
Cardiac Organoids to Model and Heal Heart Failure and Cardiomyopathies


Abstract
:1. Introduction
2. Existing Models of Cardiomyopathies
- Restrictive cardiomyopathy (RCM) is rarer and is defined by nondilated left or right ventricles with diastolic dysfunction but near to normal systolic function. Arrhythmias and conduction abnormalities often occur [28].
- Arrhythmogenic right ventricular cardiomyopathy (ARVC) involves ventricular tachycardia or fibrillation and sudden cardiac death. It is characterized by a progressive loss of right ventricular myocardium, its replacement by fibrofatty tissue, and arrhythmias [29].
3. Cardiac Organoids to Reproduce the Features of Human Cardiac Tissue
3.1. The Physiological Features of Human Cardiac Tissue
- Vascular smooth muscle cells and pericytes, which sustain the vascular network and control the vascular tone [51];
- Neurons, which convey autonomic control of cardiac function through a brain–heart axis [54];
- Other rarer cells that can have key roles in the regulation of specific functions such as regeneration or fibrotic cardiac remodeling [57].
3.2. Modeling the Cell Composition in Engineered Cardiac Tissues (ECTs)
3.2.1. Different Types of Cardiomyocytes
3.2.2. Different Types of Non-Myocyte Cells
3.3. Modeling the Complex Organization of the Heart in ECTs
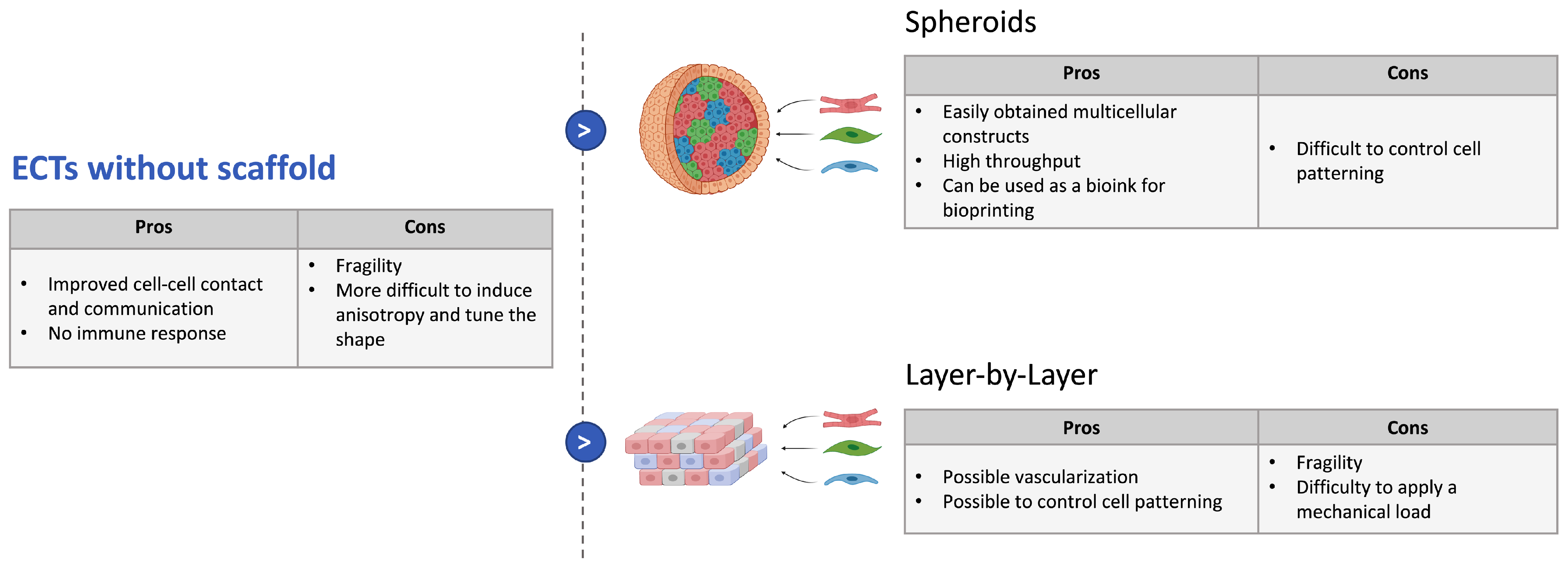
3.3.1. Strategies without Scaffold
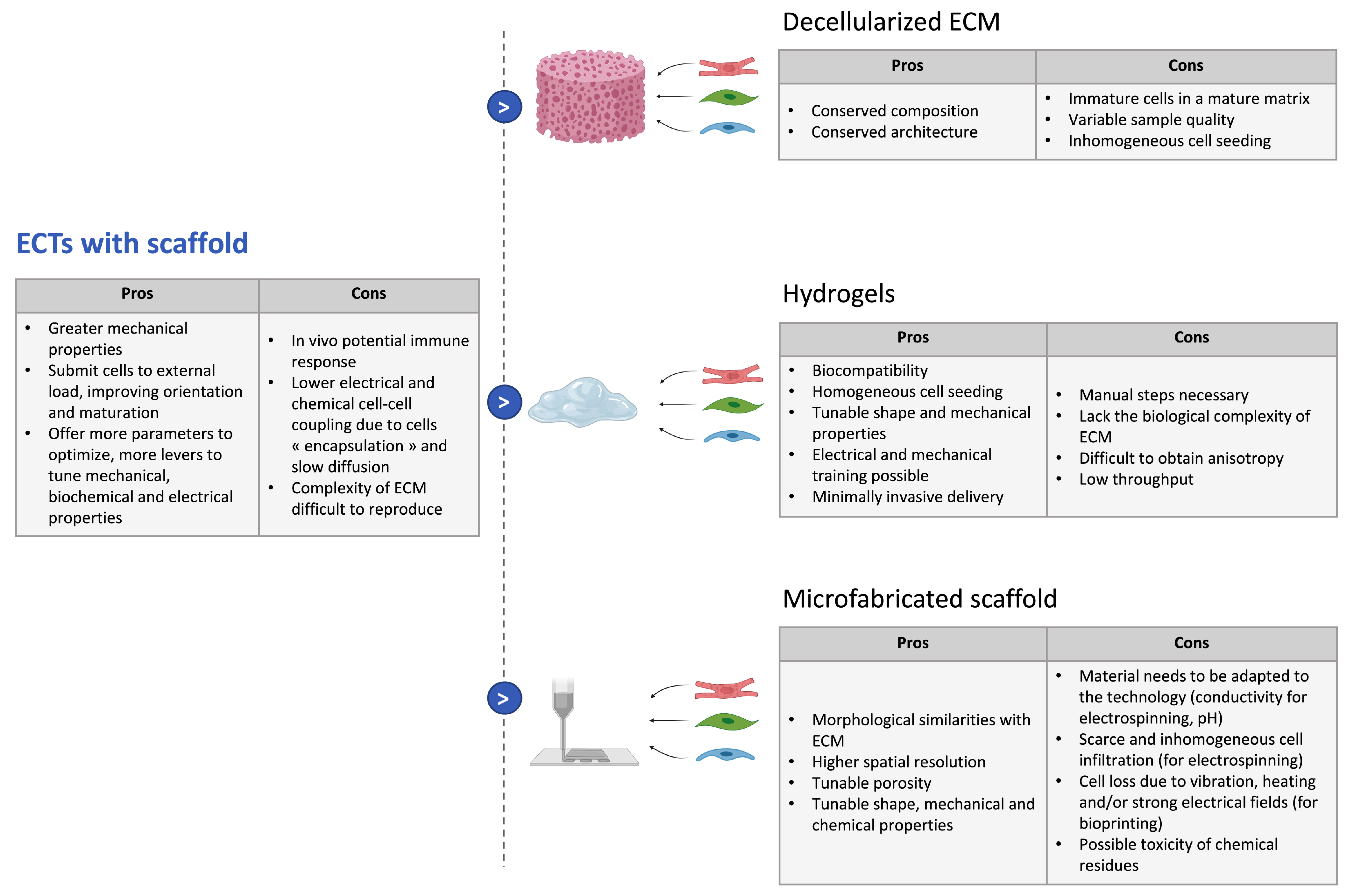
3.3.2. Strategies with Scaffolds
- Cell-seeding on decellularized extracellular matrix:As the ECM is a highly complex structural and functional environment that determines cell organization and function, decellularized ECMs have been considered ideal biomaterials in tissue engineering as they offer a solid organized scaffold [105]. This way, the extracellular matrix complex composition, bioactivity, architecture, and vascular tree are kept intact [106] and cells can be reseeded in the scaffold obtained. The decellularization protocols can be enzymatic, chemical, or physical and have been reviewed elsewhere [107]. Decellularized ECMs (dECMs) have been often used as myocardial slices, seeded with hiPSC-CMs and potentially stromal cells to form cardiac patches [108]. In [106], Guyette et al. obtained a more 3D physiological model by reseeding fibers with hiPSC-CMs, which then showed spontaneous contraction. Finally, experiments of whole heart recellularization have been carried out [109].However, the success of this approach in order to build cardiac constructs is largely dependent on the quality of the decellularization, which remains variable across the samples. Moreover, processes tend to be long and a compromise has to be made between the removal of all cells and the preservation of ECM integrity [110]. Additionally, as the ECM surrounds the cells, keeping the intricate ECM network intact may impede proper cell seeding of decellularized material. Therefore, to further maximize cell seeding, ECM postprocessing protocols have been developed and the ECM can now also be solubilized in hydrogels employed as a biological ink in bioprinting [111].
