
Somatostatin and Its Receptor System in Colorectal Cancer


Abstract
:1. Introduction
2. The SRIF System—General Comments
2.1. Somatostatin (SST) Localization and Role in Physiology
2.2. SST Secretion Control in the Gastrointestinal Tract
2.3. Somatostatin Receptor Localization and Role in Physiology
3. The SRIF System and Different Tumors
4. The SRIF System and Large Intestine in Physiology
5. The SRIF System and Inflammatory Bowel Diseases
6. The SRIF System and Colorectal Cancer
6.1. Somatostatin Tissue Expression In Vivo
6.2. Somatostatin Receptor Tissue Expression In Vivo
6.3. The SRIF System Component Expression in Colorectal Cancer In Vitro
7. The SRIF System and Colorectal Cancer Histogenesis
7.1. New Nomenclature of Neuroendocrine Neoplasms
7.2. Enteroendocrine Cells in Colorectal Cancer Histogenesis
7.3. The SRIF System and Sporadic Colorectal Cancer Histogenesis
8. Epigenetic Alterations of the SRIF System Components in Colorectal Cancer
9. Clinical Application of the SRIF System Components
9.1. Diagnostics
9.2. The SRIF System Components in Therapy of Cancers
Somatostatin Analogues in Neuroendocrine and Sporadic Colorectal Cancer Therapy
10. Conclusions and Future Challenges
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA | amino acid |
| ACL | adenylate cyclase |
| ALDH+ | aldehyde dehydrogenase |
| APC | adenomatous polyposis coli |
| APUD (DNES) cells | amine precursor uptake decarboxylation; diffuse neuroendocrine system cells |
| CGRP | calcitonin gene-related peptide |
| CD | Crohn’s disease |
| c-Met | tyrosine-protein kinase Met or hepatocyte growth factor receptor |
| CMTM3 | CKLF-like MARVEL transmembrane domain containing protein 3 |
| CRC | colorectal cancer |
| CNS | central nervous system |
| CSCs | cancer stem cells |
| CST | cortistatin |
| DAB | 3,3′-diaminobenzidine |
| DFS | disease-free survival |
| DSS | dextran sodium sulfate |
| ECL | enterochromaffin-like |
| EECs | enteroendocrine cells |
| ENS | enteric nervous system |
| ERK1/2 | extracellular signal-regulated kinase 1/2 |
| ESCs | embryonic stem cells |
| FAP | familial adenomatous coli |
| GH | growth hormone |
| GIP | gastric inhibitory peptide |
| GLP-1/-2/R | glucagon-like peptide-1, -2/Receptor |
| GPCR | G protein-coupled receptors |
| 5-HT | 5-hydroxytryptamine (serotonin) |
| IBD | inflammatory bowel diseases |
| IHC | immunohistochemistry |
| IL-6 | interleukin-6 |
| IFNγ | interferon γ |
| ISH | in situ hybridization |
| MANEC/MiNENs | mixed adenoneuroendocrine carcinoma/mixed neuroendocrine–non-neuroendocrine neoplasms |
| MAPK | mitogen-activated protein kinase (originally called ERK) |
| MDFI | MyoD family inhibitor |
| MSI-L | low-level microsatellite stability |
| NCs | neuroendocrine cells |
| NEN | neuroendocrine neoplasm |
| NET | neuroendocrine tumor |
| OCT | octreotide |
| OS | overall survival |
| PACAP | pituitary adenylate cyclase-activating peptide |
| PDAC | pancreatic ductal adenocarcinoma |
| panNET/pNET | pancreatic NET |
| PC2 | proprotein convertase 2 |
| PFS | progression-free survival |
| PNS | peripheral nervous system |
| PP | pancreatic polypeptide |
| PTP | protein phosphotyrosine phosphatase |
| P2Y(1) | purinoceptor 1; belongs to GPCR |
| PYY | peptide YY |
| RA | retinoic acid |
| SCs | stem cells |
| SH2 | Src homology 2 |
| SRCC | signet-ring cell carcinoma |
| SRIF/SRIH/SST | somatotropin-release inhibitory factor/hormone/somatostatin |
| SSAs | somatostatin analogues |
| TNBS | trinitrobenzene sulfonic acid |
| UC | ulcerative colitis |
| VIP | vasoactive intestinal peptide |
References
- Harada, S.; Morlote, D. Molecular Pathology of Colorectal Cancer. Adv. Anat. Pathol. 2020, 27, 20–26. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Fleming, M.; Ravula, S.; Tatishchev, S.F.; Wang, H.L. Colorectal carcinoma: Pathologic aspects. J. Gastrointest. Oncol. 2012, 3, 153–173. [Google Scholar] [CrossRef]
- Bosman, F.; Yan, P. Molecular pathology of colorectal cancer. Pol. J. Pathol. 2014, 65, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Munro, M.J.; Wickremesekera, S.K.; Peng, L.; Tan, S.T.; Itinteang, T. Cancer stem cells in colorectal cancer: A review. J. Clin. Pathol. 2018, 71, 110–116. [Google Scholar] [CrossRef]
- Mármol, I.; Sánchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal Carcinoma: A General Overview and Future Perspectives in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [Green Version]
- Jung, G.; Hernández-Illán, E.; Moreira, L.; Balaguer, F.; Goel, A. Epigenetics of colorectal cancer: Biomarker and therapeutic potential. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 111–130. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, T.; Okada, K.; Tabei, R. Immunoreactivity to neurohormonal polypeptide in colorectal carcinomas and tumor-neighboring mucosa, and its significance. Arch. Pathol. Lab. Med. 1996, 120, 560–568. [Google Scholar]
- Ohmori, T.; Asahi, S.; Sato, C.; Maki, F.; Masumoto, A.; Okada, K. Bcl-2 protein expression and gut neurohormonal polypeptide/amine production in colorectal carcinomas and tumor-neighboring mucosa, which closely correlate to the occurrence of tumor. Histol. Histopathol. 1999, 14, 37–44. [Google Scholar] [CrossRef]
- Modarai, S.R.; Opdenaker, L.M.; Viswanathan, V.; Fields, J.Z.; Boman, B.M. Somatostatin signaling via SSTR1 contributes to the quiescence of colon cancer stem cells. BMC Cancer 2016, 16, 941. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Ahn, K.; Emerick, B.; Modarai, S.R.; Opdenaker, L.M.; Palazzo, J.; Schleiniger, G.; Fields, J.Z.; Boman, B.M. APC mutations in human colon lead to decreased neuroendocrine maturation of ALDH+ stem cells that alters GLP-2 and SST feedback signaling: Clue to a link between WNT and retinoic acid signalling in colon cancer development. PLoS ONE 2020, 15, e0239601. [Google Scholar] [CrossRef]
- Günther, T.; Tulipano, G.; Dournaud, P.; Bousquet, C.; Csaba, Z.; Kreienkamp, H.J.; Lupp, A.; Korbonits, M.; Castaño, J.P.; Wester, H.J.; et al. International Union of Basic and Clinical Pharmacology. CV. Somatostatin Receptors: Structure, Function, Ligands, and New Nomenclature. Pharmacol. Rev. 2018, 70, 763–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casnici, C.; Lattuada, D.; Perego, C.; Franco, P.; Marelli, O. Inhibitory effect of somatostatin on human T lymphocytes proliferation. Int. J. Immunopharmacol. 1997, 19, 721–727. [Google Scholar] [CrossRef]
- Ten Bokum, A.M.; Hofland, L.J.; van Hagen, P.M. Somatostatin and somatostatin receptors in the immune system: A review. Eur. Cytokine Netw. 2000, 11, 161–176. [Google Scholar] [PubMed]
- Rosskopf, D.; Schürks, M.; Manthey, I.; Joisten, M.; Busch, S.; Siffert, W. Signal transduction of somatostatin in human B lymphoblasts. Am. J. Physiol. Cell Physiol. 2003, 284, C179–C190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbieri, F.; Bajetto, A.; Pattarozzi, A.; Gatti, M.; Würth, R.; Thellung, S.; Corsaro, A.; Villa, V.; Nizzari, M.; Florio, T. Peptide receptor targeting in cancer: The somatostatin paradigm. Int. J. Pept. 2013, 2013, 926295. [Google Scholar] [CrossRef]
- Ruscica, M.; Arvigo, M.; Steffani, L.; Ferone, D.; Magni, P. Somatostatin, somatostatin analogs and somatostatin receptor dynamics in the biology of cancer progression. Curr. Mol. Med. 2013, 13, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Leiszter, K.; Sipos, F.; Galamb, O.; Krenács, T.; Veres, G.; Wichmann, B.; Fűri, I.; Kalmár, A.; Patai, Á.V.; Tóth, K.; et al. Promoter hypermethylation-related reduced somatostatin production promotes uncontrolled cell proliferation in colorectal cancer. PLoS ONE 2015, 10, e0118332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moody, T.W.; Ramos-Alvarez, I.; Jensen, R.T. Neuropeptide G Protein-Coupled Receptors as Oncotargets. Front. Endocrinol. 2018, 9, 345. [Google Scholar] [CrossRef] [Green Version]
- Gatto, F.; Barbieri, F.; Arvigo, M.; Thellung, S.; Amarù, J.; Albertelli, M.; Ferone, D.; Florio, T. Biological and Biochemical Basis of the Differential Efficacy of First and Second Generation Somatostatin Receptor Ligands in Neuroendocrine Neoplasms. Int. J. Mol. Sci. 2019, 20, 3940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyronnet, S.; Bousquet, C.; Najib, S.; Azar, R.; Laklai, H.; Susini, C. Antitumor effects of somatostatin. Mol. Cell Endocrinol. 2008, 286, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rai, U.; Thrimawithana, T.R.; Valery, C.; Young, S.A. Therapeutic uses of somatostatin and its analogues: Current view and potential applications. Pharmacol. Ther. 2015, 152, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Cives, M.; Strosberg, J. Radionuclide Therapy for Neuroendocrine Tumors. Curr. Oncol. Rep. 2017, 19, 9. [Google Scholar] [CrossRef]
