
Bisphenol A and Its Analogues Deteriorate the Hormones Physiological Function of the Male Reproductive System: A Mini-Review




Abstract
:1. Introduction
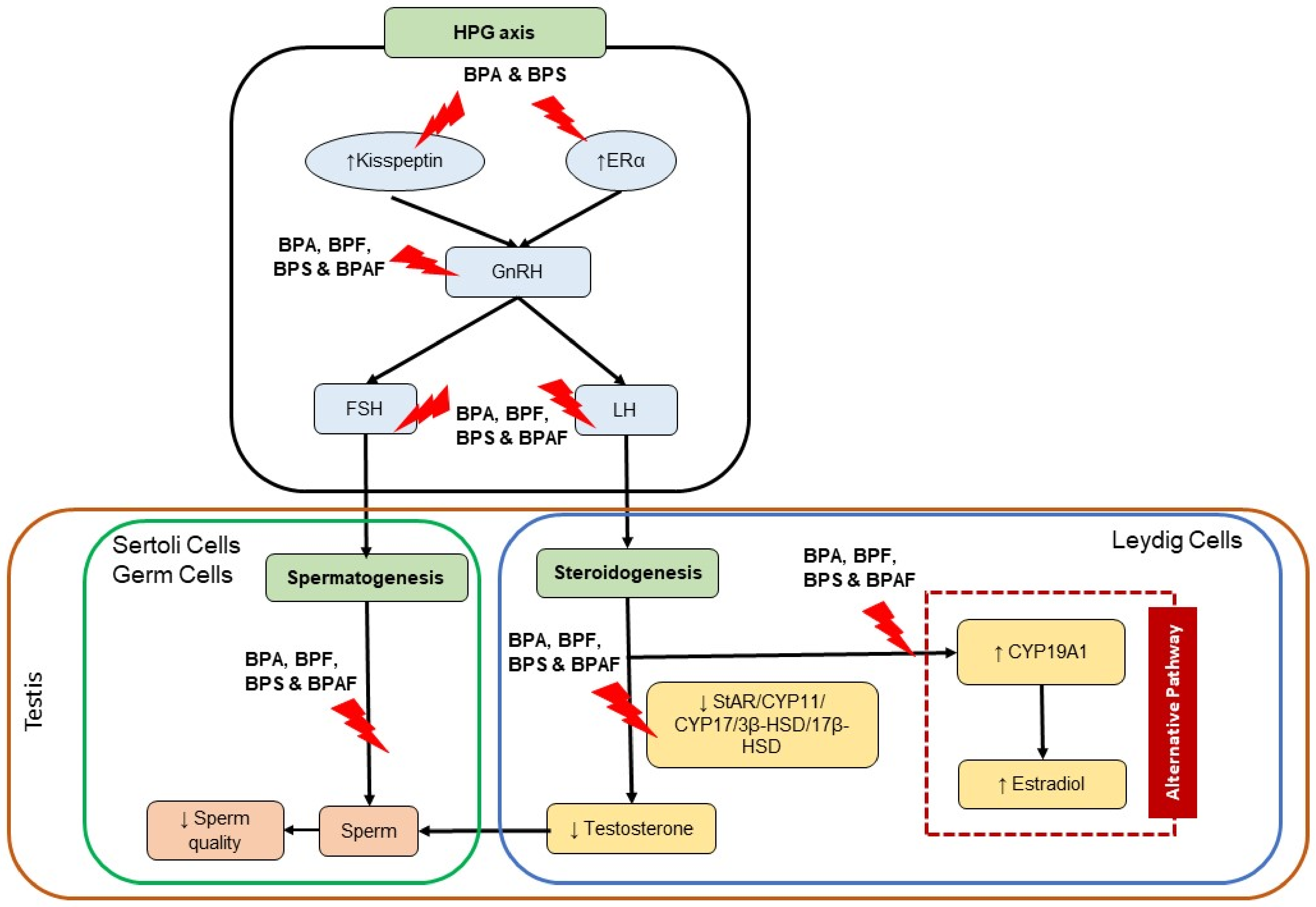
2. Bisphenols and Hypothalamus-Pituitary-Gonadal Axis in Male Reproductive System
3. Bisphenols and Steroidogenesis
4. Bisphenols and Spermatogenesis
5. The Effects of BPA and Its Analogues on Male Reproductive Hormones: Human Biological Studies Evidence
6. Discussion
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| 3β-HSD | 3β-hydroxysteroid dehydrogenase |
| 5αRed1 | 5α- reductase type 1 |
| 17β-HSD | 17β-hydroxysteroid dehydrogenase |
| AA | arachidonic acid |
| AD | androstenedione |
| AR | androgen receptor |
| ATP | adenosine 5’-triphosphate |
| BPA | bisphenol A |
| BPAF | bisphenol AF |
| BPF | bisphenol F |
| BPS | bisphenol S |
| BTB | blood-testis-barrier |
| cAMP | cyclic adenosine monophosphate |
| CatSper | sperm-specific ion calcium (Ca2+) channel |
| CYP11A1 | cytochrome P450 isoform 1A1 |
| CYP17 | cytochrome P450 isoform 17 |
| CYP19A1 | cytochrome P450 isoform 19A1 |
| COX-2 | cyclooxygenase-2 |
| DHEA | dehydroepiandrosterone |
| E2 | estradiol |
| EDCs | endocrine disrupting chemicals |
| EPA | Environmental Protection Agency |
| ER | estrogen receptor |
| ER α/β | estrogen receptor α/β |
| ERR-γ | estrogen-related receptor gamma |
| FSH | follicle-stimulating hormone |
| FSHβ | follicle-stimulating hormone beta |
| FT | free testosterone |
| GnRH | gonadotropin-releasing hormone |
| Gnrh2 | gonadotropin-releasing hormone 2 |
| GnRH3 | gonadotropin-releasing hormone 3 |
| GnRHR | GnRH receptor |
| GnRHR 1/2 | GnRH receptor 1/2 |
| GPR54 | G-protein coupled receptor 54 |
| HPG | hypothalamic–pituitary–gonadal axis |
| HSD | hydroxysteroid dehydrogenase |
| INB | inhibin B |
| IVF | in vitro fertilization |
| LC | Leydig cells |
| LH | luteinizing hormone |
| LHβ | luteinizing hormone beta |
| LHR | luteinizing hormone receptor |
| MAPK | mitogen-activated protein kinase |
| mFeTA | mouse fetal testicular cell assay |
| mRNA | messenger RNA |
| PGE-2 | prostaglandin E2 |
| PKA | protein kinase A |
| PSA | prostate-specific antigen |
| Q | quartile |
| SC | Sertoli cell |
| SHBG | Sex hormone binding globulin |
| StAR | Steroidogenic acute regulatory |
| TT | total testosterone |
| YO | years old |
| ZO-1 | zona occludens-1 |
References
- Michałowicz, J.; Bisphenol, A. Sources, toxicity and biotransformation. Environ. Toxicol. Pharmacol. 2014, 37, 738–758. [Google Scholar] [CrossRef]
- Park, B.; Kwon, J.E.; Cho, S.M.; Kim, C.W.; Lee, D.E.; Koo, Y.T.; Lee, S.H.; Lee, H.M.; Kang, S.C. Protective effect of Lespedeza cuneata ethanol extract on Bisphenol A-induced testicular dysfunction in vivo and in vitro. Biomed. Pharmacother. 2018, 102, 76–85. [Google Scholar] [CrossRef]
- Abraham, A.; Chakraborty, P. A review on sources and health impacts of bisphenol A. Rev. Environ. Health 2020, 35, 201–210. [Google Scholar] [CrossRef]
- Castillo-Sanchez, R.; Ramirez-Ricardo, J.; Martinez-Baeza, E.; Cortes-Reynosa, P.; Candanedo-Gonzales, F.; Gomez, R.; Salazar, E.P. Bisphenol A induces focal adhesions assembly and activation of FAK, Src and ERK2 via GPER in MDA-MB-231 breast cancer cells. Toxicol. Vitr. 2020, 66, 104871. [Google Scholar] [CrossRef] [PubMed]
- Talpade, J.; Shrman, K.; Sharma, R.K.; Gutham, V.; Singh, R.P.; Meena, N.S. Bisphenol A: An endocrine disruptor. J. Entomol. Zool. Stud. 2018, 6, 394–397. [Google Scholar]
- Bakar, N.A.; Salleh, M.M.; Umar, A.A.; Shapter, J.G. Design and measurement technique of surface-enhanced Raman scattering for detection of bisphenol A. Adv. Nat. Sci. Nanosci. Nanotechnol. 2017, 8, 025008. [Google Scholar] [CrossRef]
- Kang, J.-H.; Kondo, F.; Katayama, Y. Human exposure to Bisphenol A. Toxicology 2006, 226, 79–89. [Google Scholar] [CrossRef]
- Kawamura, Y.; Etoh, M.; Hirakawa, Y.; Abe, Y.; Mutsuga, M. Bisphenol A in domestic and imported canned foods in Japan. Food Addit. Contam. Part A Chem. 2014, 31, 330–340. [Google Scholar] [CrossRef]
- Pelch, K.; Wignall, J.A.; Goldstone, A.E.; Ross, P.K.; Blain, R.B.; Shapiro, A.J.; Holmgren, S.D.; Hsieh, J.H.; Svoboda, D.; Auerbach, S.S.; et al. A scoping review of the health and toxicological activity of bisphenol A (BPA) structural analogues and functional alternatives. Toxicology 2019, 424, 152235. [Google Scholar] [CrossRef]
- Bakirhan, N.K.; Ozkan, S.A. The recent electrochemical studies on bisphenol a detection in beverages. In Safety Issues in Beverage Production; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2019; Volume 18, pp. 309–333. [Google Scholar] [CrossRef]
- Bashir, A.B.; Audu, A.A. Determination of Bisphenol A Released From Polycarbonate Infant Feeding Bottles By UV-Vis Spectrophotometry. J. Chem. Soc. Niger. 2020, 45, 1120–1127. [Google Scholar] [CrossRef]
- Ugboka, U.G.; Ihedioha, J.N.; Ekere, N.R.; Okechukwu, F.O. Human health risk assessment of bisphenol A released from polycarbonate drinking water bottles and carbonated drinks exposed to sunlight in Nigeria. Int. J. Environ. Anal. Chem. 2020, 1–11. [Google Scholar] [CrossRef]
- Wang, R.; Huang, Y.; Dong, S.; Wang, P.; Su, X. The occurrence of bisphenol compounds in animal feed plastic packaging and migration into feed. Chemosphere 2021, 265, 129022. [Google Scholar] [CrossRef]
- Lalonde, B.; Garron, C. Spatial and Temporal Distribution of BPA in the Canadian Freshwater Environment. Arch. Environ. Contam. Toxicol. 2020, 78, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Liu, F.; Guo, Y.; Moon, H.B.; Nakata, H.; Wu, Q.; Kannan, K. Occurrence of eight bisphenol analogues in indoor dust from the United States and several Asian countries: Implications for human exposure. Environ. Sci. Technol. 2012, 46, 9138–9145. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Kannan, K. A survey of bisphenol A and other bisphenol analogues in foodstuffs from nine cities in China. Food Addit. Contam. Part A 2014, 31, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Rotimi, O.A.; Olawole, T.D.; De Campos, O.C.; Adelani, I.B.; Rotimi, S.O. Bisphenol A in Africa: A review of environmental and biological levels. Sci. Total Environ. 2021, 764, 142854. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, H.; Chen, Z.-L.; Wang, X.-C.; Shen, J.-M. Spatial and temporal distributions of bisphenol analogues in water and sediment from the Lanzhou section of the Yellow River, China. Arab. J. Geosci. 2020, 13, 1–8. [Google Scholar] [CrossRef]