- Seeding in liquid hydrogels:Hydrogels are 3D networks of polymers that can absorb large amounts of fluids, in general, water. Their biocompatibility, good diffusion properties, and high permeability for oxygen, nutrients, and other water-soluble metabolites make them suitable scaffolds for tissue engineering [112]. Moreover, one can control their microscale structure and can tune their mechanical properties to mimic the native ECM by introducing crosslinks and by playing on the polymer molecular weight or density [102]. The main asset of hydrogel scaffolds in tissue engineering is the fact that cells can be mixed into liquid hydrogels, allowing for homogeneous seeding throughout the scaffold, and the mix can be cast to tailor the tissue geometry for the desired application. Once the hydrogel polymerizes, it encapsulates the cells to obtain a tissue with a complex geometry and 3D cell–cell interactions. Hydrogels also enable minimally invasive delivery for cardiac repair: they have already been used for direct injection with hiPSC-CM to regenerate ischemic myocardium and allowed for better cell retention and graft viability [20,107].Several materials are used for the fabrication of hydrogel scaffolds for cardiac tissue engineering. They can be natural polymers such as fibrin, collagen, gelatin, chitosan, hyaluronic acid (HA), matrigel, or even directly dECM, as explained earlier. These natural polymers all present an inherent biocompatibility and biodegradability and can mimic components of the ECM. However, they present batch-to-batch variability and are therefore more difficult to control [113]. Synthetic polymers have also been used to build engineered cardiac tissues; the most commonly used are poly(ethylene glycol) (PEG), poly(vinyl alcohol) (PVA), and poly(2-hydroxyethyl methacrylate) (PHEMA) [114]. Those synthetic polymeric possess more reproducible physical and chemical properties but generally show lower biocompatibility and cell attachment. Cell adhesion can however be improved by molecular modifications and the addition of short peptides to mimic aspects of native cell–ECM interactions [115].To obtain a cardiac-specific anisotropic structure, ECTs made with hydrogel scaffolds can be submitted to physical cues. Mechanical stimulation has been proven to improve the contractility of engineered heart tissues and to result in enhanced maturation of hiPSC-CMs, with better sarcomere organization, improved cell–cell junctions and t-tubules network, as well as higher amplitudes for twitch force and action potentials [116]. Various types of mechanical stimulation have been studied. The most common technique is passive stretch, by seeding hiPSC-CM (potentially mixed with other cells) between two PDMS posts [80,117], but a cyclic stretch, imitating the successive filling and emptying steps of the cardiac cycle, can also further enhance the maturation and structure of the ECTs, as shown in [118]. Several studies also demonstrated that electrical stimulation induces higher force generation and improved alignment and cell–cell junction in ECTs [119,120,121]. Finally, combinations of mechanical and electrical stimulation have been implemented on ECTs, as these signals are coupled in the native heart tissue; these lead to improved functional properties compared to electrical or mechanical stimulation only [122,123]. For more examples, one can refer to other recent reviews [18,116]. Bioactive molecules can also be added in the hydrogel to regulate cell adhesion, proliferation, or differentiation or to improve vascularization [114].
- Microfabrication of solid materials:The scaffold can be patterned to better control the 3D environment of the cells and to make it more reminiscent of the endogenous cardiac tissue. This patterning can also enhance vascularization and improve cell survival when implanted in vivo. To achieve this, the fibrillar aspect of the ECM should be reproduced, and this requires complete control over the micron- to nanometer-scale features of the scaffold [124]. Several technologies of micro- and nano-fabrication have been investigated in that prospect such as solvent casting/particulate leaching, gas foaming, freeze drying, thermally induced phase separation, photolithography, electrospinning, or 3D printing. Those have been comprehensively reviewed elsewhere [124,125]. Those techniques allow us to control the chemistry, topography, elasticity, and conductivity of the scaffold. Among those processes, electrospinning and 3D bioprinting currently experience fast development. Electrospinning is a process that consists in using electrostatic forces to produce nanofibers from a polymer solution [126]. The fibers are deposited on a collector to form a nanoporous scaffold with a tailored shape composed of fibers ranging from the micro- to nano-scale and showing morphological similarities with ECM and high porosity with variable pore-size distribution. Moreover, the fibers can be functionalized with different ECM-derived proteins [127]. These properties are very attractive for tissue engineering. Indeed, in [128], iPSC-CMs were seeded on electrospun dextran vinyl sulfone (DVS) fibers with varying biochemical, architectural, and mechanical properties, and it was shown that the optimal configuration was fibronectin-functionalized DVS matrices with highly aligned fibers and low stiffness. The tissues generated on these scaffolds even demonstrated better maturation than micropatterned fibronectin lines. Many other materials and geometries were tested in other studies, and the electrospun scaffold always improved cell alignment and cardiac function [129]. Moreover, electrospinning is a technology that can be up-scaled for industrial application, which provides hope for potential off-shelf scaffolds for cardiac repair [130].Bioprinting is a layer-by-layer additive manufacturing technology that allows the user to print biological material with a defined pattern [131]. Usually, bioinks consist of previously described hydrogels seeded with cells, in which co-factors can be added. This technology allows for a greater complexity in composition and a higher spatial resolution, as it has enabled to print several inks for the same tissue. Indeed, in [132], a 3D bioprinting method based on extrusion was developed to print collagen using freeform reversible embedding of suspended hydrogels. Components of the human heart at various scales could then be reproduced. A human cardiac ventricle model could even be printed, using collagen ink to form external shells and a high-concentration cell ink composed of human embryonic stem cell-derived cardiomyocytes (hESC-CMs) and 2% cardiac fibroblasts between the walls. This ventricle showed synchronized contractions after 7 days, anisotropic action potential propagation, and wall thickening up to 14% during systole. Hence, this method gives encouraging results for the production of complex cardiac models. Several other techniques have been developed for bioprinting and have been thoroughly reviewed in a recent article [131]. Despite all of their advantages, microfabricated scaffolds have limitations: they are less straightforward to make, as they require additional optimization of the material to the chosen technique; they can contain cytotoxic chemical residues; and when the scaffold is fabricated first and the cells are seeded afterwards, it can lead to scarce and inhomogeneous cell infiltration. On the contrary, in the case of bioprinting for which the cells are mixed into the bioink, it can induce cell loss due to method-specific processes.
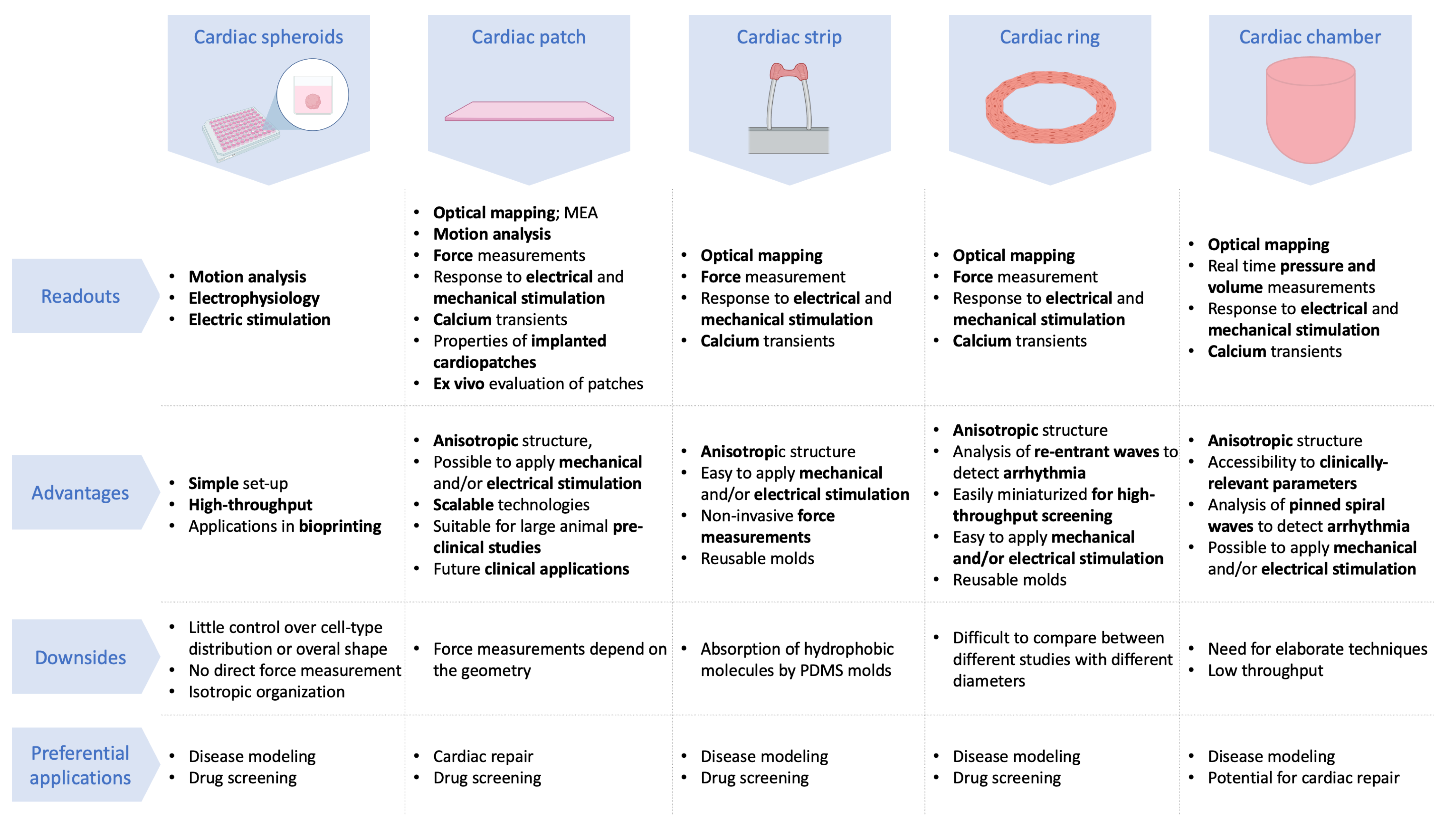
4. Different Geometries for Different Readouts and Applications
4.1. Cardiac Spheroids
4.2. Cardiac Patches
4.3. Cardiac Strips
4.4. Cardiac Rings
4.5. Cardiac Chamber
5. Limitations and Perspectives of Cardiac Organoids
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 2D | Two-dimensional |