- Shamsi, B.H.; Chatoo, M.; Xu, X.K.; Xu, X.; Chen, X.Q. Versatile Functions of Somatostatin and Somatostatin Receptors in the Gastrointestinal System. Front. Endocrinol. 2021, 12, 652363. [Google Scholar] [CrossRef]
- Dawod, M.; Gordoa, T.A.; Cives, M.; De Mestier, L.; Crona, J.; Spada, F.; Oberg, K.; Pavel, M.; Lamarca, A. Antiproliferative Systemic Therapies for Metastatic Small Bowel Neuroendocrine Tumours. Curr. Treat. Options Oncol. 2021, 22, 73. [Google Scholar] [CrossRef]
- Rinzivillo, M.; De Felice, I.; Magi, L.; Annibale, B.; Panzuto, F. Octreotide long-acting release (LAR) in combination with other therapies for treatment of neuroendocrine neoplasia: A systematic review. J. Gastrointest. Oncol. 2021, 12, 845–855. [Google Scholar] [CrossRef]
- Benali, N.; Ferjoux, G.; Puente, E.; Buscail, L.; Susini, C. Somatostatin receptors. Digestion 2000, 62 (Suppl. 1), 27–32. [Google Scholar] [CrossRef] [PubMed]
- Ampofo, E.; Nalbach, L.; Menger, M.D.; Laschke, M.W. Regulatory Mechanisms of Somatostatin Expression. Int. J. Mol. Sci. 2020, 21, 4170. [Google Scholar] [CrossRef]
- Polak, J.M.; Pearse, A.G.; Grimelius, L.; Bloom, S.R. Growth-hormone release-inhibiting hormone in gastrointestinal and pancreatic D cells. Lancet 1975, 1, 1220–1222. [Google Scholar] [CrossRef]
- Penman, E.; Wass, J.A.; Butler, M.G.; Penny, E.S.; Price, J.; Wu, P.; Rees, L.H. Distribution and characterisation of immunoreactive somatostatin in human gastrointestinal tract. Regul. Pept. 1983, 7, 53–65. [Google Scholar] [CrossRef]
- Adriaenssens, A.; Lam, B.Y.; Billing, L.; Skeffington, K.; Sewing, S.; Reimann, F.; Gribble, F. A Transcriptome-Led Exploration of Molecular Mechanisms Regulating Somatostatin-Producing D-Cells in the Gastric Epithelium. Endocrinology 2015, 156, 3924–3936. [Google Scholar] [CrossRef] [Green Version]
- Fothergill, L.J.; Furness, J.B. Diversity of enteroendocrine cells investigated at cellular and subcellular levels: The need for a new classification scheme. Histochem. Cell Biol. 2018, 150, 693–702. [Google Scholar] [CrossRef] [PubMed]
- Schubert, M.L.; Rehfeld, J.F. Gastric Peptides-Gastrin and Somatostatin. Compr. Physiol. 2019, 10, 197–228. [Google Scholar] [CrossRef] [PubMed]
- Patel, Y.C.; Reichlin, S. Somatostatin in hypothalamus, extrahypothalamic brain, and peripheral tissues of the rat. Endocrinology 1978, 102, 523–530. [Google Scholar] [CrossRef]
- Brissova, M.; Fowler, M.J.; Nicholson, W.E.; Chu, A.; Hirshberg, B.; Harlan, D.M.; Powers, A.C. Assessment of human pancreatic islet architecture and composition by laser scanning confocal microscopy. J. Histochem. Cytochem. 2005, 53, 1087–1097. [Google Scholar] [CrossRef]
- Baskin, D.G. A Historical Perspective on the Identification of Cell Types in Pancreatic Islets of Langerhans by Staining and Histochemical Techniques. J. Histochem. Cytochem. 2015, 63, 543–558. [Google Scholar] [CrossRef] [Green Version]
- Rorsman, P.; Huising, M.O. The somatostatin-secreting pancreatic δ-cell in health and disease. Nat. Rev. Endocrinol. 2018, 14, 404–414. [Google Scholar] [CrossRef]
- Weidmann, S.; Schrödl, F.; Neuhuber, W.; Brehmer, A. Quantitative estimation of putative primary afferent neurons in the myenteric plexus of human small intestine. Histochem. Cell Biol. 2007, 128, 399–407. [Google Scholar] [CrossRef]
- Gonkowski, S.; Rytel, L. Somatostatin as an Active Substance in the Mammalian Enteric Nervous System. Int. J. Mol. Sci. 2019, 20, 4461. [Google Scholar] [CrossRef] [Green Version]
- Brehmer, A. Classification of human enteric neurons. Histochem. Cell Biol. 2021, 156, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Yang, T.; Zhang, Q. δ-Cells: The Neighborhood Watch in the Islet Community. Biology 2021, 10, 74. [Google Scholar] [CrossRef]
- Francis, B.H.; Baskin, D.G.; Saunders, D.R.; Ensinck, J.W. Distribution of somatostatin-14 and somatostatin-28 gastrointestinal-pancreatic cells of rats and humans. Gastroenterology 1990, 99, 1283–1291. [Google Scholar] [CrossRef]
- Schaer, J.C.; Waser, B.; Mengod, G.; Reubi, J.C. Somatostatin receptor subtypes sst1, sst2, sst3 and sst5 expression in human pituitary, gastroentero-pancreatic and mammary tumors: Comparison of mRNA analysis with receptor autoradiography. Int. J. Cancer 1997, 70, 530–537. [Google Scholar] [CrossRef]
- Reubi, J.C.; Waser, B.; Schaer, J.C.; Laissue, J.A. Somatostatin receptor sst1-sst5 expression in normal and neoplastic human tissues using receptor autoradiography with subtype-selective ligands. Eur. J. Nucl. Med. 2001, 28, 836–846. [Google Scholar] [CrossRef]
- Corleto, V.D.; Falconi, M.; Panzuto, F.; Milione, M.; De Luca, O.; Perri, P.; Cannizzaro, R.; Bordi, C.; Pederzoli, P.; Scarpa, A.; et al. Somatostatin receptor subtypes 2 and 5 are associated with better survival in well-differentiated endocrine carcinomas. Neuroendocrinology 2009, 89, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Cives, M.; Strosberg, J.R. Gastroenteropancreatic Neuroendocrine Tumors. CA Cancer J. Clin. 2018, 68, 471–487. [Google Scholar] [CrossRef]
- Qian, Z.R.; Li, T.; Ter-Minassian, M.; Yang, J.; Chan, J.A.; Brais, L.K.; Masugi, Y.; Thiaglingam, A.; Brooks, N.; Nishihara, R.; et al. Association Between Somatostatin Receptor Expression and Clinical Outcomes in Neuroendocrine Tumors. Pancreas 2016, 45, 1386–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmona Matos, D.M.; Jang, S.; Hijaz, B.; Chang, A.W.; Lloyd, R.V.; Chen, H.; Jaskula-Sztul, R. Characterization of somatostatin receptors (SSTRs) expression and antiproliferative effect of somatostatin analogues in aggressive thyroid cancers. Surgery 2019, 165, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Klomp, M.J.; Dalm, S.U.; de Jong, M.; Feelders, R.A.; Hofland, J.; Hofland, L.J. Epigenetic regulation of somatostatin and somatostatin receptors in neuroendocrine tumors and other types of cancer. Rev. Endocr. Metab. Disord. 2021, 22, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Reubi, J.C. Somatostatin and other Peptide receptors as tools for tumor diagnosis and treatment. Neuroendocrinology 2004, 80 (Suppl. 1), 51–56. [Google Scholar] [CrossRef] [PubMed]
- Reubi, J.C. Peptide receptor expression in GEP-NET. Virchows Arch. 2007, 451 (Suppl. 1), S47–S50. [Google Scholar] [CrossRef] [PubMed]
- Jesinghaus, M.; Konukiewitz, B.; Keller, G.; Kloor, M.; Steiger, K.; Reiche, M.; Penzel, R.; Endris, V.; Arsenic, R.; Hermann, G.; et al. Colorectal mixed adenoneuroendocrine carcinomas and neuroendocrine carcinomas are genetically closely related to colorectal adenocarcinomas. Mod. Pathol. 2017, 30, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Smitha, M.C.; Maggi, M.; Orlando, C. Somatostatin receptors in non-endocrine tumours. Dig. Liver Dis. 2004, 36 (Suppl. 1), S78–S85. [Google Scholar] [CrossRef] [PubMed]
- Raggi, C.C.; Cianchi, F.; Valanzano, R.; Smith, M.C.; Serio, M.; Maggi, M.; Orlando, C. Prognostic value of somatostatin receptor subtype 2 expression in colorectal cancer. Regul. Pept. 2005, 132, 23–26. [Google Scholar] [CrossRef]
- Swatek, J.; Chibowski, D. Endocrine cells in colorectal carcinomas. Immunohistochemical study. Pol. J. Pathol. 2000, 51, 127–136. [Google Scholar] [PubMed]
- Ogimi, T.; Sadahiro, S.; Kamei, Y.; Chan, L.F.; Miyakita, H.; Saito, G.; Okada, K.; Suzuki, T.; Kajiwara, H. Distribution of Neuroendocrine Marker-Positive Cells in Colorectal Cancer Tissue and Normal Mucosal Tissue: Consideration of Histogenesis of Neuroendocrine Cancer. Oncology 2019, 97, 294–300. [Google Scholar] [CrossRef]
- Seretis, E.; Konstantinidou, A.; Arnogiannakis, N.; Xinopoulos, D.; Voloudakis-Baltatzis, I.E. Mucinous colorectal adenocarcinoma with signet-ring cells: Immunohistochemical and ultrastructural study. Ultrastruct. Pathol. 2010, 34, 337–343. [Google Scholar] [CrossRef] [PubMed]
- La Salvia, A.; Espinosa-Olarte, P.; Riesco-Martinez, M.D.C.; Anton-Pascual, B.; Garcia-Carbonero, R. Targeted Cancer Therapy: What’s New in the Field of Neuroendocrine Neoplasms? Cancers 2021, 13, 1701. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Williams, M.; Cheng, Y.Y.; Leung, W.K. Roles of Methylated DNA Biomarkers in Patients with Colorectal Cancer. Dis. Markers 2019, 2019, 2673543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.P.; Pictet, R.L.; Rutter, W.J. Human somatostatin I: Sequence of the cDNA. Proc. Natl. Acad. Sci. USA 1982, 79, 4575–4579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.P.; Rutter, W.J. Sequence of the human somatostatin I gene. Science 1984, 224, 168–171. [Google Scholar] [CrossRef] [PubMed]
- SST and SST1–5. Available online: https://www.GeneCards (accessed on 15 October 2021).