- Derakhshan, A.; Philips, E.M.; Ghassabian, A.; Santos, S.; Asimakopoulos, A.G.; Kannan, K.; Kortenkamp, A.; Jaddoe, V.W.V.; Trasande, L.; Peeters, R.P.; et al. Association of urinary bisphenols during pregnancy with maternal, cord blood and childhood thyroid function. Environ. Int. 2021, 146, 106160. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Xie, J.; Mao, L.; Zhao, M.; Bai, X.; Wen, J.; Shen, T.; Wu, P. Bisphenol analogue concentrations in human breast milk and their associations with postnatal infant growth. Environ. Pollut. 2020, 259, 113779. [Google Scholar] [CrossRef]
- Polydorou, O.; Schmidt, O.C.; Spraul, M.; Vach, K.; Schulz, S.D.; König, A.; Hellwig, E.; Gminski, R. Detection of Bisphenol A in dental wastewater after grinding of dental resin composites. Dent. Mater. 2020, 36, 1009–1018. [Google Scholar] [CrossRef]
- Cao, X.L.; Kosarac, I.; Popovic, S.; Zhou, S.; Smith, D.; Dabeka, R. LC-MS/MS analysis of bisphenol S and five other bisphenols in total diet food samples. Food Addit. Contam Part A 2019, 36, 1740–1747. [Google Scholar] [CrossRef]
- Haq, M.E.U.; Akash, M.S.H.; Sabir, S.; Mahmood, M.H.; Rehman, K. Human exposure to bisphenol A through dietary sources and development of diabetes mellitus: A cross-sectional study in Pakistani population. Environ. Sci. Pollut. Res. 2020, 27, 26262–26275. [Google Scholar] [CrossRef] [PubMed]
- Malaisé, Y.; Lencina, C.; Cartier, C.; Olier, M.; Ménard, S.; Guzylack-Piriou, L. Perinatal oral exposure to low doses of bisphenol A, S or F impairs immune functions at intestinal and systemic levels in female offspring mice. Environ. Health 2020, 19, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rolfo, A.; Nuzzo, A.M.; De Amicis, R.; Moretti, L.; Bertoli, S.; Leone, A. Fetal–maternal exposure to endocrine disruptors: Correlation with diet intake and pregnancy outcomes. Nutrients 2020, 12, 1744. [Google Scholar] [CrossRef] [PubMed]
- Adeyemi, J.A.; Gallimberti, M.; Olise, C.C.; Rocha, B.A.; Adedire, C.O.; Barbosa, F. Evaluation of bisphenol A levels in Nigerian thermal receipts and estimation of daily dermal exposure. Environ. Sci. Pollut. Res. 2020, 27, 37645–37649. [Google Scholar] [CrossRef] [PubMed]
- González, N.; Marquès, M.; Cunha, S.C.; Fernandes, J.O.; Domingo, J.L.; Nadal, M. Biomonitoring of co-exposure to bisphenols by consumers of canned foodstuffs. Environ. Int. 2020, 140, 105760. [Google Scholar] [CrossRef]
- Sánchez-Piñero, J.; Bowerbank, S.L.; Moreda-Piñeiro, J.; López-Mahía, P.; Dean, J.R. The occurrence and distribution of polycyclic aromatic hydrocarbons, bisphenol A and organophosphate flame retardants in indoor dust and soils from public open spaces: Implications for human exposure. Environ. Pollut. 2020, 266, 115372. [Google Scholar] [CrossRef]
- Zhang, H.; Quan, Q.; Zhang, M.; Zhang, N.; Zhang, W.; Zhan, M.; Xu, W.; Lu, L.; Fan, J.; Wang, Q. Occurrence of bisphenol A and its alternatives in paired urine and indoor dust from Chinese university students: Implications for human exposure. Chemosphere 2020, 247, 125987. [Google Scholar] [CrossRef]
- Hart, R.J. The Impact of Prenatal Exposure to Bisphenol A on Male Reproductive Function. Front. Endocrinol. 2020, 11, 320. [Google Scholar] [CrossRef] [PubMed]
- Meli, R.; Monnolo, A.; Annunziata, C.; Pirozzi, C.; Ferrante, M.C. Oxidative stress and BPA toxicity: An antioxidant approach for male and female reproductive dysfunction. Antioxidants 2020, 9, 405. [Google Scholar] [CrossRef]
- Shokry, D.A.; Mohamed, M.I.; Abdel-Satar, M.F.; Selim, N.E.S.; Abd El Salam, M.A. Correlation between urinary bisphenol a (BPA) levels and male reproductive functions among sample of Egyptian population. Indian J. Forensic Med. Toxicol. 2020, 14, 1821–1826. [Google Scholar] [CrossRef]
- Aljadeff, G.; Longhi, E.; Shoenfeld, Y. Bisphenol A: A notorious player in the mosaic of autoimmunity. Autoimmunity 2018, 51, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, H.S. Impact of bisphenol A on the cardiovascular system—Epidemiological and experimental evidence and molecular mechanisms. Int. J. Environ. Res. Public Health 2014, 11, 8399–8413. [Google Scholar] [CrossRef] [Green Version]
- Kharrazian, D. The potential roles of bisphenol A (BPA) pathogenesis in autoimmunity. Autoimmune Dis. 2014, 2014, 743616. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, Y.; Goldblum, R.M.; Midoro-Horiuti, T. Fetal exposure to bisphenol A as a risk factor for the development of childhood asthma: An animal model study. Environ. Health 2012, 11, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skakkebaek, N.E.; Toppari, J.; Söder, O.; Gordon, C.M.; Divall, S.; Draznin, M. The exposure of fetuses and children to endocrine disrupting chemicals: A European Society for Paediatric Endocrinology (ESPE) and Pediatric Endocrine Society (PES) call to action statement. J. Clin. Endocrinol. Metab. 2011, 96, 3056–3058. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, H.; Tu, X.; Yuan, W.; Zhou, Z.; Jin, L.; Miao, M.; Li, D.K. Bisphenol A and pubertal height growth in school-aged children. J. Expo. Sci. Environ. Epidemiol. 2019, 29, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Chianese, R.; Troisi, J.; Richards, S.; Scafuro, M.; Fasano, S.; Guida, M.; Pierantoni, R.; Meccariello, R. Bisphenol A in Reproduction: Epigenetic Effects. Curr. Med. Chem. 2018, 25, 748–770. [Google Scholar] [CrossRef] [PubMed]
- Lombó, M.; Fernández-Díez, C.; González-Rojo, S.; Herráez, M.P. Genetic and epigenetic alterations induced by bisphenol A exposure during different periods of spermatogenesis: From spermatozoa to the progeny. Sci. Rep. 2019, 9, 18029. [Google Scholar] [CrossRef] [Green Version]
- Gys, C.; Bastiaensen, M.; Malarvannan, G.; Ait Bamai, Y.; Araki, A.; Covaci, A. Short-term variability of bisphenols in spot, morning void and 24-hour urine samples. Environ. Pollut. 2021, 268, 115747. [Google Scholar] [CrossRef]
- Huang, M.; Liu, S.; Fu, L.; Jiang, X.; Yang, M. Bisphenol A and its analogues bisphenol S, bisphenol F and bisphenol AF induce oxidative stress and biomacromolecular damage in human granulosa KGN cells. Chemosphere 2020, 253, 126707. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Zhuang, T.; Shi, W.; Liang, Y.; Liao, C.; Song, M.; Jiang, G. Serum concentration of bisphenol analogues in pregnant women in China. Sci. Total Environ. 2020, 707, 136100. [Google Scholar] [CrossRef]
