
The Role of NKL Homeobox Genes in T-Cell Malignancies


Abstract
:1. T-Cell Development
2. NKL Homeobox Genes in Normal and Malignant T-Cells
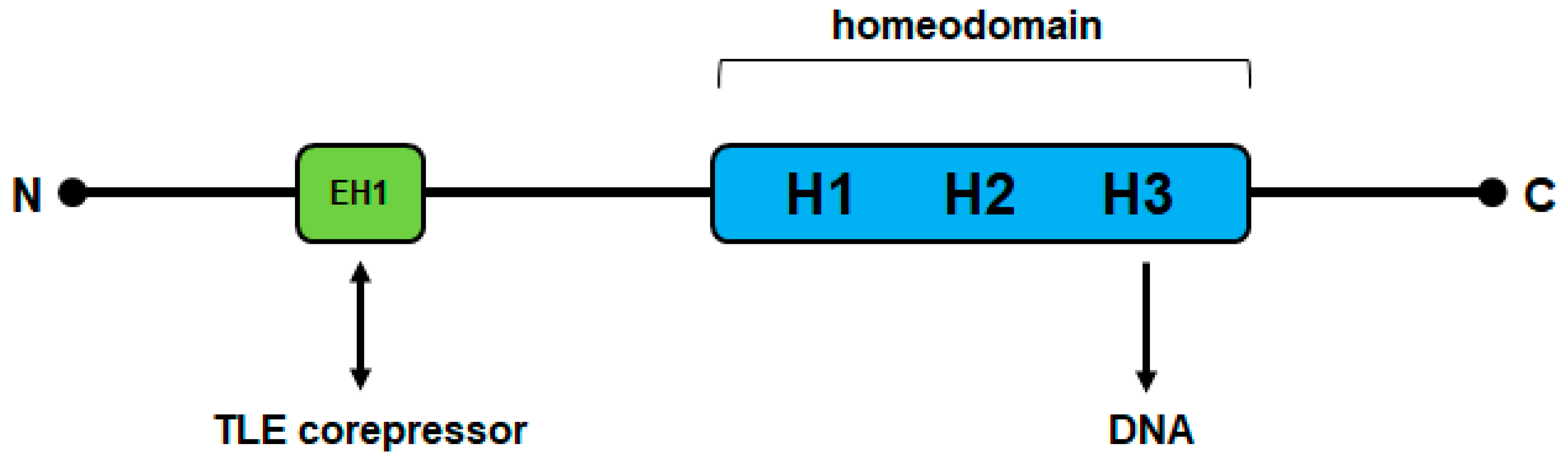
2.1. Classification of Homeobox Genes
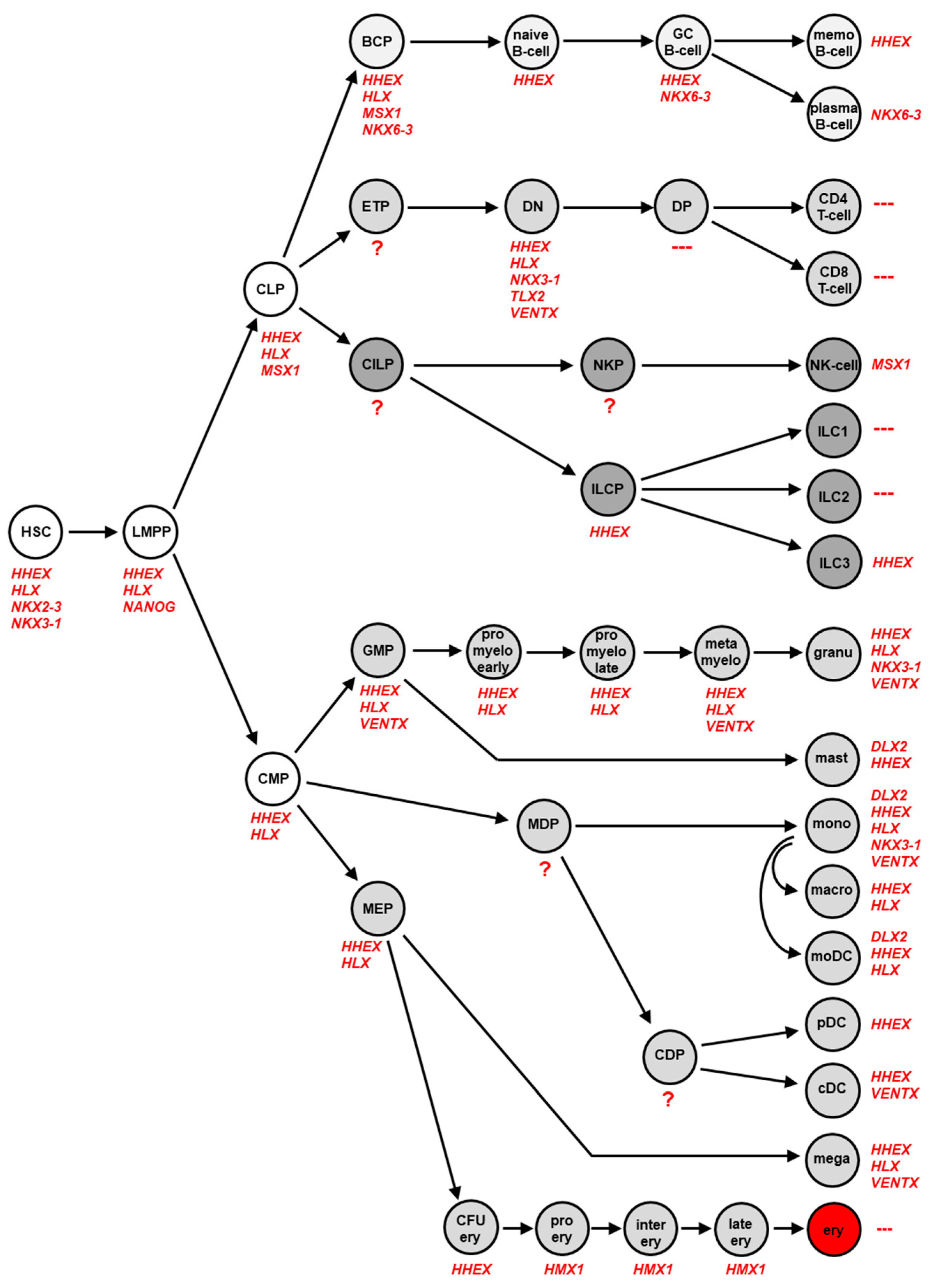
2.2. NKL-Code in Hematopoiesis
2.3. Deregulated NKL Homeobox Genes in T-Cell Acute Lymphoid Leukemia

{kind=link}
{kind=link}
| Gene | AITL | ALCL | ATLL | HSTL | NKTL | PTCL | T-ALL | T-Cell Line Models | References |
|---|---|---|---|---|---|---|---|---|---|
| BARHL1 | + | ||||||||
| BARHL2 | |||||||||
| BARX1 | |||||||||
| BARX2 | + | ||||||||
| BSX | |||||||||
| DBX1 | |||||||||
| DBX2 | |||||||||
| DLX1 | + | ||||||||
| DLX2 | + | HPB-ALL | [43] | ||||||
| DLX3 | + | ||||||||
| DLX4 | + | ||||||||
| DLX5 | |||||||||
| DLX6 | + | ||||||||
| EMX1 | |||||||||
| EMX2 | + | ||||||||
| EN1 | + | ||||||||
| EN2 | + | ||||||||
| HHEX | + | + | + | + | + | + | + | CCRF-CEM, RPMI-8402 | [43] |
| HLX | + | + | + | + | + | + | + | DEL, KI-JK, L-82, SR-786, SU-DHL-1, SUP-M2 | [21] |
| HMX1 | |||||||||
| HMX2 | + | ||||||||
| HMX3 | |||||||||
| LBX1 | + | ||||||||
| LBX2 | + | ||||||||
| MSX1 | + | + | + | + | + | + | DERL-2, DERL-7, LOUCY | [44,45] | |
| MSX2 | + | ||||||||
| NANOG | + | ||||||||
| NKX1-1 | + | + | + | + | |||||
| NKX1-2 | |||||||||
| NKX2-1 | + | + | + | + | + | RPMI-8402 | [43] | ||
| NKX2-2 | + | + | + | + | + | ||||
| NKX2-3 | + | + | + | + | + | + | + | ||
| NKX2-4 | |||||||||
| NKX2-5 | + | + | + | + | + | CCRF-CEM, PEER | [46] | ||
| NKX2-6 | |||||||||
| NKX2-8 | |||||||||
| NKX3-1 | + | + | JURKAT, MOLT-4, RPMI-8402 | [47] | |||||
| NKX3-2 | + | CCRF-CEM | [48] | ||||||
| NKX6-1 | + | + | |||||||
| NKX6-2 | |||||||||
| NKX6-3 | + | + | |||||||
| NOTO | |||||||||
| TLX1 | + | + | ALL-SIL | [49] | |||||
| TLX2 | + | ||||||||
| TLX3 | + | + | DND-41, HPB-ALL | [50] | |||||
| VAX1 | |||||||||
| VAX2 | + | ||||||||
| VENTX | |||||||||
| 48 | 7 | 9 | 6 | 6 | 11 | 11 | 25 |
2.3.1. TLX1
2.3.2. TLX3
2.3.3. NKX2-5
2.3.4. NKX2-1
2.3.5. NKX2-3
2.3.6. NKX2-4
2.3.7. NKX3-1
2.4. Deregulated NKL Homeobox Genes in T-Cell Lymphoma
2.4.1. DLX
2.4.2. HHEX
2.4.3. HLX
2.4.4. MSX1
2.4.5. NANOG
2.4.6. NKX2-2
2.5. Tumor Cell Lines as Models for Deregulated NKL Homeobox Genes
3. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rothenberg, E.V. Transcriptional control of early T and B cell developmental choices. Annu. Rev. Immunol. 2014, 32, 283–321. [Google Scholar] [CrossRef] [Green Version]
- Yui, M.A.; Rothenberg, E.V. Developmental gene networks: A triathlon on the course to T cell identity. Nat. Rev. Immunol. 2014, 14, 529–545. [Google Scholar] [CrossRef] [Green Version]
- Graux, C.; Cools, J.; Michaux, L.; Vandenberghe, P.; Hagemeijer, A. Cytogenetics and molecular genetics of T-cell acute lymphoblastic leukemia: From thymocyte to lymphoblast. Leukemia 2006, 20, 1496–1510. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, A.A.; Neuberg, D.S.; Staunton, J.; Loh, M.L.; Huard, C.; Raimondi, S.C.; Behm, F.G.; Pui, C.H.; Downing, J.R.; Gilliland, D.G.; et al. Gene expression signatures define novel oncogenic pathways in T cell acute lymphoblastic leukemia. Cancer Cell 2002, 1, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Vaquerizas, J.M.; Kummerfeld, S.K.; Teichmann, S.A.; Luscombe, N.M. A census of human transcription factors: Function, expression and evolution. Nat. Rev. Genet. 2009, 10, 252–263. [Google Scholar] [CrossRef]