- Warren, T.G.; Shields, D. Expression of preprosomatostatin in heterologous cells: Biosynthesis, posttranslational processing, and secretion of mature somatostatin. Cell 1984, 39, 547–555. [Google Scholar] [CrossRef]
- Krulich, L.; Dhariwal, A.P.; McCann, S.M. Stimulatory and inhibitory effects of purified hypothalamic extracts on growth hormone release from rat pituitary in vitro. Endocrinology 1968, 83, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Brazeau, P.; Vale, W.; Burgus, R.; Ling, N.; Butcher, M.; Rivier, J.; Guillemin, R. Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 1973, 179, 77–79. [Google Scholar] [CrossRef]
- Pradayrol, L.; Jörnvall, H.; Mutt, V.; Ribet, A. N-terminally extended somatostatin: The primary structure of somatostatin-28. FEBS Lett. 1980, 109, 55–58. [Google Scholar] [CrossRef] [Green Version]
- Reichlin, S. Somatostatin. N. Engl. J. Med. 1983, 309, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- Pittaluga, A.; Roggeri, A.; Vallarino, G.; Olivero, G. Somatostatin, a Presynaptic Modulator of Glutamatergic Signal in the Central Nervous System. Int. J. Mol. Sci. 2021, 22, 5864. [Google Scholar] [CrossRef]
- Polak, J.M.; Bloom, S.R. Somatostatin localization in tissues. Scand. J. Gastroenterol. Suppl. 1986, 119, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Merighi, A.; Polak, J.M.; Gibson, S.J.; Gulbenkian, S.; Valentino, K.L.; Peirone, S.M. Ultrastructural studies on calcitonin gene-related peptide-, tachykinins- and somatostatin-immunoreactive neurones in rat dorsal root ganglia: Evidence for the colocalization of different peptides in single secretory granules. Cell Tissue Res. 1988, 254, 101–109. [Google Scholar] [CrossRef]
- Huising, M.O.; van der Meulen, T.; Huang, J.L.; Pourhosseinzadeh, M.S.; Noguchi, G.M. The Difference δ-Cells Make in Glucose Control. Physiology 2018, 33, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Brereton, M.F.; Vergari, E.; Zhang, Q.; Clark, A. Alpha-, Delta- and PP-cells: Are They the Architectural Cornerstones of Islet Structure and Co-ordination? J. Histochem. Cytochem. 2015, 63, 575–591. [Google Scholar] [CrossRef] [Green Version]
- Da Silva Xavier, G. The Cells of the Islets of Langerhans. J. Clin. Med. 2018, 7, 54. [Google Scholar] [CrossRef] [Green Version]
- Habib, A.M.; Richards, P.; Cairns, L.S.; Rogers, G.J.; Bannon, C.A.; Parker, H.E.; Morley, T.C.; Yeo, G.S.; Reimann, F.; Gribble, F.M. Overlap of endocrine hormone expression in the mouse intestine revealed by transcriptional profiling and flow cytometry. Endocrinology 2012, 153, 3054–3065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drucker, D.J. Evolving Concepts and Translational Relevance of Enteroendocrine Cell Biology. J. Clin. Endocrinol. Metab. 2016, 101, 778–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beumer, J.; Gehart, H.; Clevers, H. Enteroendocrine Dynamics—New Tools Reveal Hormonal Plasticity in the Gut. Endocr. Rev. 2020, 41, 695–706. [Google Scholar] [CrossRef]
- Haber, A.L.; Biton, M.; Rogel, N.; Herbst, R.H.; Shekhar, K.; Smillie, C.; Burgin, G.; Delorey, T.M.; Howitt, M.R.; Katz, Y.; et al. A single-cell survey of the small intestinal epithelium. Nature 2017, 551, 333–339. [Google Scholar] [CrossRef]
- O’Briain, D.S.; Dayal, Y.; DeLellis, R.A.; Tischler, A.S.; Bendon, R.; Wolfe, H.J. Rectal carcinoids as tumors of the hindgut endocrine cells: A morphological and immunohistochemical analysis. Am. J. Surg. Pathol. 1982, 6, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Martins, P.; Fakhry, J.; de Oliveira, E.C.; Hunne, B.; Fothergill, L.J.; Ringuet, M.; Reis, D.D.; Rehfeld, J.F.; Callaghan, B.; Furness, J.B. Analysis of enteroendocrine cell populations in the human colon. Cell Tissue Res. 2017, 367, 161–168. [Google Scholar] [CrossRef]
- Gunawardene, A.R.; Corfe, B.M.; Staton, C.A. Classification and functions of enteroendocrine cells of the lower gastrointestinal tract. Int. J. Exp. Pathol. 2011, 92, 219–231. [Google Scholar] [CrossRef]
- Diwakarla, S.; Fothergill, L.J.; Fakhry, J.; Callaghan, B.; Furness, J.B. Heterogeneity of enterochromaffin cells within the gastrointestinal tract. Neurogastroenterol. Motil. 2017, 29, e13101. [Google Scholar] [CrossRef] [PubMed]
- Zabel, M. Ultrastructural localization of calcitonin, somatostatin and serotonin in parafollicular cells of rat thyroid. Histochem. J. 1984, 16, 1265–1272. [Google Scholar] [CrossRef]
- Zabel, M.; Schäfer, H. Ultrastructural localization of calcitonin and somatostatin in C cells of rabbit thyroid. Cell Tissue Res. 1986, 245, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, B.; Czykier, E.; Kasacka, I.; Kasprzak, A.; Zabel, M. Hybridocytochemical detection of mRNA for calcitonin, CGRP, NPY and somatostatin in thyroid parafollicular (C) cells in three rodent species. Folia Histochem. Et Cytobiol. 2002, 40, 183–184. [Google Scholar] [PubMed]
- Seidel, J.; Zabel, M.; Kasprzak, A.; Spachacz, R. The expression of calcitonin, calcitonin gene-related peptide and somatostatin in the thyroids of rats of different ages. Folia Morphol. 2003, 62, 485–487. [Google Scholar]
- Unger, N.; Ueberberg, B.; Schulz, S.; Saeger, W.; Mann, K.; Petersenn, S. Differential expression of somatostatin receptor subtype 1-5 proteins in numerous human normal tissues. Exp. Clin. Endocrinol. Diabetes 2012, 120, 482–489. [Google Scholar] [CrossRef]
- Bhandari, S.; Watson, N.; Long, E.; Sharpe, S.; Zhong, W.; Xu, S.Z.; Atkin, S.L. Expression of somatostatin and somatostatin receptor subtypes 1-5 in human normal and diseased kidney. J. Histochem. Cytochem. 2008, 56, 733–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Yang, W.; Li, Y.; Cong, Y. Enteroendocrine Cells: Sensing Gut Microbiota and Regulating Inflammatory Bowel Diseases. Inflamm. Bowel Dis. 2020, 26, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Svendsen, B.; Holst, J.J. Paracrine regulation of somatostatin secretion by insulin and glucagon in mouse pancreatic islets. Diabetologia 2021, 64, 142–151. [Google Scholar] [CrossRef]
- Low, M.J. Clinical endocrinology and metabolism: The somatostatin neuroendocrine system: Physiology and clinical relevance in gastrointestinal and pancreatic disorders. Best Pract. Res. Clin. Endocrinol. Metab. 2004, 18, 607–622. [Google Scholar] [CrossRef] [PubMed]
- Billing, L.J.; Larraufie, P.; Lewis, J.; Leiter, A.; Li, J.; Lam, B.; Yeo, G.S.; Goldspink, D.A.; Kay, R.G.; Gribble, F.M.; et al. Single cell transcriptomic profiling of large intestinal enteroendocrine cells in mice—Identification of selective stimuli for insulin-like peptide-5 and glucagon-like peptide-1 co-expressing cells. Mol. Metab. 2019, 29, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, M.; Ramla, D.; Seidah, N.G.; Chrétien, M. Developmental expression of the prohormone convertases PC1 and PC2 in mouse pancreatic islets. Endocrinology 1994, 135, 1651–1660. [Google Scholar] [CrossRef] [PubMed]
- Ravazzola, M.; Benoit, R.; Ling, N.; Orci, L. Prosomatostatin-derived antrin is present in gastric D cells and in portal blood. J. Clin. Investig. 1989, 83, 362–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, J.M.; Eriksson, B.; Grimelius, L.; Oberg, K.; Thim, L. Characterization of three peptides derived from prosomatostatin [prosomatostatin-(1-63)-, -(65-76)- and -(79-92)-peptides] in a human pancreatic tumour. Biochem. J. 1987, 248, 123–127. [Google Scholar] [CrossRef] [Green Version]
- Adriaenssens, A.E.; Svendsen, B.; Lam, B.Y.; Yeo, G.S.; Holst, J.J.; Reimann, F.; Gribble, F.M. Transcriptomic profiling of pancreatic alpha, beta and delta cell populations identifies delta cells as a principal target for ghrelin in mouse islets. Diabetologia 2016, 59, 2156–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Meulen, T.; Donaldson, C.J.; Cáceres, E.; Hunter, A.E.; Cowing-Zitron, C.; Pound, L.D.; Adams, M.W.; Zembrzycki, A.; Grove, K.L.; Huising, M.O. Urocortin3 mediates somatostatin-dependent negative feedback control of insulin secretion. Nat. Med. 2015, 21, 769–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, M.; Ramracheya, R.; Amisten, S.; Bengtsson, M.; Moritoh, Y.; Zhang, Q.; Johnson, P.R.; Rorsman, P. Somatostatin release, electrical activity, membrane currents and exocytosis in human pancreatic delta cells. Diabetologia 2009, 52, 1566–1578. [Google Scholar] [CrossRef] [Green Version]
- Salehi, A.; Qader, S.S.; Grapengiesser, E.; Hellman, B. Pulses of somatostatin release are slightly delayed compared with insulin and antisynchronous to glucagon. Regul. Pept. 2007, 144, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Salehi, A.; Parandeh, F.; Fredholm, B.B.; Grapengiesser, E.; Hellman, B. Absence of adenosine A1 receptors unmasks pulses of insulin release and prolongs those of glucagon and somatostatin. Life Sci. 2009, 85, 470–476. [Google Scholar] [CrossRef]
- Patel, Y.C. Somatostatin and its receptor family. Front. Neuroendocrinol. 1999, 20, 157–198. [Google Scholar] [CrossRef]
- Csaba, Z.; Peineau, S.; Dournaud, P. Molecular mechanisms of somatostatin receptor trafficking. J. Mol. Endocrinol. 2012, 48, R1–R12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corness, J.D.; Demchyshyn, L.L.; Seeman, P.; Van Tol, H.H.; Srikant, C.B.; Kent, G.; Patel, Y.C.; Niznik, H.B. A human somatostatin receptor (SSTR3), located on chromosome 22, displays preferential affinity for somatostatin-14 like peptides. FEBS Lett. 1993, 321, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Reisine, T.; Law, S.F.; Ihara, Y.; Kubota, A.; Kagimoto, S.; Seino, M.; Seino, Y.; Bell, G.I.; Seino, S. Somatostatin receptors, an expanding gene family: Cloning and functional characterization of human SSTR3, a protein coupled to adenylyl cyclase. Mol. Endocrinol. 1992, 6, 2136–2142. [Google Scholar] [CrossRef] [Green Version]