- Sharma, P.; Gautam, D.K.; Bhagat, P.; Mandal, M.B. Comparative assessment of impact of Bisphenol A and Bisphenol S on hematological parameters in rats after 4 weeks exposure. J. Crit. Rev. 2020, 7, 1250–1256. [Google Scholar]
- Andújar, N.; Gálvez-Ontiveros, Y.; Zafra-Gómez, A.; Rodrigo, L.; Álvarez-Cubero, M.J.; Aguilera, M.; Monteagudo, C.; Rivas, A. Bisphenol A analogues in food and their hormonal and obesogenic effects: A review. Nutrients 2019, 11, 2136. [Google Scholar] [CrossRef] [Green Version]
- García-Córcoles, M.T.; Cipa, M.; Rodríguez-Gómez, R.; Rivas, A.; Olea-Serrano, F.; Vílchez, J.L.; Zafra-Gómez, A. Determination of bisphenols with estrogenic activity in plastic packaged baby food samples using solid-liquid extraction and clean-up with dispersive sorbents followed by gas chromatography tandem mass spectrometry analysis. Talanta 2018, 178, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Harnett, K.G.; Chin, A.; Schuh, S.M. BPA and BPA alternatives BPS, BPAF, and TMBPF, induce cytotoxicity and apoptosis in rat and human stem cells. Ecotoxicol. Environ. Saf. 2021, 216, 112210. [Google Scholar] [CrossRef]
- Environmental Protection Agency (EPA). Bisphenol A Alternatives in Thermal Paper Final Report. Available online: https://www.epa.gov/sites/default/files/2015-08/documents/bpa_final.pdf (accessed on 10 June 2021).
- Jubaidi, F.F.; Mathialagan, R.D.; Noor, M.M.; Taib, I.S.; Budin, S.B. Monosodium glutamate daily oral supplementation: Study of its effects on male reproductive system on rat model. Syst. Biol. Reprod. Med. 2019, 65, 194–204. [Google Scholar] [CrossRef]
- Yusoff, N.A.; Mohamed, M.; Budin, S.B.; Taib, I.S. Fenitrothion impaired sexual behaviour and reproductive performance in male sprague-Dawley rats. Sains Malays. 2020, 49, 1333–1344. [Google Scholar] [CrossRef]
- Gules, O.; Yildiz, M.; Naseer, Z.; Tatar, M. Effects of folic acid on testicular toxicity induced by bisphenol-A in male Wistar rats. Biotech. Histochem. 2019, 94, 26–35. [Google Scholar] [CrossRef]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Khan, M.J. Bisphenol A analogues bisphenol B, bisphenol F, and bisphenol S induce oxidative stress, disrupt daily sperm production, and damage DNA in rat spermatozoa: A comparative in vitro and in vivo study. Toxicol. Ind. Health 2019, 35, 294–303. [Google Scholar] [CrossRef]
- Yusoff, N.A.; Budin, S.B.; Taib, I.S. Pesticide Exposures Induce Male-Mediated Reproductive Toxicity: A Review. J. Agric. Sci. 2017, 9, 122–135. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.S.; Pang, M.G. Understanding the molecular mechanisms of bisphenol A action in spermatozoa. Clin. Exp. Reprod. Med. 2019, 46, 99–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roelofs, M.J.E.; van den Berg, M.; Bovee, T.F.H.; Piersma, A.H.; Duursen, M.B.M. Structural bisphenol analogues differentially target steroidogenesis in murine MA-10 Leydig cells as well as the glucocorticoid receptor. Toxicology 2015, 329, 10–20. [Google Scholar] [CrossRef]
- Yang, Q.; Yang, X.; Liu, J.; Ren, W.; Chen, Y.; Shen, S. Effects of BPF on steroid hormone homeostasis and gene expression in the hypothalamic–pituitary–gonadal axis of zebrafish. Environ. Sci. Pollut. Res. 2017, 24, 21311–21322. [Google Scholar] [CrossRef]
- Bruks, S. Metabolism and Estrogenicity of Bisphenol A and Its Analogues: A Comparative Analysis of Experimental and Computational Data on Metabolism of Bisphenols. Bachelor’s Thesis, Umeå University, Umeå, Sweden, 24 April 2020. [Google Scholar]
- Zühlke, M.K.; Schlüter, R.; Mikolasch, A.; Henning, A.K.; Giersberg, M.; Lalk, M.; Kunze, G.; Schweder, T.; Urich, T.; Schauer, F. Biotransformation of bisphenol A analogues by the biphenyl-degrading bacterium Cupriavidus basilensis—A structure-biotransformation relationship. Appl. Microbiol. Biotechnol. 2020, 104, 3569–3583. [Google Scholar] [CrossRef]
- Cano-Nicolau, J.; Vaillant, C.; Pellegrini, E.; Charlier, T.D.; Kah, O.; Coumailleau, P. Estrogenic effects of several BPA analogs in the developing zebrafish brain. Front. Neurosci. 2016, 10, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- In, S.; Cho, H.; Lee, K.W.; Won, E.J.; Lee, Y.M. Cloning and molecular characterization of estrogen-related receptor (ERR) and vitellogenin genes in the brackish water flea Diaphanosoma celebensis exposed to bisphenol A and its structural analogues. Mar. Pollut. Bull. 2020, 154, 111063. [Google Scholar] [CrossRef]
- Lina, S.; Eliza, H.; Hashida, N.H.; Ibrahim, S.F.; Osman, K. Androgen receptor and ultrastructural features of nigella sativa oil and nicotine-treated male rat reproductive glands. Sains Malays. 2018, 47, 1827–1833. [Google Scholar] [CrossRef]
- Qiu, W.; Zhao, Y.; Yang, M.; Farajzadeh, M.; Pan, C.; Wayne, N.L. Actions of bisphenol A and bisphenol S on the reproductive neuroendocrine system during early development in zebrafish. Endocrinology 2016, 157, 636–647. [Google Scholar] [CrossRef] [Green Version]
- Matuszczak, E.; Komarowska, M.D.; Debek, W.; Hermanowicz, A. The Impact of Bisphenol A on Fertility, Reproductive System, and Development: A Review of the Literature. Int. J. Endocrinol. 2019, 2019, 4068717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alboghobeish, S.; Mahdavinia, M.; Zeidooni, L.; Samimi, A.; Oroojan, A.A.; Alizadeh, S.; Dehghani, M.A.; Ahangarpour, A.; Khorsandi, L. Efficiency of naringin against reproductive toxicity and testicular damages induced by bisphenol A in rats. Iran J. Basic Med. Sci. 2019, 22, 315–323. [Google Scholar] [CrossRef]
- Jiang, Z.; Wang, J.; Li, X.; Zhang, X. Echinacoside and Cistanche tubulosa (Schenk) R. wight ameliorate bisphenol A-induced testicular and sperm damage in rats through gonad axis regulated steroidogenic enzymes. J. Ethnopharmacol. 2016, 193, 321–328. [Google Scholar] [CrossRef]
- Zahra, Z.; Khan, M.R.; Majid, M.; Maryam, S.; Sajid, M. Gonadoprotective ability of Vincetoxicum arnottianum extract against bisphenol A-induced testicular toxicity and hormonal imbalance in male Sprague Dawley rats. Andrologia 2020, 52, e13590. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, C.; Jiang, Z.; Wang, M.; Jiang, H.; Zhang, X. Protective effect of Cordyceps militaris extract against bisphenol A induced reproductive damage. Syst. Biol. Reprod. Med. 2016, 62, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Pirzada, M.; Afsar, T.; Razak, S.; Almajwal, A.; Jahan, S. Effect of bisphenol F, an analog of bisphenol A, on the reproductive functions of male rats. Environ. Health Prev. Med. 2019, 24, 41. [Google Scholar] [CrossRef]
