
Effect of NIR Laser Therapy by MLS-MiS Source on Fibroblast Activation by Inflammatory Cytokines in Relation to Wound Healing

 ,
,  and
and 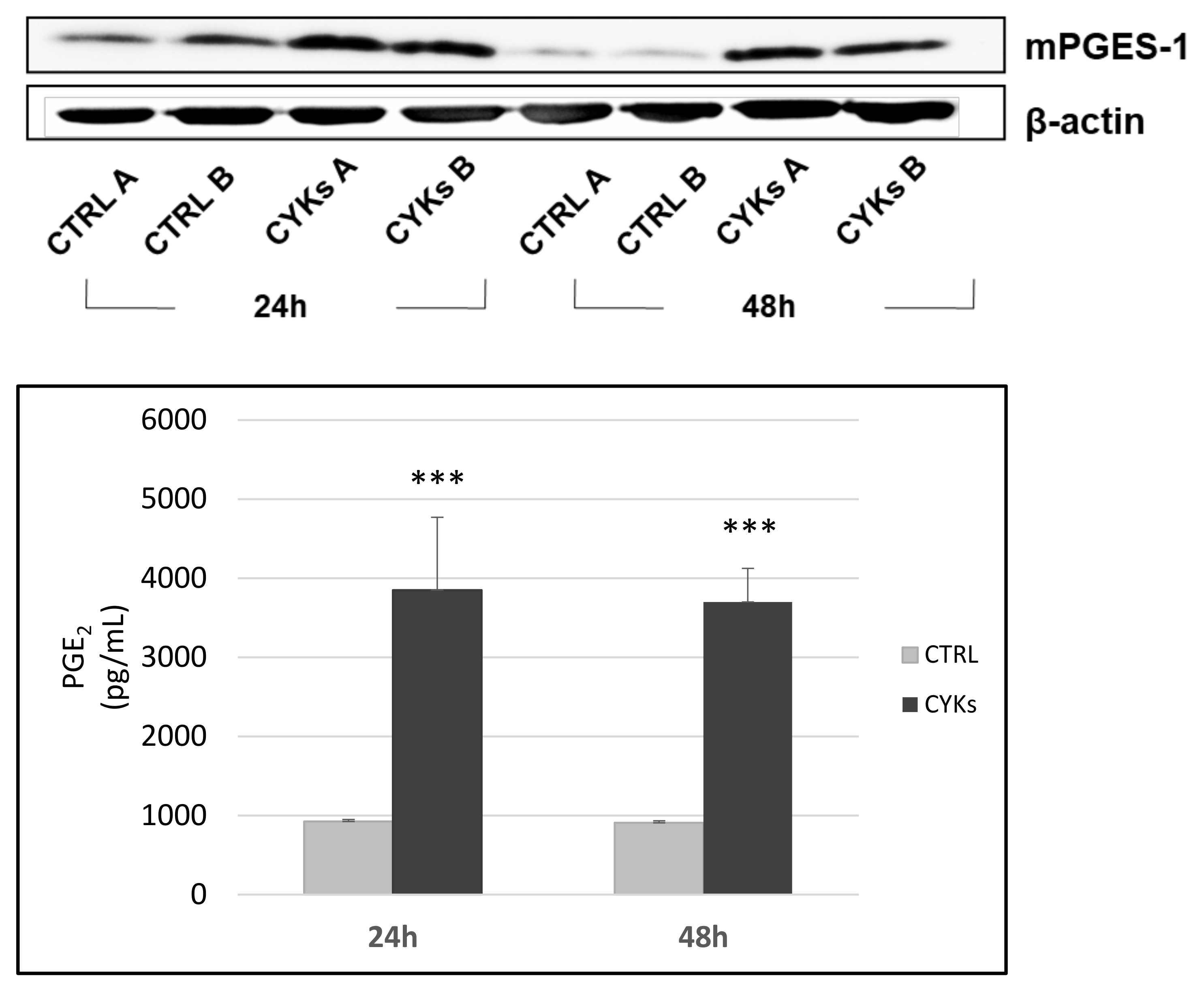{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. In Vitro Model of Inflammation
2.3. Laser Treatment
2.4. Experiment Design
- (i)
- samples stimulated with a mix of IL-1β and TNF-α for 48 h and then exposed to 3 laser treatments, repeated once a day, for 3 consecutive days under sterile conditions (CYKs + LASER Group);
- (ii)
- samples stimulated with a mix of IL-1β and TNF-α for 48 h and not exposed to laser treatments CYKs Group);
- (iii)
- samples not stimulated with a mix of IL-1β and TNF-α for 48 h and not exposed to laser treatments (CTRL Group).
2.5. Immunofluorescence Analysis
2.6. Western Blot
2.7. Immunoassays for Prostaglandin E-2 and VEGF Quantification
2.8. Statistics
3. Results
3.1. Set up of an “In Vitro” Inflammatory Model in Fibroblasts Cultures
3.2. Effect of Laser Treatment on Inflammatory Phenotype in Fibroblasts
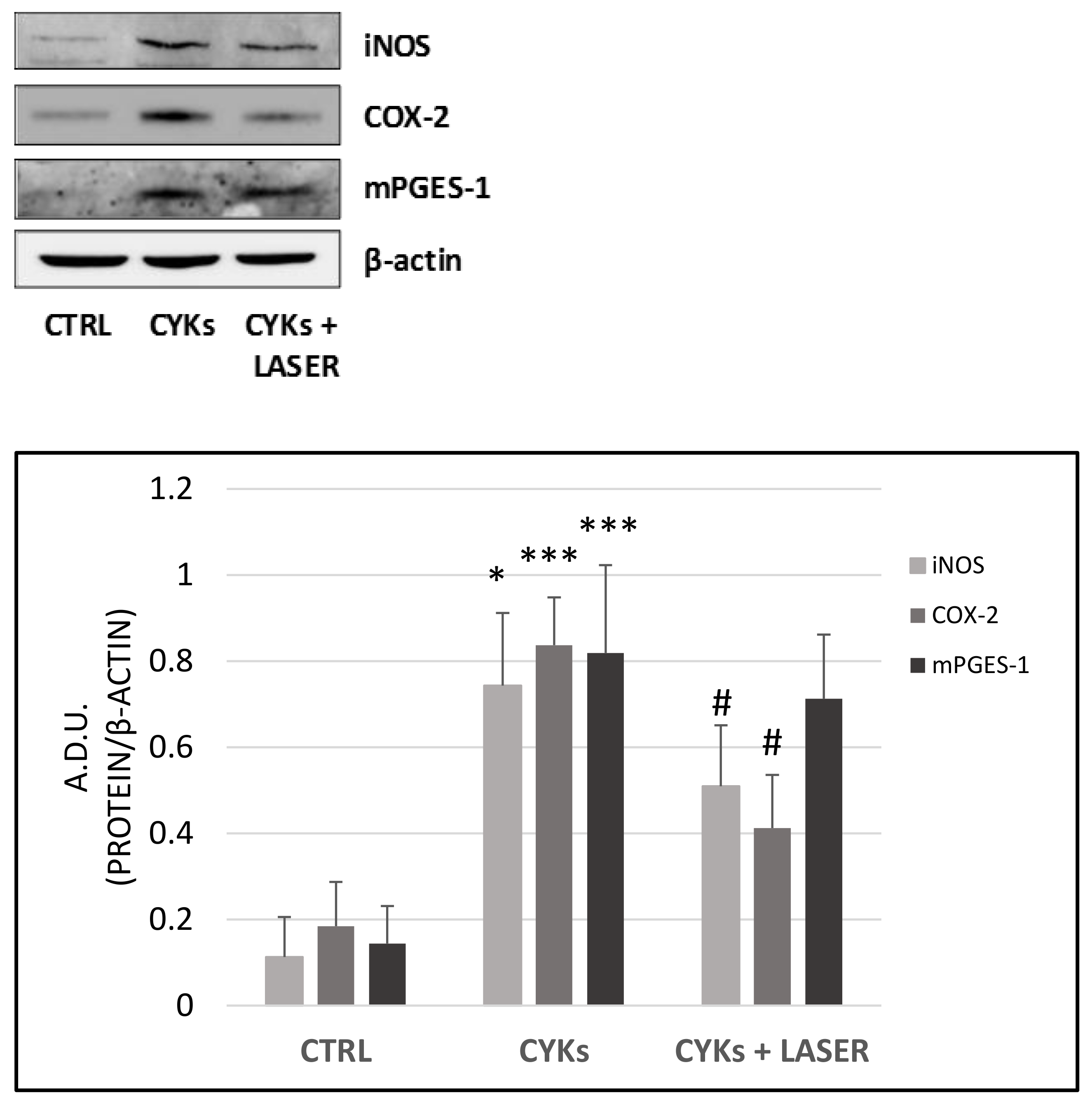
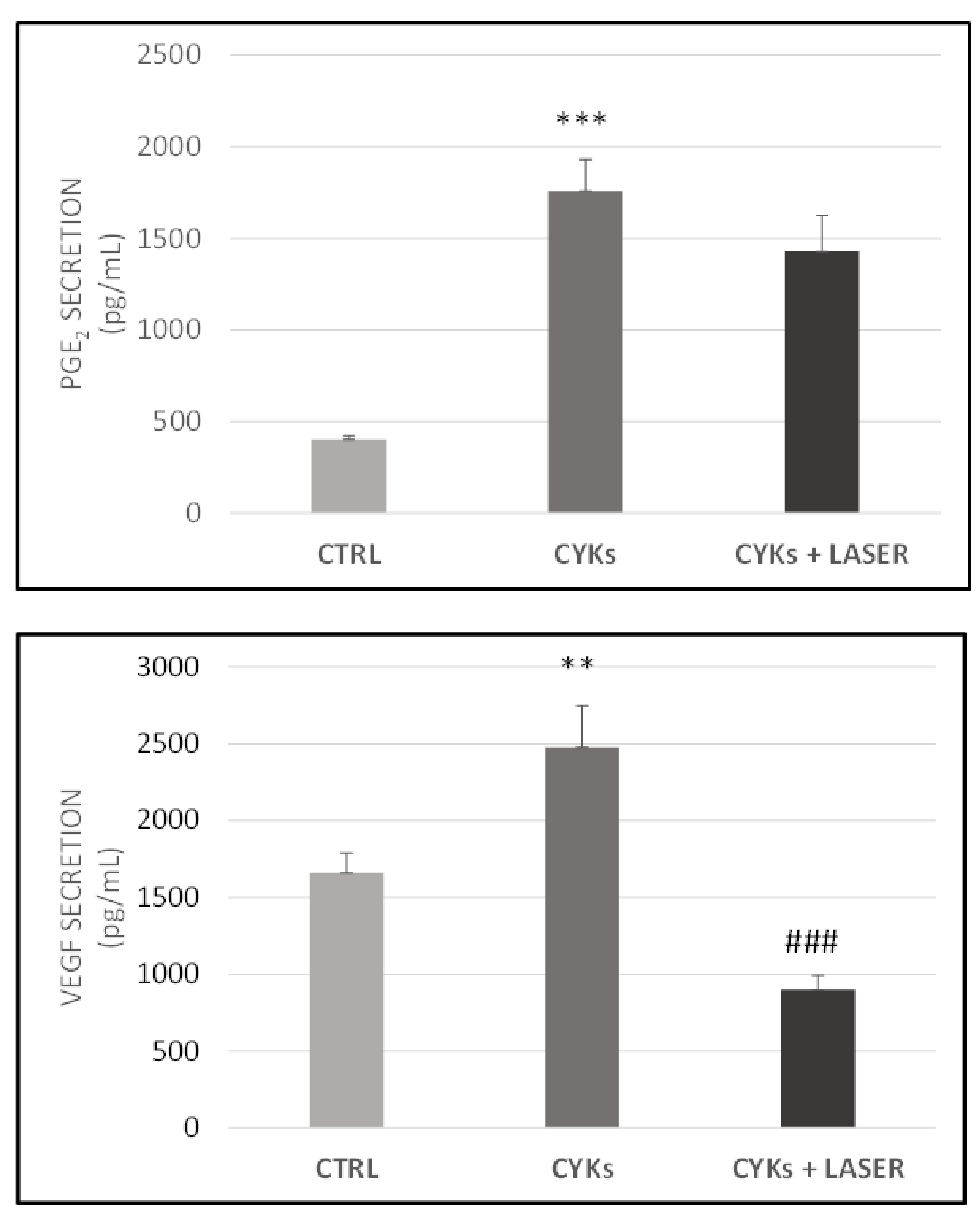
3.2.1. Expression of Inflammatory Markers
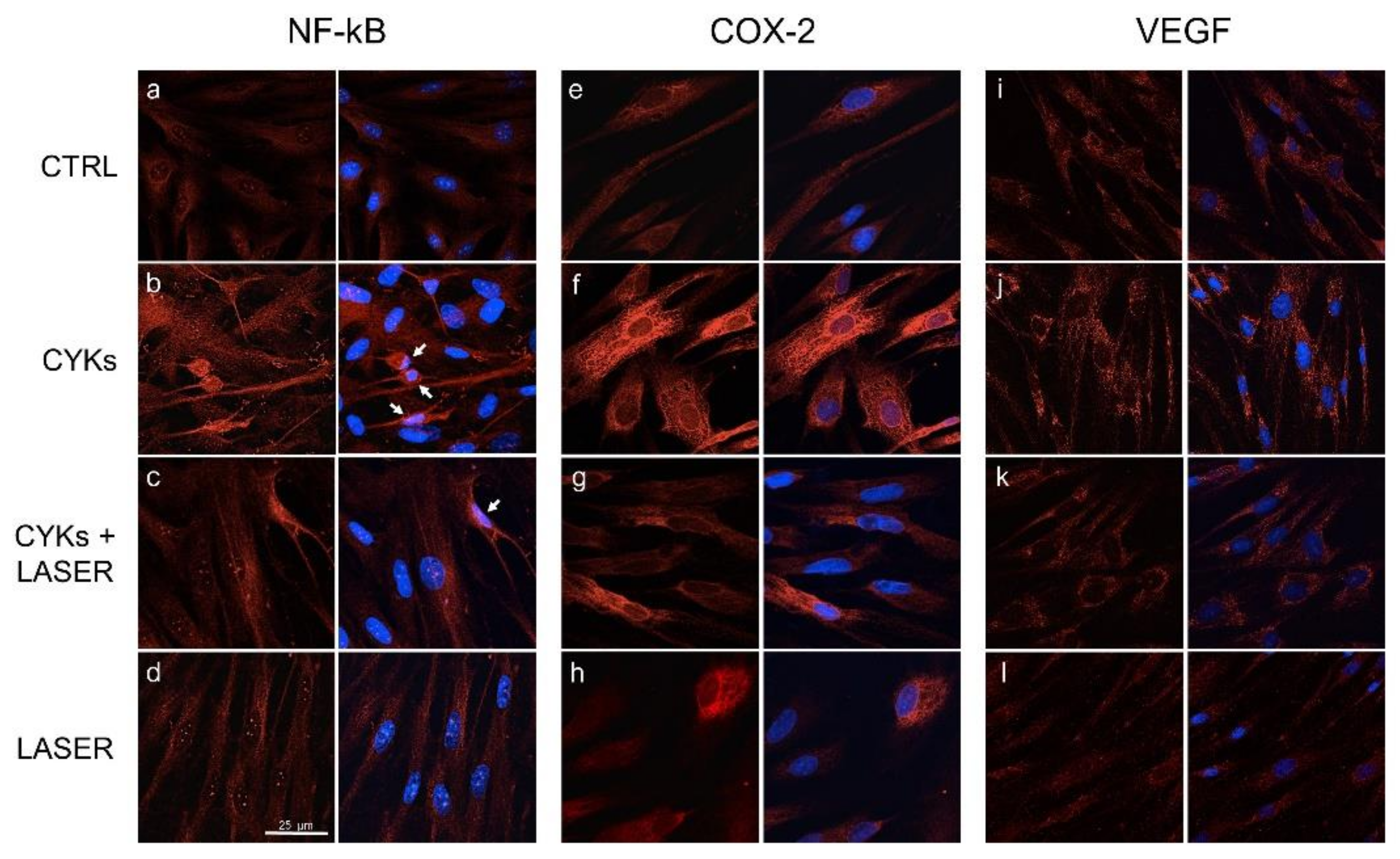
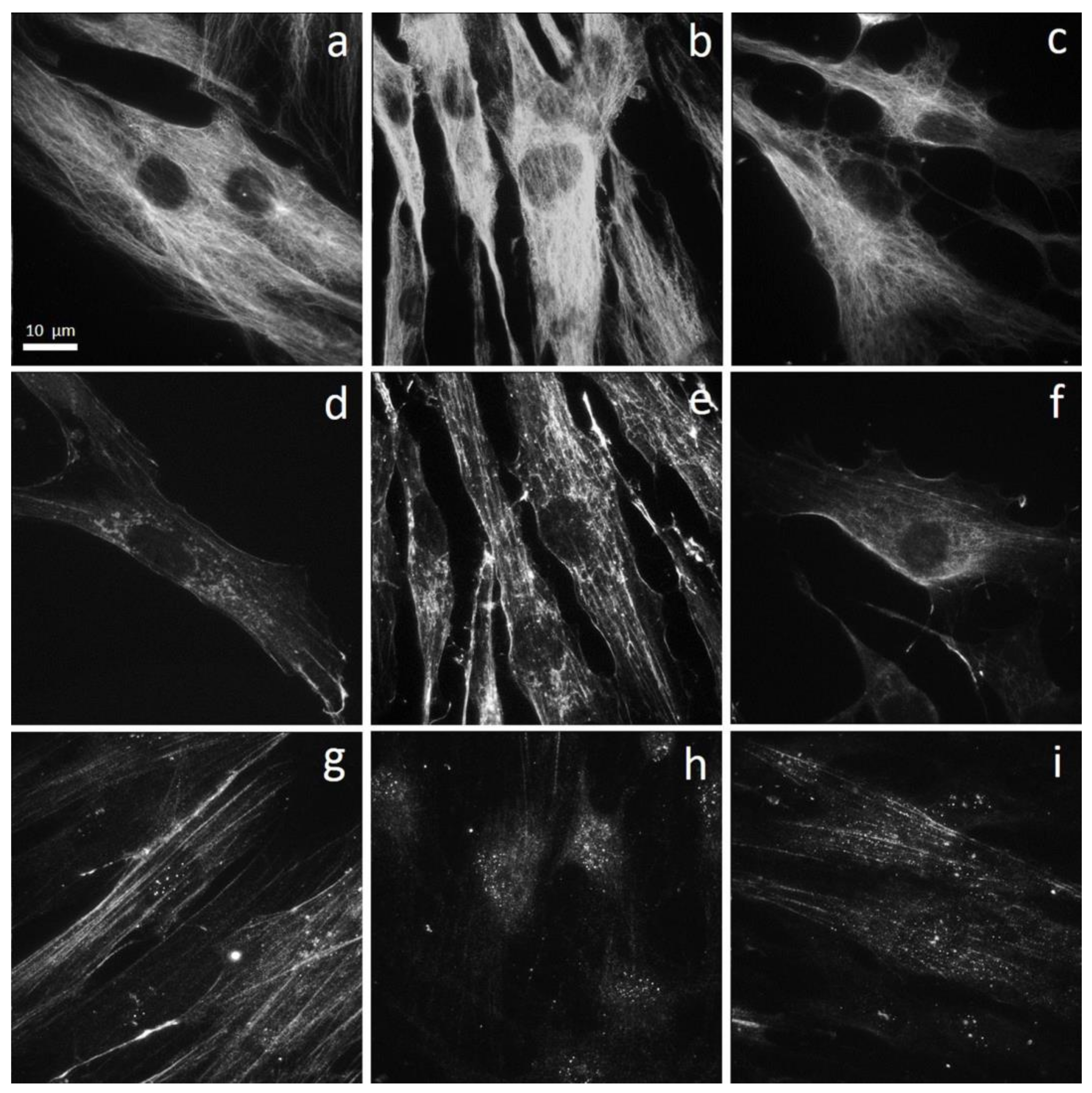
3.2.2. Morphology and Cytoskeleton Organization
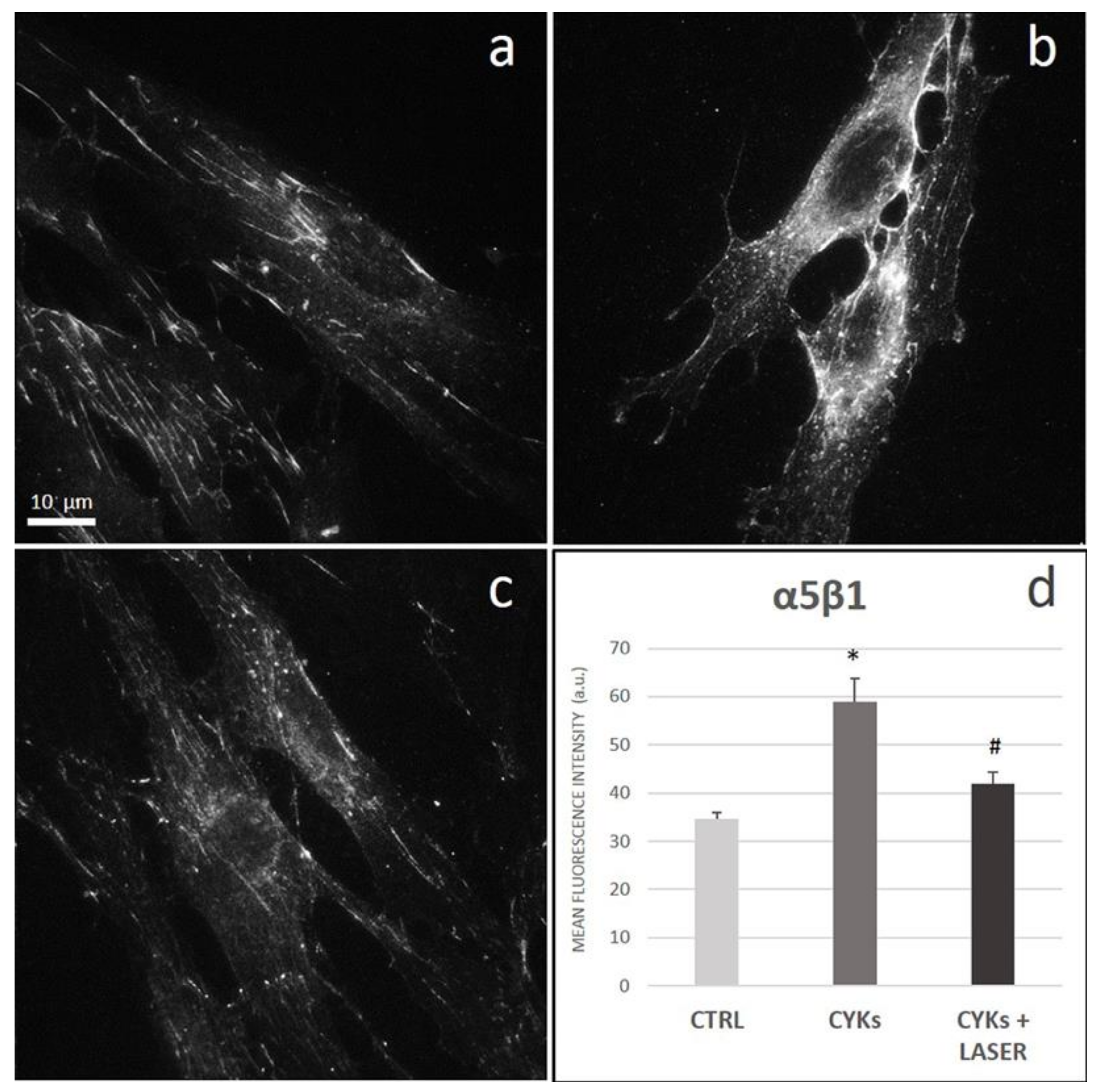
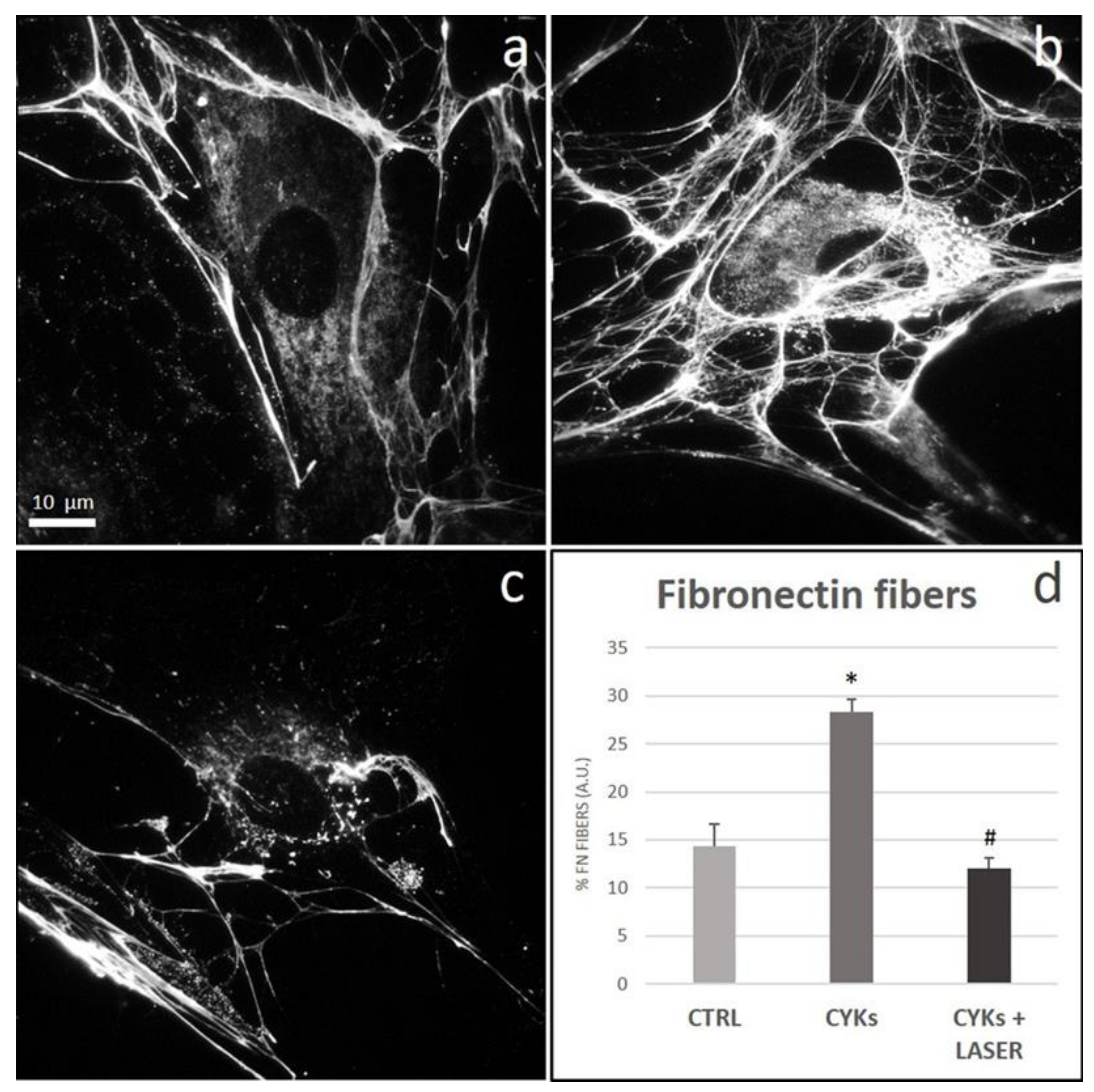
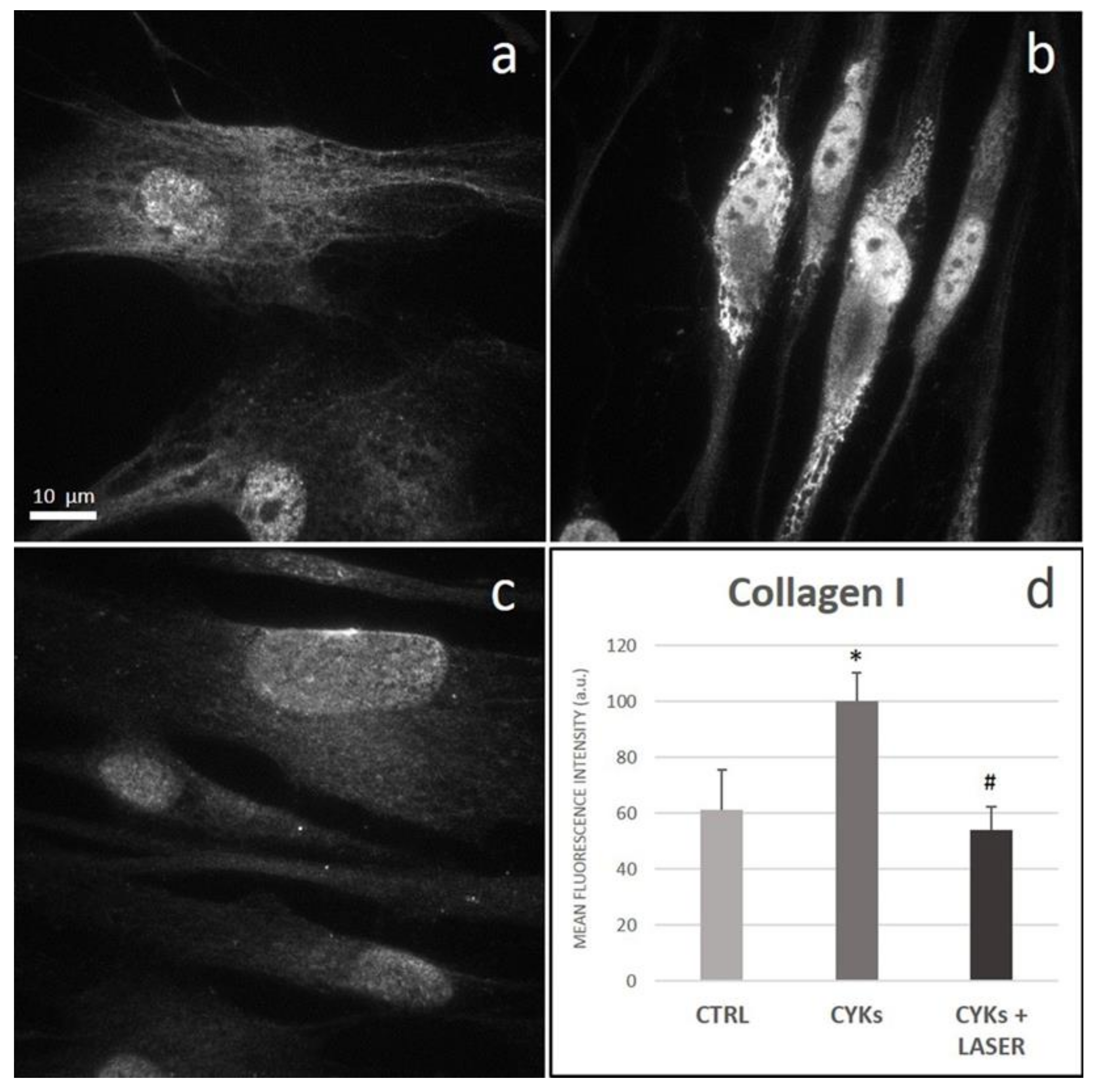
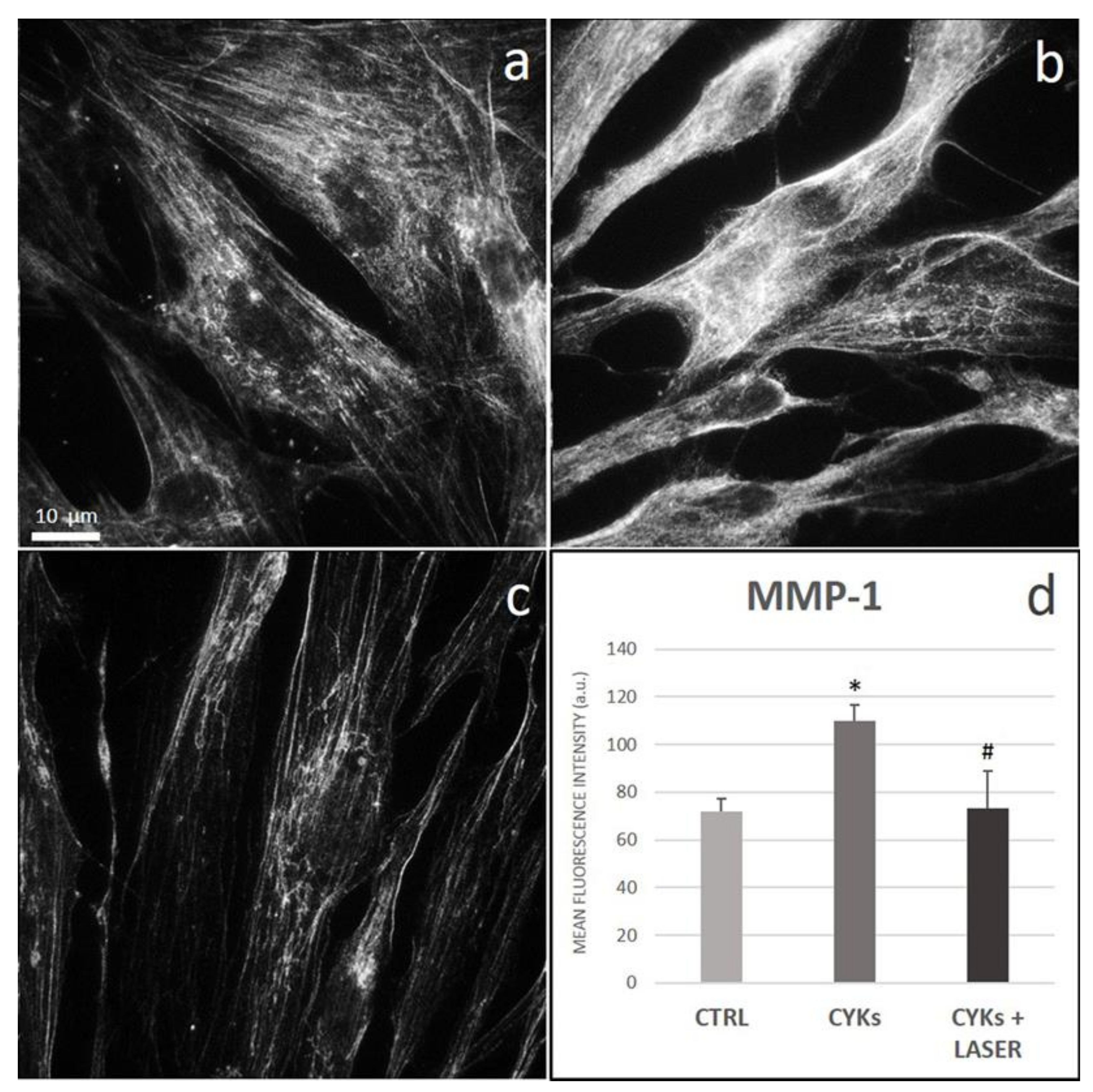
3.2.3. Extracellular Matrix Proteins and Membrane Integrin
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaw, T.J.; Martin, P. Wound repair: A showcase for cell plasticity and migration. Curr. Opin. Cell Biol. 2016, 42, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Wynn, T.A.; Vannella, K.M. Macrophages in tissue repair, regeneration, and fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Glaros, T. Macrophages and fibroblasts during inflammation, tissue damage and organ injury. Front. Biosci. 2009, 14, 3988–3993. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Sinha, M.; Datta, S.; Abas, M.; Chaffee, S.; Sen, C.K.; Roy, S. Monocyte and macrophage plasticity in tissue repair and regeneration. Am. J. Pathol. 2015, 185, 2596–2606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mescher, A.L. Macrophages and fibroblasts during inflammation and tissue repair in models of organ regeneration. Regenaration 2017, 4, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Bermudez, D.M.; Xu, J.; Herdrich, B.J.; Radu, A.; Mitchell, M.E.; Liechty, K.W. Inhibition of stromal cell-derived factor-1α further impairs diabetic wound healing. J. Vasc. Surg. 2011, 53, 774–784. [Google Scholar] [CrossRef] [Green Version]
