneumonic plague. Infect. Immun. 2008, 76, 4564–4573. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.; Griffin, K.; Hodgson, I.; Williamson, E. Protective efficacy of a fully recombinant plague vaccine in the guinea pig. Vaccine 2003, 21, 3912–3918. [Google Scholar] [CrossRef]
- Williamson, E.D.; Flick-Smith, H.C.; Lebutt, C.; Rowland, C.A.; Jones, S.M.; Waters, E.L.; Gwyther, R.J.; Miller, J.; Packer, P.J.; Irving, M. Human immune response to a plague vaccine comprising recombinant F1 and V antigens. Infect. Immun. 2005, 73, 3598–3608. [Google Scholar] [CrossRef] [Green Version]
- Andrews, G.P.; Strachan, S.T.; Benner, G.E.; Sample, A.K.; Anderson, G.W., Jr.; Adamovicz, J.J.; Welkos, S.L.; Pullen, J.K.; Friedlander, A.M. Protective efficacy of recombinant Yersinia outer proteins against bubonic plague caused by encapsulated and nonencapsulated Yersinia pestis. Infect. Immun. 1999, 67, 1533–1537. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, M.I.; Noel, B.L.; Rampersaud, R.; Mena, P.; Benach, J.L.; Bliska, J.B. Vaccination of mice with a Yop translocon complex elicits antibodies that are protective against infection with F1- Yersinia pestis. Infect. Immun. 2008, 76, 5181–5190. [Google Scholar] [CrossRef] [Green Version]
- Motin, V.L.; Nakajima, R.; Smirnov, G.B.; Brubaker, R.R. Passive immunity to yersiniae mediated by anti-recombinant V antigen and protein A-V antigen fusion peptide. Infect. Immun. 1994, 62, 4192–4201. [Google Scholar] [CrossRef] [Green Version]
- Quenee, L.E.; Berube, B.J.; Segal, J.; Elli, D.; Ciletti, N.A.; Anderson, D.; Schneewind, O. Amino acid residues 196-226 of LcrV represent a plague protective epitope. Vaccine 2010, 28, 1870–1876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amemiya, K.; Dankmeyer, J.L.; Keasey, S.L.; Trevino, S.R.; Wormald, M.M.; Halasohoris, S.A.; Ribot, W.J.; Fetterer, D.P.; Cote, C.K.; Worsham, P.L.; et al. Binding sites of anti-LcrV monoclonal antibodies are more critical than the avidities and affinities for passive protection against Yersinia pestis infection in a bubonic plague model. Antibodies 2020, 9, 37. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.; Philipovskiy, A.V.; Wulff-Strobel, C.R.; Ye, Z.; Straley, S.C. Anti-LcrV antibody inhibits delivery of Yops by Yersinia pestis KIM5 by directly promoting phagocytosis. Infect. Immun. 2005, 73, 6127–6137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisele, N.A.; Anderson, D.M. Dual-function antibodies to Yersinia pestis LcrV required for pulmonary clearance of plague. Clin. Vaccine Immunol. 2009, 16, 1720–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sofer-Podesta, C.; Ang, J.; Hackett, N.R.; Senina, S.; Perlin, D.; Crystal, R.G.; Boyer, J.L. Adenovirus-mediated delivery of an anti-V antigen monoclonal antibody protects mice against a lethal Yersinia pestis challenge. Infect. Immun. 2009, 77, 1561–1568. [Google Scholar] [CrossRef] [Green Version]
- Miller, N.C.; Quenee, L.E.; Elli, D.; Ciletti, N.A.; Schneewind, O. Polymorphisms in the lcrV gene of Yersinia enterocolitica and their effect on plague protective immunity. Infect. Immun. 2012, 80, 1572–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roggenkamp, A.; Geiger, A.M.; Leitritz, L.; Kessler, A.; Heesemann, J. Passive immunity to infection with Yersinia spp. mediated by anti-recombinant V antigen is dependent on polymorphism of V antigen. Infect. Immun. 1997, 65, 446–451. [Google Scholar] [CrossRef] [Green Version]