- Ullah, A.; Pirzada, M.; Jahan, S.; Ullah, H.; Shaheen, G.; Rehman, H.; Siddiqui, M.F.; Butt, M.A. Bisphenol A and its analogs bisphenol B, bisphenol F, and bisphenol S: Comparative in vitro and in vivo studies on the sperms and testicular tissues of rats. Chemosphere 2018, 209, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Jahan, S.; Ain, Q.U.; Shaheen, G.; Ahsan, N. Effect of bisphenol S exposure on male reproductive system of rats: A histological and biochemical study. Chemosphere 2016, 152, 383–391. [Google Scholar] [CrossRef]
- Stoker, C.; Andreoli, M.F.; Kass, L.; Bosquiazzo, V.L.; Rossetti, M.F.; Canesini, G.; Luque, E.H.; Ramos, J.G. Perinatal exposure to bisphenol A (BPA) impairs neuroendocrine mechanisms regulating food intake and kisspetin system in adult male rats. Evidences of metabolic disruptor hypothesis. Mol. Cell. Endocrinol. 2020, 499, 110614. [Google Scholar] [CrossRef]
- Bai, Y.; Chang, F.; Zhou, R.; Jin, P.P.; Matsumoto, H.; Sokabe, M.; Chen, L. Increase of anteroventral periventricular kisspeptin neurons and generation of E2-induced LH-surge system in male rats exposed perinatally to environmental dose of bisphenol-A. Endocrinology 2011, 152, 1562–1571. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Mickens, J.A.; McCaffrey, K.A.; Leyrer, S.M.; Patisaul, H.B. Neonatal Bisphenol A exposure alters sexually dimorphic gene expression in the postnatal rat hypothalamus. Neurotoxicology 2012, 33, 23–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Jiao, Z.; Zheng, S.; Li, M.; Zhang, J.; Feng, Y.; Yin, J.; Shao, B. Long-term effects of Bisphenol AF (BPAF) on hormonal balance and genes of hypothalamus-pituitary-gonad axis and liver of zebrafish (Danio rerio), and the impact on offspring. Chemosphere 2015, 128, 252–257. [Google Scholar] [CrossRef]
- Majid, M.; Ijaz, F.; Baig, M.W.; Nasir, B.; Khan, M.R.; Haq, I.U. Scientific Validation of Ethnomedicinal Use of Ipomoea batatas L. Lam. as Aphrodisiac and Gonadoprotective Agent against Bisphenol A Induced Testicular Toxicity in Male Sprague Dawley Rats. Biomed. Res. Int. 2019, 2019, 8939854. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, P.; Romano, R.M.; Kizys, M.M.L.; Oliveira, K.C.; Kasamatsu, T.; Giannocco, G.; Chiamolera, M.I.; Dias-da-Silva, M.R.; Romano, M.A. Adult exposure to bisphenol A (BPA) in Wistar rats reduces sperm quality with disruption of the hypothalamic-pituitary-testicular axis. Toxicology 2015, 329, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akintunde, J.K.; Farouk, A.A.; Mogbojuri, O. Metabolic treatment of syndrome linked with Parkinson’s disease and hypothalamus pituitary gonadal hormones by turmeric curcumin in Bisphenol-A induced neuro-testicular dysfunction of wistar rat. Biochem. Biophys. Rep. 2019, 17, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Eladak, S.; Grisin, T.; Moison, D.; Guerquin, M.J.; N’Tumba-Byn, T.; Pozzi-Gaudin, S.; Benachi, A.; Livera, G.; Rouiller-Fabre, V.; Habert, R. A new chapter in the bisphenol a story: Bisphenol S and bisphenol F are not safe alternatives to this compound. Fertil. Steril. 2015, 103, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Feng, Y.; Jiao, Z.; Shi, J.; Li, M.; Guo, Q.; Shao, B. Effects of bisphenol analogues on steroidogenic gene expression and hormone synthesis in H295R cells. Chemosphere 2016, 147, 9–19. [Google Scholar] [CrossRef]
- Jahan, S.; Ain, Q.U.; Ullah, H. Therapeutic effects of quercetin against bisphenol A induced testicular damage in male Sprague Dawley rats. Syst. Biol. Reprod. Med. 2016, 62, 114–124. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Yin, L.; Yu, K.S.; Hofmann, M.C.; Yu, X. High-content analysis provides mechanistic insights into the testicular toxicity of Bisphenol A and selected analogues in mouse spermatogonial cells. Toxicol. Sci. 2017, 155, 43–60. [Google Scholar] [CrossRef]
- de Freitas, A.T.A.G.; Ribeiro, M.A.; Pinho, C.F.; Peixoto, A.R.; Domeniconi, R.F.; Scarano, W.R. Regulatory and junctional proteins of the blood-testis barrier in human Sertoli cells are modified by monobutyl phthalate (MBP) and bisphenol A (BPA) exposure. Toxicol. Vitr. 2016, 34, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.-L.; Yin, Y.-Y.; Wang, X.-S.; Wang, P.-T. BPA Disturb TJ-Permeability of Rat Sertoli Cells During Spermatogenesis in Vitro. Prog. Mod. Biomed. 2012, 12, 1430–1434. [Google Scholar]
- Wang, H.F.; Liu, M.; Li, N.; Luo, T.; Zheng, L.P.; Zeng, X.H. Bisphenol a impairs mature sperm functions by a CatSper-relevant mechanism. Toxicol. Sci. 2016, 152, 145–154. [Google Scholar] [CrossRef]
- Rehfeld, A.; Andersson, A.M.; Skakkebæk, N.E. Bisphenol A Diglycidyl Ether (BADGE) and Bisphenol Analogs, but Not Bisphenol A (BPA), Activate the CatSper Ca2+ Channel in Human Sperm. Front. Endocrinol. 2020, 11, 324. [Google Scholar] [CrossRef]
- Zhu, N.; Zhao, M.; Song, Y.; Ding, L.; Ni, Y. The KiSS-1/GPR54 system: Essential roles in physiological homeostasis and cancer biology. Genes Dis. 2020, in press. [Google Scholar] [CrossRef]
- Dedes, I. Kisspeptins and the control of gonadotrophin secretion. Syst. Biol. Reprod. Med. 2012, 58, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Khbouz, B.; de Bournonville, C.; Court, L.; Taziaux, M.; Corona, R.; Arnal, J.F.; Lenfant, F.; Cornil, C.A. Role for the membrane estrogen receptor alpha in the sexual differentiation of the brain. Eur. J. Neurosci. 2020, 52, 2627–2645. [Google Scholar] [CrossRef]
- Weiser, M.J.; Foradori, C.D.; Handa, R.J. Estrogen receptor beta in the brain: From form to function. Brain Res. Rev. 2008, 57, 309–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimento, A.; Sirianni, R.; Casaburi, I.; Pezzi, V. Role of estrogen receptors and G protein-coupled estrogen receptor in regulation of hypothalamus-pituitary-testis axis and spermatogenesis. Front. Endocrinol. 2014, 5, 1. [Google Scholar] [CrossRef]