- Bürglin, T.R.; Affolter, M. Homeodomain proteins: An update. Chromosoma 2016, 125, 497–521. [Google Scholar] [CrossRef] [Green Version]
- Gehring, W.J.; Müller, M.; Affolter, M.; Percival-Smith, A.; Billeter, M.; Qian, Y.Q.; Otting, G.; Wüthrich, K. The structure of the homeodomain and its functional implications. Trends Genet. 1990, 6, 323–329. [Google Scholar] [CrossRef]
- Holland, P.W.; Booth, H.A.; Bruford, E.A. Classification and nomenclature of all human homeobox genes. BMC Biol. 2007, 5, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Nirenberg, M. Drosophila NK-homeobox genes. Proc. Natl. Acad. Sci. USA 1989, 86, 7716–7720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollard, S.L.; Holland, P.W. Evidence for 14 homeobox gene clusters in human genome ancestry. Curr. Biol. 2000, 10, 1059–1062. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.P. NK-2 homeobox genes and heart development. Dev. Biol. 1996, 178, 203–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.T.; Jaynes, J.B. A conserved region of engrailed, shared among all en-, gsc-, Nk1-, Nk2- and msh-class homeoproteins, mediates active transcriptional repression in vivo. Development 1996, 122, 3141–3150. [Google Scholar] [CrossRef] [PubMed]
- Jennings, B.H.; Ish-Horowicz, D. The Groucho/TLE/Grg family of transcriptional co-repressors. Genome Biol. 2008, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Fernàndez, J. The genesis and evolution of homeobox gene clusters. Nat. Rev. Genet. 2005, 6, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Jagla, K.; Bellard, M.; Frasch, M. A cluster of Drosophila homeobox genes involved in mesoderm differentiation programs. Bioessays 2001, 23, 125–133. [Google Scholar] [CrossRef]
- Lints, T.J.; Parsons, L.M.; Hartley, L.; Lyons, I.; Harvey, R.P. Nkx-2.5: A novel murine homeobox gene expressed in early heart progenitor cells and their myogenic descendants. Development 1993, 119, 419–431. [Google Scholar] [CrossRef]
- Lazzaro, D.; Price, M.; de Felice, M.; Di Lauro, R. The transcription factor TTF-1 is expressed at the onset of thyroid and lung morphogenesis and in restricted regions of the foetal brain. Development 1991, 113, 1093–1104. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Pommerenke, C.; Scherr, M.; Meyer, C.; Kaufmann, M.; Battmer, K.; MacLeod, R.A.; Drexler, H.G. NKL homeobox gene activities in hematopoietic stem cells, T-cell development and T-cell leukemia. PLoS ONE 2017, 12, e0171164. [Google Scholar] [CrossRef]
- Nagel, S.; MacLeod, R.A.F.; Meyer, C.; Kaufmann, M.; Drexler, H.G. NKL homeobox gene activities in B-cell development and lymphomas. PLoS ONE 2018, 13, e0205537. [Google Scholar] [CrossRef]
- Nagel, S.; Pommerenke, C.; Meyer, C.; Kaufmann, M.; MacLeod, R.A.F.; Drexler, H.G. NKL homeobox gene MSX1 acts like a tumor suppressor in NK-cell leukemia. Oncotarget 2017, 8, 66815–66832. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.; Pommerenke, C.; MacLeod, R.A.F.; Meyer, C.; Kaufmann, M.; Drexler, H.G. The NKL-code for innate lymphoid cells reveals deregulated expression of NKL homeobox genes HHEX and HLX in anaplastic large cell lymphoma (ALCL). Oncotarget 2020, 11, 3208–3226. [Google Scholar] [CrossRef]