| 3D | Three-dimensional |
| ARVC | Arrhythmogenic right ventricular cardiomyopathy |
| CF | Cardiac fibroblast |
| CM | Cardiomyocyte |
| DCM | Dilated cardiomyopathy |
| dECM | Decellularized extracellular matrix |
| DVS | Dextran vinyl sulfone |
| ECT | Engineered cardiac tissue |
| EC | Endothelial cell |
| EHT | Engineered heart tissue |
| FFR | Force–frequency relationship |
| HA | Hyaluronic acid |
| HCM | Hypertrophic cardiomyopathy |
| HUVEC | Human umbilical vein endothelial cells |
| hESCs | Human embryonic stem cells |
| hiPSC | Human-induced pluripotent stem cell |
| hPSC | Human pluripotent stem cells |
| -CM | Cardiomyocyte derived from |
| MBCD | Methyl-beta cyclodextrin |
| NHDF | Native human dermal fibroblast |
| PEG | Poly(ethyleneglycol) |
| PDMS | Polydimethylsiloxane |
| PHEMA | Poly(2-hydroxyethyl methacrylate) |
| PVA | Poly(vinyl alcohol) |
| RCM | Restrictive cardiomyopathy |
References
- Cahill, T.J.; Ashrafian, H.; Watkins, H. Genetic cardiomyopathies causing heart failure. Circ. Res. 2013, 113, 660–675. [Google Scholar] [CrossRef]
- Elliott, P.; Andersson, B.; Arbustini, E.; Bilinska, Z.; Cecchi, F.; Charron, P.; Dubourg, O.; Kühl, U.; Maisch, B.; McKenna, W.J.; et al. Classification of the cardiomyopathies: A position statement from the European Society of Cardiology Working Group on Myocardial and Pericardial Diseases. Eur. Heart J. 2008, 29, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Franz, W.M.; Müller, O.J.; Katus, H.A. Cardiomyopathies: From genetics to the prospect of treatment. Lancet 2001, 358, 1627–1637. [Google Scholar] [CrossRef]
- Kimura, A. Molecular genetics and pathogenesis of cardiomyopathy. J. Hum. Genet. 2016, 61, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Chien, K.R. Genotype, phenotype: Upstairs, downstairs in the family of cardiomyopathies. J. Clin. Investig. 2003, 111, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Kayvanpour, E.; Sedaghat-Hamedani, F.; Amr, A.; Lai, A.; Haas, J.; Holzer, D.B.; Frese, K.S.; Keller, A.; Jensen, K.; Katus, H.A.; et al. Genotype-phenotype associations in dilated cardiomyopathy: Meta-analysis on more than 8000 individuals. Clin. Res. Cardiol. 2017, 106, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Akinrinade, O.; Ollila, L.; Vattulainen, S.; Tallila, J.; Gentile, M.; Salmenperä, P.; Koillinen, H.; Kaartinen, M.; Nieminen, M.S.; Myllykangas, S.; et al. Genetics and genotype–phenotype correlations in Finnish patients with dilated cardiomyopathy. Eur. Heart J. 2015, 36, 2327–2337. [Google Scholar] [CrossRef]
- Lopes, L.R.; Rahman, M.S.; Elliott, P.M. A systematic review and meta-analysis of genotype–phenotype associations in patients with hypertrophic cardiomyopathy caused by sarcomeric protein mutations. Heart 2013, 99, 1800–1811. [Google Scholar] [CrossRef] [PubMed]
- Olivotto, I.; Oreziak, A.; Barriales-Villa, R.; Abraham, T.P.; Masri, A.; Garcia-Pavia, P.; Saberi, S.; Lakdawala, N.K.; Wheeler, M.T.; Owens, A.; et al. Mavacamten for treatment of symptomatic obstructive hypertrophic cardiomyopathy (EXPLORER-HCM): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2020, 396, 759–769. [Google Scholar] [CrossRef]
- Teerlink, J.R.; Diaz, R.; Felker, G.M.; McMurray, J.J.; Metra, M.; Solomon, S.D.; Adams, K.F.; Anand, I.; Arias-Mendoza, A.; Biering-Sørensen, T.; et al. Cardiac myosin activation with omecamtiv mecarbil in systolic heart failure. N. Engl. J. Med. 2021, 384, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Vermersch, E.; Jouve, C.; Hulot, J.S. CRISPR/Cas9 gene-editing strategies in cardiovascular cells. Cardiovasc. Res. 2020, 116, 894–907. [Google Scholar] [CrossRef]
- Cannatà, A.; Ali, H.; Sinagra, G.; Giacca, M. Gene therapy for the heart lessons learned and future perspectives. Circ. Res. 2020, 126, 1394–1414. [Google Scholar] [CrossRef]
- Purevjav, E. Animal Models of Cardiomyopathies. In Animal Models in Medicine and Biology; IntechOpen: London, UK, 2019; p. 1. [Google Scholar]
- Jimenez-Tellez, N.; Greenway, S.C. Cellular models for human cardiomyopathy: What is the best option? World J. Cardiol. 2019, 11, 221. [Google Scholar] [CrossRef] [PubMed]
- Kreutzer, F.P.; Meinecke, A.; Schmidt, K.; Fiedler, J.; Thum, T. Alternative strategies in cardiac preclinical research and new clinical trial formats. Cardiovasc. Res. 2021, cvab075. [Google Scholar] [CrossRef] [PubMed]
- Eschenhagen, T.; Carrier, L. Cardiomyopathy phenotypes in human-induced pluripotent stem cell-derived cardiomyocytes—A systematic review. Pflügers Arch. Eur. J. Physiol. 2019, 471, 755–768. [Google Scholar] [CrossRef] [Green Version]
- Mummery, C.L. Perspectives on the use of human induced pluripotent stem cell-derived cardiomyocytes in biomedical research. Stem Cell Rep. 2018, 11, 1306–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feric, N.T.; Radisic, M. Maturing human pluripotent stem cell-derived cardiomyocytes in human engineered cardiac tissues. Adv. Drug Deliv. Rev. 2016, 96, 110–134. [Google Scholar] [CrossRef] [Green Version]
- De Lange, W.J.; Farrell, E.T.; Kreitzer, C.R.; Jacobs, D.R.; Lang, D.; Glukhov, A.V.; Ralphe, J.C. Human iPSC-engineered cardiac tissue platform faithfully models important cardiac physiology. Am. J. Physiol. Heart Circ. Physiol. 2021, 320, H1670–H1686. [Google Scholar] [CrossRef]
- Chachques, J.C.; Trainini, J.C.; Lago, N.; Masoli, O.H.; Barisani, J.L.; Cortes-Morichetti, M.; Schussler, O.; Carpentier, A. Myocardial assistance by grafting a new bioartificial upgraded myocardium (MAGNUM clinical trial): One year follow-up. Cell Transplant. 2007, 16, 927–934. [Google Scholar] [CrossRef]
- Maron, B.J.; Towbin, J.A.; Thiene, G.; Antzelevitch, C.; Corrado, D.; Arnett, D.; Moss, A.J.; Seidman, C.E.; Young, J.B. Contemporary definitions and classification of the cardiomyopathies: An American Heart Association scientific statement from the council on clinical cardiology, heart failure and transplantation committee; quality of care and outcomes research and functional genomics and translational biology interdisciplinary working groups; and council on epidemiology and prevention. Circulation 2006, 113, 1807–1816. [Google Scholar]
- Miles, C.; Fanton, Z.; Tome, M.; Behr, E.R. Inherited cardiomyopathies. BMJ 2019, 365, l1570. [Google Scholar] [CrossRef] [Green Version]
- Sen-Chowdhry, S.; Jacoby, D.; Moon, J.C.; McKenna, W.J. Update on hypertrophic cardiomyopathy and a guide to the guidelines. Nat. Rev. Cardiol. 2016, 13, 651. [Google Scholar] [CrossRef] [PubMed]
- Sisakian, H. Cardiomyopathies: Evolution of pathogenesis concepts and potential for new therapies. World J. Cardiol. 2014, 6, 478. [Google Scholar] [CrossRef] [PubMed]
- Teekakirikul, P.; Zhu, W.; Huang, H.C.; Fung, E. Hypertrophic Cardiomyopathy: An Overview of Genetics and Management. Biomolecules 2019, 9, 878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hershberger, R.E.; Hedges, D.J.; Morales, A. Dilated cardiomyopathy: The complexity of a diverse genetic architecture. Nat. Rev. Cardiol. 2013, 10, 531. [Google Scholar] [CrossRef] [PubMed]
- Japp, A.G.; Gulati, A.; Cook, S.A.; Cowie, M.R.; Prasad, S.K. The diagnosis and evaluation of dilated cardiomyopathy. J. Am. Coll. Cardiol. 2016, 67, 2996–3010. [Google Scholar] [CrossRef]
- Muchtar, E.; Blauwet, L.A.; Gertz, M.A. Restrictive cardiomyopathy: Genetics, pathogenesis, clinical manifestations, diagnosis, and therapy. Circ. Res. 2017, 121, 819–837. [Google Scholar] [CrossRef]
- Corrado, D.; Link, M.S.; Calkins, H. Arrhythmogenic right ventricular cardiomyopathy. N. Engl. J. Med. 2017, 376, 61–72. [Google Scholar] [CrossRef]
- Hossain, M.B.; Bohlooly-Y, M.; Knöll, R. Mouse Models to Study Inherited Cardiomyopathy. In Genetic Causes of Cardiac Disease; Springer: Berlin/Heidelberg, Germany, 2019; pp. 289–312. [Google Scholar]
