
Possible Therapeutic Options for Complex Regional Pain Syndrome



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and CRPS Model
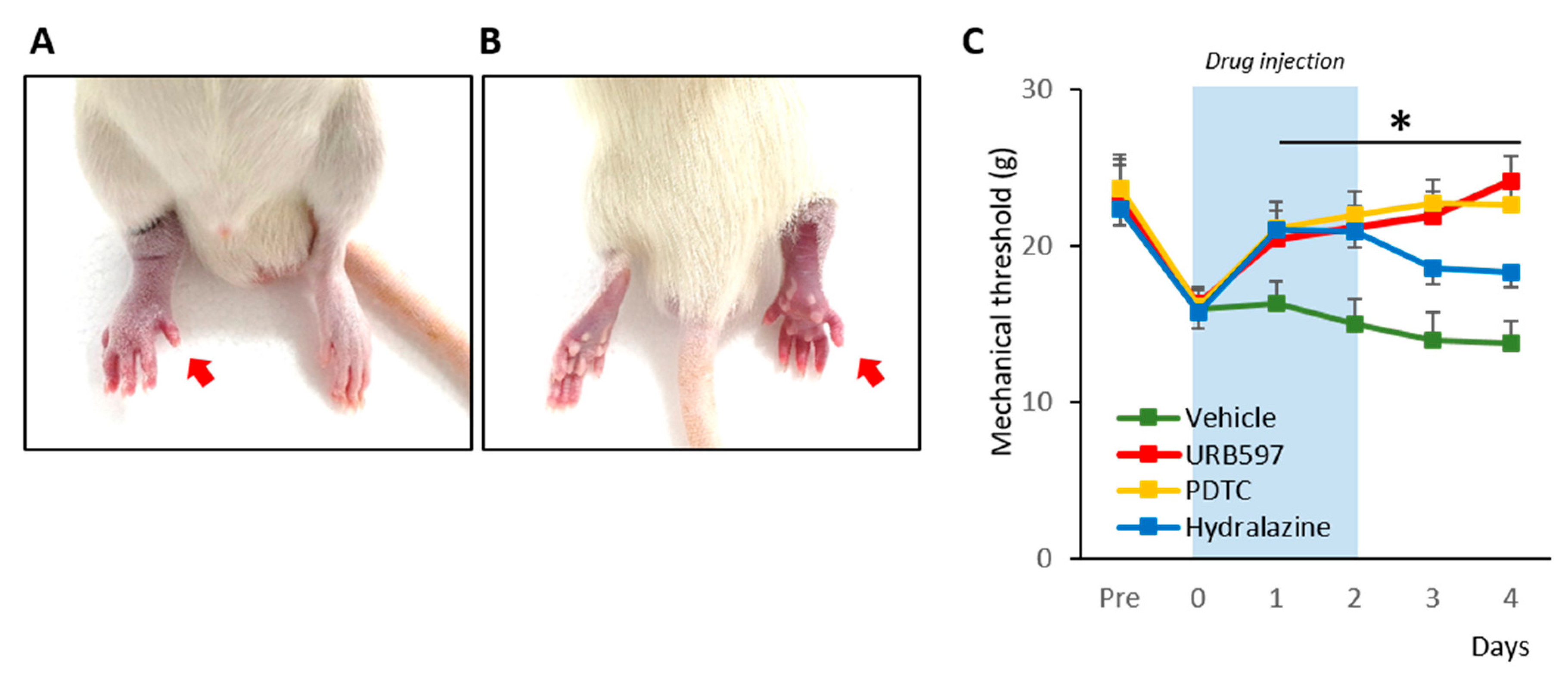
2.2. Drug Administration and Mechanical Allodynia Assessment
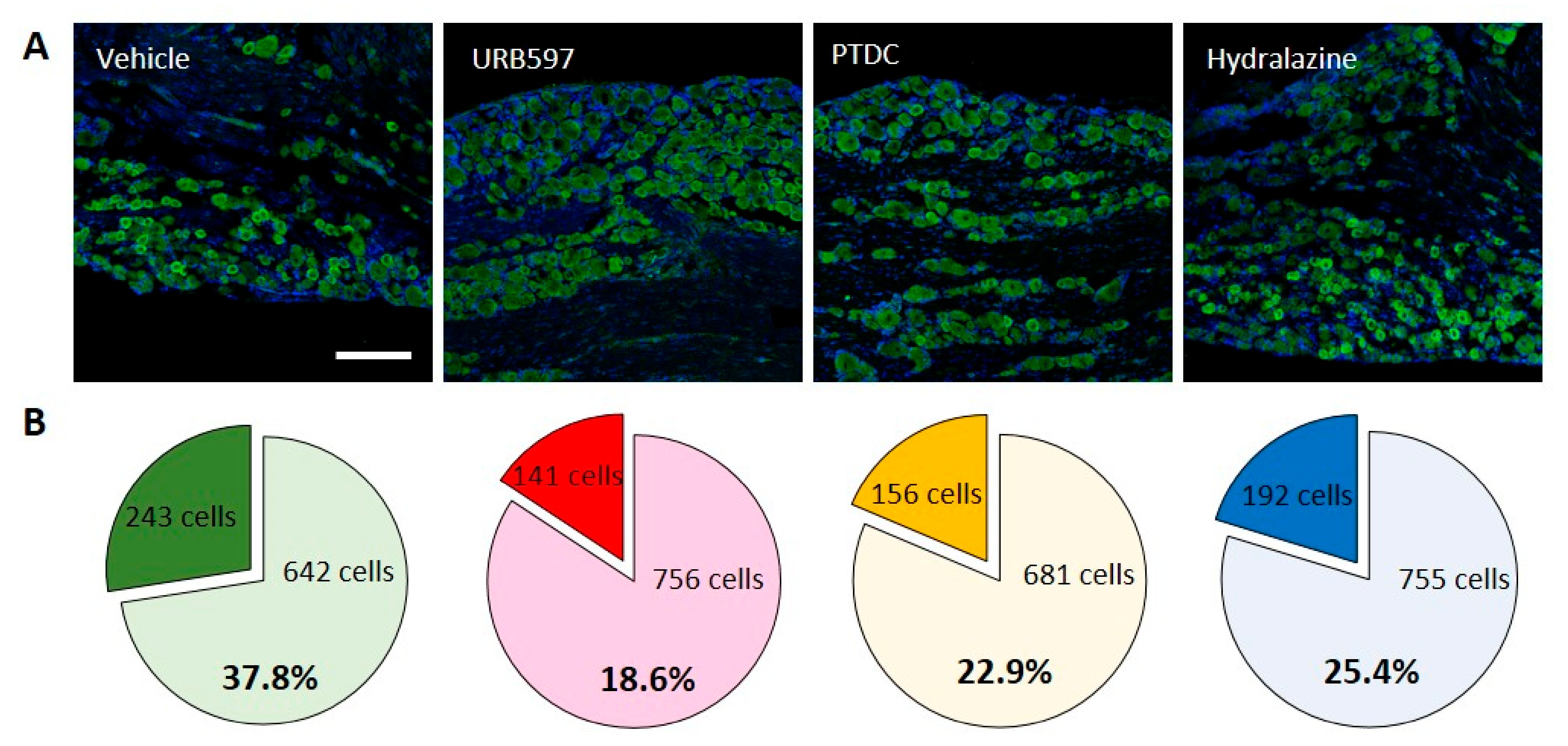
2.3. Immunohistochemistry
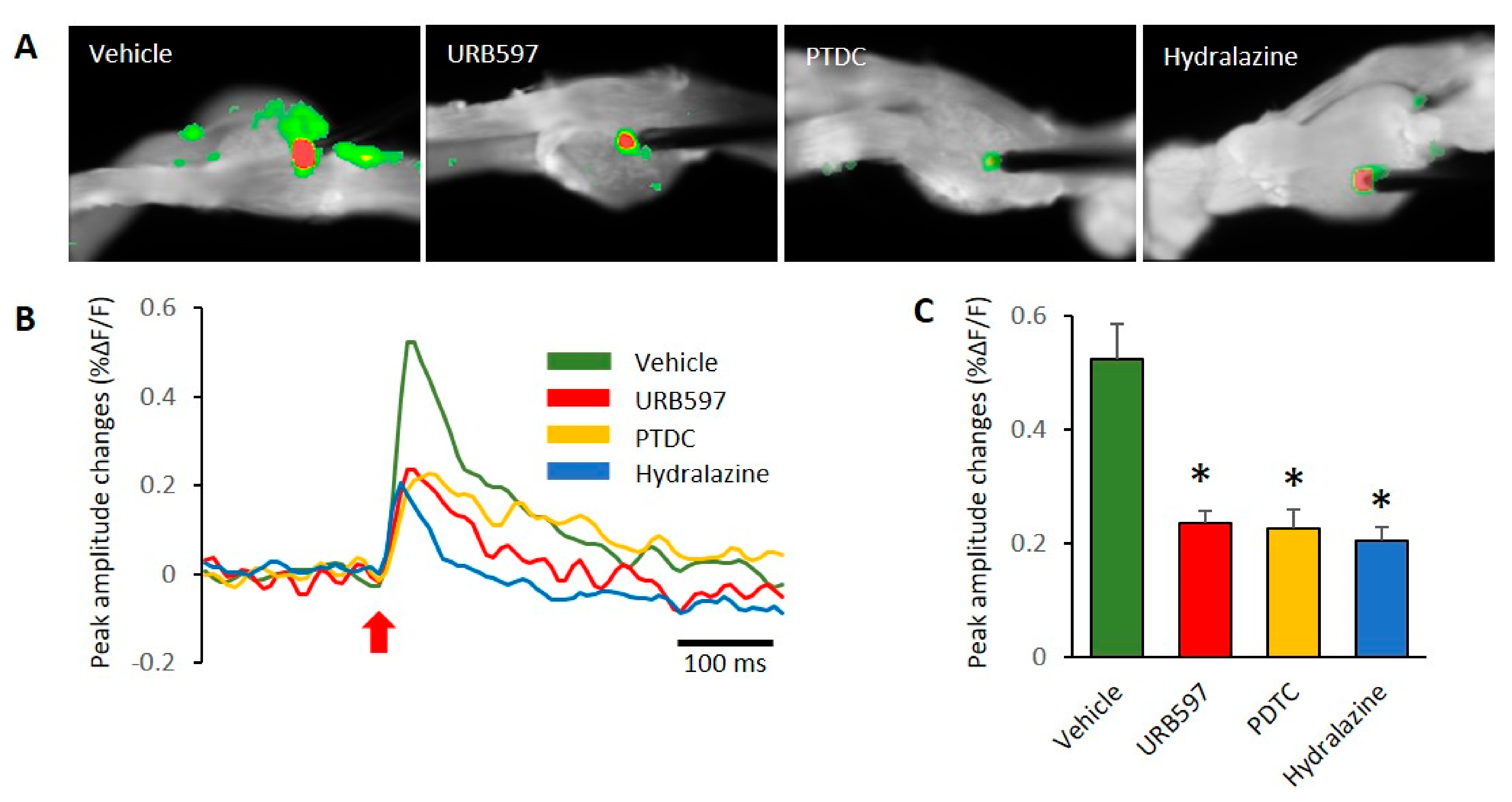
2.4. Voltage-Sensitive Dye Imaging
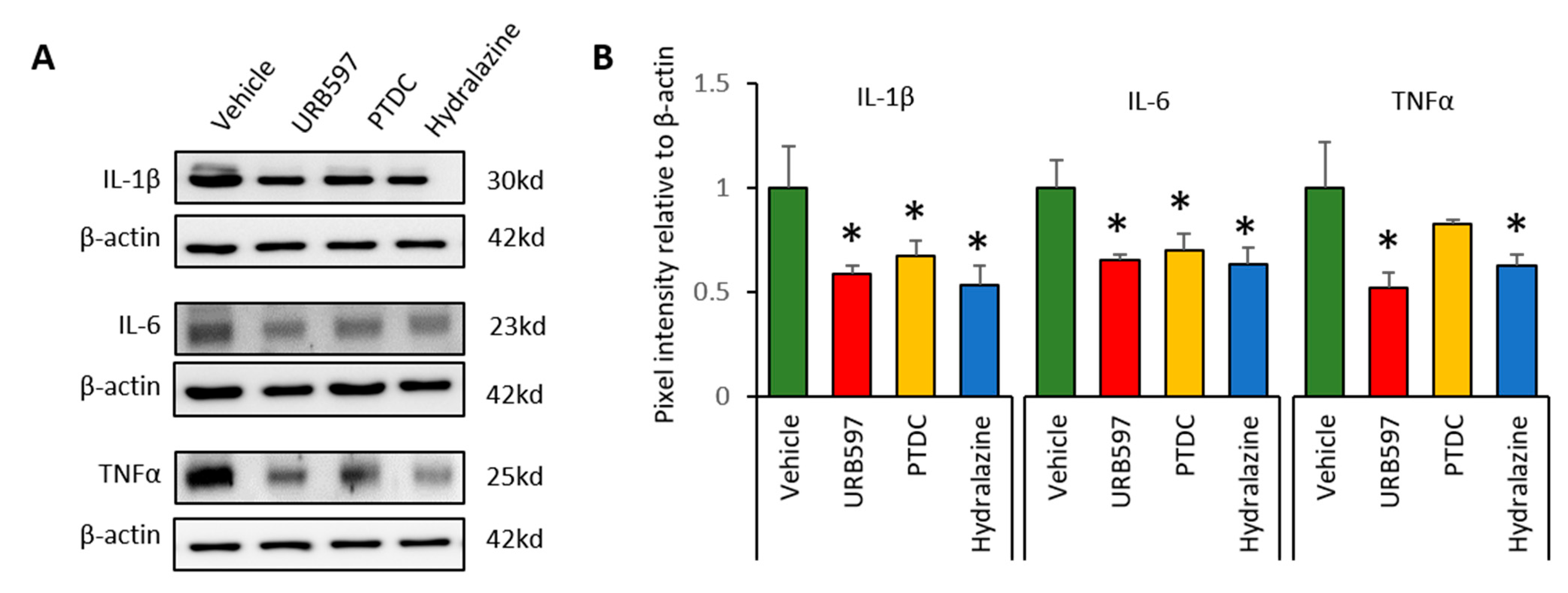
2.5. Western Blot
2.6. Statistical Analysis
3. Results
3.1. Establishment of the CPIP Model and Nocifensive Behavior Changes
3.2. Cellular Expression of Nav1.7 in DRGs
3.3. Spatial and Temporal Differences in Neural Responses after Electrical Stimulation
3.4. Expression Changes in Il-1β, Il-6, and TNFα in DRGs after Drug Application
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ott, S.; Maihöfner, C. Signs and Symptoms in 1,043 Patients with Complex Regional Pain Syndrome. J. Pain 2018, 19, 599–611. [Google Scholar] [CrossRef] [PubMed]