- Nagel, S.; Scherr, M.; MacLeod, R.A.F.; Pommerenke, C.; Koeppel, M.; Meyer, C.; Kaufmann, M.; Dallmann, I.; Drexler, H.G. NKL homeobox gene activities in normal and malignant myeloid cells. PLoS ONE 2019, 14, e0226212. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.; Pommerenke, C.; Meyer, C.; Drexler, H.G. NKL Homeobox gene VENTX is part of a regulatory network in human conventional dendritic cells. Int. J. Mol. Sci. 2021, 22, 5902. [Google Scholar] [CrossRef]
- Nagel, S. NKL-Code in Normal and Aberrant Hematopoiesis. Cancers 2021, 13, 1961. [Google Scholar] [CrossRef]
- Farlik, M.; Halbritter, F.; Müller, F.; Choudry, F.A.; Ebert, P.; Klughammer, J.; Farrow, S.; Santoro, A.; Ciaurro, V.; Mathur, A.; et al. DNA methylation dynamics of human hematopoietic stem cell differentiation. Cell Stem Cell 2016, 19, 808–822. [Google Scholar] [CrossRef] [Green Version]
- Allen, J.D.; Lints, T.; Jenkins, N.A.; Copeland, N.G.; Strasser, A.; Harvey, R.P.; Adams, J.M. Novel murine homeo box gene on chromosome 1 expressed in specific hematopoietic lineages and during embryogenesis. Genes Dev. 1991, 5, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Crompton, M.R.; Bartlett, T.J.; MacGregor, A.D.; Manfioletti, G.; Buratti, E.; Giancotti, V.; Goodwin, G.H. Identification of a novel vertebrate homeobox gene expressed in haematopoietic cells. Nucleic Acids Res. 1992, 20, 5661–5667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedford, F.K.; Ashworth, A.; Enver, T.; Wiedemann, L.M. HEX: A novel homeobox gene expressed during haematopoiesis and conserved between mouse and human. Nucleic Acids Res. 1993, 21, 1245–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manfioletti, G.; Gattei, V.; Buratti, E.; Rustighi, A.; De Iuliis, A.; Aldinucci, D.; Goodwin, G.H.; Pinto, A. Differential expression of a novel proline-rich homeobox gene (Prh) in human hematolymphopoietic cells. Blood 1995, 85, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Mack, D.L.; Leibowitz, D.S.; Cooper, S.; Ramsey, H.; Broxmeyer, H.E.; Hromas, R. Down-regulation of the myeloid homeobox protein Hex is essential for normal T-cell development. Immunology 2002, 107, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Jackson, J.T.; Nasa, C.; Shi, W.; Huntington, N.D.; Bogue, C.W.; Alexander, W.S.; McCormack, M.P. A crucial role for the homeodomain transcription factor Hhex in lymphopoiesis. Blood 2015, 125, 803–814. [Google Scholar] [CrossRef] [Green Version]
- Laidlaw, B.J.; Duan, L.; Xu, Y.; Vazquez, S.E.; Cyster, J.G. The transcription factor Hhex cooperates with the corepressor Tle3 to promote memory B cell development. Nat. Immunol. 2020, 21, 1082–1093. [Google Scholar] [CrossRef]
- Allen, J.D.; Harris, A.W.; Bath, M.L.; Strasser, A.; Scollay, R.; Adams, J.M. Perturbed development of T and B cells in mice expressing an Hlx homeobox transgene. J. Immunol. 1995, 154, 1531–1542. [Google Scholar] [PubMed]