- Wilgus, T.A.; DiPietro, L.A. Complex Rroles for VEGF in dermal wound healing. J. Investig. Dermatol. 2012, 132, 493–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernardo, M.E.; Fibbe, W.E. Mesenchymal stromal cells: Sensors and switchers of inflammation. Cell Stem Cell 2013, 13, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A. Cellular and molecular mechanisms of fibrosis. J. Pathol. 2007, 214, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desmouliere, A.; Darby, I.A.; Laverdet, B.; Bonté, F. Fibroblasts and myofibroblasts in wound healing. Clin. Cosmet. Investig. Dermatol. 2014, 7, 301–311. [Google Scholar] [CrossRef] [Green Version]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef]
- Grinnell, F.; Petroll, W.M. Cell motility and mechanics in three-dimensional collagen matrices. Annu. Rev. Cell Dev. Biol. 2010, 26, 335–361. [Google Scholar] [CrossRef]
- Dreifke, M.B.; Jayasuriya, A.A.; Jayasuriya, A.C. Current wound healing procedures and potential care. Mater. Sci. Eng. C 2015, 48, 651–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javed, Q.; Murtaza, I. Therapeutic potential of tumour necrosis factor-alpha antagonists in patients with chronic heart failure. Heart Lung Circ. 2013, 22, 323–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyadarshini, L.M.J.; Kishore Babu, E.P. Effect of low level laser therapy on diabetic foot ulcers: A randomized control trial. Int. Surg. J. 2018, 5, 1008–1015. [Google Scholar]
- Nesi-Reis, V.; Lera-Nonose, D.S.S.L.; Oyama, J.; Silva-Lalucci, M.P.P.; Demarchi, I.G.; Aristides, S.M.A.; Teixeira, J.J.V.; Silveira, T.G.V.; Lonardoni, M.V.C. Contribution of photodynamic therapy in wound healing: A systematic review. Photodiagnosis Photodyn. Ther. 2018, 21, 294–305. [Google Scholar] [CrossRef]
- Hamblin, M.R. Mechanisms and applications of the anti-inflammatory effects of photobiomodulation. AIMS Biophys. 2017, 4, 337–361. [Google Scholar] [CrossRef]
- Tantawy, S.A.; AbdelBasset, W.K.; Kamel, D.M.; Alrawaili, S.M.; Alsubaie, S.F. Laser photobiomodulation is more effective than ultrasound therapy in patients with chronic nonspecific low back pain: A comparative study. Lasers Med. Sci. 2019, 34, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Micheli, L.; Mannelli, L.D.C.; Lucarini, E.; Cialdai, F.; Vignali, L.; Ghelardini, C.; Monici, M. Photobiomodulation therapy by NIR laser in persistent pain: An analytical study in the rat. Lasers Med. Sci. 2017, 32, 1835–1846. [Google Scholar] [CrossRef] [PubMed]
