
Zinc: Multidimensional Effects on Living Organisms




Abstract
:1. Introduction
2. Prokaryotes
2.1. Zinc as Antimicrobial
2.2. Zinc Oxide Nanoparticles as Antibacterial
2.3. A Brief Overview of Zinc Homeostasis in Bacterial Pathogens
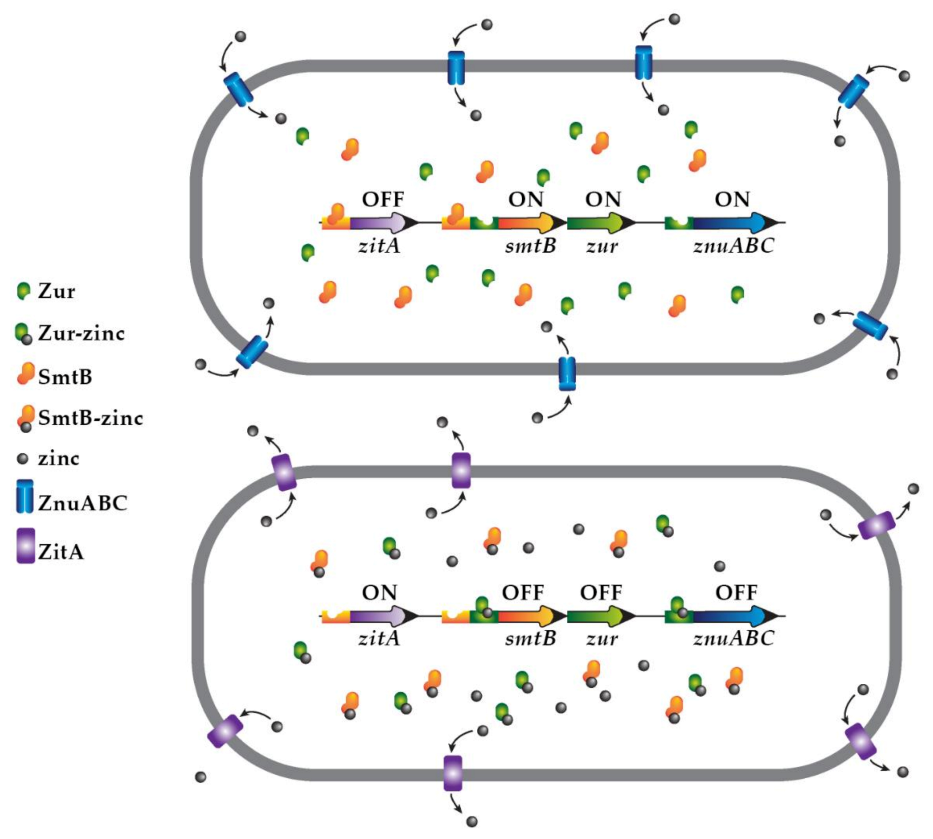
2.4. Zinc-Driven Regulation of Gene Expression
2.5. Zinc as Adjuvant to Antibiotic Treatment
3. Eukaryotes
3.1. Importance of Zinc in Eukaryotic Cells
3.2. Zinc-Rich Cells
3.3. Zinc Transporters
3.4. Zinc Interactions with Various Proteins
3.4.1. Enzymes
3.4.2. Signaling Proteins
3.4.3. Cytoskeletal Proteins
3.5. Zinc-Induced Cell Death
Zinc Mobilization in Apoptosis
3.6. Zinc Chelators
Chelators That Bind Zinc and Their Effects on Cells
3.7. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andreini, I.; Banci, C.L.; Bertini, I.; Rosato, A. Zinc through the three domains of life. J. Proteome Res. 2006, 5, 3173–3178. [Google Scholar] [CrossRef] [PubMed]
- Maxfield, L.; Crane, J.S. Zinc Deficiency; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Andreini, C.; Bertini, I. A bioinformatics view of zinc enzymes. J. Inorg. Biochem. 2012, 111, 150–156. [Google Scholar] [CrossRef]
- Banci, L.; Bertini, I.; Ciofi-Baffoni, S.; Finney, L.A.; Outten, C.E.; O’Halloran, T.V. A new zinc-protein coordination site in intracellular metal Trafficking: Solution structure of the Apo and Zn(II) forms of ZntA(46–118). J. Mol. Biol. 2002, 323, 883–897. [Google Scholar] [CrossRef]
- Debela, M.; Magdolen, V.; Grimminger, V.; Sommerhoff, C.; Messerschmidt, A.; Huber, R.; Friedrich, R.; Bode, W.; Goettig, P. Crystal structures of human tissue kallikrein 4: Activity modulation by a specific zinc binding site. J. Mol. Biol. 2006, 362, 1094–1107. [Google Scholar] [CrossRef]
- Mikhaylina, A.; Ksibe, A.Z.; Scanlan, D.J.; Blindauer, C.A. Bacterial zinc uptake regulator proteins and their regulons. Biochem. Soc. Trans. 2018, 46, 983–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, J.E. Zinc proteins: Enzymes, storage proteins, transcription factors, and replication proteins. Annu. Rev. Biochem. 1992, 61, 897–946. [Google Scholar] [CrossRef] [PubMed]
- Loh, S.N. The missing Zinc: p53 misfolding and cancer. Metallomics 2010, 2, 442–449. [Google Scholar] [CrossRef]
- Namuswe, F.; Berg, J.M. Secondary interactions involving zinc-bound ligands: Roles in structural stabilization and macromolecular interactions. J. Inorg. Biochem. 2012, 111, 146–149. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, S.E.; Ceder, S.; Bykov, V.J.N.; Wiman, K.G. p53 as a hub in cellular redox regulation and therapeutic target in cancer. J. Mol. Cell Biol. 2019, 11, 330–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buberg, M.L.; Witsø, I.L.; L’Abée-Lund, T.M.; Wasteson, Y. Zinc and Copper Reduce Conjugative Transfer of Resistance Plasmids from Extended-Spectrum Beta-Lactamase-Producing Escherichia coli. Microb. Drug Resist. 2020, 26, 842–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blencowe, D.K.; Morby, A.P. Zn(II) metabolism in prokaryotes. FEMS Microbiol. Rev. 2003, 27, 291–311. [Google Scholar] [CrossRef]
- Eide, D. Molecular biology of iron and zinc uptake in eukaryotes. Curr. Opin. Cell Biol. 1997, 9, 573–577. [Google Scholar] [CrossRef]
- Hennigar, S.R.; McClung, J.P. Nutritional immunity: Starving pathogens of trace minerals. Am. J. Lifestyle Med. 2016, 10, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Wooldridge, K.G.; Williams, P.H. Iron uptake mechanisms of pathogenic bacteria. FEMS Microbiol. Rev. 1993, 12, 325–348. [Google Scholar] [CrossRef]
- Weinberg, E.D. Nutritional immunity. Host’s attempt to withold iron from microbial invaders. JAMA 1975, 231, 39–41. [Google Scholar] [CrossRef]
- Schade, A.L.; Caroline, L. Raw hen egg white and the role of iron in growth inhibition of Shigella dysenteriae, Staphylococcus aureus, Escherichia coli and Saccharomyces cerevisiae. Science 1944, 100, 14–15. [Google Scholar] [CrossRef]
- Schade, A.L.; Caroline, L. An Iron-binding Component in Human Blood Plasma. Science 1946, 104, 340–341. [Google Scholar] [CrossRef]
- Weinberg, E.D. Iron withholding: A defense against infection and neoplasia. Physiol. Rev. 1984, 64, 65–102. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, E.D. Iron depletion: A defense against intracellular infection and neoplasia. Life Sci. 1992, 50, 1289–1297. [Google Scholar] [CrossRef]
- Kehl-Fie, T.E.; Skaar, E.P. Nutritional immunity beyond iron: A role for manganese and zinc. Curr. Opin. Chem. Biol. 2010, 14, 218–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbin, B.D.; Seeley, E.H.; Raab, A.; Feldmann, J.; Miller, M.R.; Torres, V.J.; Anderson, K.L.; Dattilo, B.M.; Dunman, P.M.; Gerads, R.; et al. Metal chelation and inhibition of bacterial growth in tissue abscesses. Science 2008, 319, 962–965. [Google Scholar] [CrossRef]
- Hood, M.I.; Mortensen, B.L.; Moore, J.L.; Zhang, Y.; Kehl-Fie, T.E.; Sugitani, N.; Chazin, W.J.; Caprioli, R.M.; Skaar, E.P. Identification of an Acinetobacter baumannii zinc acquisition system that facilitates resistance to calprotectin-mediated zinc sequestration. PLoS Pathog. 2012, 8, e1003068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen–host interface. Nat. Rev. Genet. 2012, 10, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, Z.R.; Skaar, E.P. Nutrient Zinc at the Host–Pathogen Interface. Trends Biochem. Sci. 2019, 44, 1041–1056. [Google Scholar] [CrossRef] [PubMed]
- Craig, G.M.; Evans, S.J.; Brayshaw, B.J.; Raina, S.K. A study of serum zinc, albumin, alpha-2-macroglobulin and transferrin levels in acute and long stay elderly hospital patients. Postgrad. Med. J. 1990, 66, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Foote, J.W.; Delves, H.T. Albumin bound and alpha 2-macroglobulin bound zinc concentrations in the sera of healthy adults. J. Clin. Pathol. 1984, 37, 1050–1054. [Google Scholar] [CrossRef] [Green Version]
- Foote, J.W.; Delves, H.T. Distribution of zinc amongst human serum globulins determined by gel filtration-affinity chromatography and atomic-absorption spectrophotometry. Analyst 1984, 109, 709–711. [Google Scholar] [CrossRef]
- Zygiel, E.M.; Nolan, E.M. Transition Metal Sequestration by the Host-Defense Protein Calprotectin. Annu. Rev. Biochem. 2018, 87, 621–643. [Google Scholar] [CrossRef]
- Damo, S.M.; Kehl-Fie, T.E.; Sugitani, N.; Holt, M.E.; Rathi, S.; Murphy, W.J.; Zhang, Y.; Betz, C.; Hench, L.; Günter, F.; et al. Molecular basis for manganese sequestration by calprotectin and roles in the innate immune response to invading bacterial pathogens. Proc. Natl. Acad. Sci. USA 2013, 110, 3841–3846. [Google Scholar] [CrossRef] [Green Version]
- Nakashige, T.G.; Stephan, J.R.; Cunden, L.S.; Brophy, M.B.; Wommack, A.J.; Keegan, B.C.; Shearer, J.M.; Nolan, E.M. The hexahistidine motif of host-defense protein human calprotectin contributes to zinc withholding and its functional versatility. J. Am. Chem. Soc. 2016, 138, 12243–12251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephan, J.R.; Nolan, E.M. Calcium-induced tetramerization and zinc chelation shield human calprotectin from degradation by host and bacterial extracellular proteases. Chem. Sci. 2016, 7, 1962–1975. [Google Scholar] [CrossRef] [Green Version]
- Sohnle, P.G.; Collins-Lech, C.; Wiessner, J.H. The Zinc-Reversible Antimicrobial Activity of Neutrophil Lysates and Abscess Fluid Supernatants. J. Infect. Dis. 1991, 164, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Sohnle, P.G.; Collins-Lech, C.; Wiessner, J.H. Antimicrobial Activity of an Abundant Calcium-Binding Protein in the Cytoplasm of Human Neutrophils. J. Infect. Dis. 1991, 163, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Steinbakk, M.; Naess-Andresen, C.F.; Lingaas, E.; Dale, I.; Brandtzaeg, P.; Fagerhol, M.K. Antimicrobial actions of calcium binding leucocyte L1 protein, calprotectin. Lancet 1990, 336, 763–765. [Google Scholar] [CrossRef]
- Haase, H.; Rink, L. Multiple impacts of zinc on immune function. Metallomics 2014, 6, 1175–1180. [Google Scholar] [CrossRef]
- Haase, H.; Rink, L. Zinc signals and immune function. BioFactors 2014, 40, 27–40. [Google Scholar] [CrossRef]
- Achouiti, A.; Vogl, T.; Urban, C.F.; Rohm, M.; Hommes, T.J.; van Zoelen, M.A.; Florquin, S.; Roth, J.; van ’t Veer, C.; de Vos, A.F.; et al. Myeloid-related protein-14 contributes to protective immunity in gram-negative pneumonia derived sepsis. PLoS Pathog. 2012, 8, e1002987. [Google Scholar] [CrossRef] [Green Version]
- Lopez, C.A.; Beavers, W.N.; Weiss, A.; Knippel, R.J.; Zackular, J.P.; Chazin, W.; Skaar, E.P. The immune protein calprotectin impacts Clostridioides difficile metabolism through zinc limitation. MBio 2019, 10, e02289–e19. [Google Scholar] [CrossRef] [Green Version]