- Migueles, R.P.; Shaw, L.; Rodrigues, N.P.; May, G.; Henseleit, K.; Anderson, K.G.; Goker, H.; Jones, C.M.; de Bruijn, M.F.; Brickman, J.M.; et al. Transcriptional regulation of Hhex in hematopoiesis and hematopoietic stem cell ontogeny. Dev. Biol. 2017, 424, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.E. How do genes regulate simple behaviours? Understanding how different neurons in the vertebrate spinal cord are genetically specified. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 45–66. [Google Scholar] [CrossRef]
- Hunt, P.; Gulisano, M.; Cook, M.; Sham, M.H.; Faiella, A.; Wilkinson, D.; Boncinelli, E.; Krumlauf, R. A distinct Hox code for the branchial region of the vertebrate head. Nature 1991, 353, 861–864. [Google Scholar] [CrossRef]
- Depew, M.J.; Simpson, C.A.; Morasso, M.; Rubenstein, J.L. Reassessing the Dlx code: The genetic regulation of branchial arch skeletal pattern and development. J. Anat. 2005, 207, 501–561. [Google Scholar] [CrossRef] [PubMed]
- Streit, A. The cranial sensory nervous system: Specification of sensory progenitors and placodes. In StemBook; Harvard Stem Cell Institute: Cambridge, MA, USA, 2008. [Google Scholar] [PubMed]
- Nagel, S.; Pommerenke, C.; Meyer, C.; MacLeod, R.A.F.; Drexler, H.G. Establishment of the TALE-code reveals aberrantly activated homeobox gene PBX1 in Hodgkin lymphoma. PLoS ONE 2021, 16, e0246603. [Google Scholar] [CrossRef]
- O’Neil, J.; Look, A.T. Mechanisms of transcription factor deregulation in lymphoid cell transformation. Oncogene 2007, 26, 6838–6849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eswaran, J.; Sinclair, P.; Heidenreich, O.; Irving, J.; Russell, L.J.; Hall, A.; Calado, D.P.; Harrison, C.J.; Vormoor, J. The pre-B-cell receptor checkpoint in acute lymphoblastic leukaemia. Leukemia 2015, 29, 1623–1631. [Google Scholar] [CrossRef]
- Tomolonis, J.A.; Agarwal, S.; Shohet, J.M. Neuroblastoma pathogenesis: Deregulation of embryonic neural crest development. Cell Tissue Res. 2018, 372, 245–262. [Google Scholar] [CrossRef] [Green Version]
- Quentmeier, H.; Pommerenke, C.; Dirks, W.G.; Eberth, S.; Koeppel, M.; MacLeod, R.A.F.; Nagel, S.; Steube, K.; Uphoff, C.C.; Drexler, H.G. The LL-100 panel: 100 cell lines for blood cancer studies. Sci. Rep. 2019, 9, 8218. [Google Scholar] [CrossRef]
- Nagel, S.; Pommerenke, C.; MacLeod, R.A.F.; Meyer, C.; Kaufmann, M.; Fähnrich, S.; Drexler, H.G. Deregulated expression of NKL homeobox genes in T-cell lymphomas. Oncotarget 2019, 10, 3227–3247. [Google Scholar] [CrossRef]
- Nagel, S.; Ehrentraut, S.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Repressed BMP signaling reactivates NKL homeobox gene MSX1 in a T-ALL subset. Leuk. Lymphoma 2015, 56, 480–491. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. The cardiac homeobox gene NKX2-5 is deregulated by juxtaposition with BCL11B in pediatric T-ALL cell lines via a novel t(5;14)(q35.1;q32.2). Cancer Res. 2003, 63, 5329–5334. [Google Scholar] [PubMed]
- Nagel, S.; Ehrentraut, S.; Tomasch, J.; Lienenklaus, S.; Schneider, B.; Geffers, R.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Transcriptional activation of prostate specific homeobox gene NKX3-1 in subsets of T-cell lymphoblastic leukemia (T-ALL). PLoS ONE 2012, 7, e40747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, S.; Meyer, C.; Kaufmann, M.; Zaborski, M.; MacLeod, R.A.F.; Drexler, H.G. Aberrant activity of NKL homeobox gene NKX3-2 in a T-ALL subset. PLoS ONE 2018, 13, e0197194. [Google Scholar] [CrossRef] [PubMed]