- Yeo, S.H.; Colledge, W.H. The role of Kiss1 neurons as integrators of endocrine, metabolic, and environmental factors in the hypothalamic-pituitary-gonadal axis. Front. Endocrinol. 2018, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Lin, M.-C.A.; Farajzadeh, M.; Wayne, N.L. Early Development of the Gonadotropin-Releasing Hormone Neuronal Network in Transgenic Zebrafish. Front. Endocrinol. 2013, 4, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zirkin, B.R.; Papadopoulos, V. Leydig cells: Formation, function, and regulation. Biol. Reprod. 2018, 99, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Flück, C.E.; Pandey, A.V. Testicular Steroidogenesis. In Endocrinology of the Testis and Male Reproduction, 1st ed.; Simoni, M., Huhtaniemi, I., Eds.; Springer: Cham, Switzerland, 2017; pp. 343–371. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, F.; Ye, L.; Zirkin, B.; Chen, H. Steroidogenesis in leydig cells: Effects of aging and environmental factors. Reproduction 2017, 154, R111–R122. [Google Scholar] [CrossRef] [Green Version]
- Jefcoate, C.R.; Lee, J. Cholesterol signaling in single cells: Lessons from STAR and sm-FISH. J. Mol. Endocrinol. 2018, 60, R213–R235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadopoulos, V.; Aghazadeh, Y.; Fan, J.; Campioli, E.; Zirkin, B.; Midzak, A. Translocator protein-mediated pharmacology of cholesterol transport and steroidogenesis. Mol. Cell. Endocrinol. 2015, 408, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Ronco, A.M.; Moraga, P.F.; Llanos, M.N. Arachidonic acid release from rat Leydig cells: The involvement of G protein, phospholipase A2 and regulation of cAMP production. J. Endocrinol. 2002, 172, 95–104. [Google Scholar] [CrossRef]
- Hadley, K.B.; Ryan, A.S.; Forsyth, S.; Gautier, S.; Salem, N. The essentiality of arachidonic acid in infant development. Nutrients 2016, 8, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouzer, C.A.; Marnett, L.J. Cyclooxygenases: Structural and functional insights. J. Lipid Res. 2009, 50, S29–S34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrotta, I.; Santoro, M.; Guido, C.; Avena, P.; Tripepi, S.; De Amicis, F.; Gervasi, M.C.; Aquila, S. Expression of cyclooxygenase-1 (COX-1) and COX-2 in human male gametes from normal patients, and those with varicocele and diabetes: A potential molecular marker for diagnosing male infertility disorders. J. Anat. 2012, 221, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Dyson, M.T.; Jo, Y.; Stocco, D.M. Inhibition of cyclooxygenase-2 activity enhances steroidogenesis and steroidogenic acute regulatory gene expression in MA-10 mouse Leydig cells. Endocrinology 2003, 144, 3368–3375. [Google Scholar] [CrossRef] [PubMed]
- Scott, H.M.; Mason, J.I.; Sharpe, R.M. Steroidogenesis in the fetal testis and its susceptibility to disruption by exogenous compounds. Endocr. Rev. 2009, 30, 883–925. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Mruk, D.D.; Wong, C.K.C.; Yan Cheng, C. Germ cell transport across the seminiferous epithelium during spermatogenesis. Physiology 2014, 29, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Li, M.W.M.; Mruk, D.D.; Lee, W.M.; Cheng, C.Y. Disruption of the blood-testis barrier integrity by bisphenol A in vitro: Is this a suitable model for studying blood-testis barrier dynamics? Int. J. Biochem. Cell Biol. 2009, 41, 2302–2314. [Google Scholar] [CrossRef] [Green Version]
- Kamaruzaman, K.A.; Mat Noor, M. Reversible spermatoxic effect of Andrographis paniculata methanol extract in Sprague Dawley rats. Malays. Appl. Biol. 2012, 46, 225–232. [Google Scholar]
- Sun, H.X.; Zhu, Y.; Wang, L.; Ling Liu, H.; Ling, Y.; Li, Z.L.; Sun, L.B. The Catsper channel and its roles in male fertility: A systematic review. Reprod. Biol. Endocrinol. 2017, 15, 65. [Google Scholar] [CrossRef] [Green Version]
- Scinicariello, F.; Buser, M.C. Serum testosterone concentrations and urinary bisphenol a, benzophenone-3, triclosan, and paraben levels in male and female children and adolescents: NHANES 2011–2012. Environ. Health Perspect. 2016, 124, 1898–1904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Miao, M.; Zhou, Z.; Gao, E.; Chen, J.; Wang, J.; Sun, F.; Yuan, W.; Li, D.K. Exposure to bisphenol-A and reproductive hormones among male adults. Environ. Toxicol. Pharmacol. 2015, 39, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Wu, K.; Wang, Y.; Zhu, H.; Deng, Z.; Peng, L.; Zhu, G. Association of Serum Bisphenol-A Concentration and Male Reproductive Function among Exposed Workers. Arch. Environ. Contam. Toxicol. 2015, 68, 38–45. [Google Scholar] [CrossRef]
- Lassen, T.H.; Frederiksen, H.; Jensen, T.K.; Petersen, J.H.; Joensen, U.N.; Main, K.M.; Skakkebaek, N.E.; Juul, A.; Jørgensen, N.; Andersson, A.M. Urinary bisphenol a level in young men: Association with reproductive hormones and semen quality. Environ. Health Perspect. 2014, 122, 478–484. [Google Scholar] [CrossRef] [Green Version]
- Adoamnei, E.; Mendiola, J.; Vela-Soria, F.; Fernández, M.F.; Olea, N.; Jørgensen, N.; Swan, S.H.; Torres-Cantero, A.M. Urinary bisphenol A concentrations are associated with reproductive parameters in young men. Environ. Res. 2018, 161, 122–128. [Google Scholar] [CrossRef]
- Taylor, J.A.; Richter, C.A.; Ruhlen, R.L.; Vom Saal, F.S. Estrogenic environmental chemicals and drugs: Mechanisms for effects on the developing male urogenital system. J. Steroid Biochem. Mol. Biol. 2011, 127, 83–95. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, K.K.; Peterson, K.E.; Lee, J.M.; Mercado-García, A.; Blank-Goldenberg, C.; Téllez-Rojo, M.M.; Meeker, J.D. Prenatal and peripubertal phthalates and bisphenol A in relation to sex hormones and puberty in boys. Reprod. Toxicol. 2014, 47, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Hart, R.J.; Doherty, D.A.; Keelan, J.A.; Minaee, N.S.; Thorstensen, E.B.; Dickinson, J.E.; Pennell, C.E.; Newnham, J.P.; McLachlan, R.; Norman, R.J.; et al. The impact of antenatal Bisphenol A exposure on male reproductive function at 20–22 years of age. Reprod. Biomed. Online 2018, 36, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Knez, J.; Kranvogl, R.; Breznik, B.P.; Vončina, E.; Vlaisavljević, V. Are urinary bisphenol A levels in men related to semen quality and embryo development after medically assisted reproduction? Fertil. Steril. 2014, 101, 215–221.e5. [Google Scholar] [CrossRef]
- Ghayda, R.A.; Williams, P.L.; Chavarro, J.E.; Ford, J.B.; Souter, I.; Calafat, A.M.; Hauser, R.; Mínguez-Alarcón, L. Urinary bisphenol S concentrations: Potential predictors of and associations with semen quality parameters among men attending a fertility center. Environ. Int. 2019, 131, 105050. [Google Scholar] [CrossRef] [PubMed]