- Cunden, L.S.; Brophy, M.B.; Rodriguez, G.E.; Flaxman, H.A.; Nolan, E.M. Biochemical and functional evaluation of the intramolecular disulfide bonds in the zinc-chelating antimicrobial protein human S100A7 (psoriasin). Biochemistry 2017, 56, 5726–5738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crane, J.K.; Broome, J.E.; Reddinger, R.M.; Werth, B.B. Zinc protects against Shiga-toxigenic Escherichia coli by acting on host tissues as well as on bacteria. BMC Microbiol. 2014, 14, 145. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, P.; Bolick, D.T.; Roche, J.K.; Noronha, F.; Pinheiro, C.; Kolling, G.L.; Lima, A.; Guerrant, R.L. The micronutrient zinc inhibits EAEC strain 042 adherence, biofilm formation, virulence gene expression, and epithelial cytokine responses benefiting the infected host. Virulence 2013, 4, 624–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, J.; Arentz, S.; Goldenberg, J.; Yang, G.; Beardsley, J.; Mertz, D.; Leeder, S. Rapid review protocol: Zinc for the prevention or treatment of COVID-19 and other coronavirus-related respiratory tract infections. Integr. Med. Res. 2020, 9, 100457. [Google Scholar] [CrossRef] [PubMed]
- te Velthuis, A.J.; van den Worm, S.H.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn(2+) inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef] [PubMed]
- Cordingley, M.G.; Register, R.B.; Callahan, P.L.; Garsky, V.M.; Colonno, R.J. Cleavage of small peptides in vitro by human rhinovirus 14 3C protease expressed in Escherichia coli. J. Virol. 1989, 63, 5037–5045. [Google Scholar] [CrossRef] [Green Version]
- Fenstermacher, K.J.; DeStefano, J.J. Mechanism of HIV reverse transcriptase inhibition by zinc: Formation of a highly stable enzyme-(primer-template) complex with profoundly diminished catalytic activity. J. Biol. Chem. 2011, 286, 40433–40442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crane, J.K.; Cheema, M.B.; Olyer, M.A.; Sutton, M.D. Zinc blockade of SOS response inhibits horizontal transfer of antibiotic resistance genes in enteric bacteria. Front. Cell Infect. Microbiol. 2018, 8, 410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bunnell, B.E.; Escobar, J.F.; Bair, K.L.; Sutton, M.D.; Crane, J.K. Zinc blocks SOS-induced antibiotic resistance via inhibition of RecA in Escherichia coli. PLoS ONE 2017, 12, e0178303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botella, H.; Peyron, P.; Levillain, F.; Poincloux, R.; Poquet, Y.; Brandli, I.; Wang, C.; Tailleux, L.; Tilleul, S.; Charriere, G.M.; et al. Mycobacterial p(1)-type ATPases mediate resistance to zinc poisoning in human macrophages. Cell Host Microbe. 2011, 10, 248–259. [Google Scholar] [CrossRef]
- Ong, C.L.; Gillen, C.M.; Barnett, T.C.; Walker, M.J.; McEwan, A.G. An antimicrobial role for zinc in innate immune defense against group A streptococcus. J. Infect. Dis. 2014, 209, 1500–1508. [Google Scholar] [CrossRef] [Green Version]
- Kapetanovic, R.; Bokil, N.J.; Achard, M.E.; Ong, C.L.; Peters, K.M.; Stocks, C.J.; Phan, M.D.; Monteleone, M.; Schroder, K.; Irvine, K.M.; et al. Salmonella employs multiple mechanisms to subvert the TLR-inducible zinc-mediated antimicrobial response of human macrophages. FASEB J. 2016, 30, 1901–1912. [Google Scholar] [CrossRef] [Green Version]
- McDevitt, C.A.; Ogunniyi, A.D.; Valkov, E.; Lawrence, M.C.; Kobe, B.; McEwan, A.G.; Paton, J.C. A molecular mechanism for bacterial susceptibility to zinc. PLoS Pathog. 2011, 7, e1002357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botella, H.; Stadthagen, G.; Lugo-Villarino, G.; De Chastellier, C.; Neyrolles, O. Metallobiology of host–pathogen interactions: An intoxicating new insight. Trends Microbiol. 2012, 20, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Eijkelkamp, B.A.; Morey, J.R.; Ween, M.P.; Ong, C.L.; McEwan, A.G.; Paton, J.C.; McDevitt, C.A. Extracellular zinc competitively inhibits manganese uptake and compromises oxidative stress management in Streptococcus pneumoniae. PLoS ONE 2014, 9, e89427. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.-L.Y.; Walker, M.J.; McEwan, A.G. Zinc disrupts central carbon metabolism and capsule biosynthesis in Streptococcus pyogenes. Sci. Rep. 2015, 5, 10799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makthal, N.; Kumaraswami, M. Zinc’ing it out: Zinc homeostasis mechanisms and their impact on the pathogenesis of human pathogen group A streptococcus. Metallomics 2017, 9, 1693–1702. [Google Scholar] [CrossRef]
- Crane, J.K.; Olson, R.A.; Jones, H.M.; Duffey, M.E. Release of ATP during host cell killing by enteropathogenic E. coli and its role as a secretory mediator. Am. J. Physiol. Gastrointest. Liver Physiol. 2002, 283, G74–G86. [Google Scholar] [CrossRef] [Green Version]
- Crane, J.K.; Shulgina, I.; Naeher, T.M. Ecto-5′-nucleotidase and intestinal ion secretion by enteropathogenic Escherichia coli. Purinergic Signal. 2007, 3, 233–246. [Google Scholar] [CrossRef] [Green Version]
- Barffour, M.A.; Hinnouho, G.M.; Wessells, K.R.; Kounnavong, S.; Ratsavong, K.; Sitthideth, D.; Bounheuang, B.; Sengnam, K.; Chanhthavong, B.; Arnold, C.D.; et al. Effects of therapeutic zinc supplementation for diarrhea and two preventive zinc supplementation regimens on the incidence and duration of diarrhea and acute respiratory tract infections in rural Laotian children: A randomized controlled trial. J. Glob. Health 2020, 10, 010424. [Google Scholar] [CrossRef]
- Bhutta, Z.A.; Bird, S.M.; Black, R.E.; Brown, K.H.; Gardner, J.M.; Hidayat, A.; Khatun, F.; Martorell, R.; Ninh, N.X.; Penny, M.E.; et al. Therapeutic effects of oral zinc in acute and persistent diarrhea in children in developing countries: Pooled analysis of randomized controlled trials. Am. J. Clin. Nutr. 2000, 72, 1516–1522. [Google Scholar]
- Metzler-Zebeli, B.U.; Caine, W.R.; McFall, M.; Miller, B.; Ward, T.L.; Kirkwood, R.N.; Mosenthin, R. Supplementation of diets for lactating sows with zinc amino acid complex and gastric nutriment-intubation of suckling pigs with zinc methionine on mineral status, intestinal morphology and bacterial translocation in lipopolysaccharide-challenged weaned pigs. J. Anim. Physiol. Anim. Nutr. (Berl) 2010, 94, 237–249. [Google Scholar]
- Crane, J.K.; Naeher, T.M.; Shulgina, I.; Zhu, C.; Boedeker, E.C. Effect of zinc in enteropathogenic Escherichia coli infection. Infect. Immun. 2007, 75, 5974–5984. [Google Scholar] [CrossRef] [Green Version]
- Magallon, J.; Chiem, K.; Tran, T.; Ramirez, M.S.; Jimenez, V.; Tolmasky, M.E. Restoration of susceptibility to amikacin by 8-hydroxyquinoline analogs complexed to zinc. PLoS ONE 2019, 14, e0217602. [Google Scholar] [CrossRef] [Green Version]
- Crane, J.K.; Byrd, I.W.; Boedeker, E.C. Virulence Inhibition by Zinc in Shiga-ToxigenicEscherichia coli. Infect. Immun. 2011, 79, 1696–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cirz, R.T.; Romesberg, F.E. Induction and Inhibition of Ciprofloxacin Resistance-Conferring Mutations in Hypermutator Bacteria. Antimicrob. Agents Chemother. 2006, 50, 220–225. [Google Scholar] [CrossRef] [Green Version]
- Recacha, E.; Machuca, J.; De Alba, P.D.; Ramos-Güelfo, M.; Pérez, F.M.D.; Beltrán, J.R.; Blázquez, J.; Pascual, A.; Martínez, J.M.R. Quinolone Resistance Reversion by Targeting the SOS Response. MBio 2017, 8, e00971-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plaut, R.D.; Beaber, J.W.; Zemansky, J.; Kaur, A.P.; George, M.; Biswas, B.; Henry, M.; Bishop-Lilly, K.A.; Mokashi, V.; Hannah, R.M.; et al. Genetic evidence for the involvement of the S-layer protein gene sap and the sporulation genes spo0A, spo0B, and spo0F in phage AP50c infection of Bacillus anthracis. J. Bacteriol. 2014, 196, 1143–1154. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.L.; Tran, T.; Alam, J.Y.; Herron, S.R.; Ramirez, M.S.; Tolmasky, M.E. Inhibition of aminoglycoside 6’-N-acetyltransferase type Ib by zinc: Reversal of amikacin resistance in Acinetobacter baumannii and Escherichia coli by a zinc ionophore. Antimicrob. Agents Chemother. 2014, 58, 4238–4241. [Google Scholar] [CrossRef] [Green Version]
- Kelly, P.; Besa, E.; Zyambo, K.; Louis-Auguste, J.; Lees, J.; Banda, T.; Soko, R.; Banda, R.; Amadi, B.; Watson, A. Endomicroscopic and transcriptomic analysis of impaired barrier function and Mmalabsorption in environmental enteropathy. PLoS Negl. Trop. Dis. 2016, 10, e0004600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korpe, P.S.; Petri, W.A., Jr. Environmental enteropathy: Critical implications of a poorly understood condition. Trends Mol. Med. 2012, 18, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Choudhry, N.; Scott, F.; Edgar, M.; Sanger, G.J.; Kelly, P. Reversal of pathogen-induced barrier defects in intestinal epithelial cells by contra-pathogenicity agents. Dig. Dis. Sci. 2021, 66, 88–104. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Labrie, J.; Tremblay, Y.D.; Haine, D.; Mourez, M.; Jacques, M. Zinc as an agent for the prevention of biofilm formation by pathogenic bacteria. J. Appl. Microbiol. 2013, 115, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Kolodziejczak-Radzimska, A.; Jesionowski, T. Zinc oxide-from synthesis to application: A review. Materials 2014, 7, 2833–2881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morshedtalab, Z.; Rahimi, G.; Emami-Nejad, A.; Farasat, A.; Mohammadbeygi, A.; Ghaedamini, N.; Negahdary, M. Antibacterial assessment of zinc sulfide nanoparticles against Streptococcus pyogenes and Acinetobacter baumannii. Curr. Top Med. Chem. 2020, 20, 1042–1055. [Google Scholar] [CrossRef]
- Sanchez-Lopez, E.; Gomes, D.; Esteruelas, G.; Bonilla, L.; Lopez-Machado, A.L.; Galindo, R.; Cano, A.; Espina, M.; Ettcheto, M.; Camins, A.; et al. Metal-based nanoparticles as antimicrobial agents: An overview. NanoMaterials 2020, 10, 292. [Google Scholar] [CrossRef] [Green Version]