- Halicka, M.; Vittersø, A.D.; Proulx, M.J.; Bultitude, J.H. Neuropsychological Changes in Complex Regional Pain Syndrome (CRPS). Behav. Neurol. 2020, 2020, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Stanton-Hicks, M.; Jänig, W.; Hassenbusch, S.; Haddox, J.; Boas, R.; Wilson, P. Reflex sympathetic dystrophy: Changing concepts and taxonomy. Pain 1995, 63, 127–133. [Google Scholar] [CrossRef]
- Birklein, F.; Dimova, V. Complex regional pain syndrome–up-to-date. Pain Rep. 2017, 2, e624. [Google Scholar] [CrossRef] [PubMed]
- Sigtermans, M.J.; Van Hilten, J.J.; Bauer, M.C.; Arbous, S.M.; Marinus, J.; Sarton, E.Y.; Dahan, A. Ketamine produces effective and long-term pain relief in patients with Complex Regional Pain Syndrome Type 1. Pain 2009, 145, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Nama, S.; Meenan, D.R.; Fritz, W.T. The use of sub-anesthetic intravenous ketamine and adjuvant dexmedetomidine when treating acute pain from CRPS. Pain Physician 2010, 13, 365–368. [Google Scholar] [PubMed]
- Lee, B.; Henderson, L.A.; Rae, C.D.; Di Pietro, F. CRPS Is Not Associated with Altered Sensorimotor Cortex GABA or Glutamate. Eneuro 2020, 7. [Google Scholar] [CrossRef]
- Elsamadicy, A.A.; Yang, S.; Sergesketter, A.R.; Ashraf, B.; Charalambous, L.; Kemeny, H.; Ejikeme, T.; Ren, X.; Pagadala, P.; Parente, B.; et al. Prevalence and Cost Analysis of Complex Regional Pain Syndrome (CRPS): A Role for Neuromodulation. Neuromodulation Technol. Neural Interface 2017, 21, 423–430. [Google Scholar] [CrossRef]