- Hatano, M.; Roberts, C.W.; Minden, M.; Crist, W.M.; Korsmeyer, S.J. Deregulation of a homeobox gene, HOX11, by the t(10;14) in T cell leukemia. Science 1991, 253, 79–82. [Google Scholar] [CrossRef]
- MacLeod, R.A.; Nagel, S.; Kaufmann, M.; Janssen, J.W.; Drexler, H.G. Activation of HOX11L2 by juxtaposition with 3’-BCL11B in an acute lymphoblastic leukemia cell line (HPB-ALL) with t(5;14)(q35;q32.2). Genes Chromosomes Cancer 2003, 37, 84–91. [Google Scholar] [CrossRef]
- Nagel, S.; Pommerenke, C.; Meyer, C.; MacLeod, R.A.F.; Drexler, H.G. Aberrant expression of NKL homeobox genes HMX2 and HMX3 interferes with cell differentiation in acute myeloid leukemia. PLoS ONE 2020, 15, e0240120. [Google Scholar] [CrossRef]
- Brendolan, A.; Ferretti, E.; Salsi, V.; Moses, K.; Quaggin, S.; Blasi, F.; Cleary, M.L.; Selleri, L. A Pbx1-dependent genetic and transcriptional network regulates spleen ontogeny. Development 2005, 132, 3113–3126. [Google Scholar] [CrossRef] [Green Version]
- Bernard, O.A.; Busson-LeConiat, M.; Ballerini, P.; Mauchauffé, M.; Della Valle, V.; Monni, R.; Nguyen Khac, F.; Mercher, T.; Penard-Lacronique, V.; Pasturaud, P.; et al. A new recurrent and specific cryptic translocation, t(5;14)(q35;q32), is associated with expression of the Hox11L2 gene in T acute lymphoblastic leukemia. Leukemia 2001, 15, 1495–1504. [Google Scholar] [CrossRef] [Green Version]
- Meijerink, J.P. Genetic rearrangements in relation to immunophenotype and outcome in T-cell acute lymphoblastic leukaemia. Best Pract. Res. Clin. Haematol. 2010, 23, 307–318. [Google Scholar] [CrossRef]
- De Keersmaecker, K.; Real, P.J.; Gatta, G.D.; Palomero, T.; Sulis, M.L.; Tosello, V.; Van Vlierberghe, P.; Barnes, K.; Castillo, M.; Sole, X.; et al. The TLX1 oncogene drives aneuploidy in T cell transformation. Nat. Med. 2010, 16, 1321–1327. [Google Scholar] [CrossRef] [Green Version]
- Dadi, S.; Le Noir, S.; Payet-Bornet, D.; Lhermitte, L.; Zacarias-Cabeza, J.; Bergeron, J.; Villarèse, P.; Vachez, E.; Dik, W.A.; Millien, C.; et al. TLX homeodomain oncogenes mediate T cell maturation arrest in T-ALL via interaction with ETS1 and suppression of TCRα gene expression. Cancer Cell 2012, 21, 563–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villarese, P.; Lours, C.; Trinquand, A.; Le Noir, S.; Belhocine, M.; Lhermitte, L.; Cieslak, A.; Tesio, M.; Petit, A.; LeLorch, M.; et al. TCRα rearrangements identify a subgroup of NKL-deregulated adult T-ALLs associated with favorable outcome. Leukemia 2018, 32, 61–71. [Google Scholar] [CrossRef]
- Przybylski, G.K.; Dik, W.A.; Grabarczyk, P.; Wanzeck, J.; Chudobska, P.; Jankowski, K.; von Bergh, A.; van Dongen, J.J.; Schmidt, C.A.; Langerak, A.W. The effect of a novel recombination between the homeobox gene NKX2-5 and the TRD locus in T-cell acute lymphoblastic leukemia on activation of the NKX2-5 gene. Haematologica 2006, 91, 317–321. [Google Scholar]