- Ye, Q.; Chen, W.; Huang, H.; Tang, Y.; Wang, W.; Meng, F.; Wang, H.; Zheng, Y. Iron and zinc ions, potent weapons against multidrug-resistant bacteria. Appl. Microbiol. Biotechnol. 2020, 104, 5213–5227. [Google Scholar] [CrossRef]
- Slavin, Y.N.; Asnis, J.; Hafeli, U.O.; Bach, H. Metal nanoparticles: Understanding the mechanisms behind antibacterial activity. J. Nanobiotechnol. 2017, 15, 65. [Google Scholar] [CrossRef] [PubMed]
- Stensberg, M.C.; Wei, Q.; McLamore, E.S.; Porterfield, D.M.; Wei, A.; Sepulveda, M.S. Toxicological studies on silver nanoparticles: Challenges and opportunities in assessment, monitoring and imaging. Nanomedicine (Lond) 2011, 6, 879–898. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, S.; Krishnakumar, C.; Arulmozhi, P.; Mahadevan, S.; Parameswari, N. Biosynthesis, characterization and antimicrobial activities of zinc oxide nanoparticles from leaf extract of Glycosmis pentaphylla (Retz.) DC. Microb. Pathog. 2018, 116, 44–48. [Google Scholar] [CrossRef]
- Singh, P.; Nanda, A. Antimicrobial and antifungal potential of zinc oxide nanoparticles in comparison to conventional zinc oxide particles. J. Chem. Pharm. Res. 2013, 5, 457–463. [Google Scholar]
- Yu, J.; Zhang, W.; Li, Y.; Wang, G.; Yang, L.; Jin, J.; Chen, Q.; Huang, M. Synthesis, characterization, antimicrobial activity and mechanism of a novel hydroxyapatite whisker/nano zinc oxide biomaterial. Biomed. Mater. 2014, 10, 015001. [Google Scholar] [CrossRef]
- Sirelkhatim, A.; Mahmud, S.; Seeni, A.; Kaus, N.H.M.; Ann, L.C.; Bakhori, S.K.M.; Hasan, H.; Mohamad, D. Review on zinc oxide nanoparticles: Antibacterial activity and toxicity mechanism. Nanomicro. Lett. 2015, 7, 219–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sukri, S.; Shameli, K.; Wong, M.; Teow, S.; Ismail, N. Cytotoxicity and antibacterial activities of plant-mediated synthesized zinc oxide (ZnO) nanoparticles using Punica granatum (pomegranate) fruit peels extract. J. Mol. Struct. 2019, 1189, 57–65. [Google Scholar] [CrossRef]
- Happy, A.; Soumya, M.; Venkat Kumar, S.; Rajeshkumar, S.; Sheba, R.D.; Lakshmi, T.; Deepak Nallaswamy, V. Phyto-assisted synthesis of zinc oxide nanoparticles using Cassia alata and its antibacterial activity against Escherichia coli. Biochem. Biophys. Rep. 2019, 17, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Hantke, K. Bacterial zinc uptake and regulators. Curr. Opin. Microbiol. 2005, 8, 196–202. [Google Scholar] [CrossRef]
- Waldron, K.J.; Robinson, N.J. How do bacterial cells ensure that metalloproteins get the correct metal? Nat. Rev. Genet. 2009, 7, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Chandrangsu, P.; Rensing, C.; Helmann, J.D. Metal homeostasis and resistance in bacteria. Nat. Rev. Genet. 2017, 15, 338–350. [Google Scholar] [CrossRef]
- Outten, C.E.; O’Halloran, T.V. Femtomolar sensitivity of metalloregulatory proteins controlling zinc homeostasis. Science 2001, 292, 2488–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldron, K.J.; Rutherford, J.C.; Ford, D.; Robinson, N.J. Metalloproteins and metal sensing. Nature 2009, 460, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Grim, K.P.; Radin, J.N.; Solorzano, P.K.P.; Morey, J.R.; Frye, K.A.; Ganio, K.; Neville, S.L.; McDevitt, C.A.; Kehl-Fie, T.E. Intracellular accumulation of staphylopine can sensitize Staphylococcus aureus to host-imposed zinc starvation by chelation-independent toxicity. J. Bacteriol. 2020, 202, e00014–e00020. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, D.A.; Wang, J.; Giedroc, D.P. Bacterial Strategies to Maintain Zinc Metallostasis at the Host-Pathogen Interface. J. Biol. Chem. 2016, 291, 20858–20868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blindauer, C.A. Advances in the molecular understanding of biological zinc transport. Chem. Commun. 2015, 51, 4544–4563. [Google Scholar] [CrossRef] [Green Version]
- Hesse, L.E.; Lonergan, Z.R.; Beavers, W.N.; Skaar, E.P. The Acinetobacter baumannii Znu system overcomes host-imposed nutrient zinc limitation. Infect. Immun. 2019, 87, e00746–e19. [Google Scholar] [CrossRef]
- Lonergan, Z.R.; Nairn, B.L.; Wang, J.; Hsu, Y.P.; Hesse, L.E.; Beavers, W.N.; Chazin, W.J.; Trinidad, J.C.; VanNieuwenhze, M.S.; Giedroc, D.P.; et al. An Acinetobacter baumannii, zinc-regulated peptidase maintains cell wall integrity during immune-mediated nutrient sequestration. Cell Rep. 2019, 26, 2009–2018.e6. [Google Scholar] [CrossRef] [Green Version]
- Schneider, E.; Hunke, S. ATP-binding-cassette (ABC) transport systems: Functional and structural aspects of the ATP-hydrolyzing subunits/domains. FEMS Microbiol. Rev. 1998, 22, 1–20. [Google Scholar] [CrossRef]
- Patzer, S.I.; Hantke, K. The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli. Mol. Microbiol. 1998, 28, 1199–1210. [Google Scholar] [CrossRef]
- Berntsson, R.P.; Smits, S.H.; Schmitt, L.; Slotboom, D.J.; Poolman, B. A structural classification of substrate-binding proteins. FEBS Lett. 2010, 584, 2606–2617. [Google Scholar] [CrossRef] [Green Version]
- van der Heide, T.; Poolman, B. ABC transporters: One, two or four extracytoplasmic substrate-binding sites? EMBO Rep. 2002, 3, 938–943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warner, D.M.; Levy, S.B. SoxS Increases the Expression of the Zinc Uptake System ZnuACB in an Escherichia coli Murine Pyelonephritis Model. J. Bacteriol. 2011, 194, 1177–1185. [Google Scholar] [CrossRef] [PubMed]
- Panina, E.M.; Mironov, A.A.; Gelfand, M.S. Comparative genomics of bacterial zinc regulons: Enhanced ion transport, pathogenesis, and rearrangement of ribosomal proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 9912–9917. [Google Scholar] [CrossRef] [Green Version]
- Bellotti, D.; Rowinska-Zyrek, M.; Remelli, M. Novel insights into the metal binding ability of ZinT periplasmic protein from Escherichia coli and Salmonella enterica. Dalton Trans. 2020, 49, 9393–9403. [Google Scholar] [CrossRef]
- Gabbianelli, R.; Scotti, R.; Ammendola, S.; Petrarca, P.; Nicolini, L.; Battistoni, A. Role of ZnuABC and ZinT in Escherichia coli O157:H7 zinc acquisition and interaction with epithelial cells. BMC Microbiol. 2011, 11, 36. [Google Scholar] [CrossRef] [PubMed]
- Hubert, K.; Devos, N.; Mordhorst, I.; Tans, C.; Baudoux, G.; Feron, C.; Goraj, K.; Tommassen, J.; Vogel, U.; Poolman, J.T.; et al. ZnuD, a potential candidate for a simple and universal Neisseria meningitidis vaccine. Infect. Immun. 2013, 81, 1915–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stork, M.; Grijpstra, J.; Bos, M.P.; Manas Torres, C.; Devos, N.; Poolman, J.T.; Chazin, W.J.; Tommassen, J. Zinc piracy as a mechanism of Neisseria meningitidis for evasion of nutritional immunity. PLoS Pathog. 2013, 9, e1003733. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, M.; Stork, M. Plasmid-Encoded Iron Uptake Systems. In Plasmids. Biology and Impact in Biotechnology and Discovery; Tolmasky, M.E., Alonso, J., Eds.; ASM Press: Washington, DC, USA, 2014; pp. 577–597. [Google Scholar]
- Stojnev, T.; Harichova, J.; Ferianc, P.; Nystrom, T. Function of a novel cadmium-induced yodA protein in Escherichia coli. Curr. Microbiol. 2007, 55, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Calmettes, C.; Ing, C.; Buckwalter, C.M.; El Bakkouri, M.; Chieh-Lin Lai, C.; Pogoutse, A.; Gray-Owen, S.D.; Pomes, R.; Moraes, T.F. The molecular mechanism of Zinc acquisition by the neisserial outer-membrane transporter ZnuD. Nat. Commun. 2015, 6, 7996. [Google Scholar] [CrossRef] [Green Version]
- Pederick, V.G.; Eijkelkamp, B.A.; Begg, S.L.; Ween, M.P.; McAllister, L.J.; Paton, J.C.; McDevitt, C.A. ZnuA and zinc homeostasis in Pseudomonas aeruginosa. Sci. Rep. 2015, 5, 13139. [Google Scholar] [CrossRef] [PubMed]
- Qamsari, M.M.; Rasooli, I.; Chaudhuri, S.; Astaneh, S.D.A.; Schryvers, A.B. Hybrid antigens expressing surface loops of ZnuD from Acinetobacter baumannii Is capable of inducing protection against infection. Front. Immunol. 2020, 11, 158. [Google Scholar] [CrossRef]
- Plumptre, C.D.; Eijkelkamp, B.A.; Morey, J.R.; Behr, F.; Counago, R.M.; Ogunniyi, A.D.; Kobe, B.; O’Mara, M.L.; Paton, J.C.; McDevitt, C.A. AdcA and AdcAII employ distinct zinc acquisition mechanisms and contribute additively to zinc homeostasis in Streptococcus pneumoniae. Mol. Microbiol. 2014, 91, 834–851. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.C.; Garcia-Herrero, A.; Johanson, T.H.; Krewulak, K.D.; Lau, C.K.; Peacock, R.S.; Slavinskaya, Z.; Vogel, H.J. Siderophore uptake in bacteria and the battle for iron with the host; a bird’s eye view. Biometals 2010, 23, 601–611. [Google Scholar] [CrossRef]
- Ghssein, G.; Brutesco, C.; Ouerdane, L.; Fojcik, C.; Izaute, A.; Wang, S.; Hajjar, C.; Lobinski, R.; Lemaire, D.; Richaud, P.; et al. Biosynthesis of a broad-spectrum nicotianamine-like metallophore in Staphylococcus aureus. Science 2016, 352, 1105–1109. [Google Scholar] [CrossRef]
- Grim, K.P.; San Francisco, B.; Radin, J.N.; Brazel, E.B.; Kelliher, J.L.; Parraga Solorzano, P.K.; Kim, P.C.; McDevitt, C.A.; Kehl-Fie, T.E. The metallophore staphylopine enables Staphylococcus aureus to compete with the host for zinc and overcome nutritional immunity. MBio 2017, 8, e01281–e17. [Google Scholar] [CrossRef] [Green Version]