- Irfan Maqsood, M.; Matin, M.M.; Bahrami, A.R.; Ghasroldasht, M.M. Immortality of cell lines: Challenges and advantages of establishment. Cell Biol. Int. 2013, 37, 1038–1045. [Google Scholar] [CrossRef]
- Laflamme, M.A.; Chen, K.Y.; Naumova, A.V.; Muskheli, V.; Fugate, J.A.; Dupras, S.K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Police, S.; et al. Cardiomyocytes derived from human embryonic stem cells in pro-survival factors enhance function of infarcted rat hearts. Nat. Biotechnol. 2007, 25, 1015–1024. [Google Scholar] [CrossRef]
- Bai, Q.; Desprat, R.; Klein, B.; Lemaitre, J.M.; De Vos, J. Embryonic stem cells or induced pluripotent stem cells? A DNA integrity perspective. Curr. Gene Ther. 2013, 13, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Musunuru, K.; Sheikh, F.; Gupta, R.M.; Houser, S.R.; Maher, K.O.; Milan, D.J.; Terzic, A.; Wu, J.C. Induced pluripotent stem cells for cardiovascular disease modeling and precision medicine: A scientific statement from the American Heart Association. Circ. Genom. Precis. Med. 2018, 11, e000043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodehl, A.; Ebbinghaus, H.; Deutsch, M.A.; Gummert, J.; Gärtner, A.; Ratnavadivel, S.; Milting, H. Human induced pluripotent stem-cell-derived cardiomyocytes as models for genetic cardiomyopathies. Int. J. Mol. Sci. 2019, 20, 4381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, N.; Liu, C.; Wu, J.C. Translation of human-induced pluripotent stem cells: From clinical trial in a dish to precision medicine. J. Am. Coll. Cardiol. 2016, 67, 2161–2176. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Lau, Y.M.; Cai, Z.J.; Lai, W.H.; Wong, L.Y.; Tse, H.F.; Ng, K.M.; Siu, C.W. Modeling treatment response for lamin A/C related dilated cardiomyopathy in human induced pluripotent stem cells. J. Am. Heart Assoc. 2017, 6, e005677. [Google Scholar] [CrossRef]
- Sewanan, L.R.; Campbell, S.G. Modelling sarcomeric cardiomyopathies with human cardiomyocytes derived from induced pluripotent stem cells. J. Physiol. 2020, 598, 2909–2922. [Google Scholar] [CrossRef] [Green Version]
- Elitt, M.S.; Barbar, L.; Tesar, P.J. Drug screening for human genetic diseases using iPSC models. Hum. Mol. Genet. 2018, 27, R89–R98. [Google Scholar] [CrossRef] [Green Version]
- Farkhondeh, A.; Li, R.; Gorshkov, K.; Chen, K.G.; Might, M.; Rodems, S.; Lo, D.C.; Zheng, W. Induced pluripotent stem cells for neural drug discovery. Drug Discov. Today 2019, 24, 992–999. [Google Scholar] [CrossRef]
- Colatsky, T.; Fermini, B.; Gintant, G.; Pierson, J.B.; Sager, P.; Sekino, Y.; Strauss, D.G.; Stockbridge, N. The comprehensive in vitro proarrhythmia assay (CiPA) initiative—Update on progress. J. Pharmacol. Toxicol. Methods 2016, 81, 15–20. [Google Scholar] [CrossRef] [Green Version]
- Gintant, G.; Burridge, P.; Gepstein, L.; Harding, S.; Herron, T.; Hong, C.; Jalife, J.; Wu, J.C.; American Heart Association Council on Basic Cardiovascular Sciences. Use of Human Induced Pluripotent Stem Cell–Derived Cardiomyocytes in Preclinical Cancer Drug Cardiotoxicity Testing: A Scientific Statement From the American Heart Association. Circ. Res. 2019, 125, e75–e92. [Google Scholar] [CrossRef]
- Sachinidis, A. Cardiotoxicity and heart failure: Lessons from human-induced pluripotent stem cell-derived cardiomyocytes and anticancer drugs. Cells 2020, 9, 1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.S.; Macadangdang, J.; Leung, W.; Laflamme, M.A.; Kim, D.H. Human iPSC-derived cardiomyocytes and tissue engineering strategies for disease modeling and drug screening. Biotechnol. Adv. 2017, 35, 77–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronaldson-Bouchard, K.; Ma, S.P.; Yeager, K.; Chen, T.; Song, L.; Sirabella, D.; Morikawa, K.; Teles, D.; Yazawa, M.; Vunjak-Novakovic, G. Advanced maturation of human cardiac tissue grown from pluripotent stem cells. Nature 2018, 556, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, Z.; Liu, Y.; Guo, C.; Gong, Y.; Yang, S.; Ma, M.; Li, Z.; Gao, W.Q.; He, Z. Generation, characterization, and potential therapeutic applications of cardiomyocytes from various stem cells. Stem Cells Dev. 2012, 21, 2095–2110. [Google Scholar] [CrossRef]
- Liau, B.; Zhang, D.; Bursac, N. Functional cardiac tissue engineering. Regen. Med. 2012, 7, 187–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, A.R.; Ilinykh, A.; Ivey, M.J.; Kuwabara, J.T.; D’antoni, M.L.; Debuque, R.; Chandran, A.; Wang, L.; Arora, K.; Rosenthal, N.A.; et al. Revisiting cardiac cellular composition. Circ. Res. 2016, 118, 400–409. [Google Scholar] [CrossRef] [Green Version]
- Devalla, H.D.; Passier, R. Cardiac differentiation of pluripotent stem cells and implications for modeling the heart in health and disease. Sci. Transl. Med. 2018, 10, eaah5457. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, H.; Wei, R.; Ma, J.; Zhao, Y.; Lian, Z.; Liu, Z. Endothelial cells regulate cardiac myocyte reorganisation through β1-integrin signalling. Cell. Physiol. Biochem. 2015, 35, 1808–1820. [Google Scholar] [CrossRef]
- Michel, J.B.; Li, Z.; Lacolley, P. Smooth muscle cells and vascular diseases. Cardiovasc. Res. 2012, 95, 135–137. [Google Scholar] [CrossRef] [Green Version]
- Baudino, T.A.; Carver, W.; Giles, W.; Borg, T.K. Cardiac fibroblasts: Friend or foe? Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1015–H1026. [Google Scholar] [CrossRef]
- Ivey, M.J.; Tallquist, M.D. Defining the cardiac fibroblast. Circ. J. 2016, 80, CJ-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achanta, S.; Gorky, J.; Leung, C.; Moss, A.; Robbins, S.; Eisenman, L.; Chen, J.; Tappan, S.; Heal, M.; Farahani, N.; et al. A comprehensive integrated anatomical and molecular atlas of rat intrinsic cardiac nervous system. Iscience 2020, 23, 101140. [Google Scholar] [CrossRef]
- Nahrendorf, M. Myeloid cells in cardiovascular organs. J. Intern. Med. 2019, 285, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, X.; Xia, L.; Su, Z. Friend or foe of innate lymphoid cells in inflammation-associated cardiovascular disease. Immunology 2020, 162, 368–376. [Google Scholar] [CrossRef]
- Yaniz-Galende, E.; Roux, M.; Nadaud, S.; Mougenot, N.; Bouvet, M.; Claude, O.; Lebreton, G.; Blanc, C.; Pinet, F.; Atassi, F.; et al. Fibrogenic potential of PW1/Peg3 expressing cardiac stem cells. J. Am. Coll. Cardiol. 2017, 70, 728–741. [Google Scholar] [CrossRef] [PubMed]
- Litviňuková, M.; Talavera-López, C.; Maatz, H.; Reichart, D.; Worth, C.L.; Lindberg, E.L.; Kanda, M.; Polanski, K.; Heinig, M.; Lee, M.; et al. Cells of the adult human heart. Nature 2020, 588, 466–472. [Google Scholar] [CrossRef]
- Schwach, V.; Passier, R. Native cardiac environment and its impact on engineering cardiac tissue. Biomater. Sci. 2019, 7, 3566–3580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rienks, M.; Papageorgiou, A.P.; Frangogiannis, N.G.; Heymans, S. Myocardial extracellular matrix: An ever-changing and diverse entity. Circ. Res. 2014, 114, 872–888. [Google Scholar] [CrossRef] [Green Version]
- Ariyasinghe, N.R.; Lyra-Leite, D.M.; McCain, M.L. Engineering cardiac microphysiological systems to model pathological extracellular matrix remodeling. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H771–H789. [Google Scholar] [CrossRef]
- Hasenfuss, G.; Mulieri, L.A.; Blanchard, E.M.; Holubarsch, C.; Leavitt, B.; Ittleman, F.; Alpert, N. Energetics of isometric force development in control and volume-overload human myocardium. Comparison with animal species. Circ. Res. 1991, 68, 836–846. [Google Scholar] [CrossRef] [Green Version]
- Klabunde, R. Cardiovascular Physiology Concepts; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2011. [Google Scholar]
- Janssen, P.M. Myocardial contraction-relaxation coupling. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H1741–H1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cain, B.S.; Meldrum, D.R.; Meng, X.; Shames, B.D.; Harken, A.H. Calcium preconditioning in human myocardium. Ann. Thorac. Surg. 1998, 65, 1065–1070. [Google Scholar] [CrossRef]