- Goldstone, A.E.; Chen, Z.; Perry, M.J.; Kannan, K.; Louis, G.M.B. Urinary bisphenol A and semen quality, the LIFE study. Reprod. Toxicol. 2015, 51, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Benson, T.E.; Gaml-Sørensen, A.; Ernst, A.; Brix, N.; Hougaard, K.S.; Hærvig, K.K.; Ellekilde Bonde, J.P.; Tøttenborg, S.S.; Lindh, C.H.; Ramlau-Hansen, C.H.; et al. Urinary Bisphenol A, F and S Levels and Semen Quality in Young Adult Danish Men. Int. J. Environ. Res. Public Health 2021, 8, 1742. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Type of Bisphenol | Purity (Manufacturer) | Dose (Route) | Animal | Duration of Exposure | Findings | Author |
|---|---|---|---|---|---|---|
| BPA | 99% (Sigma-Aldrich, St. Louis, MO, USA) | 50 µg/kg/bw (drinking water) | Adult male Wistar rats | Perinatal exposure (From gestation day 9 until lactation day 20) | ↑ Kiss1 mRNA expression (hypothalamus) ↓ ERα, ERβ (hypothalamus) ↑ testosterone (blood) ↓ estradiol (blood) | [71] |
| BPA | >99% (Sigma-Aldrich) | 2 µg/kg/bw (s.c) | Offspring male SD | Perinatal exposure Day 10 of gestation until day 7 of lactation | ↑ Kiss1 mRNA expression (brain) ↑ GnRH neuron ↑ LH, estradiol (blood) ↓ testosterone (blood) | [72] |
| BPA | - | 50 mg/kg/bw (s.c.) | Male pup Long Evans rats | 2 days Postnatal day 0–2 | ↑ expression of Kiss1 (brain) ↑ expression of ERα (brain) | [73] |
| BPA | MERCK, Kenilworth, NJ, USA | 50 mg/kg/bw (i.p.) | Adult male SD rats | Alternate day until 21 days | ↓ FSH, LH, testosterone (serum) ↑ estradiol (serum) | [75] |
| BPA | Sigma-Aldrich | 5 or 25 mg/kg/bw (oral gavage) | Adult male Wistar rats | 40 days | ↑ expression of Gnrhr, Lhb, Fshb, ERβ, AR mRNA (pituitary) ↓ Gnrh, ERα (hypothalamus) ↓ FSH, LH, testosterone (blood) ↑ estradiol (blood) | [76] |
| BPA | >99% (Sigma-Aldrich) | 50 mg/kg/bw (oral gavage) | Adult male Wistar rats | 14 days | ↓ FSH, LH, testosterone (serum) | [77] |
| BPA | Sigma-Aldrich | 50 mg/kg/bw (oral gavage) | Adult male Wistar rats | 30 days | ↓ FSH, LH, testosterone, E2 (plasma) | [64] |
| BPA | Sigma-Aldrich | 200 mg/kg (oral gavage) | Adult male SD rats | 42 days | ↓ FSH, LH, testosterone (blood) | [65] |
| BPA | - | 25 mg/kg/bw (i.p.) | Adult male SD rats | Alternate day for 30 days | ↓ FSH, LH, testosterone (plasma) | [66] |
| BPA | Gracia chengdu chemical technology co, Chengdu, Sichuan, China. | 200 mg/kg (oral gavage) | Adult male SD rats | 28 days | ↓ FSH, LH, testosterone (blood) | [67] |
| BPF | 99% purity (Santa Cruz Biotechnologie, Dallas, TX, USA) | 1, 5, 25, 50, and 100 mg/kg/bw (Oral gavage) | Adult male SD rats | 28 days | ↓ FSH, LH, testosterone (plasma) ↓ testicular testosterone | [68] |
| BPA/BPS | >98% | 1000 μg/L (BPA) 100 μg/L (BPS) | Transgenic zebrafish (embryo) | 2 h of post fertilization until 25 or 20 h of post fertilization | BPA: ↑ number of GnRH3 neuron at 25 hpf (hypothalamus) ↑ expression of Kiss1 mRNA, Kiss1 receptor, gnrh3, lhβ, and fshβ after 120 h post fertilization (embryo) ↑ expression of ERα at 25 hpf BPS: ↑ number of GnRH3 neuron at 25 hpf (hypothalamus) ↑ expression of mRNA Kiss1 and gnrh3 at 25 hpf (hypothalamus) ↑ expression of ERα 25 hpf | [62] |
| BPF/BPS | 99% purity (Santa Cruz Biotechnologie) | 5, 50, and 500 mg/kg/bw (Oral gavage) | Adult male SD rats | 28 days | BPF: ↓ testicular and plasma testosterone BPS: ↓ testicular and plasma testosterone | [69] |
| BPF | > 98% (J&K Scientific Ltd., Newark, DE, USA) | 0.1 and 1 mg/L in aquarium water Renewed 50% of water every 2 days | Male Zebrafish | 21 days | ↑ expression of GnRH2, GnRH3, GnRHR1, and GnRHR2 (Brain) ↑ FSHR, LHR (testis) ↓ testosterone (testis) ↑ estradiol (testis) | [56] |
| BPS/BPF/ BPAF | 98% (Sigma-Aldrich) | In vitro: BPAF: 0.076 µm BPA: 2.8 µm BPF: 10.6 µm | Zebrafish (embryo and larvae) | From day 1 of hdf until 7-dpf larva | BPAF, BPA, BPF: ↑ affinity toward binding of ERα (in vitro) BPS: No affinity toward the ERα receptor (in vitro) | [59] |
| BPS | - | 1, 50 µg/kg/bw (oral gavage) | Adult male SD rats | 28 days | ↓ testicular and plasma testosterone | [70] |
| BPAF | 99% (Sigma-Aldrich) | 25 and 125 µg/L | Offspring male zebrafish (embryo) | 120 days (exposure during embryo and larva stages) | ↑ gnrh2, fshβ, lhβ, and cyp19b in 125 µg/L ↑ estradiol ↓ testosterone | [74] |
| Type of Bisphenol | Purity (Manufacturer) | Dose (Route) | Animal | Duration of Exposure | Findings | Author |
|---|---|---|---|---|---|---|
| BPA | 99% (Sigma-Aldrich) | 50 mg/kg/bw (oral gavage) | Adult male Wistar rats | 14 days | ↓ testosterone (serum) | [80] |
| BPA | 99% (Sigma-Aldrich) | 50 µg/kg/bw (drinking water) | Adult male Wistar rats | Perinatal exposure (gestation day 9 until lactation day 20) | ↑ testosterone (blood) ↓ estradiol (blood) | [71] |
| BPA | >99% (Sigma-Aldrich) | 2 µg/kg/bw (s.c) | Offspring male SD rats | Perinatal exposure Day 10 of gestation until day 7 of lactation | ↑ estradiol (blood) ↓ testosterone (blood) | [72] |
| BPA | Sigma-Aldrich | 5 or 25 mg/kg/bw (oral gavage) | Adult male Wistar rats | 40 days | ↓ testosterone (blood) ↑ estradiol (blood) | [76] |
| BPA | MERCKS | 50 mg/kg/bw (i.p.) | Adult male SD rats | Alternate day until 21 days | ↓ testosterone (serum) ↑ estradiol (serum) | [75] |
| BPA | Gracia chengdu chemical technology co. | 200 mg/kg (oral gavage) | Adult male SD rats | 28 days | ↓ testosterone (blood) ↓ expression of mRNA StAR, CYP11A1, 3β-HSD, CYP17A1 (testis) | [67] |
| BPA | Sigma-Aldrich | 200 mg/kg (oral gavage) | Adult male SD rats | 42 days | ↓ testosterone (blood) ↓ expression of protein StAR, CYP11A1, 17β-HSD, 3β-HSD, CYP17A1 (testis) | [65] |
| BPA | Sigma-Aldrich | 50 mg/kg/bw (oral gavage) | Adult male Wistar rats | 30 days | ↓ testosterone, estradiol (plasma) | [64] |
| BPA | - | 25 mg/kg/bw (i.p.) | Adult male SD rats | Alternate day of 30 days | ↓ testosterone (plasma) | [66] |
| BPF | 99% (Santa Cruz Biotechnologie) | 1, 5, 25, 50, and 100 mg/kg/bw | Adult male SD rats | 28 days | ↓ testosterone (plasma) ↓ testosterone (testis) | [68] |
| BPF/BPS | 99% (Santa Cruz Biotechnologie) | 5, 50, and 500 mg/kg/bw | Adult male SD rats | 28 days | BPF: ↓ testicular and plasma testosterone BPS: ↓ testicular and plasma testosterone | [69] |