- Skerjanc, I.S.; Petropoulos, H.; Ridgeway, A.G.; Wilton, S. Myocyte enhancer factor 2C and Nkx2-5 up-regulate each other’s expression and initiate cardiomyogenesis in P19 cells. J. Biol. Chem. 1998, 273, 34904–34910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, S.; Meyer, C.; Quentmeier, H.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. MEF2C is activated by multiple mechanisms in a subset of T-acute lymphoblastic leukemia cell lines. Leukemia 2008, 22, 600–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homminga, I.; Pieters, R.; Langerak, A.W.; de Rooi, J.J.; Stubbs, A.; Verstegen, M.; Vuerhard, M.; Buijs-Gladdines, J.; Kooi, C.; Klous, P.; et al. Integrated transcript and genome analyses reveal NKX2-1 and MEF2C as potential oncogenes in T cell acute lymphoblastic leukemia. Cancer Cell 2011, 19, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Venturini, L.; Meyer, C.; Kaufmann, M.; Scherr, M.; Drexler, H.G.; Macleod, R.A. Transcriptional deregulation of oncogenic myocyte enhancer factor 2C in T-cell acute lymphoblastic leukemia. Leuk. Lymphoma 2011, 52, 290–297. [Google Scholar] [CrossRef]
- Nagel, S.; Ehrentraut, S.; Tomasch, J.; Quentmeier, H.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Ectopic expression of homeobox gene NKX2-1 in diffuse large B-cell lymphoma is mediated by aberrant chromatin modifications. PLoS ONE 2013, 8, e61447. [Google Scholar] [CrossRef]
- Le Noir, S.; Ben Abdelali, R.; Lelorch, M.; Bergeron, J.; Sungalee, S.; Payet-Bornet, D.; Villarèse, P.; Petit, A.; Callens, C.; Lhermitte, L.; et al. Extensive molecular mapping of TCRα/δ- and TCRβ-involved chromosomal translocations reveals distinct mechanisms of oncogene activation in T-ALL. Blood 2012, 120, 3298–3309. [Google Scholar] [CrossRef] [Green Version]
- Small, E.M.; Vokes, S.A.; Garriock, R.J.; Li, D.; Krieg, P.A. Developmental expression of the Xenopus Nkx2-1 and Nkx2-4 genes. Mech. Dev. 2000, 96, 259–262. [Google Scholar] [CrossRef]
- Manoli, M.; Driever, W. nkx2.1 and nkx2.4 genes function partially redundant during development of the zebrafish hypothalamus, preoptic region, and pallidum. Front. Neuroanat. 2014, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Kusy, S.; Gerby, B.; Goardon, N.; Gault, N.; Ferri, F.; Gérard, D.; Armstrong, F.; Ballerini, P.; Cayuela, J.M.; Baruchel, A.; et al. NKX3.1 is a direct TAL1 target gene that mediates proliferation of TAL1-expressing human T cell acute lymphoblastic leukemia. J. Exp. Med. 2010, 207, 2141–2156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soulier, J.; Clappier, E.; Cayuela, J.M.; Regnault, A.; García-Peydró, M.; Dombret, H.; Baruchel, A.; Toribio, M.L.; Sigaux, F. HOXA genes are included in genetic and biologic networks defining human acute T-cell leukemia (T-ALL). Blood 2005, 106, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Testa, J.R. DLX Genes: Roles in Development and Cancer. Cancers 2021, 13, 3005. [Google Scholar] [CrossRef]
- Sunwoo, J.B.; Kim, S.; Yang, L.; Naik, T.; Higuchi, D.A.; Rubenstein, J.L.; Yokoyama, W.M. Distal-less homeobox transcription factors regulate development and maturation of natural killer cells. Proc. Natl. Acad. Sci. USA 2008, 105, 10877–10882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starkova, J.; Gadgil, S.; Qiu, Y.H.; Zhang, N.; Hermanova, I.; Kornblau, S.M.; Drabkin, H.A. Up-regulation of homeodomain genes, DLX1 and DLX2, by FLT3 signaling. Haematologica 2011, 96, 820–828. [Google Scholar] [CrossRef]