- Kitazawa, T. Effect of extracellular calcium on contractile activation in guinea-pig ventricular muscle. J. Physiol. 1984, 355, 635–659. [Google Scholar] [CrossRef]
- Weinberger, F.; Mannhardt, I.; Eschenhagen, T. Engineering cardiac muscle tissue: A maturating field of research. Circ. Res. 2017, 120, 1487–1500. [Google Scholar] [CrossRef] [PubMed]
- Karakikes, I.; Senyei, G.D.; Hansen, J.; Kong, C.W.; Azeloglu, E.U.; Stillitano, F.; Lieu, D.K.; Wang, J.; Ren, L.; Hulot, J.S.; et al. Small molecule-mediated directed differentiation of human embryonic stem cells toward ventricular cardiomyocytes. Stem Cells Transl. Med. 2014, 3, 18–31. [Google Scholar] [CrossRef]
- Lian, X.; Zhang, J.; Azarin, S.M.; Zhu, K.; Hazeltine, L.B.; Bao, X.; Hsiao, C.; Kamp, T.J.; Palecek, S.P. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/β-catenin signaling under fully defined conditions. Nat. Protoc. 2013, 8, 162–175. [Google Scholar] [CrossRef] [Green Version]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Batalov, I.; Feinberg, A.W. Differentiation of cardiomyocytes from human pluripotent stem cells using monolayer culture: Supplementary issue: Stem cell biology. Biomark. Insights 2015, 10, BMI-S20050. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Rafatian, N.; Feric, N.T.; Cox, B.J.; Aschar-Sobbi, R.; Wang, E.Y.; Aggarwal, P.; Zhang, B.; Conant, G.; Ronaldson-Bouchard, K.; et al. A platform for generation of chamber-specific cardiac tissues and disease modeling. Cell 2019, 176, 913–927. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, R.; Nakayama, K.; Itoh, M.; Kamohara, K.; Furukawa, K.; Oyama, J.i.; Node, K.; Morita, S. Development of a three-dimensional pre-vascularized scaffold-free contractile cardiac patch for treating heart disease. J. Heart Lung Transplant. 2016, 35, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Pitaktong, I.; Lui, C.; Lowenthal, J.; Mattson, G.; Jung, W.H.; Bai, Y.; Yeung, E.; Ong, C.S.; Chen, Y.; Gerecht, S.; et al. Early vascular cells improve microvascularization within 3D cardiac spheroids. Tissue Eng. Part C Methods 2020, 26, 80–90. [Google Scholar] [CrossRef]
- Saini, H.; Navaei, A.; Van Putten, A.; Nikkhah, M. 3D cardiac microtissues encapsulated with the co-culture of cardiomyocytes and cardiac fibroblasts. Adv. Healthc. Mater. 2015, 4, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Giacomelli, E.; Meraviglia, V.; Campostrini, G.; Cochrane, A.; Cao, X.; van Helden, R.W.; Garcia, A.K.; Mircea, M.; Kostidis, S.; Davis, R.P.; et al. Human-iPSC-derived cardiac stromal cells enhance maturation in 3D cardiac microtissues and reveal non-cardiomyocyte contributions to heart disease. Cell Stem Cell 2020, 26, 862–879. [Google Scholar] [CrossRef]
- Caspi, O.; Lesman, A.; Basevitch, Y.; Gepstein, A.; Arbel, G.; Habib, I.H.M.; Gepstein, L.; Levenberg, S. Tissue engineering of vascularized cardiac muscle from human embryonic stem cells. Circ. Res. 2007, 100, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Amano, Y.; Nishiguchi, A.; Matsusaki, M.; Iseoka, H.; Miyagawa, S.; Sawa, Y.; Seo, M.; Yamaguchi, T.; Akashi, M. Development of vascularized iPSC derived 3D-cardiomyocyte tissues by filtration Layer-by-Layer technique and their application for pharmaceutical assays. Acta Biomater. 2016, 33, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Stevens, K.; Kreutziger, K.; Dupras, S.; Korte, F.; Regnier, M.; Muskheli, V.; Nourse, M.; Bendixen, K.; Reinecke, H.; Murry, C. Physiological function and transplantation of scaffold-free and vascularized human cardiac muscle tissue. Proc. Natl. Acad. Sci. USA 2009, 106, 16568–16573. [Google Scholar] [CrossRef] [Green Version]
- Tulloch, N.L.; Muskheli, V.; Razumova, M.V.; Korte, F.S.; Regnier, M.; Hauch, K.D.; Pabon, L.; Reinecke, H.; Murry, C.E. Growth of engineered human myocardium with mechanical loading and vascular coculture. Circ. Res. 2011, 109, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Fujita, B.; Zimmermann, W.H. Myocardial tissue engineering strategies for heart repair: Current state of the art. Interact. Cardiovasc. Thorac. Surg. 2018, 27, 916–920. [Google Scholar] [CrossRef]
- Filippo Buono, M.; von Boehmer, L.; Strang, J.; Hoerstrup, S.P.; Emmert, M.Y.; Nugraha, B. Human Cardiac Organoids for Modeling Genetic Cardiomyopathy. Cells 2020, 9, 1733. [Google Scholar] [CrossRef]
- Shum, A.M.; Che, H.; Wong, A.O.T.; Zhang, C.; Wu, H.; Chan, C.W.; Costa, K.; Khine, M.; Kong, C.W.; Li, R.A. A micropatterned human pluripotent stem cell-based ventricular cardiac anisotropic sheet for visualizing drug-induced arrhythmogenicity. Adv. Mater. 2017, 29, 1602448. [Google Scholar] [CrossRef]
- Zuppinger, C. 3D culture for cardiac cells. Biochim. Biophys. Acta BBA Mol. Cell Res. 2016, 1863, 1873–1881. [Google Scholar] [CrossRef] [PubMed]
- Eschenhagen, T.; Zimmermann, W.H. Engineering myocardial tissue. Circ. Res. 2005, 97, 1220–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauchamp, P.; Moritz, W.; Kelm, J.M.; Ullrich, N.D.; Agarkova, I.; Anson, B.D.; Suter, T.M.; Zuppinger, C. Development and characterization of a scaffold-free 3D spheroid model of induced pluripotent stem cell-derived human cardiomyocytes. Tissue Eng. Part C Methods 2015, 21, 852–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polonchuk, L.; Chabria, M.; Badi, L.; Hoflack, J.C.; Figtree, G.; Davies, M.J.; Gentile, C. Cardiac spheroids as promising in vitro models to study the human heart microenvironment. Sci. Rep. 2017, 7, 7005. [Google Scholar] [CrossRef]
- Daly, A.C.; Davidson, M.D.; Burdick, J.A. 3D bioprinting of high cell-density heterogeneous tissue models through spheroid fusion within self-healing hydrogels. Nat. Commun. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Miwa, T.; Idiris, A.; Kumagai, H. A novel cardiac differentiation method of a large number and uniformly-sized spheroids using microfabricated culture vessels. Regen. Ther. 2020, 15, 18–26. [Google Scholar] [CrossRef]
- Zuppinger, C. Measurement of contractility and calcium release in cardiac spheroids. In Calcium-Binding Proteins of the EF-Hand Superfamily; Springer: Berlin/Heidelberg, Germany, 2019; pp. 41–52. [Google Scholar]
- Beauchamp, P.; Jackson, C.B.; Ozhathil, L.C.; Agarkova, I.; Galindo, C.L.; Sawyer, D.B.; Suter, T.M.; Zuppinger, C. 3D co-culture of hiPSC-derived cardiomyocytes with cardiac fibroblasts improves tissue-like features of cardiac spheroids. Front. Mol. Biosci. 2020, 7, 14. [Google Scholar] [CrossRef]
- Yan, Y.; Bejoy, J.; Xia, J.; Griffin, K.; Guan, J.; Li, Y. Cell population balance of cardiovascular spheroids derived from human induced pluripotent stem cells. Sci. Rep. 2019, 9, 1295. [Google Scholar] [CrossRef]
- Giacomelli, E.; Bellin, M.; Sala, L.; Van Meer, B.J.; Tertoolen, L.G.; Orlova, V.V.; Mummery, C.L. Three-dimensional cardiac microtissues composed of cardiomyocytes and endothelial cells co-differentiated from human pluripotent stem cells. Development 2017, 144, 1008–1017. [Google Scholar] [CrossRef] [Green Version]
- Arai, K.; Murata, D.; Verissimo, A.R.; Mukae, Y.; Itoh, M.; Nakamura, A.; Morita, S.; Nakayama, K. Fabrication of scaffold-free tubular cardiac constructs using a Bio-3D printer. PLoS ONE 2018, 13, e0209162. [Google Scholar] [CrossRef] [Green Version]
- Ong, C.S.; Pitaktong, I.; Hibino, N. Principles of Spheroid Preparation for Creation of 3D Cardiac Tissue Using Biomaterial-Free Bioprinting. In 3D Bioprinting; Springer: Berlin/Heidelberg, Germany, 2020; pp. 183–197. [Google Scholar]
- Moldovan, N.I. Progress in scaffold-free bioprinting for cardiovascular medicine. J. Cell. Mol. Med. 2018, 22, 2964–2969. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Yamato, M.; Isoi, Y.; Akutsu, T.; Setomaru, T.; Abe, K.; Kikuchi, A.; Umezu, M.; Okano, T. Fabrication of pulsatile cardiac tissue grafts using a novel 3-dimensional cell sheet manipulation technique and temperature-responsive cell culture surfaces. Circ. Res. 2002, 90, e40–e48. [Google Scholar] [CrossRef] [Green Version]