- Cotler, H.B.; Chow, R.T.; Hamblin, M.R.; Carroll, J. The use of low level laser therapy (LLLT) For musculoskeletal pain. MOJ Orthop. Rheumatol. 2015, 2, 00068. [Google Scholar] [CrossRef]
- Enwemeka, C.S.; Parker, J.C.; Dowdy, D.S.; Harkness, E.E.; Harkness, L.E.; Woodruff, L.D. The efficacy of low-power lasers in tissue repair and pain control: A meta-analysis study. Photomed. Laser Surg. 2004, 22, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Monici, M.; Cialdai, F.; Ranaldi, F.; Paoli, P.; Boscaro, F.; Moneti, G.; Caselli, A. Effect of IR laser on myoblasts: A proteomic study. Mol. BioSyst. 2013, 9, 1147. [Google Scholar] [CrossRef]
- De Medeiros, M.L.; Araújo-Filho, I.; Da Silva, E.M.N.; Queiroz, W.S.D.S.; Soares, C.D.; De Carvalho, M.G.F.; Maciel, M.A.M. Effect of low-level laser therapy on angiogenesis and matrix metalloproteinase-2 immunoexpression in wound repair. Lasers Med. Sci. 2016, 32, 35–43. [Google Scholar] [CrossRef]
- Lee, J.-H.; Chiang, M.-H.; Chen, P.-H.; Ho, M.-L.; Lee, H.-E.; Wang, Y.-H. Anti-inflammatory effects of low-level laser therapy on human periodontal ligament cells: In vitro study. Lasers Med. Sci. 2018, 33, 469–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakata, S.; Kunimatsu, R.; Tsuka, Y.; Nakatani, A.; Hiraki, T.; Gunji, H.; Hirose, N.; Yanoshita, M.; Putranti, N.A.R.; Tanimoto, K. High-frequency near-infrared diode laser irradiation attenuates IL-1β;-induced expression of inflammatory cytokines and matrix metalloproteinases in human primary chondrocytes. J. Clin. Med. 2020, 9, 881. [Google Scholar] [CrossRef] [Green Version]
- Parks, W.C.; Wilson, C.L.; López-Boado, Y.S. Matrix metalloproteinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol. 2004, 4, 617–629. [Google Scholar] [CrossRef]
- Manicone, A.M.; McGuire, J.K. Matrix Metalloproteinases as Modulators of Inflammation. Semin. Cell Dev. Biol. 2008, 19, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Ciccone, V.; Monti, M.; Antonini, G.; Mattoli, L.; Burico, M.; Marini, F.; Maidecchi, A.; Morbidelli, L. Efficacy of AdipoDren® in reducing interleu-kin-1-induced lymphatic endothelial hyperpermeability. J. Vasc. Res. 2016, 53, 255–268. [Google Scholar] [CrossRef]