- Daniel, C.; Dewitte, A.; Poiret, S.; Marceau, M.; Simonet, M.; Marceau, L.; Descombes, G.; Boutillier, D.; Bennaceur, N.; Bontemps-Gallo, S.; et al. Polymorphism in the yersinia LcrV antigen enables immune escape from the protection conferred by an LcrV-secreting Lactococcus lactis in a pseudotuberculosis mouse model. Front. Immunol. 2019, 10, 1830. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.W., Jr.; Worsham, P.L.; Bolt, C.R.; Andrews, G.P.; Welkos, S.L.; Friedlander, A.M. Protection of mice from fatal bubonic and pneumonic plague by passive immunization with monoclonal antibodies against the F1 protein of Yersinia pestis. Am. J. Trop. Med. Hyg. 1997, 56, 471–473. [Google Scholar] [CrossRef]
- Hill, J.; Copse, C.; Leary, S.; Stagg, A.J.; Williamson, E.D.; Titball, R.W. Synergistic protection of mice against plague with monoclonal antibodies specific for the F1 and V antigens of Yersinia pestis. Infect. Immun. 2003, 71, 2234–2238. [Google Scholar] [CrossRef] [Green Version]
- Hill, J.; Eyles, J.E.; Elvin, S.J.; Healey, G.D.; Lukaszewski, R.A.; Titball, R.W. Administration of antibody to the lung protects mice against pneumonic plague. Infect. Immun. 2006, 74, 3068–3070. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Zhu, Z.; Dankmeyer, J.L.; Wormald, M.M.; Fast, R.L.; Worsham, P.L.; Cote, C.K.; Amemiya, K.; Dimitrov, D.S. Human anti-plague monoclonal antibodies protect mice from Yersinia pestis in a bubonic plague model. PLoS ONE 2010, 5, e13047. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Ren, J.; Zhang, J.; Song, X.; Lui, S.; Chi, X.; Chen, Y.; Wem, Z.; Li, J.; Chen, W. Identification and characterization of a neutralizing monoclonal antibody that provides complete protection against Yersinia pestis. PLoS ONE 2017, 12, e0177012. [Google Scholar] [CrossRef]
- Lillo, A.M.; Velappan, N.; Kelliher, J.M.; Watts, A.J.; Merriman, S.P.; Vuyisich, G.; Lilley, L.M.; Coombs, K.E.; Mastren, T.; Teshima, M.; et al. Development of anti-Yersinia pestis human antibodies with features required for diagnostic and therapeutic applications. Immunotargets Ther. 2020, 9, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Navalkele, B.D.; Chopra, T. Bezlotoxumab: An emerging monoclonal antibody therapy for prevention of recurrent Clostridium difficile infection. Biologics 2018, 12, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, D.P.; Wilcox, M.H. Antibodies for treatment of Clostridium difficile infection. Clin. Vaccine Immunol. 2014, 21, 913–923. [Google Scholar] [CrossRef]
- Chen, Z.; Moayeri, M.; Purcell, R. Monoclonal antibody therapies against anthrax. Toxins 2011, 3, 1004–1019. [Google Scholar] [CrossRef] [Green Version]
- Pelfrene, E.; Mura, M.; Cavaleiro Sanches, A.; Cavaleri, M. Monoclonal antibodies as anti-infective products: A promising future? Clin. Microbiol. Infect. 2019, 25, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Schweizer, H.P. Mechanisms of antibiotic resistance in Burkholderia pseudomallei: Implications for treatment of melioidosis. Future Microbiol. 2012, 7, 1389–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarrad, A.M.; Karoli, T.; Blaskovich, M.A.; Lyras, D.; Cooper, M.A. Clostridium difficile drug pipeline: Challenges in discovery and development of new agents. J. Med. Chem. 2015, 58, 5164–5185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merriman, H. Chapter 13—Infectious diseases. In Acute Care Handbook for Physical Therapists, 4th ed.; Paz, J.C., West, M.P., Eds.; W.B. Saunders: St. Louis, MO, USA, 2014; pp. 313–334. [Google Scholar]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pachori, P.; Gothalwal, R.; Gandhi, P. Emergence of antibiotic resistance Pseudomonas aeruginosa in intensive care unit; a critical review. Genes Dis. 2019, 6, 109–119. [Google Scholar] [CrossRef] [PubMed]
- González-González, E.; Alvarez, M.M.; Márquez-Ipiña, A.R.; Trujillo-de Santiago, G.; Rodríguez-Martínez, L.M.; Annabi, N.; Khademhosseini, A. Anti-Ebola therapies based on monoclonal antibodies: Current state and challenges ahead. Crit. Rev. Biotechnol. 2017, 37, 53–68. [Google Scholar] [CrossRef]