- Lhospice, S.; Gomez, N.O.; Ouerdane, L.; Brutesco, C.; Ghssein, G.; Hajjar, C.; Liratni, A.; Wang, S.; Richaud, P.; Bleves, S.; et al. Pseudomonas aeruginosa zinc uptake in chelating environment is primarily mediated by the metallophore pseudopaline. Sci. Rep. 2017, 7, 17132. [Google Scholar] [CrossRef] [Green Version]
- McFarlane, J.S.; Lamb, A.L. Biosynthesis of an Opine Metallophore byPseudomonas aeruginosa. Biochemistry 2017, 56, 5967–5971. [Google Scholar] [CrossRef]
- Mastropasqua, M.C.; D’Orazio, M.; Cerasi, M.; Pacello, F.; Gismondi, A.; Canini, A.; Canuti, L.; Consalvo, A.; Ciavardelli, D.; Chirullo, B.; et al. Growth of Pseudomonas aeruginosa in zinc poor environments is promoted by a nicotianamine-related metallophore. Mol. Microbiol. 2017, 106, 543–561. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Chai, J.; Love, J.; Fu, D. Selective electrodiffusion of zinc ions in a Zrt-, Irt-like protein, ZIPB. J. Biol. Chem. 2010, 285, 39013–39020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudek, L.; Pearson, L.A.; Michalczyk, A.; Neilan, B.A.; Ackland, M.L. Functional characterization of the twin ZIP/SLC39 metal transporters, NpunF3111 and NpunF2202 in Nostoc punctiforme. Appl. Microbiol. Biotechnol. 2013, 97, 8649–8662. [Google Scholar] [CrossRef] [PubMed]
- Cerasi, M.; Liu, J.Z.; Ammendola, S.; Poe, P.; Petrarca, A.J.; Pesciaroli, M.; Pasquali, P.; Raffatellu, M.; Battistoni, A. The ZupT transporter plays an important role in zinc homeostasis and contributes to Salmonella enterica Virulence. Metallomics 2014, 6, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Grass, G.; Franke, S.; Taudte, N.; Nies, D.H.; Kucharski, L.M.; Maguire, M.E.; Rensing, C. The metal permease ZupT from Escherichia coli is a transporter with a broad substrate spectrum. J. Bacteriol. 2005, 187, 1604–1611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolaj-Robin, O.; Russell, D.; Hayes, K.A.; Pembroke, J.T.; Soulimane, T. Cation diffusion facilitator family: Structure and function. FEBS Lett. 2015, 589, 1283–1295. [Google Scholar] [CrossRef]
- Nies, D.H. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef]
- Nairn, B.L.; Lonergan, Z.R.; Wang, J.; Braymer, J.J.; Zhang, Y.; Calcutt, M.W.; Lisher, J.P.; Gilston, B.A.; Chazin, W.J.; de Crecy-Lagard, V.; et al. The response of Acinetobacter baumannii to zinc starvation. Cell Host Microbe. 2016, 19, 826–836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montanini, B.; Blaudez, D.; Jeandroz, S.; Sanders, D.; Chalot, M. Phylogenetic and functional analysis of the Cation Diffusion Facilitator (CDF) family: Improved sigNature and prediction of substrate specificity. BMC Genomics 2007, 8, 107. [Google Scholar] [CrossRef] [Green Version]
- Udagedara, S.R.; La Porta, D.M.; Spehar, C.; Purohit, G.; Hein, M.J.A.; Fatmous, M.E.; Casas Garcia, G.P.; Ganio, K.; McDevitt, C.A.; Maher, M.J. Structural and functional characterizations of the C-terminal domains of CzcD proteins. J. Inorg. Biochem. 2020, 208, 111087. [Google Scholar] [CrossRef] [PubMed]
- Bublitz, M.; Morth, J.P.; Nissen, P. P-type ATPases at a glance. J. Cell Sci. 2011, 124, 2515–2519. [Google Scholar] [CrossRef] [Green Version]
- Klein, J.S.; Lewinson, O. Bacterial ATP-driven transporters of transition metals: Physiological roles, mechanisms of action, and roles in bacterial Virulence. Metallomics 2011, 3, 1098–1108. [Google Scholar] [CrossRef]
- Wang, K.; Sitsel, O.; Meloni, G.; Autzen, H.E.; Andersson, M.; Klymchuk, T.; Nielsen, A.M.; Rees, D.C.; Nissen, P.; Gourdon, P. Structure and mechanism of Zn2+-transporting P-type ATPases. Nature 2014, 514, 518–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimunda, D.; Subramanian, P.; Stemmler, T.; Arguello, J.M. A tetrahedral coordination of Zinc during transmembrane transport by P-type Zn(2+)-ATPases. Biochim. Biophys. Acta 2012, 1818, 1374–1377. [Google Scholar] [CrossRef] [Green Version]
- Okkeri, J.; Haltia, T. The metal-binding sites of the zinc-transporting P-type ATPase of Escherichia coli. Lys693 and Asp714 in the seventh and eighth transmembrane segments of ZntA contribute to the coupling of metal binding and ATPase activity. Biochim. Et Biophys. Acta (Bba) Gen. Subj. 2006, 1757, 1485–1495. [Google Scholar] [CrossRef] [Green Version]
- Argüello, J.M.; González-Guerrero, M.; Raimunda, D. Bacterial Transition Metal P1B-ATPases: Transport Mechanism and Roles in Virulence. Biochemistry 2011, 50, 9940–9949. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.T.; Smith, K.P.; Rosenzweig, A.C. Diversity of the metal-transporting P1B-type ATPases. JBIC J. Biol. Inorg. Chem. 2014, 19, 947–960. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, M.; Hayashi, H.; Ohta, T. Chromosome-determined zinc-responsible operon czr in Staphylococcus aureus strain. Microbiol. Immunol. 1999, 43, 115–125. [Google Scholar] [CrossRef]
- Grass, G.; Fan, B.; Rosen, B.P.; Franke, S.; Nies, D.H.; Rensing, C. ZitB (YbgR), a member of the cation diffusion facilitator family, is an additional zinc transporter in Escherichia coli. J. Bacteriol. 2001, 183, 4664–4667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, Y.; Fu, D. Kinetic Study of the Antiport Mechanism of an Escherichia coli Zinc Transporter, ZitB. J. Biol. Chem. 2004, 279, 12043–12050. [Google Scholar] [CrossRef] [Green Version]
- Anton, A.; Weltrowski, A.; Haney, C.J.; Franke, S.; Grass, G.; Rensing, C.; Nies, D.H. Characteristics of zinc transport by two bacterial cation diffusion facilitators from Ralstonia metallidurans CH34 and Escherichia coli. J. Bacteriol. 2004, 186, 7499–7507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coudray, N.; Valvo, S.; Hu, M.; Lasala, R.; Kim, C.; Vink, M.; Zhou, M.; Provasi, D.; Filizola, M.; Tao, J.; et al. Inward-facing conformation of the zinc transporter YiiP revealed by cryoelectron microscopy. Proc. Natl. Acad. Sci. USA 2013, 110, 2140–2145. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Grass, G.; Haney, C.J.; Fan, B.; Rosen, B.P.; Anton, A.; Nies, D.H.; Rensing, C. Functional analysis of the Escherichia coli zinc transporter ZitB. FEMS Microbiol. Lett. 2002, 215, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Guffanti, A.A.; Wei, Y.; Rood, S.V.; Krulwich, T.A. An antiport mechanism for a member of the cation diffusion facilitator family: Divalent cations efflux in exchange for K+ and H+. Mol. Microbiol. 2002, 45, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Chai, J.; Fu, D. Structural basis for autoregulation of the zinc transporter YiiP. Nat. Struct. Mol. Biol. 2009, 16, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Fu, D. Structure of the Zinc Transporter YiiP. Science 2007, 317, 1746–1748. [Google Scholar] [CrossRef] [Green Version]
- Cherezov, V.; Hofer, N.; Szebenyi, D.M.; Kolaj, O.; Wall, J.G.; Gillilan, R.; Srinivasan, V.; Jaroniec, C.P.; Caffrey, M. Insights into the mode of action of a putative zinc transporter CzrB in Thermus thermophilus. Structure 2008, 16, 1378–1388. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, T.; Hattori, M.; Tanaka, Y.; Ishitani, R.; Nureki, O. Crystal structure of the cytosolic domain of the cation diffusion facilitator family protein. Proteins 2009, 76, 768–771. [Google Scholar] [CrossRef] [PubMed]
- Zeytuni, N.; Uebe, R.; Maes, M.; Davidov, G.; Baram, M.; Raschdorf, O.; Nadav-Tsubery, M.; Kolusheva, S.; Bitton, R.; Goobes, G.; et al. Cation diffusion facilitators transport initiation and regulation is mediated by cation induced conformational changes of the cytoplasmic domain. PLoS ONE 2014, 9, e92141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, D.; Soulimane, T. Evidence for zinc and cadmium binding in a CDF transporter lacking the cytoplasmic domain. FEBS Lett. 2012, 586, 4332–4338. [Google Scholar] [CrossRef]
- Prince, R.W.; Cox, C.D.; Vasil, M.L. Coordinate regulation of siderophore and exotoxin A production: Molecular cloning and sequencing of the Pseudomonas aeruginosa fur gene. J. Bacteriol. 1993, 175, 2589–2598. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.-H.; Nies, D.H.; McEvoy, M.M.; Rensing, C. Switch or Funnel: How RND-Type Transport Systems Control Periplasmic Metal Homeostasis. J. Bacteriol. 2011, 193, 2381–2387. [Google Scholar] [CrossRef] [Green Version]
- Alquethamy, S.F.; Adams, F.G.; Naidu, V.; Khorvash, M.; Pederick, V.G.; Zang, M.; Paton, J.C.; Paulsen, I.T.; Hassan, K.A.; Cain, A.K.; et al. The Role of zinc efflux during Acinetobacter baumannii infection. ACS Infect Dis. 2020, 6, 150–158. [Google Scholar] [CrossRef]
- Meni, A.; Yukl, E.T. Structural Features Mediating Zinc Binding and Transfer in the AztABCD Zinc Transporter System. Biomolecules 2020, 10, 1156. [Google Scholar] [CrossRef] [PubMed]
- Dyla, M.; Hansen, S.B.; Nissen, P.; Kjaergaard, M. Structural dynamics of P-type ATPase ion pumps. Biochem. Soc. Trans. 2019, 47, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Bagg, A.; Neilands, J.B. Ferric uptake regulation protein acts as a repressor, employing iron(II) as a cofactor to bind the operator of an iron transport operon in Escherichia coli. Biochemistry 1987, 26, 5471–5477. [Google Scholar] [CrossRef]
- Hantke, K. Selection procedure for deregulated iron transport mutants (fur) in Escherichia coli K 12: Fur not only affects iron metabolism. Mol. Genet. Genom. 1987, 210, 135–139. [Google Scholar] [CrossRef]