- Ciccone, V.; Terzuoli, E.; Donnini, S.; Giachetti, A.; Morbidelli, L.; Ziche, M. Stemness marker ALDH1A1 promotes tumor angio-genesis via retinoic acid/HIF-1α/VEGF signalling in MCF-7 breast cancer cells. J. Exp. Clin. Cancer Res. 2018, 37, 311. [Google Scholar] [CrossRef]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The Role of CC-cmokines in the regulation of angiogenesis. Int. J. Mol. Sci. 2016, 17, 1856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meiring, J.C.; Akhmanova, A. Microtubules keep large cells in shape. J. Cell Biol. 2020, 219. [Google Scholar] [CrossRef]
- Olczyk, P.; Mencner, Ł.; Komosinska-Vassev, K. The role of the extracellular matrix components in cutaneous wound healing. BioMed Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lenselink, A.E. Role of fibronectin in normal wound healing. Int. Wound J. 2013, 12, 313–316. [Google Scholar] [CrossRef]
- Caley, M.P.; Martins, V.L.; O’Toole, E.A. Metalloproteinases and wound healing. Adv. Wound Care 2015, 4, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Theiss, A.L.; Simmons, J.G.; Jobin, C.; Lund, P.K. Tumor necrosis factor (TNF) α; increases collagen accumulation and proliferation in intestinal myofibroblasts via TNF receptor 2. J. Biol. Chem. 2005, 280, 36099–36109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jiang, J.; Xie, J.; Xu, C.; Wang, C.; Yin, L.; Yang, L.; Sung, K.L.P. Combined effects of tumor necrosis factor α and inter-leukin 1β on lysyl oxidase and matrix metalloproteinase expression in human knee synovial fibroblasts in vitro. Exp. Ther. Med. 2017, 14, 5258–5266. [Google Scholar]
- Zhu, Y.; Zhu, M.; Lance, P. iNOS signaling interacts with COX-2 pathway in colonic fibroblasts. Exp. Cell Res. 2012, 318, 2116–2127. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, L.M.; Pansani, T.N.; Hebling, J.; Costa, C.A.D.S.; Basso, F.G. Photobiomodulation of inflammatory-cytokine-related effects in a 3-D culture model with gingival fibroblasts. Lasers Med. Sci. 2020, 35, 1205–1212. [Google Scholar] [CrossRef]
- Cunha, J.L.S.; Carvalho, F.M.D.A.D.; Filho, R.N.P.; Ribeiro, M.A.G.; De Albuquerque-Júnior, R.L.C. Effects of different protocols of low-level laser therapy on collagen deposition in wound healing. Braz. Dent. J. 2019, 30, 317–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maloney, J.P.; Gao, L. Proinflammatory cytokines increase vascular endothelial growth factor expression in alveolar epithelial cells. Mediat. Inflamm. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bao, P.; Kodra, A.; Tomic-Canic, M.; Golinko, M.S.; Ehrlich, H.P.; Brem, H. The role of vascular endothelial growth factor in wound healing. J. Surg. Res. 2009, 153, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojadinovic, O.K.A.; Golinko, M.; Tomic-Canic, M.; Brem, H. A novel, non-angiogenic mechanism of VEGF: Stimulation of keratinocyte and fibroblast migration. Wound Repair Regen. 2007, 15, A30. [Google Scholar]
- Szymanska, J.; Goralczyk, K.; Klawe, J.J.; Lukowicz, M.; Michalska, M.; Goralczyk, B.; Zalewski, P.; Newton, J.L.; Gryko, L.; Zajac, A.; et al. Phototherapy with low-level laser influences the proliferation of endothelial cells and vascular endothelial growth factor and transforming growth factor-beta secretion. J. Physiol. Pharmacol. 2013, 64. [Google Scholar]
- Cury, V.; Moretti, A.I.S.; Assis, L.; Bossini, P.; Crusca, J.D.S.; Neto, C.B.; Fangel, R.; De Souza, H.P.; Hamblin, M.R.; Parizotto, N.A. Low level laser therapy increases angiogenesis in a model of ischemic skin flap in rats mediated by VEGF, HIF-1α and MMP-2. J. Photochem. Photobiol. B Biol. 2013, 125, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhmanova, A.; Steinmetz, M.O. Control of microtubule organization and dynamics: Two ends in the limelight. Nat. Rev. Mol. Cell Biol. 2015, 16, 711–726. [Google Scholar] [CrossRef]
- Ricci, R.; Pazos, M.C.; Borges, R.E.; Pacheco-Soares, C. Biomodulation with low-level laser radiation induces changes in endothelial cell actin filaments and cytoskeletal organization. J. Photochem. Photobiol. B Biol. 2009, 95, 6–8. [Google Scholar] [CrossRef]
- Sandbo, N.; Dulin, N. Actin cytoskeleton in myofibroblast differentiation: Ultrastructure defining form and driving function. Transl. Res. 2011, 158, 181–196. [Google Scholar] [CrossRef] [Green Version]
- Storch, K.N.; Taatjes, D.J.; Bouffard, N.A.; Locknar, S.; Bishop, N.M.; Langevin, H.M. Alpha smooth muscle actin distribution in cytoplasm and nuclear invaginations of connective tissue fibroblasts. Histochem. Cell Biol. 2007, 127, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Langevin, H.M.; Storch, K.N.; Snapp, R.R.; Bouffard, N.A.; Badger, G.J.; Howe, A.K.; Taatjes, D.J. Tissue stretch induces nuclear remodeling in connective tissue fibroblasts. Histochem. Cell Biol. 2010, 133, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Malhas, A.N.; Vaux, D.J. The nuclear envelope and its involvement in cellular stress responses. Biochem. Soc. Trans. 