- Sekine, H.; Shimizu, T.; Hobo, K.; Sekiya, S.; Yang, J.; Yamato, M.; Kurosawa, H.; Kobayashi, E.; Okano, T. Endothelial cell coculture within tissue-engineered cardiomyocyte sheets enhances neovascularization and improves cardiac function of ischemic hearts. Circulation 2008, 118, S145–S152. [Google Scholar] [CrossRef] [Green Version]
- Jiao, A.; Trosper, N.E.; Yang, H.S.; Kim, J.; Tsui, J.H.; Frankel, S.D.; Murry, C.E.; Kim, D.H. Thermoresponsive nanofabricated substratum for the engineering of three-dimensional tissues with layer-by-layer architectural control. ACS Nano 2014, 8, 4430–4439. [Google Scholar] [CrossRef] [PubMed]
- Rogozhnikov, D.; O’Brien, P.J.; Elahipanah, S.; Yousaf, M.N. Scaffold free bio-orthogonal assembly of 3-dimensional cardiac tissue via cell surface engineering. Sci. Rep. 2016, 6, 39806. [Google Scholar] [CrossRef] [Green Version]
- Capulli, A.; MacQueen, L.; Sheehy, S.P.; Parker, K. Fibrous scaffolds for building hearts and heart parts. Adv. Drug Deliv. Rev. 2016, 96, 83–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Ferguson, M.; Kamp, T.J.; Zhao, F. Constructing biomimetic cardiac tissues: A review of scaffold materials for engineering cardiac patches. Emergent Mater. 2019, 2, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, N.J.; Coulombe, K.L. Physiologically inspired cardiac scaffolds for tailored in vivo function and heart regeneration. Biomed. Mater. 2015, 10, 034003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouten, C.; Dankers, P.; Driessen-Mol, A.; Pedron, S.; Brizard, A.; Baaijens, F. Substrates for cardiovascular tissue engineering. Adv. Drug Deliv. Rev. 2011, 63, 221–241. [Google Scholar] [CrossRef]
- Bejleri, D.; Davis, M.E. Decellularized extracellular matrix materials for cardiac repair and regeneration. Adv. Healthc. Mater. 2019, 8, 1801217. [Google Scholar] [CrossRef]
- Guyette, J.P.; Charest, J.M.; Mills, R.W.; Jank, B.J.; Moser, P.T.; Gilpin, S.E.; Gershlak, J.R.; Okamoto, T.; Gonzalez, G.; Milan, D.J.; et al. Bioengineering human myocardium on native extracellular matrix. Circ. Res. 2016, 118, 56–72. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.S.; Kim, H.; Gao, G.; Jang, J.; Cho, D.W. Decellularized extracellular matrix: A step towards the next generation source for bioink manufacturing. Biofabrication 2017, 9, 034104. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, H.; Bai, A.; Jiang, W.; Li, X.; Wang, X.; Mao, Y.; Lu, C.; Qian, R.; Guo, F.; et al. Functional engineered human cardiac patches prepared from nature’s platform improve heart function after acute myocardial infarction. Biomaterials 2016, 105, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Ott, H.C.; Matthiesen, T.S.; Goh, S.K.; Black, L.D.; Kren, S.M.; Netoff, T.I.; Taylor, D.A. Perfusion-decellularized matrix: Using nature’s platform to engineer a bioartificial heart. Nat. Med. 2008, 14, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Mendibil, U.; Ruiz-Hernandez, R.; Retegi-Carrion, S.; Garcia-Urquia, N.; Olalde-Graells, B.; Abarrategi, A. Tissue-specific decellularization methods: Rationale and strategies to achieve regenerative compounds. Int. J. Mol. Sci. 2020, 21, 5447. [Google Scholar] [CrossRef]
- Saldin, L.T.; Cramer, M.C.; Velankar, S.S.; White, L.J.; Badylak, S.F. Extracellular matrix hydrogels from decellularized tissues: Structure and function. Acta Biomater. 2017, 49, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saludas, L.; Pascual-Gil, S.; Prósper, F.; Garbayo, E.; Blanco-Prieto, M. Hydrogel based approaches for cardiac tissue engineering. Int. J. Pharm. 2017, 523, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Marchant, R.E. Design properties of hydrogel tissue-engineering scaffolds. Expert Rev. Med. Devices 2011, 8, 607–626. [Google Scholar] [CrossRef]
- Unal, A.Z.; West, J.L. Synthetic ECM: Bioactive synthetic hydrogels for 3D tissue engineering. Bioconjug. Chem. 2020, 31, 2253–2271. [Google Scholar] [CrossRef]
- Tallawi, M.; Rosellini, E.; Barbani, N.; Cascone, M.G.; Rai, R.; Saint-Pierre, G.; Boccaccini, A.R. Strategies for the chemical and biological functionalization of scaffolds for cardiac tissue engineering: A review. J. R. Soc. Interface 2015, 12, 20150254. [Google Scholar] [CrossRef]
- Hendrickson, T.; Mancino, C.; Whitney, L.; Tsao, C.; Rahimi, M.; Taraballi, F. Mimicking cardiac tissue complexity through physical cues: A review on cardiac tissue engineering approaches. Nanomed. Nanotechnol. Biol. Med. 2021, 33, 102367. [Google Scholar] [CrossRef]
- Turnbull, I.C.; Karakikes, I.; Serrao, G.W.; Backeris, P.; Lee, J.J.; Xie, C.; Senyei, G.; Gordon, R.E.; Li, R.A.; Akar, F.G.; et al. Advancing functional engineered cardiac tissues toward a preclinical model of human myocardium. FASEB J. 2014, 28, 644–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihic, A.; Li, J.; Miyagi, Y.; Gagliardi, M.; Li, S.H.; Zu, J.; Weisel, R.D.; Keller, G.; Li, R.K. The effect of cyclic stretch on maturation and 3D tissue formation of human embryonic stem cell-derived cardiomyocytes. Biomaterials 2014, 35, 2798–2808. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.S.; Miklas, J.W.; Liu, J.; Aschar-Sobbi, R.; Xiao, Y.; Zhang, B.; Jiang, J.; Massé, S.; Gagliardi, M.; Hsieh, A.; et al. Biowire: A platform for maturation of human pluripotent stem cell–derived cardiomyocytes. Nat. Methods 2013, 10, 781–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirt, M.N.; Boeddinghaus, J.; Mitchell, A.; Schaaf, S.; Börnchen, C.; Müller, C.; Schulz, H.; Hubner, N.; Stenzig, J.; Stoehr, A.; et al. Functional improvement and maturation of rat and human engineered heart tissue by chronic electrical stimulation. J. Mol. Cell. Cardiol. 2014, 74, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, J.L.; Tulloch, N.L.; Razumova, M.V.; Saiget, M.; Muskheli, V.; Pabon, L.; Reinecke, H.; Regnier, M.; Murry, C.E. Mechanical stress conditioning and electrical stimulation promote contractility and force maturation of induced pluripotent stem cell-derived human cardiac tissue. Circulation 2016, 134, 1557–1567. [Google Scholar] [CrossRef]
- Wang, B.; Wang, G.; To, F.; Butler, J.R.; Claude, A.; McLaughlin, R.M.; Williams, L.N.; de Jongh Curry, A.L.; Liao, J. Myocardial scaffold-based cardiac tissue engineering: Application of coordinated mechanical and electrical stimulations. Langmuir 2013, 29, 11109–11117. [Google Scholar] [CrossRef]
- Morgan, K.Y.; Black, L.D., III. Mimicking isovolumic contraction with combined electromechanical stimulation improves the development of engineered cardiac constructs. Tissue Eng. Part A 2014, 20, 1654–1667. [Google Scholar] [CrossRef] [Green Version]
- Kitsara, M.; Kontziampasis, D.; Agbulut, O.; Chen, Y. Heart on a chip: Micro-nanofabrication and microfluidics steering the future of cardiac tissue engineering. Microelectron. Eng. 2019, 203, 44–62. [Google Scholar] [CrossRef]
- Boffito, M.; Sartori, S.; Ciardelli, G. Polymeric scaffolds for cardiac tissue engineering: Requirements and fabrication technologies. Polym. Int. 2014, 63, 2–11. [Google Scholar] [CrossRef]
- Doshi, J.; Reneker, D.H. Electrospinning process and applications of electrospun fibers. J. Electrost. 1995, 35, 151–160. [Google Scholar] [CrossRef]
- Muniyandi, P.; Palaninathan, V.; Veeranarayanan, S.; Ukai, T.; Maekawa, T.; Hanajiri, T.; Mohamed, M.S. ECM mimetic electrospun porous poly (l-lactic acid)(PLLA) scaffolds as potential substrates for cardiac tissue engineering. Polymers 2020, 12, 451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DePalma, S.J.; Davidson, C.D.; Stis, A.E.; Helms, A.S.; Baker, B.M. Microenvironmental determinants of organized iPSC-cardiomyocyte tissues on synthetic fibrous matrices. Biomater. Sci. 2021, 9, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Suh, T.C.; Amanah, A.Y.; Gluck, J.M. Electrospun Scaffolds and Induced Pluripotent Stem Cell-Derived Cardiomyocytes for Cardiac Tissue Engineering Applications. Bioengineering 2020, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Persano, L.; Camposeo, A.; Tekmen, C.; Pisignano, D. Industrial upscaling of electrospinning and applications of polymer nanofibers: A review. Macromol. Mater. Eng. 2013, 298, 504–520. [Google Scholar] [CrossRef]