- Pinochet-Barros, A.; Helmann, J.D. Redox Sensing by Fe2+in Bacterial Fur Family Metalloregulators. Antioxid. Redox Signal. 2018, 29, 1858–1871. [Google Scholar] [CrossRef]
- Tolmasky, M.E.; Wertheimer, A.M.; Actis, L.A.; Crosa, J.H. Characterization of the Vibrio anguillarum fur gene: Role in regulation of expression of the FatA outer membrane protein and catechols. J. Bacteriol. 1994, 176, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Staggs, T.M.; Perry, R.D. Identification and cloning of a fur regulatory gene in Yersinia pestis. J. Bacteriol. 1991, 173, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Litwin, C.M.; Boyko, S.A.; Calderwood, S.B. Cloning, sequencing, and transcriptional regulation of the Vibrio cholerae fur gene. J. Bacteriol. 1992, 174, 1897–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniel, C.; Haentjens, S.; Bissinger, M.C.; Courcol, R.J. Characterization of the Acinetobacter baumannii Fur regulator: Cloning and sequencing of the fur homolog gene. FEMS Microbiol. Lett. 1999, 170, 199–209. [Google Scholar] [CrossRef]
- Hantke, K. Iron and metal regulation in bacteria. Curr. Opin. Microbiol. 2001, 4, 172–177. [Google Scholar] [CrossRef]
- Coy, M.; Neilands, J.B. Structural dynamics and functional domains of the Fur protein. Biochemistry 1991, 30, 8201–8210. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.A.; Marletta, M.A. Metal Binding Characteristics and Role of Iron Oxidation in the Ferric Uptake Regulator fromEscherichia coli. Biochemistry 2005, 44, 13553–13559. [Google Scholar] [CrossRef]
- Waldbeser, L.S.; Tolmasky, M.E.; Actis, L.A.; Crosa, J.H. Mechanisms for negative regulation by iron of the fatA outer membrane protein gene expression in Vibrio anguillarum. J. Biol. Chem. 1993, 268, 10433–10439. [Google Scholar] [CrossRef]
- Salinas, P.C.; Tolmasky, M.E.; Crosa, J.H. Regulation of the iron uptake system in Vibrio anguillarum: Evidence for a cooperative effect between two transcriptional activators. Proc. Natl. Acad. Sci. USA 1989, 86, 3529–3533. [Google Scholar] [CrossRef] [Green Version]
- Troxell, B.; Hassan, H.M. Transcriptional regulation by Ferric Uptake Regulator (Fur) in pathogenic bacteria. Front. Cell Infect. Microbiol. 2013, 3, 59. [Google Scholar] [CrossRef] [Green Version]
- Fillat, M.F. The FUR (ferric uptake regulator) superfamily: Diversity and versatility of key transcriptional regulators. Arch. Biochem. Biophys. 2014, 546, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.L.; Tang, D.J.; Liao, Q.; Li, H.C.; Chen, Q.; He, Y.Q.; Feng, J.X.; Jiang, B.L.; Lu, G.T.; Chen, B.; et al. The Zur of Xanthomonas campestris functions as a repressor and an activator of putative zinc homeostasis genes via recognizing two distinct sequences within its target promoters. Nucleic Acids Res. 2008, 36, 4295–4309. [Google Scholar] [CrossRef] [Green Version]
- Gilston, B.A.; Wang, S.; Marcus, M.D.; Canalizo-Hernandez, M.A.; Swindell, E.P.; Xue, Y.; Mondragon, A.; O’Halloran, T.V. Structural and mechanistic basis of zinc regulation across the E. coli Zur regulon. PLoS Biol. 2014, 12, e1001987. [Google Scholar] [CrossRef]
- Lucarelli, D.; Russo, S.; Garman, E.; Milano, A.; Meyer-Klaucke, W.; Pohl, E. Crystal structure and function of the zinc uptake regulator FurB from Mycobacterium tuberculosis. J. Biol. Chem. 2007, 282, 9914–9922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.H.; Jung, H.J.; An, Y.J.; Cho, Y.B.; Cha, S.S.; Roe, J.H. Graded expression of zinc-responsive genes through two regulatory zinc-binding sites in Zur. Proc. Natl. Acad. Sci. USA 2011, 108, 5045–5050. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, B.L.; Rathi, S.; Chazin, W.J.; Skaar, E.P. Acinetobacter baumannii response to host-mediated zinc limitation requires the transcriptional regulator Zur. J. Bacteriol. 2014, 196, 2616–2626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, J.A.; Foster, S.J. Zur: A Zn(2+)-responsive regulatory element of Staphylococcus aureus. Microbiology (Reading) 2001, 147, 1259–1266. [Google Scholar] [CrossRef] [Green Version]
- Sanson, M.; Makthal, N.; Flores, A.R.; Olsen, R.J.; Musser, J.M.; Kumaraswami, M. Adhesin competence repressor (AdcR) from Streptococcus pyogenes controls adaptive responses to zinc limitation and contributes to Virulence. Nucleic Acids Res. 2015, 43, 418–432. [Google Scholar] [CrossRef]
- Makthal, N.; Do, H.; Wendel, B.M.; Olsen, R.J.; Helmann, J.D.; Musser, J.M.; Kumaraswami, M. Group A Streptococcus AdcR regulon participates in bacterial defense against host-mediated zinc sequestration and contributes to Virulence. Infect. Immun. 2020, 88, e00097–e20. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Caballero, H.; Guerra, A.J.; Jacobsen, F.E.; Kazmierczak, K.M.; Cowart, D.; Koppolu, U.M.; Scott, R.A.; Winkler, M.E.; Giedroc, D.P. The metalloregulatory zinc site in Streptococcus pneumoniae AdcR, a zinc-activated MarR family repressor. J. Mol. Biol. 2010, 403, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Toewiwat, N.; Whangsuk, P.; Ploypradith, W.; Mongkolsuk, S.; Loprasert, S. Cefoperazone induces esterase B expression by EstR and esterase B enhances cefoperazone activity at the periplasm. Int. J. Med. Microbiol. 2020, 310, 151396. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Li, M.; Liu, L.; Zhang, J.; Zhang, Y.; Guo, J.; Zhao, T.; Cao, S.; Li, Z.; Zhang, H. Functional analysis of Brucella reveals transcriptional regulation of MarR. Microb. Pathog. 2020, 144, 104201. [Google Scholar] [CrossRef]
- Guerra, A.J.; Dann, C.E., 3rd; Giedroc, D.P. Crystal structure of the zinc-dependent MarR family transcriptional regulator AdcR in the Zn(II)-bound state. J. Am. Chem. Soc. 2011, 133, 19614–19617. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.E.; Edmonds, K.A.; Bruce, K.E.; Campanello, G.C.; Eijkelkamp, B.A.; Brazel, E.B.; McDevitt, C.A.; Winkler, M.E.; Giedroc, D.P. The zinc efflux activator SczA protects Streptococcus pneumoniae serotype 2 D39 from intracellular zinc toxicity. Mol. Microbiol. 2017, 104, 636–651. [Google Scholar] [CrossRef] [Green Version]
- Kloosterman, T.G.; van der Kooi-Pol, M.M.; Bijlsma, J.J.; Kuipers, O.P. The novel transcriptional regulator SczA mediates protection against Zn2+ stress by activation of the Zn2+-resistance gene czcD in Streptococcus pneumoniae. Mol. Microbiol. 2007, 65, 1049–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canneva, F.; Branzoni, M.; Riccardi, G.; Provvedi, R.; Milano, A. Rv2358 and FurB: Two transcriptional regulators from Mycobacterium tuberculosis which respond to zinc. J. Bacteriol. 2005, 187, 5837–5840. [Google Scholar] [CrossRef] [Green Version]
- Eckelt, E.; Jarek, M.; Fromke, C.; Meens, J.; Goethe, R. Identification of a lineage specific zinc responsive genomic island in Mycobacterium avium ssp. paratuberculosis. BMC Genom. 2014, 15, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milano, A.; Branzoni, M.; Canneva, F.; Profumo, A.; Riccardi, G. The Mycobacterium tuberculosis Rv2358-furB operon is induced by zinc. Res. Microbiol. 2004, 155, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, M.S.; Nikolaidis, N.; Tolmasky, M.E. Rise and dissemination of aminoglycoside resistance: The aac(6′)-Ib paradigm. Front. Microbiol. 2013, 4, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, F.; Yan, X.; Shakya, T.; Baettig, O.M.; Ait-Mohand-Brunet, S.; Berghuis, A.M.; Wright, G.D.; Auclair, K. Synthesis and structure-activity relationships of truncated bisubstrate inhibitors of aminoglycoside 6’-N-acetyltransferases. J. Med. Chem. 2006, 49, 5273–5281. [Google Scholar] [CrossRef]
- Vong, K.; Auclair, K. Understanding and overcoming aminoglycoside resistance caused by N-6′-acetyltransferase. MedChemComm 2012, 3, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Goethe, E.; Laarmann, K.; Luhrs, J.; Jarek, M.; Meens, J.; Lewin, A.; Goethe, R. Critical role of Zur and SmtB in zinc homeostasis of Mycobacterium smegmatis. MSystems 2020, 5, e00880–e19. [Google Scholar] [CrossRef] [PubMed]
- Houghton, J.L.; Green, K.D.; Chen, W.; Garneau-Tsodikova, S. The future of aminoglycosides: The end or renaissance? ChemBiochem 2010, 11, 880–902. [Google Scholar] [CrossRef] [Green Version]
- Block, M.; Blanchard, D.L. Aminoglycosides; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Ramirez, M.S.; Tolmasky, M.E. Amikacin: Uses, Resistance, and Prospects for Inhibition. Molecules 2017, 22, 2267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, M.S.; Tolmasky, M.E. Aminoglycoside modifying enzymes. Drug Resist. Updat. 2010, 13, 151–171. [Google Scholar] [CrossRef] [Green Version]
- Tolmasky, M.E. Strategies to prolong the useful life of existing antibiotics and help overcoming the antibiotic resistance crisis. In Frontiers in Clinical Drug Research-Anti Infectives; Atta-ur-Rhaman, Ed.; Bentham Books: Sharjah, United Arab Emirates, 2017; pp. 1–27. [Google Scholar]
- Papp-Wallace, K.M.; Bonomo, R.A. New beta-lactamase inhibitors in the clinic. Infect. Dis. Clin. North Am. 2016, 30, 441–464. [Google Scholar] [CrossRef]