- Yamada, Y.; Kagimoto, S.; Kubota, A.; Yasuda, K.; Masuda, K.; Someya, Y.; Ihara, Y.; Li, Q.; Imura, H.; Seino, S.; et al. Cloning, functional expression and pharmacological characterization of a fourth (hSSTR4) and a fifth (hSSTR5) human somatostatin receptor subtype. Biochem. Biophys. Res. Commun. 1993, 195, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Florio, T. Somatostatin/somatostatin receptor signalling: Phosphotyrosine phosphatases. Mol. Cell Endocrinol. 2008, 286, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Florio, T. Molecular mechanisms of the antiproliferative activity of somatostatin receptors (SSTRs) in neuroendocrine tumors. Front. Biosci. 2008, 13, 822–840. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Ye, Z.; Wang, F.; Qin, Y.; Xu, X.; Yu, X.; Ji, S. Role of Somatostatin Receptor in Pancreatic Neuroendocrine Tumor Development, Diagnosis, and Therapy. Front. Endocrinol. 2021, 12, 679000. [Google Scholar] [CrossRef] [PubMed]
- Solarski, M.; Williams, D.; Mehrabian, M.; Wang, H.; Wille, H.; Schmitt-Ulms, G. The human brain somatostatin interactome: SST binds selectively to P-type family ATPases. PLoS ONE 2019, 14, e0217392. [Google Scholar] [CrossRef] [PubMed]
- Reubi, J.C.; Mazzucchelli, L.; Hennig, I.; Laissue, J.A. Local up-regulation of neuropeptide receptors in host blood vessels around human colorectal cancers. Gastroenterology 1996, 110, 1719–1726. [Google Scholar] [CrossRef]
- Taniyama, Y.; Suzuki, T.; Mikami, Y.; Moriya, T.; Satomi, S.; Sasano, H. Systemic distribution of somatostatin receptor subtypes in human: An immunohistochemical study. Endocr. J. 2005, 52, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Bhanat, E.; Koch, C.A.; Parmar, R.; Garla, V.; Vijayakumar, V. Somatostatin receptor expression in non-classical locations-clinical relevance? Rev. Endocr. Metab. Disord. 2018, 19, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Ferone, D.; Pivonello, R.; Van Hagen, P.M.; Dalm, V.A.; Lichtenauer-Kaligis, E.G.; Waaijers, M.; Van Koetsveld, P.M.; Mooy, D.M.; Colao, A.; Minuto, F.; et al. Quantitative and functional expression of somatostatin receptor subtypes in human thymocytes. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E1056–E1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weckbecker, G.; Lewis, I.; Albert, R.; Schmid, H.A.; Hoyer, D.; Bruns, C. Opportunities in somatostatin research: Biological, chemical and therapeutic aspects. Nat. Rev. Drug Discov. 2003, 2, 999–1017. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, S.; Schulz, S. Intracellular trafficking of somatostatin receptors. Mol. Cell Endocrinol. 2008, 286, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Vuaroqueaux, V.; Dutour, A.; Briard, N.; Monges, G.; Grino, M.; Oliver, C.; Ouafik, L. No loss of sst receptors gene expression in advanced stages of colorectal cancer. Eur. J. Endocrinol. 1999, 140, 362–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuaroqueaux, V.; Dutour, A.; Bourhim, N.; Ouafik, L.; Monges, G.; Briard, N.; Sauze, N.; Oliver, C.; Grino, M. Increased expression of the mRNA encoding the somatostatin receptor subtype five in human colorectal adenocarcinoma. J. Mol. Endocrinol. 2000, 24, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Casini Raggi, C.; Calabrò, A.; Renzi, D.; Briganti, V.; Cianchi, F.; Messerini, L.; Valanzano, R.; Cameron Smith, M.; Cortesini, C.; Tonelli, F.; et al. Quantitative evaluation of somatostatin receptor subtype 2 expression in sporadic colorectal tumor and in the corresponding normal mucosa. Clin. Cancer Res. 2002, 8, 419–427. [Google Scholar]
- Qiu, C.Z.; Wang, C.; Huang, Z.X.; Zhu, S.Z.; Wu, Y.Y.; Qiu, J.L. Relationship between somatostatin receptor subtype expression and clinicopathology, Ki-67, Bcl-2 and p53 in colorectal cancer. World J. Gastroenterol. 2006, 12, 2011–2015. [Google Scholar] [CrossRef] [PubMed]
- Evangelou, I.; Petraki, C.; Msaouel, P.; Scorilas, A.; Sdrolia, E.; Padazi, G.; Koborozos, V.; Koutsilieris, M. Immunohistochemical expression of somatostatin receptor subtypes 2 and 5 in colorectal cancer. Eur. J. Clin. Investig. 2012, 42, 777–783. [Google Scholar] [CrossRef]
- Yerci, O.; Sehitoglu, I.; Ugras, N.; Cubukcu, E.; Yuce, S.; Bedir, R.; Cure, E. Somatostatin receptor 2 and 5 expressions in gastroenteropancreatic neuroendocrine tumors in Turkey. Asian Pac. J. Cancer Prev. 2015, 16, 4377–4381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamora, V.; Cabanne, A.; Salanova, R.; Bestani, C.; Domenichini, E.; Marmissolle, F.; Giacomi, N.; O’Connor, J.; Méndez, G.; Roca, E. Buenos Aires and La Plata Argentina Argentum Working Group. Immunohistochemical expression of somatostatin receptors in digestive endocrine tumours. Dig. Liver Dis. 2010, 42, 220–225. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.; Jin, K.; Fang, C.; Lin, Y.; Xue, L.; Feng, S.; Zhou, Z.; Shao, C.; Chen, M.; et al. Somatostatin receptor expression indicates improved prognosis in gastroenteropancreatic neuroendocrine neoplasm, and octreotide long-acting release is effective and safe in Chinese patients with advanced gastroenteropancreatic neuroendocrine tumors. Oncol. Lett. 2017, 13, 1165–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antelo, G.; Hierro, C.; Fernández, J.P.; Baena, E.; Bugés, C.; Layos, L.; Manzano, J.L.; Caro, M.; Mesia, R. Rectal neuroendocrine carcinoma: Case report of a rare entity and perspective review of promising agents. Drugs Context. 2020, 9, 2020-2-4. [Google Scholar] [CrossRef]
- Tanaka, S.; Yamasaki, S.; Matsushita, H.; Ozawa, Y.; Kurosaki, A.; Takeuchi, K.; Hoshihara, Y.; Doi, T.; Watanabe, G.; Kawaminami, K. Duodenal somatostatinoma: A case report and review of 31 cases with special reference to the relationship between tumor size and metastasis. Pathol. Int. 2000, 50, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.A.; Choi, W.H.; Kim, C.N.; Moon, Y.S.; Chang, S.H.; Lee, H.R. Duodenal somatostatinoma: A case report and review. Korean J. Intern. Med. 2011, 26, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.Y.; Gong, Y.F.; Zhuang, H.K.; Zhou, Z.X.; Huang, S.Z.; Zou, Y.P.; Huang, B.W.; Sun, Z.H.; Zhang, C.Z.; Tang, Y.Q.; et al. Pancreatic neuroendocrine tumors: A review of serum biomarkers, staging, and management. World J. Gastroenterol. 2020, 26, 2305–2322. [Google Scholar] [CrossRef] [PubMed]
- Reubi, J.C.; Kvols, L.; Krenning, E.; Lamberts, S.W. In vitro and in vivo detection of somatostatin receptors in human malignant tissues. Acta Oncol. 1991, 30, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Okuwaki, K.; Kida, M.; Mikami, T.; Yamauchi, H.; Imaizumi, H.; Miyazawa, S.; Iwai, T.; Takezawa, M.; Saegusa, M.; Watanabe, M.; et al. Clinicopathologic characteristics of pancreatic neuroendocrine tumors and relation of somatostatin receptor type 2A to outcomes. Cancer 2013, 119, 4094–4102. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Binderup, T.; Langer, S.W.; Kjaer, A.; Knigge, P.; Grøndahl, V.; Melchior, L.; Federspiel, B.; Knigge, U. P53, Somatostatin receptor 2a and Chromogranin A immunostaining as prognostic markers in high grade gastroenteropancreatic neuroendocrine neoplasms. BMC Cancer 2020, 20, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reubi, J.C.; Waser, B.; Lamberts, S.W.; Mengod, G. Somatostatin (SRIH) messenger ribonucleic acid expression in human neuroendocrine and brain tumors using in situ hybridization histochemistry: Comparison with SRIH receptor content. J. Clin. Endocrinol. Metab. 1993, 76, 642–647. [Google Scholar] [CrossRef]
- Neonakis, E.; Thomas, G.A.; Davies, H.G.; Wheeler, M.H.; Williams, E.D. Expression of calcitonin and somatostatin peptide and mRNA in medullary thyroid carcinoma. World J. Surg. 1994, 18, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Pacini, F.; Basolo, F.; Elisei, R.; Fugazzola, L.; Cola, A.; Pinchera, A. Medullary thyroid cancer. An immunohistochemical and humoral study using six separate antigens. Am. J. Clin. Pathol. 1991, 95, 300–308. [Google Scholar] [CrossRef]
- Kasprzak, A.; Zabel, M.; Surdyk-Zasada, J.; Seidel, J. Hybridocytochemical detection of mRNA for calcitonin, CGRP, somatostatin and NPY in cultured cells of medullary thyroid carcinoma using immunomax technique. Folia Histochem. Et Cytobiol. 1999, 37, 59–60. [Google Scholar] [PubMed]
- Gabalec, F.; Drastikova, M.; Cesak, T.; Netuka, D.; Masopust, V.; Machac, J.; Marek, J.; Cap, J.; Beranek, M. Dopamine 2 and somatostatin 1-5 receptors coexpression in clinically non-functioning pituitary adenomas. Physiol. Res. 2015, 64, 369–377. [Google Scholar] [CrossRef]
- Behling, F.; Honegger, J.; Skardelly, M.; Gepfner-Tuma, I.; Tabatabai, G.; Tatagiba, M.; Schittenhelm, J. High Expression of Somatostatin Receptors 2A, 3, and 5 in Corticotroph Pituitary Adenoma. Int. J. Endocrinol. 2018, 2018, 1763735. [Google Scholar] [CrossRef]
- Unger, N.; Serdiuk, I.; Sheu, S.Y.; Walz, M.K.; Schulz, S.; Saeger, W.; Schmid, K.W.; Mann, K.; Petersenn, S. Immunohistochemical localization of somatostatin receptor subtypes in benign and malignant adrenal tumours. Clin. Endocrinol. 2008, 68, 850–877. [Google Scholar] [CrossRef] [PubMed]
- Schulz, S.; Schmitt, J.; Quednow, C.; Roessner, A.; Weise, W. Immunohistochemical detection of somatostatin receptors in human ovarian tumors. Gynecol. Oncol. 2002, 84, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Pinzani, P.; Orlando, C.; Raggi, C.C.; Distante, V.; Valanzano, R.; Tricarico, C.; Maggi, M.; Serio, M.; Pazzagli, M. Type-2 somatostatin receptor mRNA levels in breast and colon cancer determined by a quantitative RT-PCR assay based on dual label fluorogenic probe and the TaqMan technology. Regul. Pept. 2001, 99, 79–86. [Google Scholar] [CrossRef]
- Manoochehri, M.; Wu, Y.; Giese, N.A.; Strobel, O.; Kutschmann, S.; Haller, F.; Hoheisel, J.D.; Moskalev, E.A.; Hackert, T.; Bauer, A.S. SST gene hypermethylation acts as a pan-cancer marker for pancreatic ductal adenocarcinoma and multiple other tumors: Toward its use for blood-based diagnosis. Mol. Oncol. 2020, 14, 1252–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naitou, K.; Shiina, T.; Nakamori, H.; Sano, Y.; Shimaoka, H.; Shimizu, Y. Colokinetic effect of somatostatin in the spinal defecation center in rats. J. Physiol. Sci. 2018, 68, 243–251. [Google Scholar] [CrossRef] [PubMed]