- Bailey, M.J.; Broecker, F.; Freyn, A.W.; Choi, A.; Brown, J.A.; Federova, N.; Simon, V.; Lim, J.K.; Evans, M.J.; García-Sastre, A.; et al. Human monoclonal antibodies potently neutralize Zika virus and select for escape mutations on the lateral ridge of the envelope protein. J. Virol. 2019, 93, e00405-19. [Google Scholar] [CrossRef] [Green Version]
- Marovich, M.; Mascola, J.R.; Cohen, M.S. Monoclonal antibodies for prevention and treatment of COVID-19. JAMA 2020, 324, 131–132. [Google Scholar] [CrossRef] [PubMed]
- Jahanshahlu, L.; Rezaei, N. Monoclonal antibody as a potential anti-COVID-19. Biomed. Pharmacother. 2020, 129, 110337. [Google Scholar] [CrossRef]
- Lülf, S.; Matz, J.; Rouyez, M.C.; Järviluoma, A.; Saksela, K.; Benichou, S.; Geyer, M. Structural basis for the inhibition of HIV-1 Nef by a high-affinity binding single-domain antibody. Retrovirology 2014, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Teran, C.; Tiruthani, K.; McSweeney, M.; Ma, A.; Pickles, R.; Lai, S.K. Challenges and opportunities for antiviral monoclonal antibodies as COVID-19 therapy. Adv. Drug Deliv. Rev. 2021, 169, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Pecetta, S.; Finco, O.; Seubert, A. Quantum leap of monoclonal antibody (mAb) discovery and development in the COVID-19 era. Semin. Immunol. 2020, 50, 101427. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Antigen a | mAb Description | Antibody/Therapeutic Administration | Y. pestis Challenge Strain d | Challenge Dose (LD50) | Ref. | ||
|---|---|---|---|---|---|---|---|
| Route | Concentration | Schedule | |||||
| F1; LcrV; F1V + LcrV | human IgG1 | IP | 500 µg | 24 h pre–120 h post | CO92 | 25–40 | 202 |
| LcrV | mouse | IP | 350 µg | 24 h pre | GB | 12 | 156 |
| LcrV b | mouse IgG1, IgG2a | IP | 1–500 µg | 24 h pre | CO92 or C12 | 21–39 | 192 |
| LcrV; F1 + LcrV | mouse IgG1 | IP | 0.7–100 µg | 4 h pre–96 post | GB | 9.6–91,000 | 200 |
| LcrV | mouse IgG1 | IP | 200 µg | 1 h pre | CO92 | 20 | 191 |
| F1 c | mouse IgG1 | IP | 125–500 µg | 6 h or 24 h pre | CO92 | 48–54 | 199 |
| F1 | not reported | IV | 100 µg | 24 h pre | 141 | 600 | 203 |
| Antigen a | mAb Description | Antibody/Therapeutic Administration | Y. pestis Challenge Strain f | Challenge Dose (LD50) | Ref. | ||
|---|---|---|---|---|---|---|---|
| Route | Concentration | Schedule | |||||
| LcrV | mouse IgG1 | IP | 35 µg | 4 h pre–96 h post | GB | 88 | 156 |
| LcrV b | mouse IgG1 | IP | 200 or 400 µg | 1 h pre | CO92 g | 15–20 | 194 |
| LcrV c | mouse IgG2b | IP, IV | 100–500 µg, 1011 pu e | 94 h pre–24 h post | CO92 g | 363–9080 | 195 |
| F1 d | mouse IgG1 | IP | 125–500 µg | 6 h or 24 h pre | CO92 | 29–74 | 199 |
| F1 + LcrV | mouse IgG1 | IT | 77.5 µg (each) | 2 h post | GB | 27 | 200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosario-Acevedo, R.; Biryukov, S.S.; Bozue, J.A.; Cote, C.K. Plague Prevention and Therapy: Perspectives on Current and Future Strategies. Biomedicines 2021, 9, 1421. https://doi.org/10.3390/biomedicines9101421
Rosario-Acevedo R, Biryukov SS, Bozue JA, Cote CK. Plague Prevention and Therapy: Perspectives on Current and Future Strategies. Biomedicines. 2021; 9(10):1421. https://doi.org/10.3390/biomedicines9101421
Chicago/Turabian StyleRosario-Acevedo, Raysa, Sergei S. Biryukov, Joel A. Bozue, and Christopher K. Cote. 2021. "Plague Prevention and Therapy: Perspectives on Current and Future Strategies" Biomedicines 9, no. 10: 1421. https://doi.org/10.3390/biomedicines9101421
APA StyleRosario-Acevedo, R., Biryukov, S. S., Bozue, J. A., & Cote, C. K. (2021). Plague Prevention and Therapy: Perspectives on Current and Future Strategies. Biomedicines, 9(10), 1421. https://doi.org/10.3390/biomedicines9101421