- Daigle, D.M.; McKay, G.A.; Thompson, P.R.; Wright, G.D. Aminoglycoside antibiotic phosphotransferases are also serine protein kinases. Chem. Biol. 1999, 6, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Chiem, K.; Fuentes, B.A.; Lin, D.L.; Tran, T.; Jackson, A.; Ramirez, M.S.; Tolmasky, M.E. Inhibition of aminoglycoside 6’-N-acetyltransferase type Ib-mediated amikacin resistance in Klebsiella pneumoniae by zinc and copper pyrithione. Antimicrob. Agents Chemother. 2015, 59, 5851–5853. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.L.; Tran, T.; Adams, C.; Alam, J.Y.; Herron, S.R.; Tolmasky, M.E. Inhibitors of the aminoglycoside 6’-N-acetyltransferase type Ib [AAC(6’)-Ib] identified by in silico molecular docking. Bioorg. Med. Chem. Lett. 2013, 23, 5694–5698. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.; Chiem, K.; Jani, S.; Arivett, B.A.; Lin, D.L.; Lad, R.; Jimenez, V.; Farone, M.B.; Debevec, G.; Santos, R.; et al. Tolmasky. Identification of a small molecule inhibitor of the aminoglycoside 6’-N-acetyltransferase type Ib [AAC(6’)-Ib] using mixture-based combinatorial libraries. Int. J. Antimicrob. Agents 2018, 51, 752–761. [Google Scholar] [CrossRef]
- Li, Y.; Green, K.D.; Johnson, B.R.; Garneau-Tsodikova, S. Inhibition of aminoglycoside acetyltransferase resistance enzymes by metal salts. Antimicrob. Agents Chemother. 2015, 59, 4148–4156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiem, K.; Hue, F.; Magallon, J.; Tolmasky, M.E. Inhibition of aminoglycoside 6’-N-acetyltransferase type Ib-mediated amikacin resistance by zinc complexed with clioquinol, an ionophore active against tumors and neurodegenerative diseases. Int. J. Antimicrob. Agents 2018, 51, 271–273. [Google Scholar] [CrossRef]
- Ahmed, S.; Sony, S.A.; Chowdhury, M.B.; Ullah, M.M.; Paul, S.; Hossain, T. Retention of antibiotic activity against resistant bacteria harbouring aminoglycoside-N-acetyltransferase enzyme by adjuvants: A combination of in-silico and in-vitro study. Sci. Rep. 2020, 10, 19381. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Sripada, L.; Singh, R. Side effects of antibiotics during bacterial infection: Mitochondria, the main target in host cell. Mitochondrion 2014, 16, 50–54. [Google Scholar] [CrossRef]
- Sensi, S.L.; Ton-That, D.; Sullivan, P.G.; Jonas, E.A.; Gee, K.R.; Kaczmarek, L.K.; Weiss, J.H. Modulation of mitochondrial function by endogenous Zn2+ pools. Proc. Natl. Acad. Sci. USA 2003, 100, 6157–6162. [Google Scholar] [CrossRef] [Green Version]
- Sensi, S.L.; Yin, H.Z.; Carriedo, S.G.; Rao, S.S.; Weiss, J.H. Preferential Zn2+ influx through Ca2+-permeable AMPA/kainate channels triggers prolonged mitochondrial superoxide production. Proc. Natl. Acad. Sci. USA 1999, 96, 2414–2419. [Google Scholar] [CrossRef] [Green Version]
- Frederickson, C.J. Neurobiology of Zinc and Zinc-Containing Neurons. Int. Rev. Neurobiol. 1989, 31, 145–238. [Google Scholar] [CrossRef] [PubMed]
- Bettger, W.J.; O’Dell, B.L. A critical physiological role of zinc in the structure and function of biomembranes. Life Sci. 1981, 28, 1425–1438. [Google Scholar] [CrossRef]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [CrossRef]
- Brand, I.A.; Kleineke, J. Intracellular Zinc Movement and Its Effect on the Carbohydrate Metabolism of Isolated Rat Hepatocytes. J. Biol. Chem. 1996, 271, 1941–1949. [Google Scholar] [CrossRef] [Green Version]
- Simons, T.J.B. Intracellular free zinc and zinc buffering in human red blood cells. J. Membr. Biol. 1991, 123, 63–71. [Google Scholar] [CrossRef]
- Simons, T.J. Measurement of free Zn2+ ion concentration with the fluorescent probe mag-fura-2 (furaptra). J. Biochem. Biophys. Methods 1993, 27, 25–37. [Google Scholar] [CrossRef]
- Zalewski, P.D.; Forbes, I.J.; Betts, W.H. Correlation of apoptosis with change in intracellular labile Zn(II) using zinquin [(2-methyl-8-p-toluenesulphonamido-6-quinolyloxy)acetic acid], a new specific fluorescent probe for Zn(II). Biochem. J. 1993, 296, 403–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, R.J.P.A. Introduction to the Biochemistry of Zinc; Springer International Publishing: Berlin, Germany, 1989; pp. 15–31. [Google Scholar]
- Palmiter, R.D.; Cole, T.B.; Findley, S.D. ZnT-2, a mammalian protein that confers resistance to zinc by facilitating vesicular sequestration. EMBO J. 1996, 15, 1784–1791. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Yokoyama, M.; Koh, J. Zinc neurotoxicity in cortical cell culture. Neuroscience 1988, 24, 67–79. [Google Scholar] [CrossRef]
- Provinciali, M.; Di Stefano, G.; Fabris, N. Dose-dependent opposite effect of zinc on apoptosis in mouse thymocytes. Int. J. Immunopharmacol. 1995, 17, 735–744. [Google Scholar] [CrossRef]
- Telford, W.G.; Fraker, P.J. Preferential induction of apoptosis in mouse CD4+CD8+ alpha beta TCRloCD3 epsilon lo thymocytes by zinc. J. Cell Physiol. 1995, 164, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, M.; Koh, J.; Choi, D. Brief exposure to zinc is toxic to cortical neurons. Neurosci. Lett. 1986, 71, 351–355. [Google Scholar] [CrossRef]
- Palmiter, R. Constitutive Expression of Metallothionein-III (MT-III), but Not MT-I, Inhibits Growth When Cells Become Zinc Deficient. Toxicol. Appl. Pharm. 1995, 135, 139–146. [Google Scholar] [CrossRef]
- Palmiter, R.D. Protection against zinc toxicity by metallothionein and zinc transporter. Proc. Natl. Acad. Sci. USA 2004, 101, 4918–4923. [Google Scholar] [CrossRef] [Green Version]
- Palmiter, R.D.; Cole, T.B.; Quaife, C.J.; Findley, S.D. ZnT-3, a putative transporter of zinc into synaptic vesicles. Proc. Natl. Acad. Sci. USA 1996, 93, 14934–14939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmiter, R.; Findley, S. Cloning and functional characterization of a mammalian zinc transporter that confers resistance to zinc. Embo J. 1995, 14, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Cuajungco, M.P.; Lees, G.J. Zinc Metabolism in the Brain: Relevance to Human Neurodegenerative Disorders. Neurobiol. Dis. 1997, 4, 137–169. [Google Scholar] [CrossRef] [Green Version]
- Maret, W. Zinc in Cellular Regulation: The Nature and Significance of “Zinc Signals”. Int. J. Mol. Sci 2017, 18, 2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, R.J.; Cousins, R.J. Mammalian Zinc Transporters. J. Nutr. 1998, 128, 667–670. [Google Scholar] [CrossRef] [Green Version]
- Howell, G.A.; Welch, M.G.; Frederickson, C.J. Stimulation-induced uptake and release of zinc in hippocampal slices. Nat. Cell Biol. 1984, 308, 736–738. [Google Scholar] [CrossRef]
- Kambe, T.; Tsuji, T.; Hashimoto, A.; Itsumura, N. The Physiological, Biochemical, and Molecular Roles of Zinc Transporters in Zinc Homeostasis and Metabolism. Physiol. Rev. 2015, 95, 749–784. [Google Scholar] [CrossRef]
- Aniksztejn, L.; Charton, G.; Ben-Ari, Y. Selective release of endogenous zinc from the hippocampal mossy fibers in situ. Brain Res. 1987, 404, 58–64. [Google Scholar] [CrossRef]
- Assaf, S.Y.; Chung, S.-H. Release of endogenous Zn2+ from brain tissue during activity. Nat. Cell Biol. 1984, 308, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Charton, G.; Rovira, C.; Ben-Ari, Y.; Leviel, V. Spontaneous and evoked release of endogenous Zn2+ in the hippocampal mossy fiber zone of the rat in situ. Exp. Brain Res. 1985, 58, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Bin, B.H.; Fukada, T.; Hosaka, T.; Yamasaki, S.; Ohashi, W.; Hojyo, S.; Miyai, T.; Nishida, K.; Yokoyama, S.; Hirano, T. Biochemical characterization of human ZIP13 protein: A homo-dimerized zinc transporter involved in the spondylocheiro dysplastic Ehlers-Danlos syndrome. J. Biol. Chem. 2011, 286, 40255–40265. [Google Scholar] [CrossRef] [Green Version]
- Kukic, I.; Lee, J.K.; Coblentz, J.; Kelleher, S.L.; Kiselyov, K. Zinc-dependent lysosomal enlargement in TRPML1-deficient cells involves MTF-1 transcription factor and ZnT4 (Slc30a4) transporter. Biochem. J. 2013, 451, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Raffaniello, R.D.; Wapnir, R.A. Zinc-induced metallothionein synthesis by Caco-2 cells. Biochem. Med. Metab. Biol. 1991, 45, 101–107. [Google Scholar] [CrossRef]
- Thakran, P.; Leuschen, M.P.; Ebadi, M. Metallothionein induction in rat hippocampal neurons in primary culture. Vivo 1989, 3, 191–197. [Google Scholar]
- Valentine, R.A.; Jackson, K.A.; Christie, G.R.; Mathers, J.C.; Taylor, P.M.; Ford, D. ZnT5 variant B is a bidirectional zinc transporter and mediates zinc uptake in human intestinal Caco-2 cells. J. Biol. Chem. 2007, 282, 14389–14393. [Google Scholar] [CrossRef] [Green Version]
- Cuajungco, M.P.; Lees, G.J. Nitric oxide generators produce accumulation of chelatable zinc in hippocampal neuronal perikarya. Brain Res. 1998, 799, 118–129. [Google Scholar] [CrossRef]
- Jacob, C.; Maret, W.; Vallee, B.L. Control of zinc transfer between thionein, metallothionein, and zinc proteins. Proc. Natl. Acad. Sci. USA 1998, 95, 3489–3494. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.-J.; Maret, W.; Vallee, B.L. The glutathione redox couple modulates zinc transfer from metallothionein to zinc-depleted sorbitol dehydrogenase. Proc. Natl. Acad. Sci. USA 1998, 95, 3483–3488. [Google Scholar] [CrossRef] [Green Version]
- Wensink, J.; Molenaar, A.J.; Woroniecka, U.D.; Van den Hamer, C.J. Zinc uptake into synaptosomes. J. Neurochem. 1988, 50, 782–789. [Google Scholar] [CrossRef]
- Colvin, R.A.; Holmes, W.R.; Fontaine, C.P.; Maret, W. Cytosolic zinc buffering and muffling: Their role in intracellular zinc homeostasis. Metallomics 2010, 2, 306–317. [Google Scholar] [CrossRef]