- Tan, Y.; Timakhov, R.A.; Rao, M.; Altomare, D.A.; Xu, J.; Liu, Z.; Gao, Q.; Jhanwar, S.C.; Di Cristofano, A.; Wiest, D.L.; et al. A novel recurrent chromosomal inversion implicates the homeobox gene Dlx5 in T-cell lymphomas from Lck-Akt2 transgenic mice. Cancer Res. 2008, 68, 1296–1302. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Sementino, E.; Xu, J.; Pei, J.; Liu, Z.; Ito, T.K.; Cai, K.Q.; Peri, S.; Klein-Szanto, A.J.; Wiest, D.L.; et al. The homeoprotein Dlx5 drives murine T-cell lymphomagenesis by directly transactivating Notch and upregulating Akt signaling. Oncotarget 2017, 8, 14941–14956. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Sementino, E.; Liu, Z.; Cai, K.Q.; Testa, J.R. Wnt signaling mediates oncogenic synergy between Akt and Dlx5 in T-cell lymphomagenesis by enhancing cholesterol synthesis. Sci. Rep. 2020, 10, 15837. [Google Scholar] [CrossRef]
- Schleussner, N.; Merkel, O.; Costanza, M.; Liang, H.C.; Hummel, F.; Romagnani, C.; Durek, P.; Anagnostopoulos, I.; Hummel, M.; Jöhrens, K.; et al. The AP-1-BATF and -BATF3 module is essential for growth, survival and TH17/ILC3 skewing of anaplastic large cell lymphoma. Leukemia 2018, 32, 1994–2007. [Google Scholar] [CrossRef]
- Crescenzo, R.; Abate, F.; Lasorsa, E.; Tabbo’, F.; Gaudiano, M.; Chiesa, N.; Di Giacomo, F.; Spaccarotella, E.; Barbarossa, L.; Ercole, E.; et al. Convergent mutations and kinase fusions lead to oncogenic STAT3 activation in anaplastic large cell lymphoma. Cancer Cell 2015, 27, 516–532. [Google Scholar] [CrossRef] [Green Version]
- Nagel, S.; Pommerenke, C.; Meyer, C.; Kaufmann, M.; MacLeod, R.A.F.; Drexler, H.G. Aberrant expression of NKL homeobox gene HLX in Hodgkin lymphoma. Oncotarget 2018, 9, 14338–14353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagel, S.; Uphoff, C.C.; Dirks, W.G.; Pommerenke, C.; Meyer, C.; Drexler, H.G. Epstein-Barr virus (EBV) activates NKL homeobox gene HLX in DLBCL. PLoS ONE 2019, 14, e0216898. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Ehrentraut, S.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Oncogenic deregulation of NKL homeobox gene MSX1 in mantle cell lymphoma. Leuk. Lymphoma 2014, 55, 1893–1903. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.; Pommerenke, C.; Meyer, C.; Kaufmann, M.; Drexler, H.G.; MacLeod, R.A. Deregulation of polycomb repressor complex 1 modifier AUTS2 in T-cell leukemia. Oncotarget 2016, 7, 45398–45413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grocott, T.; Tambalo, M.; Streit, A. The peripheral sensory nervous system in the vertebrate head: A gene regulatory perspective. Dev. Biol. 2012, 370, 3–23. [Google Scholar] [CrossRef]
- Saunders, A.; Faiola, F.; Wang, J. Concise review: Pursuing self-renewal and pluripotency with the stem cell factor Nanog. STEM CELLS 2013, 31, 1227–1236. [Google Scholar] [CrossRef] [Green Version]
- Das, B.; Pal, B.; Bhuyan, R.; Li, H.; Sarma, A.; Gayan, S.; Talukdar, J.; Sandhya, S.; Bhuyan, S.; Gogoi, G.; et al. MYC Regulates the HIF2α Stemness Pathway via Nanog and Sox2 to Maintain Self-Renewal in Cancer Stem Cells versus Non-Stem Cancer Cells. Cancer Res. 2019, 79, 4015–4025. [Google Scholar] [CrossRef] [PubMed]
- Mariyath, M.P.M.; Shahi, M.H.; Farheen, S.; Tayyab, M.; Khanam, N.; Ali, A. Novel Homeodomain Transcription Factor Nkx2.2 in the Brain Tumor Development. Curr. Cancer Drug Targets 2020, 20, 335–340. [Google Scholar] [CrossRef]