- De Fontgalland, D.; Wattchow, D.A.; Costa, M.; Brookes, S.J. Immunohistochemical characterization of the innervation of human colonic mesenteric and submucosal blood vessels. Neurogastroenterol. Motil. 2008, 20, 1212–1226. [Google Scholar] [CrossRef]
- Warhurst, G.; Higgs, N.B.; Fakhoury, H.; Warhurst, A.C.; Garde, J.; Coy, D.H. Somatostatin receptor subtype 2 mediates somatostatin inhibition of ion secretion in rat distal colon. Gastroenterology 1996, 111, 325–333. [Google Scholar] [CrossRef]
- Warhurst, G.; Barbezat, G.O.; Higgs, N.B.; Reyl-Desmars, F.; Lewin, M.J.; Coy, D.H.; Ross, I.; Grigor, M.R. Expression of somatostatin receptor genes and their role in inhibiting Cl- secretion in HT-29cl.19A colonocytes. Am. J. Physiol. 1995, 269, G729–G736. [Google Scholar] [CrossRef]
- Corleto, V.D. Somatostatin and the gastrointestinal tract. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 63–68. [Google Scholar] [CrossRef]
- Song, S.; Li, X.; Geng, C.; Li, Y.; Wang, C. Somatostatin stimulates colonic MUC2 expression through SSTR5-Notch-Hes1 signaling pathway. Biochem. Biophys. Res. Commun. 2020, 521, 1070–1076. [Google Scholar] [CrossRef]
- Van Bergeijk, J.D.; Wilson, J.H. Somatostatin in inflammatory bowel disease. Mediat. Inflamm. 1997, 6, 303–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Salhy, M.; Solomon, T.; Hausken, T.; Gilja, O.H.; Hatlebakk, J.G. Gastrointestinal neuroendocrine peptides/amines in inflammatory bowel disease. World J. Gastroenterol. 2017, 23, 5068–5085. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.R.; Chang, D.K. Colorectal cancer in inflammatory bowel disease: The risk, pathogenesis, prevention and diagnosis. World J. Gastroenterol. 2014, 7, 9872–9881. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, A.; Adamek, A. The Neuropeptide System and Colorectal Cancer Liver Metastases: Mechanisms and Management. Int. J. Mol. Sci. 2020, 21, 3494. [Google Scholar] [CrossRef]
- Binimelis, J.; Webb, S.M.; Monés, J.; Serrano, J.; Casamitjana, R.; Elena, M.; Peinado, M.A.; Vilardell, F.; De Leiva, A. Circulating immunoreactive somatostatin in gastrointestinal diseases. Decrease after vagotomy and enhancement in active ulcerative colitis, irritable bowel syndrome, and duodenal ulcer. Scand. J. Gastroenterol. 1987, 22, 931–937. [Google Scholar] [CrossRef]
- Koch, T.R.; Carney, J.A.; Morris, V.A.; Go, V.L. Somatostatin in the idiopathic inflammatory bowel diseases. Dis. Colon Rectum 1988, 31, 198–203. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M.; Mazzawi, T.; Umezawa, K.; Gilja, O.H. Enteroendocrine cells, stem cells and differentiation progenitors in rats with TNBS-induced colitis. Int. J. Mol. Med. 2016, 38, 1743–1751. [Google Scholar] [CrossRef] [Green Version]
- El-Salhy, M.; Hatlebakk, J.G.; Gilja, O.H. Abnormalities in endocrine and immune cells are correlated in dextran-sulfate-sodium-induced colitis in rats. Mol. Med. Rep. 2017, 15, 12–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elson, C.O.; Sartor, R.B.; Tennyson, G.S.; Riddell, R.H. Experimental models of inflammatory bowel disease. Gastroenterology 1995, 109, 1344–1367. [Google Scholar] [CrossRef]
- Antoniou, E.; Margonis, G.A.; Angelou, A.; Pikouli, A.; Argiri, P.; Karavokyros, I.; Papalois, A.; Pikoulis, E. The TNBS-induced colitis animal model: An overview. Ann. Med. Surg. 2016, 11, 9–15. [Google Scholar] [CrossRef]
- Watanabe, T.; Kubota, Y.; Sawada, T.; Muto, T. Distribution and quantification of somatostatin in inflammatory disease. Dis. Colon Rectum 1992, 35, 488–494. [Google Scholar] [CrossRef]
- De Fontgalland, D.; Brookes, S.J.; Gibbins, I.; Sia, T.C.; Wattchow, D.A. The neurochemical changes in the innervation of human colonic mesenteric and submucosal blood vessels in ulcerative colitis and Crohn’s disease. Neurogastroenterol. Motil. 2014, 26, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Tertychnyi, A.S.; Akhrieva, K.M.; Zayratyants, O.V.; Selivanova, L.S.; Birg, T.M. Somatostatin receptor expression in the ileal mucosa of patients with inflammatory bowel disease. Arkh. Patol. 2016, 78, 19–24, In Russian. [Google Scholar] [CrossRef]
- Caruso, M.L.; Di Pinto, F.; Ignazzi, A.; Coletta, S.; Valentini, A.M.; Cavalcanti, E.; De Michele, F. Increased nerve twigs in small intestinal mucosa with programmed cell death-ligand 1 and somatostatin receptor type 2A expression in recurrent Crohn disease: A case report. Medicine 2018, 97, e13492. [Google Scholar] [CrossRef] [PubMed]
- De Bruïne, A.P.; Wiggers, T.; Beek, C.; Volovics, A.; von Meyenfeldt, M.; Arends, J.W.; Bosman, F.T. Endocrine cells in colorectal adenocarcinomas: Incidence, hormone profile and prognostic relevance. Int. J. Cancer 1993, 54, 765–771. [Google Scholar] [CrossRef]
- Seretis, E.C.; Agnantis, N.J.; Golematis, V.C.; Voloudakis-Balatzis, I.E. Electron immunocytochemical demonstration of serotonin, vasoactive intestinal polypeptide, bombesin, somatostatin and glucagon in mirror biopsies from primary colorectal adenocarcinoma. J. Exp. Clin. Cancer Res. 2004, 23, 477–484. [Google Scholar]
- Godlewski, J.; Kaleczyc, J. Somatostatin, substance P and calcitonin gene-related peptide-positive intramural nerve structures of the human large intestine affected by carcinoma. Folia Histochem. Et Cytobiol. 2010, 48, 475–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoda, T.; Ishikawa, E.; Sano, T.; Watanabe, K.; Ikegami, M. Histopathological and immunohistochemical study of neuroendocrine tumors of the rectum. Acta Pathol. 1984, 34, 1059–1077. [Google Scholar] [CrossRef]
- Federspiel, B.H.; Burke, A.P.; Sobin, L.H.; Shekitka, K.M. Rectal and colonic carcinoids. A clinicopathologic study of 84 cases. Cancer 1990, 65, 135–140. [Google Scholar] [CrossRef]
- Sereti, E.; Gavriil, A.; Agnantis, N.; Golematis, V.C.; Voloudakis-Baltatzis, I.E. Immunoelectron study of somatostatin, gastrin and glucagon in human colorectal adenocarcinomas and liver metastases. Anticancer Res. 2002, 22, 2117–2123. [Google Scholar] [PubMed]
- Buscail, L.; Saint-Laurent, N.; Chastre, E.; Vaillant, J.C.; Gespach, C.; Capella, G.; Kalthoff, H.; Lluis, F.; Vaysse, N.; Susini, C. Loss of sst2 somatostatin receptor gene expression in human pancreatic and colorectal cancer. Cancer Res. 1996, 56, 1823–1827. [Google Scholar] [PubMed]
- Laws, S.A.; Gough, A.C.; Evans, A.A.; Bains, M.A.; Primrose, J.N. Somatostatin receptor subtype mRNA expression in human colorectal cancer and normal colonic mucosae. Br. J. Cancer 1997, 75, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Juhlin, C.C.; Falhammar, H.; Kjellman, M.; Åhlén, J.; Welin, S.; Calissendorff, J. Highly proliferative anal neuroendocrine carcinoma: Molecular and clinical features of a rare, recurrent case in complete remission. BMC Gastroenterol. 2020, 20, 290. [Google Scholar] [CrossRef]
- Colucci, R.; Blandizzi, C.; Ghisu, N.; Florio, T.; Del Tacca, M. Somatostatin inhibits colon cancer cell growth through cyclooxygenase-2 downregulation. Br. J. Pharmacol. 2008, 155, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Hohla, F.; Buchholz, S.; Schally, A.V.; Krishan, A.; Rick, F.G.; Szalontay, L.; Papadia, A.; Halmos, G.; Koster, F.; Aigner, E.; et al. Targeted cytotoxic somatostatin analog AN-162 inhibits growth of human colon carcinomas and increases sensitivity of doxorubicin resistant murine leukemia cells. Cancer Lett. 2010, 294, 35–42. [Google Scholar] [CrossRef]
- Ayiomamitis, G.D.; Notas, G.; Zaravinos, A.; Drygiannakis, I.; Georgiadou, M.; Sfakianaki, O.; Mastrodimou, N.; Thermos, K.; Kouroumalis, E. Effects of octreotide and insulin on colon cancer cellular proliferation and correlation with hTERT activity. Oncoscience 2014, 1, 457–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björk, J.; Nilsson, J.; Hultcrantz, R.; Johansson, C. Growth-regulatory effects of sensory neuropeptides, epidermal growth factor, insulin, and somatostatin on the non-transformed intestinal epithelial cell line IEC-6 and the colon cancer cell line HT 29. Scand. J. Gastroenterol. 1993, 28, 879–884. [Google Scholar] [CrossRef]
- Chen, J.S.; Liang, Q.M.; Li, H.S.; Yang, J.; Wang, S.; Long, J.W. Octreotide inhibits growth of colonic cancer SW480 cells by modulating the Wnt/P-catenin pathway. Pharmazie 2009, 64, 126–131. [Google Scholar] [PubMed]
- Wang, S.; Bao, Z.; Liang, Q.M.; Long, J.W.; Xiao, Z.S.; Jiang, Z.J.; Liu, B.; Yang, J.; Long, Z.X. Octreotide stimulates somatostatin receptor-induced apoptosis of SW480 colon cancer cells by activation of glycogen synthase kinase-3β, A Wnt/β-catenin pathway modulator. Hepatogastroenterology 2013, 60, 1639–1646. [Google Scholar] [PubMed]
- Pobłocki, J.; Jasińska, A.; Syrenicz, A.; Andrysiak-Mamos, E.; Szczuko, M. The Neuroendocrine Neoplasms of the Digestive Tract: Diagnosis, Treatment and Nutrition. Nutrients 2020, 12, 1437. [Google Scholar] [CrossRef] [PubMed]
- Bonds, M.; Rocha, F.G. Neuroendocrine Tumors of the Pancreatobiliary and Gastrointestinal Tracts. Surg. Clin. N. Am. 2020, 100, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Nagtegaal, I.D.; Odze, R.D.; Klimstra, D.; Paradis, V.; Rugge, M.; Schirmacher, P.; Washington, K.M.; Carneiro, F.; Cree, I.A. WHO Classification of Tumours Editorial Board. The 2019 WHO classification of tumours of the digestive system. Histopathology 2020, 76, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Hrabe, J. Neuroendocrine Tumors of the Appendix, Colon, and Rectum. Surg. Oncol. Clin. N. Am. 2020, 29, 267–279. [Google Scholar] [CrossRef]
- Ahmed, M. Gastrointestinal neuroendocrine tumors in 2020. World J. Gastrointest. Oncol. 2020, 12, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Dasari, A. Novel therapeutics for patients with well-differentiated gastroenteropancreatic neuroendocrine tumors. Ther. Adv. Med. Oncol. 2021, 13, 17588359211018047. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zheng-Pywell, R.; Chen, H.A.; Bibb, J.A.; Chen, H.; Rose, J.B. Management of Gastrointestinal Neuroendocrine Tumors. Clin. Med. Insights Endocrinol. Diabetes 2019, 12, 1179551419884058. [Google Scholar] [CrossRef] [PubMed]