- Cousins, R.J.; Liuzzi, J.P.; Lichten, L.A. Mammalian Zinc Transport, Trafficking, and Signals. J. Biol. Chem. 2006, 281, 24085–24089. [Google Scholar] [CrossRef] [Green Version]
- Eide, D.J. Zinc transporters and the cellular Trafficking of zinc. Biochim. Et Biophys. Acta (Bba) Bioenerg. 2006, 1763, 711–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuajungco, M.P.; Basilio, L.C.; Silva, J.; Hart, T.; Tringali, J.; Chen, C.C.; Biel, M.; Grimm, C. Cellular Zinc Levels Are Modulated by TRPML1-TMEM163 Interaction. Traffic 2014, 15, 1247–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waberer, L.; Henrich, E.; Peetz, O.; Morgner, N.; Dotsch, V.; Bernhard, F.; Volknandt, W. The synaptic vesicle protein SV31 assembles into a dimer and transports Zn(2). J. Neurochem. 2017, 140, 280–293. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, V.B.; Ali, S.; Escobar, A.; Cuajungco, M.P. Transmembrane 163 (TMEM163) protein effluxes zinc. Arch. Biochem. Biophys. 2019, 677, 108166. [Google Scholar] [CrossRef]
- Ohana, E.; Hoch, E.; Keasar, C.; Kambe, T.; Yifrach, O.; Hershfinkel, M.; Sekler, I. Identification of the Zn2+ binding site and mode of operation of a mammalian Zn2+ transporter. J. Biol. Chem. 2009, 284, 17677–17686. [Google Scholar] [CrossRef] [Green Version]
- Maret, W. Metallothionein redox biology in the cytoprotective and cytotoxic functions of zinc. Exp. Gerontol. 2008, 43, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Krezoski, S.K.; Villalobos, J.; Shaw, C.F., 3rd; Petering, D.H. Kinetic lability of zinc bound to metallothionein in Ehrlich cells. Biochem. J. 1988, 255, 483–491. [Google Scholar]
- Beaulieu, C.; Dyck, R.; Cynader, M. Enrichment of glutamate in zinc-containing terminals of the cat visual cortex. NeuroReport 1992, 3, 861–864. [Google Scholar] [CrossRef]
- Frederickson, C.J.; Klitenick, M.A.; Manton, W.I.; Kirkpatrick, J.B. Cytoarchitectonic distribution of zinc in the hippocampus of man and the rat. Brain Res. 1983, 273, 335–339. [Google Scholar] [CrossRef]
- Haug, F.M. Electron microscopical localization of the zinc in hippocampal mossy fibre synapses by a modified sulfide silver procedure. Histochem. Cell Biol. 1967, 8, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Pe´rez-Clausell, J.; Danscher, G. Intravesicular localization of zinc in rat telencephalic boutons. A histochemical study. Brain Res. 1985, 337, 91–98. [Google Scholar] [CrossRef]
- Bresink, I.; Ebert, B.; Parsons, C.G.; Mutschler, E. Zinc changes AMPA receptor properties: Results of binding studies and patch clamp recordings. Neuropharmacology 1996, 35, 503–509. [Google Scholar] [CrossRef]
- Christine, C.; Choi, D. Effect of zinc on NMDA receptor-mediated channel currents in cortical neurons. J. Neurosci. 1990, 10, 108–116. [Google Scholar] [CrossRef] [Green Version]
- Dreixler, J.C.; Leonard, J.P. Subunit-specific enhancement of glutamate receptor responses by zinc. Mol. Brain Res. 1994, 22, 144–150. [Google Scholar] [CrossRef]
- Xie, X.; Gerber, U.; Gahwiler, B.H.; Smart, T.G. Interaction of zinc with ionotropic and metabotropic glutamate receptors in rat hippocampal slices. Neurosci. Lett. 1993, 159, 46–50. [Google Scholar] [CrossRef]
- Yin, H.Z.; Weiss, J.H. Zn2+ permeates Ca2+permeable AMPA/kainate channels and triggers selective neural injury. NeuroReport 1995, 6, 2553–2556. [Google Scholar] [CrossRef]
- Legendre, P.; Westbrook, G.L. Noncompetitive inhibition of gamma-aminobutyric acidA channels by Zn. Mol. Pharm. 1991, 39, 267–274. [Google Scholar]
- Smart, T.G.; Moss, S.J.; Xie, X.; Huganir, R.L. GABAA receptors are differentially sensitive to zinc: Dependence on subunit composition. Br. J. Pharmacol. 1991, 103, 1837–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, M.L.; Vyklicky, L.; Westbrook, G.L. Modulation of excitatory amino acid receptors by group IIB metal cations in cultured mouse hippocampal neurones. J. Physiol. 1989, 415, 329–350. [Google Scholar] [CrossRef]
- Mayer, M.L.; Vyklicky, L., Jr. The action of zinc on synaptic transmission and neuronal excitability in cultures of mouse hippocampus. J. Physiol. 1989, 415, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Cloues, R. Properties of ATP-gated channels recorded from rat sympathetic neurons: Voltage dependence and regulation by Zn2+ ions. J. Neurophysiol. 1995, 73, 312–319. [Google Scholar] [CrossRef]
- A Connor, M.; Chavkin, C. Ionic zinc may function as an endogenous ligand for the haloperidol-sensitive sigma 2 receptor in rat brain. Mol. Pharm. 1992, 42, 471–479. [Google Scholar]
- Kumamoto, E.; Murata, Y. Glycine current in rat septal cholinergic neuron in culture: Monophasic positive modulation by Zn2+. J. Neurophysiol. 1996, 76, 227–241. [Google Scholar] [CrossRef]
- Fabris, N.; Mocchegiani, E. Zinc, human diseases and aging. Aging Clin. Exp. Res. 1995, 7, 77–93. [Google Scholar] [CrossRef]
- Hexum, T.D. Studies on the reaction catalyzed by transport (Na, K) adenosine triphosphatase—I. Biochem. Pharm. 1974, 23, 3441–3447. [Google Scholar] [CrossRef]
- Hogstrand, C.; Verbost, P.M.; Wendelaar Bonga, S.E. Inhibition of human erythrocyte Ca2+-ATPase by Zn2+. Toxicology 1999, 133, 139–145. [Google Scholar] [CrossRef]
- Link, T.A.; von Jagow, G. Zinc ions inhibit the QP center of bovine heart mitochondrial bc1 complex by blocking a protonatable group. J. Biol. Chem. 1995, 270, 25001–25006. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.; Hogstrand, C.; Maret, W. Picomolar concentrations of free zinc(II) ions regulate receptor protein-tyrosine phosphatase beta activity. J. Biol. Chem. 2012, 287, 9322–9326. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, S.R.; Bishop, W.R.; Kirschmeier, P.; George, S.J.; Cramer, S.P.; Hendrickson, W.A. Identification and characterization of zinc binding sites in protein kinase C. Science 1991, 254, 1776–1779. [Google Scholar] [CrossRef]
- Perry, D.K.; Smyth, M.J.; Stennicke, H.R.; Salvesen, G.S.; Duriez, P.; Poirier, G.G.; Hannun, Y.A. Zinc is a potent inhibitor of the apoptotic protease, caspase-A novel target for zinc in the inhibition of apoptosis. J. Biol. Chem. 1997, 272, 18530–18533. [Google Scholar] [CrossRef] [Green Version]
- Velázquez-Delgado, E.M.; Hardy, J.A. Zinc-mediated Allosteric Inhibition of Caspase. J. Biol. Chem. 2012, 287, 36000–36011. [Google Scholar] [CrossRef] [Green Version]
- Posewitz, M.C.; Wilcox, D.E. Properties of the Sp1 Zinc Finger 3 Peptide: Coordination Chemistry, Redox Reactions, and Metal Binding Competition with Metallothionein. Chem. Res. Toxicol. 1995, 8, 1020–1028. [Google Scholar] [CrossRef]
- Piątek, K.; Hartwig, A.; Bal, W. Physiological levels of glutathione enhance Zn(II) binding by a Cys4 zinc finger. Biochem. Biophys. Res. Commun. 2009, 389, 265–268. [Google Scholar] [CrossRef] [PubMed]
- Maret, W. Oxidative metal release from metallothionein via zinc-thiol/disulfide interchange. Proc. Natl. Acad. Sci. USA 1994, 91, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Baudier, J.; Haglid, K.; Haiech, J.; Gerard, D. Zinc ion binding to human brain calcium binding proteins, calmodulin and S100b protein. Biochem. Biophys. Res. Commun. 1983, 114, 1138–1146. [Google Scholar] [CrossRef]
- Eagle, G.R.; Zombola, R.R.; Himes, R.H. Tubulin-zinc interactions: Binding and polymerization studies. Biochemistry 1983, 22, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Cuajungco, M.P.; Lees, G.J. Zinc and Alzheimer’s disease: Is there a direct link? Brain Res. 1997, 23, 219–236. [Google Scholar] [CrossRef]
- Donaldson, J.; St-Pierre, T.; Minnich, J.; Barbeau, A. Seizures in Rats Associated with Divalent Cation Inhibition of Na+–K+-ATP’ase. Can. J. Biochem. 1971, 49, 1217–1224. [Google Scholar] [CrossRef]
- Zhang, G.H.; Yamaguchi, M.; Kimura, S.; Higham, S.; Kraus-Friedmann, N. Effects of heavy metal on rat liver microsomal Ca2(+)-ATPase and Ca2+ sequestering. Relation to SH groups. J. Biol. Chem. 1990, 265, 2184–2189. [Google Scholar] [CrossRef]
- Lees, G.; Leong, W. Brain lesions induced by specific and non-specific inhibitors of sodium-potassium ATPase. Brain Res. 1994, 649, 225–233. [Google Scholar] [CrossRef]
- Lees, G.; Leong, W. The sodium-potassium ATPase inhibitor ouabain is neurotoxic in the rat substantia nigra and striatum. Neurosci. Lett. 1995, 188, 113–116. [Google Scholar] [CrossRef]
- Kleiner, D.; Von Jagow, G. On the inhibition of mitochondrial electron transport by Zn2+ ions. FEBS Lett. 1972, 20, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Skulachev, V.P.; Chistyakov, V.V.; Jasaitis, A.A.; Smirnova, E.G. Inhibition of the respiratory chain by zinc ions. Biochem. Biophys. Res. Commun. 1967, 26, 1–6. [Google Scholar] [CrossRef]
- Gabrielsson, B.; Robson, T.; Norris, D.; Chung, S.H. Effects of divalent metal ions on the uptake of glutamate and GABA from synaptosomal fractions. Brain Res. 1986, 384, 218–223. [Google Scholar] [CrossRef]
- Velasco, I.; Tapia, R.; Massieu, L. Inhibition of glutamate uptake induces progressive accumulation of extracellular glutamate and neuronal damage in rat cortical cultures. J. Neurosci. Res. 1996, 44, 551–561. [Google Scholar] [CrossRef]
- Shen, Z.; Haragopal, H.; Li, Y.V. Zinc modulates synaptic transmission by differentially regulating synaptic glutamate homeostasis in hippocampus. Eur. J. Neurosci. 2020, 52, 3710–3722. [Google Scholar] [CrossRef]