2011, 39, 1795–1798. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, M.T.; Han, Y.-P.; Yan, C.; Shaw, M.C.; Garner, W.L. TNF-α suppresses α-smooth muscle actin expression in human dermal fibroblasts: An implication for abnormal wound healing. J. Investig. Dermatol. 2007, 127, 2645–2655. [Google Scholar] [CrossRef] [Green Version]
- Houreld, N.N.; Ayuk, S.M.; Abrahamse, H. Expression of genes in normal fibroblast cells (WS1) in response to irradiation at 660nm. J. Photochem. Photobiol. B Biol. 2014, 130, 146–152. [Google Scholar] [CrossRef]
- Chellini, F.; Tani, A.; Zecchi-Orlandini, S.; Giannelli, M.; Sassoli, C. In vitro evidences of different fibroblast morpho-functional responses to red, near-infrared and violet-blue photobiomodulation: Clues for addressing wound healing. Appl. Sci. 2020, 10, 7878. [Google Scholar] [CrossRef]
- Illescas-Montes, R.; Melguizo-Rodríguez, L.; García-Martínez, O.; De Luna-Bertos, E.; Manzano-Moreno, F.J.; Ruiz, C.; Ramos-Torrecillas, J. Human fibroblast gene expression modulation using 940 NM diode laser. Sci. Rep. 2019, 9, 12037–12047. [Google Scholar] [CrossRef] [PubMed]
- Medrado, A.R.; Andrade, Z.A. Influence of low level laser therapy on wound healing and its biological action upon myofibroblasts. Lasers Surg. Med. 2003, 32, 239–244. [Google Scholar] [CrossRef]
- El Ayadi, A.; Jay, J.W.; Prasai, A. Current approaches targeting the wound healing phases to attenuate fibrosis and scarring. Int. J. Mol. Sci. 2020, 21, 1105. [Google Scholar] [CrossRef] [Green Version]
- Koivisto, L.; Heino, J.; Häkkinen, L.; Larjava, H. Integrins in wound healing. Adv. Wound Care 2014, 3, 762–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakkinen, L.; Heino, J.; Koivisto, L.; Larjava, H. Altered interaction of human granulation-tissue fibroblasts with fibronectin is regulated by alpha 5 beta 1 integrin. Biochim. Biophys. Acta 1994, 1224, 33–42. [Google Scholar] [CrossRef]
- Robinson, E.E.; Zazzali, K.M.; Corbett, S.A.; Foty, R.A. alpha5beta1 integrin mediates strong tissue cohesion. J. Cell Sci. 2002, 116, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Limb, G.A.; Matter, K.; Murphy, G.; Cambrey, A.D.; Bishop, P.N.; Morris, G.E.; Khaw, P.T. Matrix metalloproteinase-1 associates with intracellular organelles and confers resistance to lamin A/C degradation during apoptosis. Am. J. Pathol. 2005, 166, 1555–1563. [Google Scholar] [CrossRef] [Green Version]