- Schimmack, S.; Svejda, B.; Lawrence, B.; Kidd, M.; Modlin, I.M. The diversity and commonalities of gastroenteropancreatic neuroendocrine tumors. Langenbecks Arch. Surg. 2011, 396, 273–298. [Google Scholar] [CrossRef]
- Sun, M.H. Neuroendocrine differentiation in sporadic CRC and hereditary nonpolyosis colorectal cancer. Dis. Markers 2004, 20, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Klöppel, G. Classification and pathology of gastroenteropancreatic neuroendocrine neoplasms. Endocr. Relat. Cancer 2011, 18 (Suppl. 1), S1–S16. [Google Scholar] [CrossRef] [PubMed]
- Basturk, O.; Yang, Z.; Tang, L.H.; Hruban, R.H.; Adsay, V.; McCall, C.M.; Krasinskas, A.M.; Jang, K.T.; Frankel, W.L.; Balci, S.; et al. The high-grade (WHO G3) pancreatic neuroendocrine tumor category is morphologically and biologically heterogenous and includes both well differentiated and poorly differentiated neoplasms. Am. J. Surg. Pathol. 2015, 39, 683–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramage, J.K.; Valle, J.W.; Nieveen van Dijkum, E.J.M.; Sundin, A.; Pascher, A.; Couvelard, A.; Kloeppel, G. the ENETS 2016 Munich Advisory Board Participants; ENETS 2016 Munich Advisory Board Participants. Colorectal Neuroendocrine Neoplasms: Areas of Unmet Need. Neuroendocrinology 2019, 108, 45–53. [Google Scholar] [CrossRef]
- Pellat, A.; Cottereau, A.S.; Terris, B.; Coriat, R. Neuroendocrine Carcinomas of the Digestive Tract: What Is New? Cancers 2021, 13, 3766. [Google Scholar] [CrossRef]
- Assarzadegan, N.; Montgomery, E. What is New in the 2019 World Health Organization (WHO) Classification of Tumors of the Digestive System: Review of Selected Updates on Neuroendocrine Neoplasms, Appendiceal Tumors, and Molecular Testing. Arch. Pathol. Lab. Med. 2021, 145, 664–677. [Google Scholar] [CrossRef] [Green Version]
- La Rosa, S.; Sessa, F.; Uccella, S. Mixed Neuroendocrine-Nonneuroendocrine Neoplasms (MiNENs): Unifying the Concept of a Heterogeneous Group of Neoplasms. Endocr. Pathol. 2016, 27, 284–311. [Google Scholar] [CrossRef]
- Manhas, J.; Bhattacharya, A.; Agrawal, S.K.; Gupta, B.; Das, P.; Deo, S.V.; Pal, S.; Sen, S. Characterization of cancer stem cells from different grades of human colorectal cancer. Tumour Biol. 2016, 37, 14069–14081. [Google Scholar] [CrossRef]
- Boman, B.M.; Fields, J.Z.; Bonham-Carter, O.; Runquist, O.A. Computer modeling implicates stem cell overproduction in colon cancer initiation. Cancer Res. 2001, 61, 8408–8411. [Google Scholar]
- Boman, B.M.; Fields, J.Z.; Cavanaugh, K.L.; Guetter, A.; Runquist, O.A. How dysregulated colonic crypt dynamics cause stem cell overpopulation and initiate colon cancer. Cancer Res. 2008, 68, 3304–3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleist, B.; Poetsch, M. Neuroendocrine differentiation: The mysterious fellow of colorectal cancer. World J. Gastroenterol. 2015, 21, 11740–11747. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, P.; Schönfelder, J.; Ahnert-Hilger, G.; Foss, H.D.; Heine, B.; Schindler, I.; Stein, H.; Berger, G.; Zeitz, M.; Scherübl, H. Expression of neuroendocrine markers: A signature of human undifferentiated carcinoma of the colon and rectum. Virchows Arch. 2002, 441, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Volante, M.; Marci, V.; Andrejevic-Blant, S.; Tavaglione, V.; Sculli, M.C.; Tampellini, M.; Papotti, M. Increased neuroendocrine cells in resected metastases compared to primary colorectal adenocarcinomas. Virchows Arch. 2010, 457, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Shia, J.; Tickoo, S.K.; Guillem, J.G.; Qin, J.; Nissan, A.; Hoos, A.; Stojadinovic, A.; Ruo, L.; Wong, W.D.; Paty, P.B.; et al. Increased endocrine cells in treated rectal adenocarcinomas: A possible reflection of endocrine differentiation in tumor cells induced by chemotherapy and radiotherapy. Am. J. Surg. Pathol. 2002, 26, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Dasari, A.; Shen, C.; Halperin, D.; Zhao, B.; Zhou, S.; Xu, Y.; Shih, T.; Yao, J.C. Trends in the Incidence, Prevalence, and Survival Outcomes in Patients with Neuroendocrine Tumors in the United States. JAMA Oncol. 2017, 3, 1335–1342. [Google Scholar] [CrossRef]
- Bertani, E.; Ravizza, D.; Milione, M.; Massironi, S.; Grana, C.M.; Zerini, D.; Piccioli, A.N.; Spinoglio, G.; Fazio, N. Neuroendocrine neoplasms of rectum: A management update. Cancer Treat. Rev. 2018, 66, 45–55. [Google Scholar] [CrossRef]
- Modlin, I.M.; Lye, K.D.; Kidd, M. A 5-decade analysis of 13,715 carcinoid tumors. Cancer 2003, 97, 934–959. [Google Scholar] [CrossRef] [PubMed]
- Byrne, R.M.; Pommier, R.F. Small Bowel and Colorectal Carcinoids. Clin. Colon Rectal Surg. 2018, 31, 301–308. [Google Scholar] [CrossRef]
- Gaspar, R.; Santos-Antunes, J.; Marques, M.; Liberal, R.; Melo, D.; Pereira, P.; Lopes, S.; Macedo, G. Mixed Adenoneuroendocrine Tumor of the Rectum in an Ulcerative Colitis Patient. GE Port. J. Gastroenterol. 2019, 26, 125–127. [Google Scholar] [CrossRef]
- Takizawa, N.; Ohishi, Y.; Hirahashi, M.; Takahashi, S.; Nakamura, K.; Tanaka, M.; Oki, E.; Takayanagi, R.; Oda, Y. Molecular characteristics of colorectal neuroendocrine carcinoma; similarities with adenocarcinoma rather than neuroendocrine tumor. Hum. Pathol. 2015, 46, 1890–1900. [Google Scholar] [CrossRef]
- Frizziero, M.; Wang, X.; Chakrabarty, B.; Childs, A.; Luong, T.V.; Walter, T.; Khan, M.S.; Morgan, M.; Christian, A.; Elshafie, M.; et al. Retrospective study on mixed neuroendocrine non-neuroendocrine neoplasms from five European centres. World J. Gastroenterol. 2019, 25, 5991–6005. [Google Scholar] [CrossRef]
- Watanabe, J.; Suwa, Y.; Ota, M.; Ishibe, A.; Masui, H.; Nagahori, K.; Tsuura, Y.; Endo, I. Clinicopathological and Prognostic Evaluations of Mixed Adenoneuroendocrine Carcinoma of the Colon and Rectum: A Case-Matched Study. Dis. Colon Rectum 2016, 59, 1160–1167. [Google Scholar] [CrossRef]
- Li, Y.; Yau, A.; Schaeffer, D.; Magliocco, A.; Gui, X.; Urbanski, S.; Waghray, R.; Owen, D.; Gao, Z.H. Colorectal glandular-neuroendocrine mixed tumor: Pathologic spectrum and clinical implications. Am. J. Surg. Pathol. 2011, 35, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Sheng, H.; Wei, X.; Mao, M.; He, J.; Luo, T.; Lu, S.; Zhou, L.; Huang, Z.; Yang, A. Adenocarcinoma with mixed subtypes is a rare but aggressive histologic subtype in colorectal cancer. BMC Cancer 2019, 19, 1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Kaneko, M.; Nozawa, H.; Emoto, S.; Murono, K.; Otani, K.; Sasaki, K.; Nishikawa, T.; Kiyomatsu, T.; Hata, K.; et al. Diagnosis, Assessment, and Therapeutic Strategy for Colorectal Mixed Adenoneuroendocrine Carcinoma. Neuroendocrinology 2017, 105, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Boman, B.M.; Guetter, A.; Boman, R.M.; Runquist, O.A. Autocatalytic Tissue Polymerization Reaction Mechanism in Colorectal Cancer Development and Growth. Cancers 2020, 12, 460. [Google Scholar] [CrossRef] [Green Version]
- Mori, Y.; Cai, K.; Cheng, Y.; Wang, S.; Paun, B.; Hamilton, J.P.; Jin, Z.; Sato, F.; Berki, A.T.; Kan, T.; et al. A genome-wide search identifies epigenetic silencing of somatostatin, tachykinin-1, and 5 other genes in colon cancer. Gastroenterology 2006, 131, 797–808. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chew, M.H.; Tham, C.K.; Tang, C.L.; Ong, S.Y.; Zhao, Y. Methylation of serum SST gene is an independent prognostic marker in colorectal cancer. Am. J. Cancer Res. 2016, 6, 2098–2108. [Google Scholar]
- Liu, J.; Li, H.; Sun, L.; Wang, Z.; Xing, C.; Yuan, Y. Aberrantly methylated-differentially expressed genes and pathways in colorectal cancer. Cancer Cell Int. 2017, 17, 75. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, C.; Bi, X.; Zhou, C.; Huang, T.; Ni, C.; Yang, P.; Chen, S.; Ye, M.; Duan, S. DNA methylation of CMTM3, SSTR2, and MDFI genes in colorectal cancer. Gene 2017, 630, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Seifert, J.K.; Görges, R.; Bockisch, A.; Junginger, T. 111-Indium-DTPA-Octreotid-Szintigraphie bei kolorektalen Lebermetastasen [111-indium DTPA octreotide scintigraphy in colorectal liver metastases]. Langenbecks Arch. Chir. 1997, 382, 332–336. (In German) [Google Scholar] [PubMed]
- Binderup, T.; Knigge, U.; Mellon Mogensen, A.; Palnaes Hansen, C.; Kjaer, A. Quantitative gene expression of somatostatin receptors and noradrenaline transporter underlying scintigraphic results in patients with neuroendocrine tumors. Neuroendocrinology 2008, 87, 223–232. [Google Scholar] [CrossRef]
- Eychenne, R.; Bouvry, C.; Bourgeois, M.; Loyer, P.; Benoist, E.; Lepareur, N. Overview of Radiolabeled Somatostatin Analogs for Cancer Imaging and Therapy. Molecules 2020, 25, 4012. [Google Scholar] [CrossRef]
- Diamantopoulos, L.N.; Laskaratos, F.M.; Kalligeros, M.; Shah, R.; Navalkissoor, S.; Gnanasegaran, G.; Banks, J.; Smith, J.; Jacobs, B.; Galanopoulos, M.; et al. Antiproliferative Effect of Above-Label Doses of Somatostatin Analogs for the Management of Gastroenteropancreatic Neuroendocrine Tumors. Neuroendocrinology 2021, 111, 650–659. [Google Scholar] [CrossRef]
- Duclos, V.; Iep, A.; Gomez, L.; Goldfarb, L.; Besson, F.L. PET Molecular Imaging: A Holistic Review of Current Practice and Emerging Perspectives for Diagnosis, Therapeutic Evaluation and Prognosis in Clinical Oncology. Int. J. Mol. Sci. 2021, 22, 4159. [Google Scholar] [CrossRef]
- Krenning, E.P.; Bakker, W.H.; Breeman, W.A.; Koper, J.W.; Kooij, P.P.; Ausema, L.; Lameris, J.S.; Reubi, J.C.; Lamberts, S.W. Localisation of endocrine-related tumours with radioiodinated analogue of somatostatin. Lancet 1989, 1, 242–244. [Google Scholar] [CrossRef]