- Liu, N.; Ye, X.; Yao, B.; Zhao, M.; Wu, P.; Liu, G.; Zhuang, D.; Jiang, H.; Chen, X.; He, Y.; et al. Advances in 3D bioprinting technology for cardiac tissue engineering and regeneration. Bioact. Mater. 2021, 6, 1388–1401. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Hudson, A.; Shiwarski, D.; Tashman, J.; Hinton, T.; Yerneni, S.; Bliley, J.; Campbell, P.; Feinberg, A. 3D bioprinting of collagen to rebuild components of the human heart. Science 2019, 365, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Fournier, N.; Doillon, C.J. Biological molecule-impregnated polyester: An in vivo angiogenesis study. Biomaterials 1996, 17, 1659–1665. [Google Scholar] [CrossRef]
- Perets, A.; Baruch, Y.; Weisbuch, F.; Shoshany, G.; Neufeld, G.; Cohen, S. Enhancing the vascularization of three-dimensional porous alginate scaffolds by incorporating controlled release basic fibroblast growth factor microspheres. J. Biomed. Mater. Res. Part Off. J. Soc. Biomater. Jpn. Soc. Biomater. Aust. Soc. Biomater. Korean Soc. Biomater. 2003, 65, 489–497. [Google Scholar] [CrossRef]
- Sakaguchi, K.; Shimizu, T.; Horaguchi, S.; Sekine, H.; Yamato, M.; Umezu, M.; Okano, T. In vitro engineering of vascularized tissue surrogates. Sci. Rep. 2013, 3, 1316. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, W.H.; Didié, M.; Döker, S.; Melnychenko, I.; Naito, H.; Rogge, C.; Tiburcy, M.; Eschenhagen, T. Heart muscle engineering: An update on cardiac muscle replacement therapy. Cardiovasc. Res. 2006, 71, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bates, R.C.; Edwards, N.S.; Yates, J.D. Spheroids and cell survival. Crit. Rev. Oncol. 2000, 36, 61–74. [Google Scholar] [CrossRef]
- Zuppinger, C. 3D cardiac cell culture: A critical review of current technologies and applications. Front. Cardiovasc. Med. 2019, 6, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J. Engineered tissue patch for cardiac cell therapy. Curr. Treat. Options Cardiovasc. Med. 2015, 17, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shadrin, I.Y.; Allen, B.W.; Qian, Y.; Jackman, C.P.; Carlson, A.L.; Juhas, M.E.; Bursac, N. Cardiopatch platform enables maturation and scale-up of human pluripotent stem cell-derived engineered heart tissues. Nat. Commun. 2017, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tiburcy, M.; Hudson, J.E.; Balfanz, P.; Schlick, S.; Meyer, T.; Chang Liao, M.L.; Levent, E.; Raad, F.; Zeidler, S.; Wingender, E.; et al. Defined engineered human myocardium with advanced maturation for applications in heart failure modeling and repair. Circulation 2017, 135, 1832–1847. [Google Scholar] [CrossRef] [PubMed]
- Bian, W.; Jackman, C.P.; Bursac, N. Controlling the structural and functional anisotropy of engineered cardiac tissues. Biofabrication 2014, 6, 024109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Shadrin, I.Y.; Lam, J.; Xian, H.Q.; Snodgrass, H.R.; Bursac, N. Tissue-engineered cardiac patch for advanced functional maturation of human ESC-derived cardiomyocytes. Biomaterials 2013, 34, 5813–5820. [Google Scholar] [CrossRef] [Green Version]
- Takeda, M.; Miyagawa, S.; Ito, E.; Harada, A.; Mochizuki-Oda, N.; Matsusaki, M.; Akashi, M.; Sawa, Y. Development of a drug screening system using three-dimensional cardiac tissues containing multiple cell types. Sci. Rep. 2021, 11, 5654. [Google Scholar] [CrossRef]
- Jackman, C.P.; Ganapathi, A.M.; Asfour, H.; Qian, Y.; Allen, B.W.; Li, Y.; Bursac, N. Engineered cardiac tissue patch maintains structural and electrical properties after epicardial implantation. Biomaterials 2018, 159, 48–58. [Google Scholar] [CrossRef]
- Gao, L.; Gregorich, Z.R.; Zhu, W.; Mattapally, S.; Oduk, Y.; Lou, X.; Kannappan, R.; Borovjagin, A.V.; Walcott, G.P.; Pollard, A.E.; et al. Large cardiac muscle patches engineered from human induced-pluripotent stem cell–derived cardiac cells improve recovery from myocardial infarction in swine. Circulation 2018, 137, 1712–1730. [Google Scholar] [CrossRef]
- Feiner, R.; Engel, L.; Fleischer, S.; Malki, M.; Gal, I.; Shapira, A.; Shacham-Diamand, Y.; Dvir, T. Engineered hybrid cardiac patches with multifunctional electronics for online monitoring and regulation of tissue function. Nat. Mater. 2016, 15, 679–685. [Google Scholar] [CrossRef] [Green Version]
- Hansen, A.; Eder, A.; Bönstrup, M.; Flato, M.; Mewe, M.; Schaaf, S.; Aksehirlioglu, B.; Schwörer, A.; Uebeler, J.; Eschenhagen, T. Development of a drug screening platform based on engineered heart tissue. Circ. Res. 2010, 107, 35. [Google Scholar] [CrossRef] [Green Version]
- Feric, N.T.; Pallotta, I.; Singh, R.; Bogdanowicz, D.R.; Gustilo, M.M.; Chaudhary, K.W.; Willette, R.N.; Chendrimada, T.P.; Xu, X.; Graziano, M.P.; et al. Engineered cardiac tissues generated in the Biowire II: A platform for human-based drug discovery. Toxicol. Sci. 2019, 172, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Serrao, G.W.; Turnbull, I.C.; Ancukiewicz, D.; Kim, D.E.; Kao, E.; Cashman, T.J.; Hadri, L.; Hajjar, R.J.; Costa, K.D. Myocyte-depleted engineered cardiac tissues support therapeutic potential of mesenchymal stem cells. Tissue Eng. Part A 2012, 18, 1322–1333. [Google Scholar] [CrossRef] [Green Version]
- Kensah, G.; Gruh, I.; Viering, J.; Schumann, H.; Dahlmann, J.; Meyer, H.; Skvorc, D.; Bär, A.; Akhyari, P.; Heisterkamp, A.; et al. A novel miniaturized multimodal bioreactor for continuous in situ assessment of bioartificial cardiac tissue during stimulation and maturation. Tissue Eng. Part C Methods 2011, 17, 463–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cashman, T.J.; Josowitz, R.; Johnson, B.V.; Gelb, B.D.; Costa, K.D. Human engineered cardiac tissues created using induced pluripotent stem cells reveal functional characteristics of BRAF-mediated hypertrophic cardiomyopathy. PLoS ONE 2016, 11, e0146697. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.; Liang, T.; Massé, S.; Khan, S.; Hatkar, R.; Keller, G.; Nanthakumar, K.; Nunes, S.S. A 3-D Human Model of Complex Cardiac Arrhythmias. Acta Biomater. 2021, in press. [Google Scholar] [CrossRef]
- Mannhardt, I.; Breckwoldt, K.; Letuffe-Brenière, D.; Schaaf, S.; Schulz, H.; Neuber, C.; Benzin, A.; Werner, T.; Eder, A.; Schulze, T.; et al. Human engineered heart tissue: Analysis of contractile force. Stem Cell Rep. 2016, 7, 29–42. [Google Scholar] [CrossRef] [Green Version]
- Nattel, S. New ideas about atrial fibrillation 50 years on. Nature 2002, 415, 219–226. [Google Scholar] [CrossRef]
- Colman, M.A. Arrhythmia mechanisms and spontaneous calcium release: Bi-directional coupling between re-entrant and focal excitation. PLoS Comput. Biol. 2019, 15, e1007260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhang, L.; Yu, L.; Minami, I.; Miyagawa, S.; Hörning, M.; Dong, J.; Qiao, J.; Qu, X.; Hua, Y.; et al. Circulating re-entrant waves promote maturation of hiPSC-derived cardiomyocytes in self-organized tissue ring. Commun. Biol. 2020, 3, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Goldfracht, I.; Efraim, Y.; Shinnawi, R.; Kovalev, E.; Huber, I.; Gepstein, A.; Arbel, G.; Shaheen, N.; Tiburcy, M.; Zimmermann, W.H.; et al. Engineered heart tissue models from hiPSC-derived cardiomyocytes and cardiac ECM for disease modeling and drug testing applications. Acta Biomater. 2019, 92, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Goldfracht, I.; Protze, S.; Shiti, A.; Setter, N.; Gruber, A.; Shaheen, N.; Nartiss, Y.; Keller, G.; Gepstein, L. Generating ring-shaped engineered heart tissues from ventricular and atrial human pluripotent stem cell-derived cardiomyocytes. Nat. Commun. 2020, 11, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacQueen, L.A.; Sheehy, S.P.; Chantre, C.O.; Zimmerman, J.F.; Pasqualini, F.S.; Liu, X.; Goss, J.A.; Campbell, P.H.; Gonzalez, G.M.; Park, S.J.; et al. A tissue-engineered scale model of the heart ventricle. Nat. Biomed. Eng. 2018, 2, 930–941. [Google Scholar] [CrossRef]