| BPF | > 98% (J&K Scientific Ltd) | 0.1 and 1 mg/L in aquarium water | Male Zebrafish | 21 days | ↓ testosterone (homogenate) ↑ estradiol (homogenate) ↑ expression of CYP19A1b (aromatase)(Brain) ↑ expression of mRNA CYP11A, CYP19A (testis) ↓ expression of mRNA StAR, CYP17, 17βHSD (testis) | [56] |
| BPS/BPF/ BPAF | 98% (Sigma-Aldrich) | In vivo: 1 µm In vitro: BPAF: 0.076 µm BPA:2.8 µm BPF: 10.6 µm | Zebrafish (embryo and larvae) | From day 1 of hdf until 7-dpf larva | BPAF, BPF, BPS: ↑ expression of mRNA CYP19A1 gene in 7-dpf (embryo) | [59] |
| BPS | - | 1, 50 µg/kg/bw (oral gavage) | Adult male SD rats | 28 days | ↓ testicular and plasma testosterone | [70] |
| BPAF | 99% (Sigma–Aldrich) | 25 and 125 µg/L | Offspring male zebrafish (embryo) | 120 days through exposure of embryo and larva stages - | ↑ estradiol ↓ testosterone ↑ CYP19b (brain) ↑ expression of mRNA CYP19A and CYP11A1 (testis) ↓ expression of mRNA StAR and CYP17 (testis) | [74] |
| BPF/BPS | BPF (>99%) BPS (>98%) | BPF: 0.01–100 µM BPS: 0.01–30 µM | MA-10 Leydig cell culture | 48 h | BPF, BPS: ↑ testosterone secretion ↑ expression of 5αRed1 | [55] |
| BPF/BPS | - | 10, 100, 1000, 10,000 nmol/L | Mouse fetal testis assay (mFeTA) | 1–3 days | BPF, BPS (10 000 nmol/L): ↓ testosterone secretion ↓ expression of mRNA StAR, HSD3β1 and CYP17A1 | [78] |
| BPF/BPS/ BPAF | BPF (99%) BPS (98%) BPAF (99%) | 0.1, 1, 10, 30, 50 and 70 µM | Human adrenocortical carcinoma cell line (H295R) | - | BPF: ↑ estradiol and progesterone secretion (dose-dependent manner) ↓ expression of mRNA HSD3β2 (50 µM) and CYP17A1 (dose-dependent manner) BPS: ↓ testosterone secretion (dose-dependent manner) ↓ expression of mRNA CYP17A1 BPAF: ↓ testosterone secretion (dose-dependent manner) ↑ progesterone secretion ↓ expression of mRNA CYP17A1, HSD3β2 | [79] |
| Type of Bisphenol | Purity (Manufacturer) | Dose (Route) | Animal | Duration of Exposure | Findings | Author |
|---|---|---|---|---|---|---|
| BPA | MERCK | 50 mg/kg/bw (i.p.) | Adult male SD rats | Alternate day until 21 days | Sperm: ↓ sperm count, motility, viability | [75] |
| BPA | Sigma-Aldrich | 50 mg/kg/bw (Oral gavage) | Adult male SD rats | 52 days | Histopathology: Vacuolated and degeneration of germ cells | [80] |
| BPA | Sigma-Aldrich | 5 or 25 mg/kg/bw (Oral gavage) | Adult male Wistar rats | 40 days | Sperm: ↓ total and daily sperm production, integrity of acrosome, plasma membrane and mitochondria activity in sperm | [76] |
| BPA | 99% (Sigma-Aldrich) | 50 mg/kg/bw | Adult male Wistar rats | 14 days | Histopathology: Leydig cells mild edema Spermatocyte depletion Spermatogenesis from weak to arrest. Sperm: ↓ daily sperm production, sperm count, sperm motility | [77] |
| BPA | Sigma-Aldrich | 50 mg/kg/bw (Oral gavage) | Adult male Wistar rats | 30 days | Histopathology: ↓ diameter and epithelial height of seminiferous tubule Atrophy and separation of germinal epithelium Sperm: ↓ sperm count | [64] |
| BPA | Sigma-Aldrich | 200 mg/kg (Oral gavage) | Adult male SD rats | 42 days | Sperm: ↓ sperm count, daily sperm production, motility | [65] |
| BPA | - | 25 mg/kg/bw (i.p.) | Adult male SD rats | Alternate day of 30 days | Histopathology: Degeneration and vacuolation of germ cells Sperm: ↓ sperm count, motility, and viability | [66] |
| BPA | Gracia chengdu chemical technology co. | 200 mg/kg (Oral gavage) | Adult male SD rats | 28 days | Histopathology: ↓ quantity of mature sperm, Longer spermatid Disorganization of germ cells Sperm: ↓ sperm count, motility | [67] |
| BPF | 99% (Santa Cruz Biotechnologie) | 1, 5, 25, 50, 100 mg/kg/bw | Adult male SD rats | 28 days | Histopathology: ↓ germinal epithelial height Absence of sperm in lumen | [68] |
| BPF/BPS | 99% (Santa Cruz Biotechnologie) | 5, 50, 500 mg/kg/bw | Adult male SD rats | 28 days | Histopathology: BPF: Seminiferous tubules irregular Longer spermatid BPS: Absence of sperm in lumen | [69] |
| BPS | 99% (Santa Cruz Biotechnologie) | 25, 50 µg/kg/bw (Oral gavage) | Adult male SD rats | 28 days | Histopathology: ↓ epithelial of seminiferous tubules Spermatid become longer. | [70] |
| BPA/BPAF /BPS | BPA (>99%) BPS (98%) BPAF (98%) | 25, 50, 100 µM | C18-4 spermatogonial cell line | 24–72 h | BPA: ↓ the cell viability after 24 h (100 µM) ↑ DNA damage after 48 h (50 µM) BPAF: ↓ the cell viability after 24 h (50 µM) ↑ DNA damage after 24 h (25 µM) BPS: ↓ the cell viability after 24 h (100 µM) ↑ DNA damage after 24 h (50 µM) | [81] |
| BPA | Sigma-Aldrich | 10 and 50 mg/kg/bw | Adult male wistar rat | - | ↓ occludin (10 mg/kg for 11 weeks and 50 mg/kg for 4 weeks) ↓ nectin-3 (50 mg/kg for 2–4 weeks) | [105] |
| BPA | - | 20 µM | Sertoli cells | - | ↓ occludin (after 48 h) ↓ Z0-1 (after 6 and 48 h) ↓ cells viability after 6 and 48 h ↓ androgen receptor after 6 and 48 h | [82] |
| BPA | Sigma-Aldrich | 25 and 100 µM | Sertoli cells isolated from 20 days of wistar rats | - | ↓ occludin and ZO-1 (both doses) ↑ conexxin (both dosage) Significantly perturb the tight junction barrier at dosage 100 µM (p < 0.05) | [105] |
| BPA | Sigma-aldrich (US) | 10, 50,250 µg/kg/D (Oral gavage) | Sperm of C57BL/6 mice | 8 weeks | ↓ sperm motility (p < 0.05) ↓ Progesterone-induced acrosome reaction (p < 0.05) | [84] |
| BPG, BPAF, BPC, BADGE, BPB | Sigma-aldrich (MO,US) | Ca2+ signal: 10 µM Progesterone-induced Ca2+ signal: BPF: 5 µM BPAF&BPBP: 10 µM BPC, BADGE, BPB: 50 µM | Healthy human semen | - | ↑ Ca2+ signaling ↓ Progesterone-induced Ca2+ signal | [85] |
| Type of Bispheno | Study Design | Study Population (Age) (Project Name) | Country (Sample Population) | Biological Sample | [Bisphenol] Detected in Biological Sample (Mean/ Median) | Findings | Beta Coefficient | Significant Values | Author |
|---|---|---|---|---|---|---|---|---|---|
| BPA | Cross sectional study | Male children (6–11 y.o) Male Adolescents (12–19 y.o) (NHANES Project) | USA (n = 588) | Urine Serum | Mean: male children 1.74 ng/mL (urine) Mean: male adolescents 1.94 ng/mL (urine) | Reproductive hormones: No association between BPA and reproductive hormones in male children across the quartiles. Increased BPA level caused a significant decrease in TT in male adolescents across the quartiles. | Q2: β = −49.34% Q3: β = −36.87% Q4: β = −53.70% | p < 0.05 | [108] |
| BPA | Cross sectional study | Male worker of epoxy resin manufacturer (16–63 y.o) | Shanghai, China (n = 592) | Urine Serum | Median occupational exposure: 685.9 µg/g Cr (urine) Median non-occupational exposure: 4.2 µg/gCr (urine) | Reproductive hormones: Increased level of BPA cause significant increase in:
Increased level of BPA cause significant decreased in levels:
| [109] | ||
| β = 0.0589 ng/mL | p < 0.001 | ||||||||
| β = 0.0293 nmol/L | p < 0.01 | ||||||||
| β = 0.0362 pg/mL | p < 0.001 | ||||||||
| β = −0.0367 ng/mL | p < 0.001 | ||||||||
| β = −0.024 mIU/mL | p < 0.05 | ||||||||
| BPA | Cross sectional study | Male worker of epoxy resin manu-facturer | Guang-dong, China (n = 559) | Serum | Median workers: 8.75 ng/mL (serum) Median non-workers: 3.37 ng/mL (serum) | Reproductive hormones: No association between workers and non-workers on the level of SHBG, TT, INB and AD | [110] | ||
| Increased exposure time caused significant decreased in median AD level among workers. | - | p < 0.001 | |||||||
| Increased exposure time caused significant increase in median SHBG level among workers. | - | p < 0.05 | |||||||
| Increased BPA level caused significant increase in median SHBG level among workers. | β = 2.79 nmol/L | p < 0.05 | |||||||
| Increased of BPA level caused significant decreased in median AD level among workers. | β = −0.18 ng/mL | p < 0.001 | |||||||
| BPA | Cross sectional study | Young men | Denmark (n = 308) | Urine Serum Semen | Median: 3.74 ng/mL (Osm)(urine) | Reproductive hormones: Increased level of BPA caused significant increase in:
| [111] | ||
| β = 0.7 nmol/L | p < 0.01 | ||||||||
| β = 2.7% | p < 0.05 | ||||||||
| β = 3.5% | p < 0.05 | ||||||||
| β = 2.7% | p < 0.05 | ||||||||
| Sperm characteristics: Increased level of BPA caused a significant decreased in percentage of progressive motile spermatozoa across the quartiles. | β = −1.82% | p < 0.01 | |||||||
| BPA | Cross sectional study | Young men (18–23 y.o) | Spain (n = 215) | Urine Serum Semen | Mean: 1.8 µg/g (urine) | Reproductive hormones: Increased level of BPA caused significant increase in LH level across the quartiles. No association between BPA and FSH, FT, SHBG, INB and E2 across the quartile. | β = 0.07 IU/L | p < 0.01 | [112] |
| Sperm characteristics: Increased level of BPA caused significant decreased in sperm characteristic across the quartiles:
| |||||||||
| β = −0.04 Mill./mL | p < 0.01 | ||||||||
| β = −0.05 Mill. | p < 0.01 | ||||||||
| BPA | Retro-spective cohort | Pregnant woman Male chil-dren (8–14 y.o) (ELEMENT project) | Mexico (n = 118) | Urine Urine Serum | Mean: 0.7 ng/mL (urine) Mean: 1.1 ng/mL (urine) | Reproductive hormones: No association between prenatal urinary BPA and the boy sex hormones in the level of SHBG, INB, TT, E2, DHEAS and FT | - | - | [114] |
| No association between child-hood urinary BPA and the boy sex hormones in the level of SHBG, INB, TT, E2, DHEAS and FT | - | - | |||||||
| BPA | Retro-spective cohort | Pregnant woman (Week 18 & 34) Young men (20–22 y.o) | Australia (n = 423) | Serum (mother) Semen | Median: 0.25 µg/L (serum) | Sperm characteristics: Maternal exposure of BPA caused significant changes in sperm characteristics of young men such as increased in the sperm concentration and motility. | - | p < 0.05 | [115] |
| BPA | Pro-spective cohort | Men IVF patient (34.05 y.o) | Slovenia (n = 149) | Semen | Mean: 1.33 ng/mg (urine) | Sperm characteristics: Increase concentration of BPA cause significant decrease in:
| [116] | ||
| β = −0.219, R2 = 0.071 | p = 0.047 | ||||||||
| β = −0.241, R2 = 0.092 | p = 0.039 | ||||||||
| β = −0.273, R2 = 0.075 | p = 0.043 | ||||||||
| β = −0.266, R2 = 0.052 | p = 0.026 | ||||||||
| BPA | Pro-spective cohort | Fertile men (>18 y.o) | Michigan and Texas (n = 418) | Urine Semen | Mean: 0.51 µg/g (urine) | Sperm characteristics: Increased level of BPA caused significant decreased in sperm DNA fragmentation. No association between BPA and semen analysis (sperm morphology, sperm concentration, total sperm count, semen volume). | β = −0.0544 | p < 0.05 | [118] |
| BPA/ BPF/ BPS | Cross sectional study | Young men (18–20 y.o) FEPOS | Denmark (n = 556) | Urine Semen | BPA (urine) Q1: <0.68 ng/mL Q3: 1.3–2.74 ng/mL BPF (urine) Q1: <0.06 ng/mL Q3: 0.14–0.34 ng/mL BPS Q1: <0.03 ng/mL Q3: 0.06–0.17 ng/mL (urine) | Sperm characteristics: Percentage of motile spermatozoa in Q3 is significantly higher compared to Q1 in BPA and BPF exposures. Volume of semen per ejaculate in Q3 is significantly lower compared to Q1 in BPA and BPS exposures. No association between (BPA, BPF, and BPS) with the other semen analysis (sperm concentration, total sperm count, normal sperm morphology, motility and ejaculate volume). | β = 1.07% β = −0.87 mL | p < 0.05 p < 0.05 | [119] |
| BPS | Cross sectional study | Infertile patient (18–56 y.o) | Boston, MA, USA (n = 158) | Urine Semen | Mean BPA: 0.77 µg/L (urine) Mean BPS: 0.37 µg/L (urine) | Semen characteristics: Volume of semen per ejaculate in Q2 is significantly higher compared to Q1 in BPS exposure. Sperm concentration in Q3 is significantly lower compared to Q1 in BPS exposure. Increased level of BPS caused significant decreased in sperm quality among obese/overweight men (BMI >25 kg/m2):
| β = 3.0 mL β = −29.2 mil/mL | p < 0.05 p < 0.05 | [117] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shamhari, A.‘A.; Abd Hamid, Z.; Budin, S.B.; Shamsudin, N.J.; Taib, I.S. Bisphenol A and Its Analogues Deteriorate the Hormones Physiological Function of the Male Reproductive System: A Mini-Review. Biomedicines 2021, 9, 1744. https://doi.org/10.3390/biomedicines9111744
Shamhari A‘A, Abd Hamid Z, Budin SB, Shamsudin NJ, Taib IS. Bisphenol A and Its Analogues Deteriorate the Hormones Physiological Function of the Male Reproductive System: A Mini-Review. Biomedicines. 2021; 9(11):1744. https://doi.org/10.3390/biomedicines9111744
Chicago/Turabian StyleShamhari, Asma’ ‘Afifah, Zariyantey Abd Hamid, Siti Balkis Budin, Nurul Jehan Shamsudin, and Izatus Shima Taib. 2021. "Bisphenol A and Its Analogues Deteriorate the Hormones Physiological Function of the Male Reproductive System: A Mini-Review" Biomedicines 9, no. 11: 1744. https://doi.org/10.3390/biomedicines9111744
APA StyleShamhari, A. ‘A., Abd Hamid, Z., Budin, S. B., Shamsudin, N. J., & Taib, I. S. (2021). Bisphenol A and Its Analogues Deteriorate the Hormones Physiological Function of the Male Reproductive System: A Mini-Review. Biomedicines, 9(11), 1744. https://doi.org/10.3390/biomedicines9111744