- Krenning, E.P.; Kwekkeboom, D.J.; Bakker, W.H.; Breeman, W.A.; Kooij, P.P.; Oei, H.Y.; van Hagen, M.; Postema, P.T.; de Jong, M.; Reubi, J.C.; et al. Somatostatin receptor scintigraphy with [111In-DTPA-D-Phe1]- and [123I-Tyr3]-octreotide: The Rotterdam experience with more than 1000 patients. Eur. J. Nucl. Med. 1993, 20, 716–731. [Google Scholar] [CrossRef] [Green Version]
- De Herder, W.W.; Kwekkeboom, D.J.; Valkema, R.; Feelders, R.A.; van Aken, M.O.; Lamberts, S.W.; van der Lely, A.J.; Krenning, E.P. Neuroendocrine tumors and somatostatin: Imaging techniques. J. Endocrinol. Investig. 2005, 28, 132–136. [Google Scholar]
- De Herder, W.W.; Kwekkeboom, D.J.; Feelders, R.A.; van Aken, M.O.; Lamberts, S.W.; van der Lely, A.J.; Krenning, E.P. Somatostatin receptor imaging for neuroendocrine tumors. Pituitary 2006, 9, 243–248. [Google Scholar] [CrossRef] [PubMed]
- De Herder, W.W. GEP-NETS update: Functional localisation and scintigraphy in neuroendocrine tumours of the gastrointestinal tract and pancreas (GEP-NETs). Eur. J. Endocrinol. 2014, 170, R173–R183. [Google Scholar] [CrossRef] [Green Version]
- Kwekkeboom, D.J.; Kam, B.L.; van Essen, M.; Teunissen, J.J.; van Eijck, C.H.; Valkema, R.; de Jong, M.; de Herder, W.W.; Krenning, E.P. Somatostatin-receptor-based imaging and therapy of gastroenteropancreatic neuroendocrine tumors. Endocr. Relat. Cancer 2010, 17, R53–R73. [Google Scholar] [CrossRef] [Green Version]
- Olsen, I.H.; Langer, S.W.; Federspiel, B.H.; Oxbøl, J.; Loft, A.; Berthelsen, A.K.; Mortensen, J.; Oturai, P.; Knigge, U.; Kjær, A. (68)Ga-DOTATOC PET and gene expression profile in patients with neuroendocrine carcinomas: Strong correlation between PET tracer uptake and gene expression of somatostatin receptor subtype 2. Am. J. Nucl. Med. Mol. Imaging 2016, 6, 59–72. [Google Scholar] [PubMed]
- Lodge, M.A.; Solnes, L.B.; Chaudhry, M.A.; Wahl, R.L. Prospective Within-Patient Assessment of the Impact of an Unlabeled Octreotide Pre-dose on the Biodistribution and Tumor Uptake of 68Ga DOTATOC as Assessed by Dynamic Whole-body PET in Patients with Neuroendocrine Tumors: Implications for Diagnosis and Therapy. Mol. Imaging Biol. 2021, 23, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.L.; Pavlakis, N.; Schembri, G.P.; Bernard, E.J.; Hsiao, E.; Hayes, A.; Barnes, T.; Diakos, C.; Khasraw, M.; Samra, J.; et al. Dual Somatostatin Receptor/FDG PET/CT Imaging in Metastatic Neuroendocrine Tumours: Proposal for a Novel Grading Scheme with Prognostic Significance. Theranostics 2017, 7, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Yu, J.; Li, J.; Shen, L.; Li, N.; Zhu, H.; Zhai, S.; Zhang, Y.; Yang, Z.; Lu, M. Clinical and Prognostic Value of PET/CT Imaging with Combination of 68Ga-DOTATATE and 18F-FDG in Gastroenteropancreatic Neuroendocrine Neoplasms. Contrast Media Mol. Imaging 2018, 2018, 2340389. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, M.; Muehllechner, P.; Decristoforo, C.; von Guggenberg, E.; Kendler, D.; Prommegger, R.; Profanter, C.; Moncayo, R.; Virgolini, I. 99mTc-EDDA/HYNIC-Tyr(3)-octreotide for staging and follow-up of patients with neuroendocrine gastro-entero-pancreatic tumors. Q. J. Nucl. Med. Mol. Imaging 2005, 49, 237–244. [Google Scholar] [PubMed]
- Cwikla, J.B.; Mikolajczak, R.; Pawlak, D.; Buscombe, J.R.; Nasierowska-Guttmejer, A.; Bator, A.; Maecke, H.R.; Walecki, J. Initial direct comparison of 99mTc-TOC and 99mTc-TATE in identifying sites of disease in patients with proven GEP NETs. J. Nucl. Med. 2008, 49, 1060–1065. [Google Scholar] [CrossRef] [Green Version]
- Artiko, V.; Afgan, A.; Petrović, J.; Radović, B.; Petrović, N.; Vlajković, M.; Šobić-Šaranović, D.; Obradović, V. Evaluation of neuroendocrine tumors with 99mTc-EDDA/HYNIC TOC. Nucl. Med. Rev. Cent. East Eur. 2016, 19, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Liepe, K.; Becker, A. 99mTc-Hynic-TOC imaging in the diagnostic of neuroendocrine tumors. World J. Nucl. Med. 2018, 17, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Rufini, V.; Calcagni, M.L.; Baum, R.P. Imaging of neuroendocrine tumors. Semin. Nucl. Med. 2006, 36, 228–247. [Google Scholar] [CrossRef] [PubMed]
- Banzo, J.; Vidal-Sicat, S.; Prats, E.; Galofré, G.; Razola, P.; Mañé, S.; Ubieto, M.A.; Abós, M.D. In-111 DTPA octreotide scintigraphy and intraoperative gamma probe detection in the diagnosis and treatment of residual lymph node metastases of a rectal carcinoid tumor. Clin. Nucl. Med. 2005, 30, 308–311. [Google Scholar] [CrossRef] [PubMed]
- Kostenich, G.; Oron-Herman, M.; Kimel, S.; Livnah, N.; Tsarfaty, I.; Orenstein, A. Diagnostic targeting of colon cancer using a novel fluorescent somatostatin conjugate in a mouse xenograft model. Int. J. Cancer 2008, 122, 2044–2049. [Google Scholar] [CrossRef]
- Strosberg, J.; El-Haddad, G.; Wolin, E.; Hendifar, A.; Yao, J.; Chasen, B.; Mittra, E.; Kunz, P.L.; Kulke, M.H.; Jacene, H.; et al. Phase 3 Trial of 177Lu-Dotatate for Midgut Neuroendocrine Tumors. N. Engl. J. Med. 2017, 376, 125–135. [Google Scholar] [CrossRef]
- Mansi, R.; Nicolas, G.P.; Del Pozzo, L.; Abid, K.A.; Grouzmann, E.; Fani, M. Evaluation of a New 177Lu-Labeled Somatostatin Analog for the Treatment of Tumors Expressing Somatostatin Receptor Subtypes 2 and 5. Molecules 2020, 25, 4155. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Lu, C.; Yang, Y.; Shi, X.; Wang, H.; Yang, N.; Yang, K.; Zhang, X. Current Status and Trends in Peptide Receptor Radionuclide Therapy in the Past 20 Years (2000–2019): A Bibliometric Study. Front. Pharmacol. 2021, 12, 624534. [Google Scholar] [CrossRef]
- Ayo, A.; Laakkonen, P. Peptide-Based Strategies for Targeted Tumor Treatment and Imaging. Pharmaceutics 2021, 13, 481. [Google Scholar] [CrossRef]
- Kwekkeboom, D.J.; de Herder, W.W.; Kam, B.L.; van Eijck, C.H.; van Essen, M.; Kooij, P.P.; Feelders, R.A.; van Aken, M.O.; Krenning, E.P. Treatment with the radiolabeled somatostatin analog [177 Lu-DOTA 0,Tyr3]octreotate: Toxicity, efficacy, and survival. J. Clin. Oncol. 2008, 26, 2124–2130. [Google Scholar] [CrossRef] [Green Version]
- Weckbecker, G.; Raulf, F.; Stolz, B.; Bruns, C. Somatostatin analogs for diagnosis and treatment of cancer. Pharmacol. Ther. 1993, 60, 245–264. [Google Scholar] [CrossRef]
- Yordanova, A.; Biersack, H.J.; Ahmadzadehfar, H. Advances in Molecular Imaging and Radionuclide Therapy of Neuroendocrine Tumors. J. Clin. Med. 2020, 9, 3679. [Google Scholar] [CrossRef]
- Pavel, M.; de Herder, W.W. ENETS Consensus Guidelines for the Standard of Care in Neuroendocrine Tumors. Neuroendocrinology 2017, 105, 193–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinke, A.; Müller, H.H.; Schade-Brittinger, C.; Klose, K.J.; Barth, P.; Wied, M.; Mayer, C.; Aminossadati, B.; Pape, U.F.; Bläker, M.; et al. Placebo-controlled, double-blind, prospective, randomized study on the effect of octreotide LAR in the control of tumor growth in patients with metastatic neuroendocrine midgut tumors: A report from the PROMID Study Group. J. Clin. Oncol. 2009, 27, 4656–4663. [Google Scholar] [CrossRef] [PubMed]
- Oberg, K. Cancer: Antitumor effects of octreotide LAR, a somatostatin analog. Nat. Rev. Endocrinol. 2010, 6, 188–189. [Google Scholar] [CrossRef]
- Caplin, M.E.; Pavel, M.; Ćwikła, J.B.; Phan, A.T.; Raderer, M.; Sedláčková, E.; Cadiot, G.; Wolin, E.M.; Capdevila, J.; Wall, L.; et al. Anti-tumour effects of lanreotide for pancreatic and intestinal neuroendocrine tumours: The CLARINET open-label extension study. Endocr. Relat. Cancer 2016, 23, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Pusceddu, S.; Buzzoni, R.; de Braud, F. The underestimated role of somatostatin analogs in the NETTER-1 trial. Future Oncol. 2017, 13, 1287–1289. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Jacobson, O.; Niu, G.; Kiesewetter, D.O.; Wang, Z.; Zhu, G.; Ma, Y.; Liu, G.; Chen, X. Evans Blue Attachment Enhances Somatostatin Receptor Subtype-2 Imaging and Radiotherapy. Theranostics 2018, 8, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Wang, S.; Han, S.; Kim, S.H.; Kusnetzow, A.K.; Nguyen, J.; Rico-Bautista, E.; Tan, H.; Betz, S.F.; Scott Struthers, R.; et al. Discovery of nonpeptide 3,4-dihydroquinazoline-4-carboxamides as potent and selective sst2 agonists. Bioorg. Med. Chem. Lett. 2020, 30, 127391. [Google Scholar] [CrossRef]
- Reubi, J.C.; Waser, B.; Mäcke, H.; Rivier, J. Highly Increased 125I-JR11 Antagonist Binding In Vitro Reveals Novel Indications for sst2 Targeting in Human Cancers. J. Nucl. Med. 2017, 58, 300–306. [Google Scholar] [CrossRef] [Green Version]
- Kong, G.; Grozinsky-Glasberg, S.; Hofman, M.S.; Akhurst, T.; Meirovitz, A.; Maimon, O.; Krausz, Y.; Godefroy, J.; Michael, M.; Gross, D.J.; et al. Highly favourable outcomes with peptide receptor radionuclide therapy (PRRT) for metastatic rectal neuroendocrine neoplasia (NEN). Eur. J. Nucl. Med. Mol. Imaging 2019, 46, 718–727. [Google Scholar] [CrossRef]
- Dasgupta, P. Somatostatin analogues: Multiple roles in cellular proliferation, neoplasia, and angiogenesis. Pharmacol. Ther. 2004, 102, 61–85. [Google Scholar] [CrossRef]
- Abdellatif, A.A.H.; Ibrahim, M.A.; Amin, M.A.; Maswadeh, H.; Alwehaibi, M.N.; Al-Harbi, S.N.; Alharbi, Z.A.; Mohammed, H.A.; Mehany, A.B.M.; Saleem, I. Cetuximab Conjugated with Octreotide and Entrapped Calcium Alginate-beads for Targeting Somatostatin Receptors. Sci. Rep. 2020, 10, 4736. [Google Scholar] [CrossRef] [PubMed]
- Szepeshazi, K.; Schally, A.V.; Halmos, G.; Armatis, P.; Hebert, F.; Sun, B.; Feil, A.; Kiaris, H.; Nagy, A. Targeted cytotoxic somatostatin analogue AN-238 inhibits somatostatin receptor-positive experimental colon cancers independently of their p53 status. Cancer Res. 2002, 62, 781–788. [Google Scholar]