- Li, R.A.; Keung, W.; Cashman, T.J.; Backeris, P.C.; Johnson, B.V.; Bardot, E.S.; Wong, A.O.; Chan, P.K.; Chan, C.W.; Costa, K.D. Bioengineering an electro-mechanically functional miniature ventricular heart chamber from human pluripotent stem cells. Biomaterials 2018, 163, 116–127. [Google Scholar] [CrossRef]
- Tzatzalos, E.; Abilez, O.J.; Shukla, P.; Wu, J.C. Engineered heart tissues and induced pluripotent stem cells: Macro-and microstructures for disease modeling, drug screening, and translational studies. Adv. Drug Deliv. Rev. 2016, 96, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, R.E.; Anzai, T.; Chanthra, N.; Uosaki, H. A brief review of current maturation methods for human induced pluripotent stem cells-derived cardiomyocytes. Front. Cell Dev. Biol. 2020, 8, 178. [Google Scholar] [CrossRef] [Green Version]
- Kolanowski, T.J.; Antos, C.L.; Guan, K. Making human cardiomyocytes up to date: Derivation, maturation state and perspectives. Int. J. Cardiol. 2017, 241, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, T.P.; Pinto, S.N.; Santos, J.I.; Fernandes, T.G.; Fernandes, F.; Diogo, M.M.; Prieto, M.; Cabral, J.M. Biophysical study of human induced Pluripotent Stem Cell-Derived cardiomyocyte structural maturation during long-term culture. Biochem. Biophys. Res. Commun. 2018, 499, 611–617. [Google Scholar] [CrossRef]
- Feyen, D.A.; McKeithan, W.L.; Bruyneel, A.A.; Spiering, S.; Hörmann, L.; Ulmer, B.; Zhang, H.; Briganti, F.; Schweizer, M.; Hegyi, B.; et al. Metabolic maturation media improve physiological function of human iPSC-derived cardiomyocytes. Cell Rep. 2020, 32, 107925. [Google Scholar] [CrossRef] [PubMed]
- Ye, L.; Zhang, X.; Zhou, Q.; Tan, B.; Xu, H.; Yi, Q.; Yan, L.; Xie, M.; Zhang, Y.; Tian, J.; et al. Activation of AMPK Promotes Maturation of Cardiomyocytes Derived From Human Induced Pluripotent Stem Cells. Front. Cell Dev. Biol. 2021, 9, 644667. [Google Scholar] [CrossRef]
- Ni, X.; Xu, K.; Zhao, Y.; Li, J.; Wang, L.; Yu, F.; Li, G. Single-cell analysis reveals the purification and maturation effects of glucose starvation in hiPSC-CMs. Biochem. Biophys. Res. Commun. 2021, 534, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Block, T.; Creech, J.; da Rocha, A.M.; Marinkovic, M.; Ponce-Balbuena, D.; Jiménez-Vázquez, E.N.; Griffey, S.; Herron, T.J. Human perinatal stem cell derived extracellular matrix enables rapid maturation of hiPSC-CM structural and functional phenotypes. Sci. Rep. 2020, 10, 19071. [Google Scholar] [CrossRef] [PubMed]
- Howe, R.; Isaacson, K.; Albert, J.; Coutifaris, C. Embryonic heart rate in human pregnancy. J. Ultrasound Med. 1991, 10, 367–371. [Google Scholar] [CrossRef]
- Crestani, T.; Steichen, C.; Neri, E.; Rodrigues, M.; Fonseca-Alaniz, M.H.; Ormrod, B.; Holt, M.R.; Pandey, P.; Harding, S.; Ehler, E.; et al. Electrical stimulation applied during differentiation drives the hiPSC-CMs towards a mature cardiac conduction-like cells. Biochem. Biophys. Res. Commun. 2020, 533, 376–382. [Google Scholar] [CrossRef] [PubMed]
- Tracy, E.P.; Gettler, B.C.; Zakhari, J.S.; Schwartz, R.J.; Williams, S.K.; Birla, R.K. 3D bioprinting the cardiac purkinje system using human adipogenic mesenchymal stem cell derived Purkinje cells. Cardiovasc. Eng. Technol. 2020, 11, 587–604. [Google Scholar] [CrossRef]
- Zhao, Y.; Rafatian, N.; Wang, E.Y.; Feric, N.T.; Lai, B.F.; Knee-Walden, E.J.; Backx, P.H.; Radisic, M. Engineering microenvironment for human cardiac tissue assembly in heart-on-a-chip platform. Matrix Biol. 2020, 85, 189–204. [Google Scholar] [CrossRef]
- Massai, D.; Pisani, G.; Isu, G.; Rodriguez Ruiz, A.; Cerino, G.; Galluzzi, R.; Pisanu, A.; Tonoli, A.; Bignardi, C.; Audenino, A.L.; et al. Bioreactor platform for biomimetic culture and in situ monitoring of the mechanical response of in vitro engineered models of cardiac tissue. Front. Bioeng. Biotechnol. 2020, 8, 733. [Google Scholar] [CrossRef]
- Pitoulis, F.G.; Nunez-Toldra, R.; Xiao, K.; Kit-Anan, W.; Mitzka, S.; Jabbour, R.J.; Harding, S.E.; Perbellini, F.; Thum, T.; de Tombe, P.P.; et al. Remodelling of adult cardiac tissue subjected to physiological and pathological mechanical load in vitro. Cardiovasc. Res. 2021, cvab084. [Google Scholar] [CrossRef]
- Mathur, A.; Loskill, P.; Shao, K.; Huebsch, N.; Hong, S.; Marcus, S.G.; Marks, N.; Mandegar, M.; Conklin, B.R.; Lee, L.P.; et al. Human iPSC-based cardiac microphysiological system for drug screening applications. Sci. Rep. 2015, 5, 8883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Wang, T.; Wang, P.; Hu, N. High-throughput assessment of drug cardiac safety using a high-speed impedance detection technology-based heart-on-a-chip. Micromachines 2016, 7, 122. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Reference | Shape | Cardiac Cells | Supporting Cells | Scaffold | Contractility Performances | FFR | Frank-Starling |
|---|---|---|---|---|---|---|---|
| Zhang et al., 2013 [143] | Cardiac patch | hESC-CMs | None | Fibrinogen Matrigel | 11.8 ± 4.5 mN/mm | ND | Yes |
| Shadrin et al., 2017 [140] | Cardiac patch | hiPSC-CMs | None | Fibrinogen Matrigel | 13.3 ± 1.0 mN/mm | Flat to negative | Yes |
| Turnbull et al., 2014 [117] | Cardiac strip | hiPSC-CMs | None | Bovine collagen type I Matrigel | 0.57 mN/mm | Negative | Yes |
| Zhao et al., 2019 [72] | Cardiac strip | hiPSC-CMs (ventricular or atrial) | None | Rat tail collagen Matrigel | 0.051 ± 0.025 mN/mm | Positive | ND |
| Mannhardt et al., 2016 [154] | Cardiac strip | hiPSC-CMs | None | Fibrin Matrigel | 0.5 mN/mm | Flat | Yes |
| Ronaldson-Bouchard et al., 2018 [45] | Cardiac strip | hiPSC-CMs | Human dermal fibroblasts | Fibrinogen Thrombin | ∼2.5 mN/mm | Positive | ND |
| Tulloch et al., 2011 [80] | Cardiac strip | hESC-CMs hIPSC-CMs | Stromal | Collagen | 0.08 mN/mm | ND | Yes |
| Guyette et al., 2016 [106] | Cardiac strip | hiPSC-CMs | None | Decellularized ECM | 124.1 ± 94.7 µN | Flat | ND |
| Goldfracht et al., 2020 [159] | Cardiac ring | hESC-CMs (ventricular or atrial) | None | Bovine collagen | 0.92 ± 0.09 mN/mm (ventricular) 0.19 ± 0.04 mN/mm (atrial) | ND | Yes |
| Li et al., 2020 [157] | Cardiac ring | hiPSC-CMs | None | None | 0.23 ± 0.12 mN/mm (0 ReW) 0.54 ± 0.15 mN/mm (2 ReW) | ND | Yes |
| Tiburcy et al., 2017 [141] | Cardiac ring | hESC-CMs | Fibroblasts | Rat tail collagen Matrigel | 6.2 ± 0.8 mN/mm | Positive | Yes |
| Li et al., 2019 [161] | Cardiac chamber | hESC-CMs | Fibroblasts | Bovine collagen type I Matrigel | Pressure ∼50 m Hg | ND | ND |
| Guyette et al., 2016 [106] | Cardiac chamber | hiPSC-CMs | None | Decellularized ECM | Pressure: 2.4 ± 0.1 mm Hg | ND | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seguret, M.; Vermersch, E.; Jouve, C.; Hulot, J.-S. Cardiac Organoids to Model and Heal Heart Failure and Cardiomyopathies. Biomedicines 2021, 9, 563. https://doi.org/10.3390/biomedicines9050563
Seguret M, Vermersch E, Jouve C, Hulot J-S. Cardiac Organoids to Model and Heal Heart Failure and Cardiomyopathies. Biomedicines. 2021; 9(5):563. https://doi.org/10.3390/biomedicines9050563
Chicago/Turabian StyleSeguret, Magali, Eva Vermersch, Charlène Jouve, and Jean-Sébastien Hulot. 2021. "Cardiac Organoids to Model and Heal Heart Failure and Cardiomyopathies" Biomedicines 9, no. 5: 563. https://doi.org/10.3390/biomedicines9050563
APA StyleSeguret, M., Vermersch, E., Jouve, C., & Hulot, J. -S. (2021). Cardiac Organoids to Model and Heal Heart Failure and Cardiomyopathies. Biomedicines, 9(5), 563. https://doi.org/10.3390/biomedicines9050563