- Bildyug, N. Matrix metalloproteinases: An emerging role in regulation of actin microfilament system. Biomol. Concepts 2016, 7, 321–329. [Google Scholar] [CrossRef]
- Distler, J.H.W.; Schett, G.; Gay, S.; Distler, O. The controversial role of tumor necrosis factor α in fibrotic diseases. Arthritis Rheum. 2008, 58, 2228–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thampatty, B.P.; Li, H.; Im, H.-J.; Wang, J.H.-C. EP4 receptor regulates collagen type-I, MMP-1, and MMP-3 gene expression in human tendon fibroblasts in response to IL-1β treatment. Gene 2007, 386, 154–161. [Google Scholar] [CrossRef] [Green Version]
- Lindner, D.; Zietsch, C.; Becher, P.M.; Schulze, K.; Schultheiss, H.-P.; Tschöpe, C.; Westermann, D. Differential expression of matrix metalloproteases in human fibroblasts with different origins. Biochem. Res. Int. 2012, 2012, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mauviel, A.; Heino, J.; Kähäri, V.-M.; Hartmann, D.-J.; Eacute; Loyau, R. Comparative effects of interleukin-1 and tumor necrosis factor-α on collagen production and corresponding procollagen mrna levels in human dermal fibroblasts. J. Investig. Dermatol. 1991, 96, 243–249. [Google Scholar] [CrossRef] [Green Version]
- Chizzolini, C.; Parel, Y.; De Luca, C.; Tyndall, A.; Åkesson, A.; Scheja, A.; Dayer, J.-M. Systemic sclerosis Th2 cells inhibit collagen production by dermal fibroblasts via membrane-associated tumor necrosis factor α. Arthritis Rheum. 2003, 48, 2593–2604. [Google Scholar] [CrossRef] [Green Version]
- Greenwel, P.; Tanaka, S.; Penkov, D.; Zhang, W.; Olive, M.; Moll, J.; Vinson, C.; Di Liberto, M.; Ramirez, F. Tumor necrosis factor alpha inhibits type I collagen synthesis through repressive ccaat/enhancer-binding proteins. Mol. Cell. Biol. 2000, 20, 912–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vesey, D.A.; Cheung, C.; Cuttle, L.; Endre, Z.; Gobe, G.C.; Johnson, D.W. Interleukin-1β stimulates human renal fibroblast proliferation and matrix protein production by means of a transforming growth factor-β-dependent mechanism. J. Lab. Clin. Med. 2002, 140, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Monici, M.; Cialdai, F. The role of physical Factors in cell differentiation, tissue repair and regeneration. In Tissue Regeneration-From Basic Biology to Clinical Application; IntechOpen: London, UK, 2012; pp. 13–34. [Google Scholar]