- Nguyen, K.; Parry, J.J.; Rogers, B.E.; Anderson, C.J. Evaluation of copper-64-labeled somatostatin agonists and antagonist in SSTr2-transfected cell lines that are positive and negative for p53: Implications for cancer therapy. Nucl. Med. Biol. 2012, 39, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behnammanesh, H.; Erfani, M.; Hajiramezanali, M.; Jokar, S.; Geramifar, P.; Sabzevari, O.; Amini, M.; Mazidi, S.M.; Beiki, D. Preclinical study of a new 177Lu-labeled somatostatin receptor antagonist in HT-29 human colorectal cancer cells. Asia Ocean J. Nucl. Med. Biol. 2020, 8, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Emery, P.T.; Higgs, N.B.; Warhurst, A.C.; Carlson, G.L.; Warhurst, G. Anti-secretory properties of non-peptide somatostatin receptor agonists in isolated rat colon: Luminal activity and possible interaction with P-glycoprotein. Br. J. Pharmacol. 2002, 135, 1443–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, O.L.; Mekhael, M.M.; Ahmed, A.; Cuthbertson, D.J.; Pritchard, D.M. Frequency and Causes of False-Positive Elevated Plasma Concentrations of Fasting Gut Hormones in a Specialist Neuroendocrine Tumor Center. Front. Endocrinol. 2020, 11, 606264. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| SRIF System Member | Gene Location and Size (kb) | No. of Transcripts | Protein | Mechanisms of Action/Biological Effects in GI Tract | Ref. |
|---|---|---|---|---|---|
| SST | SS1 gene, 3q27.3; size: 1483 bases | pre-mRNA contains an intron flanked by two exons; 1 transcript (splice variant) and 262 orthologues | preprosomatostatin (116 AA) and prosomatostatin (92 AA), which in turn is C-terminally processed to generate the cyclic peptides SST-14 AA and SST-28 AA (N-terminal extension); m.m. 12736 Da; q.s.: nd | (+)SST1–5 (GPCRs family); (−)several hormones secretion; (−)exocrine secretion; (−)gallbladder contraction; (−)small intestinal segmentation; (−)gastric emptying;regulation of gastric acid, digestive enzymes, bile, colonic fluid, and motility; local immunomodulatory actions | [12,28,32,37,41,60,61,62,63,65,66,75,88,90,91] |
| CST | CORT gene; 1p36.3–1p36.2; size: 1968 bases | cleavage of preproCST (112 AA) gives rise to multiple mature products, CST-14, CST-17 and CST-29; m.m. 11532 Da, q.s.: nd | CST may activate SST1–5 and GPCRs other than SST1–5; share several biologic properties with SST | [12] | |
| SST1 | 14q13; Size: 5164 bases | an intronless gene | 391 AA; m.m. 42686 Da; q.s.: interacts with SKB1 | binds both form of SST, with slightly higher affinity for SST-14; (−)proliferation; (−)secretion; (−)intestinal Cl− secretion | [12,27,62,106,107] |
| SST2 | 17q25.1; size: 11,624 bases | consists of two exons | 369 AA; spliced into SST2A and 2B; human tissues exclusively contain the unspliced SST2A variant; m.m. 41333Da; q.s.: homo- and heterodimer with SST3 and SST5 | binds both forms of SST, with slightly higher affinity for SST-14; (−)proliferation; (−)secretion; (+)apoptosis; (−)exocrine secretion: gastric acid and intestinal Cl− secretion | [12,27,62,106,107] |
| SST3 | 22q13.1; size: 16,215 bases | spreads over eight exons | 418 AA; m.m. 45847 Da; q.s.: homodimer and heterodimer with SST2 | binds both forms of SST, with slightly higher affinity for SST-14; (−)proliferation; (−)secretion; (+)apoptosis; gastric and intestinal relaxation | [12,27,62,102,103,106,107] |
| SST4 | 20p11.2; size: 3926 bases | 388 AA; m.m. 42003 Da; q.s.: nd | binds both forms of SST, with slightly higher affinity for SST-14; (−)proliferation; (−)secretion | [12,27,62,104,106,107] | |
| SST5 | 16p13.3; size: 8708 bases | 364 AA; m.m. 39202 Da; q.s.: heterodimer with SST2 | binds both forms of SST, with a 10-fold higher affinity for SST-28; (−)proliferation; (−)secretion, e.g., amylase secretion; (−)colonic contraction | [12,27,62,104,106,107] |
| SRIF System Member | Cellular Localization | The Main Results | Material and Methods | Ref. |
|---|---|---|---|---|
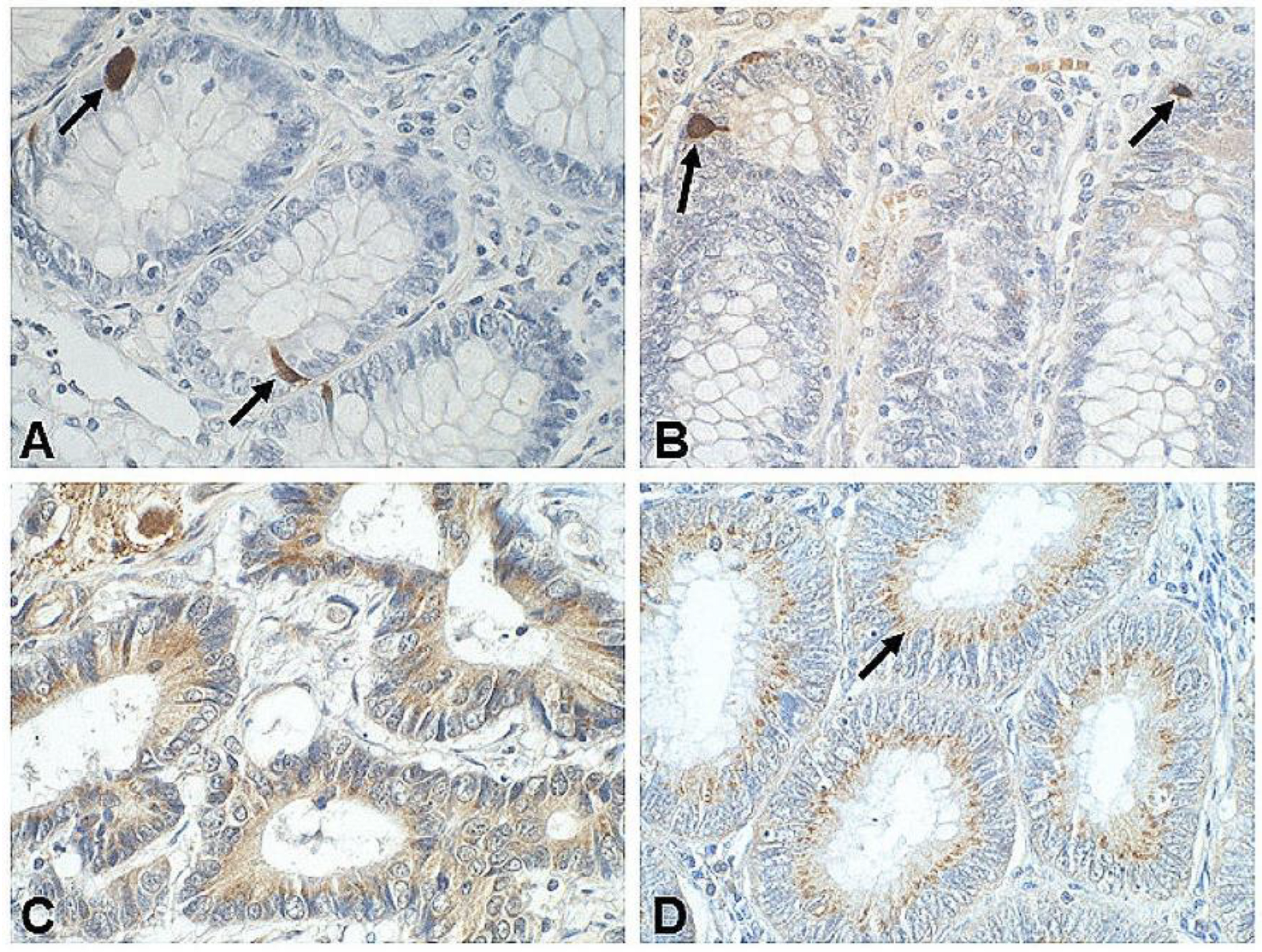
| SST | tumor cells | (+)in 18/23 tumors | n = 32 NETs of rectum (n = 27 as typical carcinoids); IHC | [163] |
| tumor cells | (+)in 3% tumors | n = 3 carcinoids of the distal sigmoid colon, and n = 81 of the rectum; IHC | [164] | |
| tumor endocrine cells | (+)in 10% of chromogranin A-positive CRC | n = 350 CRC; IHC | [160] | |
| carcinoma cells | (+)in 82% CRC vs. 90% tumor-neighboring mucosa | n = 100 CRC (39 colonic and 61 rectal), and surrounding mucosa; IHC | [8] | |
| tumor cells; normal colonocytes | (+)in 84.6% CRC vs. 88.5% tumor-neighboring mucosa | n = 53 advanced CRC, and tumor-neighboring mucosa; IHC | [9] | |
| tumor EECs in glandular structures | (+)in 4/57 CRC; in 2 cases co-expressed with serotonin and glucagon | n = 57 CRC; IHC | [55] | |
| carcinoma cells | ↑in well-differentiated vs. poorly differentiated tumors; ♣(↓expression) | n = 35 CRC; n = 25 LM; IHC | [165] | |
| D cells | low expression | n = 90 mirror biopsies of CRC; iEM | [161] | |
| carcinoma cells | (−)expression | n = 1 mucinous CRC with signet-ring cells (stage D); IHC; iEM | [57] | |
| ENS structures | frequency of (+)neurons/fibers in the intact vs. CRC areasNS | n = 15 CRC; IHC; IF | [162] | |
| D cells; tumor cells | ↓mRNA in CRC vs. C (adults); ↑ratio of (+)cells in C (children) vs. CRC | n = 34 CRC, n = 6 C (children), n = 41 C (adults) (TMA); n = 13 CRC, n = 14 C (children), n = 20 (adults) (IHC); n = 12 CRC, n = 12 C (children), n = 12 C (adults) (RT-PCR) | [18] | |
| (+)in all the C; (−)in matching CRC samples | 5 samples of C, and matched CRC; RT-PCR | [10] | ||
| SST1 | heterogeneously expressed in both C and CRC; (+) in liver M | n = 47 CRC and C; RT-PCR | [166] | |
| tumor cells; normal colonocytes | expressed infrequently | n = 32 pairs of CRC and C; RT-PCR; ISH | [167] | |
| (+)mRNA frequently expressed in C and CRC | CRC and C; RT-PCR | [115] | ||
| immune cells of the lp close to the tumor; stromal cells; epithelial cells | (+)mRNA expressed in C and CRC | CRC and C; ISH; image analysis system | [116] | |
| tumor cells; normal colonocytes | the predominant subtype in CRC and C; ♦↑(Dukes stage); ♦↑(lymph node M) | n = 127 CRC and n = 40 C; IHC | [118] | |
| (+)in all the C; (+)in 3/5 CRC | 5 samples of C and matched CRC tumor tissue; RT-PCR | [10] | ||
| SST2 | (+)mRNA (50% in Dukes stage B, and 20% in Dukes stage C), (−)in stage D CRC | n = 47 CRC and controls, RT-PCR | [166] | |
| tumor cells; normal colon crypt cells | (+)in nearly 90% CRC and C | n = 32 pairs of CRC and C; RT-PCR; ISH | [167] | |
| (+)mRNA frequently expressed in C and CRC; no loss in advanced CRC stages | CRC and C; RT-PCR | [115] | ||
| tumor cells; immune cells; epithelial cells; stromal cells | low mRNA expression in C and CRC | CRC and C; ISH; image analysis system | [116] | |
| (+)in 100% tumors; CRC vs. CNS; loss of SST2 mRNA in patient’s tumor with ↑CEA level vs. patients with low CEA; normal/tumor mRNA ratio inversely related to CEA levels | n = 26 CRC and corresponding C; qRT-PCR | [138] | ||
| (+)in 100% tumors; CRC vs. CNS; loss of SST2 mRNA in tumor in patients with ↑CEA level vs. C | n = 100 CRC and corresponding C; RT-PCR; ISH | [117] | ||
| ↑mRNA; # (↑CRD and shorter DFS) | qRT-PCR | [54] | ||
| tumor cells; normal colon crypt cells | the second subtype in C and CRC; ↑in moderately to well vs. poorly differentiated CRC; ↓in the ulcerative type of CRC | n = 127 CRC and n = 40 C; IHC | [118] | |
| tumor cells | ↑in lower-grade and rectum located tumors; ♦, #, (+) (longer survival rate); ♦,↓liver M | n = 81 CRC; IHC | [119] | |
| tumor cells | (+)in 100% tumors | n = 3 rectal NETs; IHC | [120] | |
| (−)SST2A | n = 19 colorectal MANECs and n = 8 CRC poorly differentiated NETs; IHC | [52] | ||
| tumor cells | (+)expression | n = 1 small cell anal NEC; IHC | [168] | |
| SST3 | rarely or not expressed | n = 47 CRC and C; RT-PCR | [166] | |
| tumor cells; normal colonocytes | expressed infrequently | n = 32 pairs of CRC and C; RT-PCR; ISH | [167] | |
| tumor cells; transformed epithelial cells; stromal cells | low mRNA expression in C and CRC | CRC and C; ISH; image analysis system | [116] | |
| tumor cells; normal colon crypt cells | ♦,↓(Dukes stage) | n = 127 CRC and n = 40 C; IHC | [118] | |
| SST4 | heterogeneously expressed in C and CRC; (+) in liver M | n = 47 CRC and C; RT-PCR | [166] | |
| tumor cells; normal colonocytes | expressed infrequently | n = 32 pairs of CRC and C; RT-PCR; ISH | [167] | |
| stromal cells; immune cells in CT close to the tumor; epithelial cells | low mRNA expression in both CRC and C | CRC and C; ISH; image analysis system | [116] | |
| tumor cells; normal colonocytes | ↑in moderately to well vs. poorly differentiated CRC; ↑frequency in tumor cells (18.9%) vs. C (2.5%) | n = 127 CRC and n = 40 C; IHC | [118] | |
| SST5 | heterogeneously expressed in C and CRC | n = 47 CRC and C; RT-PCR | [166] | |
| tumor cells; normal colonocytes | (+)in 46% CRC and 45% C; (+)in 75% (Dukes A and B stage) vs. 31% (Dukes C and D); ↓in M (11%) vs. all tumor samples (56%) | n = 32 pairs of CRC and C; RT-PCR; ISH | [167] | |
| (+)mRNA very frequently expressed in C and CRC; no loss in advanced CRC stages; ↑frequency in the left CRC vs. C | CRC and C; RT-PCR | [115] | ||
| tumor cells; epithelial cells; stromal cells | (+)the mRNA was the predominant subtype expressed in C and CRC; ↑in CRC vs. C | CRC and C; ISH; image analysis system | [116] | |
| tumor cells; normal colonocytes | the second subtype in C and CRC; ↑in moderately to well vs. poorly differentiated CRC; ♦,↓(Dukes stage) | n = 127 CRC and n = 40 C; IHC | [118] | |
| tumor cells | (+)(longer survival rate); ↓, #, liver M | n = 81 CRC; IHC | [119] | |
| tumor cells | (+)in 66.6% tumors | n = 3 rectal NETs; IHC | [120] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasprzak, A. Somatostatin and Its Receptor System in Colorectal Cancer. Biomedicines 2021, 9, 1743. https://doi.org/10.3390/biomedicines9111743
Kasprzak A. Somatostatin and Its Receptor System in Colorectal Cancer. Biomedicines. 2021; 9(11):1743. https://doi.org/10.3390/biomedicines9111743
Chicago/Turabian StyleKasprzak, Aldona. 2021. "Somatostatin and Its Receptor System in Colorectal Cancer" Biomedicines 9, no. 11: 1743. https://doi.org/10.3390/biomedicines9111743
APA StyleKasprzak, A. (2021). Somatostatin and Its Receptor System in Colorectal Cancer. Biomedicines, 9(11), 1743. https://doi.org/10.3390/biomedicines9111743