- Habener, J.F.; Kemp, D.M.; Thomas, M.K. Minireview: Transcriptional regulation in pancreatic development. Endocrinology 2005, 146, 1025–1034. [Google Scholar] [CrossRef]
- Drexler, H.G.; MacLeod, R.A. Malignant hematopoietic cell lines: In vitro models for the study of anaplastic large-cell lymphoma. Leukemia 2004, 18, 1569–1571. [Google Scholar] [CrossRef]
- Drexler, H.G.; Matsuo, Y.; MacLeod, R.A. Malignant hematopoietic cell lines: In vitro models for the study of erythroleukemia. Leuk. Res. 2004, 28, 1243–1251. [Google Scholar] [CrossRef]
- Drexler, H.G.; Quentmeier, H.; MacLeod, R.A.; Uphoff, C.C.; Hu, Z.B. Leukemia cell lines: In vitro models for the study of acute promyelocytic leukemia. Leuk. Res. 1995, 19, 681–691. [Google Scholar] [CrossRef]
- Drexler, H.G. Leukemia cell lines: In vitro models for the study of chronic myeloid leukemia. Leuk. Res. 1994, 18, 919–927. [Google Scholar] [CrossRef]
- Matsuo, Y.; Drexler, H.G. Establishment and characterization of human B cell precursor-leukemia cell lines. Leuk. Res. 1998, 22, 567–579. [Google Scholar] [CrossRef]
- Drexler, H.G.; Eberth, S.; Nagel, S.; MacLeod, R.A. Malignant hematopoietic cell lines: In vitro models for double-hit B-cell lymphomas. Leuk. Lymphoma 2016, 57, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G.; Pommerenke, C.; Eberth, S.; Nagel, S. Hodgkin lymphoma cell lines: To separate the wheat from the chaff. Biol. Chem. 2018, 399, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Drexler, H.G. Malignant hematopoietic cell lines: In vitro models for the study of myelodysplastic syndromes. Leuk. Res. 2000, 24, 109–115. [Google Scholar] [CrossRef]
- Drexler, H.G.; Matsuo, Y. Malignant hematopoietic cell lines: In vitro models for the study of multiple myeloma and plasma cell leukemia. Leuk. Res. 2000, 24, 681–703. [Google Scholar] [CrossRef]
- Drexler, H.G.; Matsuo, Y. Malignant hematopoietic cell lines: In vitro models for the study of natural killer cell leukemia-lymphoma. Leukemia 2000, 14, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Carbone, A.; Cesarman, E.; Gloghini, A.; Drexler, H.G. Understanding pathogenetic aspects and clinical presentation of primary effusion lymphoma through its derived cell lines. AIDS 2010, 24, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Drexler, H.G.; Ehrentraut, S.; Nagel, S.; Eberth, S.; MacLeod, R.A. Malignant hematopoietic cell lines: In vitro models for the study of primary mediastinal B-cell lymphomas. Leuk. Res. 2015, 39, 18–29. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagel, S. The Role of NKL Homeobox Genes in T-Cell Malignancies. Biomedicines 2021, 9, 1676. https://doi.org/10.3390/biomedicines9111676
Nagel S. The Role of NKL Homeobox Genes in T-Cell Malignancies. Biomedicines. 2021; 9(11):1676. https://doi.org/10.3390/biomedicines9111676
Chicago/Turabian StyleNagel, Stefan. 2021. "The Role of NKL Homeobox Genes in T-Cell Malignancies" Biomedicines 9, no. 11: 1676. https://doi.org/10.3390/biomedicines9111676
APA StyleNagel, S. (2021). The Role of NKL Homeobox Genes in T-Cell Malignancies. Biomedicines, 9(11), 1676. https://doi.org/10.3390/biomedicines9111676