- Maret, W. Inhibitory zinc sites in enzymes. Biometals 2013, 26, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Heizmann, C.W.; Cox, A.J. New perspectives on S100 proteins: A multi-functional Ca 2+ -, Zn 2+ - and Cu 2+ -binding protein family. Biometals 1998, 11, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Moroz, O.V.; Burkitt, W.; Wittkowski, H.; He, W.; Ianoul, A.; Novitskaya, V.; Xie, J.; Polyakova, O.; Lednev, I.K.; Shekhtman, A.; et al. Both Ca2+ and Zn2+ are essential for S100A12 protein oligomerization and function. BMC Biochem. 2009, 10, 11–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moroz, O.V.; Wilson, K.S.; Bronstein, I.B. The role of zinc in the S100 proteins: Insights from the X-ray structures. Amino Acids 2010, 41, 761–772. [Google Scholar] [CrossRef]
- Nickolson, V.; Veldstra, H. The influence of various cations on the binding of colchicine by rat brain homogenates. Stabilization of intact neurotubules by zinc and cadmium ions. FEBS Lett. 1972, 23, 309–313. [Google Scholar] [CrossRef] [Green Version]
- Kress, Y.; Gaskin, F.; Brosnan, C.F.; Levine, S. Effects of zinc on the cytoskeletal proteins in the central nervous system of the rat. Brain Res. 1981, 220, 139–149. [Google Scholar] [CrossRef]
- Hedberg, K.K.; Birrell, G.B.; Mobley, P.L.; Griffith, O.H. Transition metal chelator TPEN counteracts phorbol ester-induced actin cytoskeletal disruption in C6 rat glioma cells without inhibiting activation or translocation of protein kinase C. J. Cell. Physiol. 1994, 158, 337–346. [Google Scholar] [CrossRef]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer. 1972, 26, 239–257. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Xu, Y.; Nguyen, Q.; Novikoff, P.M.; Czaja, M.J. Induction of hepatoma cell apoptosis by c-myc requires zinc and occurs in the absence of DNA fragmentation. Am. J. Physiol. Content 1996, 270, 60. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.S.; Stockwell, B.R. Ferroptosis: Death by Lipid Peroxidation. Trends Cell Biol. 2016, 26, 165–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalewski, P.; Forbes, I.; Seamark, R.; Borlinghaus, R.; Betts, W.; Lincoln, S.; Ward, A. Flux of intracellular labile zinc during apoptosis (gene-directed cell death) revealed by a specific chemical probe, Zinquin. Chem. Biol. 1994, 1, 153–161. [Google Scholar] [CrossRef]
- Fliss, H.; Ménard, M. Hypochlorous acid-induced mobilization of zinc from metalloproteins. Arch. Biochem. Biophys. 1991, 287, 175–179. [Google Scholar] [CrossRef]
- Fliss, H.; Ménard, M. Oxidant-induced mobilization of zinc from metallothionein. Arch. Biochem. Biophys. 1992, 293, 195–199. [Google Scholar] [CrossRef]
- Fliss, H.; Ménard, M.; Desai, M. Hypochlorous acid mobilizes cellular zinc. Can. J. Physiol. Pharm. 1991, 69, 1686–1691. [Google Scholar] [CrossRef]
- Baba, A.; Kihara, T.; Sawada, T.; Iwata, H. Excitatory amino acids enhance dissociation of zinc from soluble protein in cytosol of rat hippocampus. Brain Res. 1989, 486, 372–375. [Google Scholar] [CrossRef]
- Colletti, G.A.; Miedel, M.T.; Quinn, J.S.; Andharia, N.; Weisz, O.A.; Kiselyov, K. Loss of Lysosomal Ion Channel Transient Receptor Potential Channel Mucolipin-1 (TRPML1) Leads to Cathepsin B-dependent Apoptosis. J. Biol. Chem. 2012, 287, 8082–8091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatachalam, K.; Long, A.A.; Elsaesser, R.; Nikolaeva, D.; Broadie, K.; Montell, C. Motor Deficit in a Drosophila Model of Mucolipidosis Type IV due to Defective Clearance of Apoptotic Cells. Cell 2008, 135, 838–851. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Zhou, Y.; Lind, S.E.; Ding, W.-Q. Clioquinol targets zinc to lysosomes in human cancer cells. Biochem. J. 2008, 417, 133–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chacon, J.; Rosas, L.; Cuajungco, M.P. ZnT3 expression levels are down-regulated in the brain of Mcoln1 knockout mice. Mol. Brain 2019, 12, 24. [Google Scholar] [CrossRef]
- Koh, J.-Y.; Suh, S.W.; Gwag, B.J.; He, Y.Y.; Hsu, C.Y.; Choi, D.W.; Novelli, M.R.; Williamson, J.A.; Tomlinson, I.P.M.; Elia, G.; et al. The Role of Zinc in Selective Neuronal Death After Transient Global Cerebral Ischemia. Science 1996, 272, 1013–1016. [Google Scholar] [CrossRef] [PubMed]
- Frederickson, C.; Hernández, M.; McGinty, J. Translocation of zinc may contribute to seizure-induced death of neurons. Brain Res. 1989, 480, 317–321. [Google Scholar] [CrossRef]
- Charriaut-Marlangue, C.; Aggoun-Zouaoui, D.; Represa, A.; Ben-Ari, Y. Apoptotic features of selective neuronal death in ischemia, epilepsy and gpI20 toxicity. Trends Neurosci. 1996, 19, 109–114. [Google Scholar] [CrossRef]
- Arslan, P.; Di Virgilio, F.; Beltrame, M.; Tsien, R.Y.; Pozzan, T. Cytosolic Ca2+ homeostasis in Ehrlich and Yoshida carcinomas. A new, membrane-permeant chelator of heavy metals reveals that these ascites tumor cell lines have normal cytosolic free Ca2+. J. Biol. Chem. 1985, 260, 2719–2727. [Google Scholar] [CrossRef]
- May, P.M.; Bulman, R.A. 5 The Present Status of Chelating Agents in Medicine. Prog. Med. Chem. 1983, 20, 225–336. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D. Historical Background and Fundamental Concepts; Elsevier BV: Berlin, Germany, 1964; pp. 1–50. [Google Scholar]
- Gibson, W.; Hardy, W.; Groom, M. The effect and mode of action of zinc pyrithione on cell growth. II. in vivo studies. Food Chem. Toxicol. 1985, 23, 103–110. [Google Scholar] [CrossRef]
- Gibson, W.; Chamberlain, M.; Parsons, J.; Brunskill, J.; Leftwich, D.; Lock, S.; Safford, R. The effect and mode of action of zinc pyrithione on cell growth. I. in vitro studies. Food Chem. Toxicol. 1985, 23, 93–102. [Google Scholar] [CrossRef]
- Park, M.-H.; Lee, S.-J.; Byun, H.-R.; Kim, Y.; Oh, Y.J.; Koh, J.-Y.; Hwang, J.J. Clioquinol induces autophagy in cultured astrocytes and neurons by acting as a zinc ionophore. Neurobiol. Dis. 2011, 42, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.; Hall, A. 2 Clinically Useful Chelators of Tripositive Elements. Prog. Med. Chem. 1991, 28, 41–173. [Google Scholar] [CrossRef]
- Cuajungco, M.P.; Lees, G.J. Diverse effects of metal chelating agents on the neuronal cytotoxicity of zinc in the hippocampus. Brain Res. 1998, 799, 97–107. [Google Scholar] [CrossRef]
- Anderegg, G.; Hubmann, E.; Podder, N.G.; Wenk, F. Pyridine derivatives as complexing agents XI: Thermodynamics of metal complex formation with bis-, tris-, and tetrakis[(2-pyridyl)methyl]-amines. Helv. Chim. Acta 1977, 60, 123–140. [Google Scholar] [CrossRef]
- Treves, S.; Trentini, P.L.; Ascanelli, M.; Bucci, G.; Di Virgilio, F. Apoptosis Is Dependent on Intracellular Zinc and Independent of Intracellular Calcium in Lymphocytes. Exp. Cell Res. 1994, 211, 339–343. [Google Scholar] [CrossRef] [PubMed]
- A Lazebnik, Y.; Cole, S.; A Cooke, C.; Nelson, W.G.; Earnshaw, W.C. Nuclear events of apoptosis in vitro in cell-free mitotic extracts: A model system for analysis of the active phase of apoptosis. J. Cell Biol. 1993, 123, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Chow, S.C.; McCabe, M.J.; Orrenius, S. Lack of Ca2+ involvement in thymocyte apoptosis induced by chelation of intracellular Zn2+. Lab. Investig. 1995, 73, 111–117. [Google Scholar] [PubMed]
- Cuajungco, M.P.; Lees, G.J. Prevention of zinc neurotoxicity in vivo by N,N,N’,N’-tetrakis (2-pyridylmethyl) ethylene-diamine (TPEN). Neuroreport 1996, 7, 1301–1304. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Koh, J.Y. ZINC AND BRAIN INJURY. Annu. Rev. Neurosci. 1998, 21, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Weiss, J.H.; Koh, J.-Y.; Christine, C.W.; Kurth, M.C. Glutamate Neurotoxicity, Calcium, and Zinc. Ann. N. Y. Acad. Sci. 1989, 568, 219–224. [Google Scholar] [CrossRef]


{kind=link}
| Protein Name | Binding Affinity * [Reference] |
|---|---|
| Superoxide dismutase (Cu/Zn) | Kd = 5 × 10−5 M [261] |
| Na+/K+ ATPase | Kd = 3 × 10−6 M [262] |
| Ca2+ ATPase | Ki = 8 × 10−12 M [263] |
| Mitochondrial complex III | Kd = 1 × 10−7 M [264] |
| Protein tyrosine phosphatase | Ki = 2 × 10−13 M [265] |
| Protein kinase C | Kd = 1 × 10−13 M [266] |
| Caspase-3 | Ki = 1 × 10−13 M [267] |
| Caspase-6 | Ki = 1 × 10−13 M [268] |
| Sp1 | Kd = 1 × 10−10 M [269] |
| Glutathione | Kd = 5 × 10−7 M [270] |
| Metallothionein | Kd = 1 × 10−12 M [271] |
| Calmodulin | Kd = 8 × 10−5 M [272] |
| NMDA Receptor | Kd = 1 × 10−7 M [256] |
| GABA receptor | Kd = 1 × 10−7 M [257] |
| Tubulin | Kd = 2 × 10−7 M [273] |
| Heme | Kd = 2 × 10−8 M [206] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuajungco, M.P.; Ramirez, M.S.; Tolmasky, M.E. Zinc: Multidimensional Effects on Living Organisms. Biomedicines 2021, 9, 208. https://doi.org/10.3390/biomedicines9020208
Cuajungco MP, Ramirez MS, Tolmasky ME. Zinc: Multidimensional Effects on Living Organisms. Biomedicines. 2021; 9(2):208. https://doi.org/10.3390/biomedicines9020208
Chicago/Turabian StyleCuajungco, Math P., Maria Soledad Ramirez, and Marcelo E. Tolmasky. 2021. "Zinc: Multidimensional Effects on Living Organisms" Biomedicines 9, no. 2: 208. https://doi.org/10.3390/biomedicines9020208
APA StyleCuajungco, M. P., Ramirez, M. S., & Tolmasky, M. E. (2021). Zinc: Multidimensional Effects on Living Organisms. Biomedicines, 9(2), 208. https://doi.org/10.3390/biomedicines9020208