- Houreld, N.N.; Ayuk, S.M.; Abrahamse, H. Cell adhesion molecules are mediated by photobiomodulation at 660 nm in diabetic wounded fibroblast cells. Cells 2018, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Keshri, G.K.; Gupta, A.; Yadav, A.; Sharma, S.K.; Singh, S.B. Photobiomodulation with pulsed and continuous wave near-infrared laser (810 NM, al-GA-as) Augments dermal wound healing in immunosuppressed rats. PLoS ONE 2016, 11, e0166705. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, D.H.; Abrahamse, H. The role of laser fluence in cell viability, proliferation, and membrane integrity of wounded human skin fibroblasts following helium-neon laser irradiation. Lasers Surg. Med. 2006, 38, 74–83. [Google Scholar] [CrossRef]
- Lee, J.-H.; Jekal, S.-J.; Kwon, A.P.-S. 630 nm light emitting diode irradiation improves dermal wound healing in rats. J. Korean Phys. Ther. 2015, 27, 140–146. [Google Scholar] [CrossRef]
- Squarzoni, P.; Bani, D.; Cialdai, F.; Monici, M. NIR laser therapy in the management of feline stomatitis. SM Dermatol. J. 2017, 3, 1–11. [Google Scholar] [CrossRef]
- Micheli, L.; Cialdai, F.; Pacini, A.; Branca, J.J.V.; Morbidelli, L.; Ciccone, V.; Lucarini, E.; Ghelardini, C.; Monici, M.; Mannelli, L.D.C. Effect of NIR laser therapy by MLS-MiS source against neuropathic pain in rats: In vivo and ex vivo analysis. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Genah, S.; Cialdai, F.; Ciccone, V.; Sereni, E.; Morbidelli, L.; Monici, M. Effect of NIR Laser Therapy by MLS-MiS Source on Fibroblast Activation by Inflammatory Cytokines in Relation to Wound Healing. Biomedicines 2021, 9, 307. https://doi.org/10.3390/biomedicines9030307
Genah S, Cialdai F, Ciccone V, Sereni E, Morbidelli L, Monici M. Effect of NIR Laser Therapy by MLS-MiS Source on Fibroblast Activation by Inflammatory Cytokines in Relation to Wound Healing. Biomedicines. 2021; 9(3):307. https://doi.org/10.3390/biomedicines9030307
Chicago/Turabian StyleGenah, Shirley, Francesca Cialdai, Valerio Ciccone, Elettra Sereni, Lucia Morbidelli, and Monica Monici. 2021. "Effect of NIR Laser Therapy by MLS-MiS Source on Fibroblast Activation by Inflammatory Cytokines in Relation to Wound Healing" Biomedicines 9, no. 3: 307. https://doi.org/10.3390/biomedicines9030307
APA StyleGenah, S., Cialdai, F., Ciccone, V., Sereni, E., Morbidelli, L., & Monici, M. (2021). Effect of NIR Laser Therapy by MLS-MiS Source on Fibroblast Activation by Inflammatory Cytokines in Relation to Wound Healing. Biomedicines, 9(3), 307. https://doi.org/10.3390/biomedicines9030307