- Cuhadar, U.; Gentry, C.; Vastani, N.; Sensi, S.; Bevan, S.; Goebel, A.; Andersson, D.A. Autoantibodies produce pain in complex regional pain syndrome by sensitizing nociceptors. Pain 2019, 160, 2855–2865. [Google Scholar] [CrossRef]
- Coderre, T.J.; Bennett, G.J. A Hypothesis for the Cause of Complex Regional Pain Syndrome-Type I (Reflex Sympathetic Dystrophy): Pain Due to Deep-Tissue Microvascular Pathology. Pain Med. 2010, 11, 1224–1238. [Google Scholar] [CrossRef] [Green Version]
- Kortekaas, M.C.; Niehof, S.P.; Stolker, R.J.; Huygen, F.J. Pathophysiological Mechanisms Involved in Vasomotor Disturbances in Complex Regional Pain Syndrome and Implications for Therapy: A Review. Pain Pract. 2015, 16, 905–914. [Google Scholar] [CrossRef]
- Poree, L.; Krames, E.; Pope, J.; Deer, T.R.; Levy, R.; Schultz, L. Spinal Cord Stimulation as Treatment for Complex Regional Pain Syndrome Should Be Considered Earlier Than Last Resort Therapy. Neuromodulation Technol. Neural Interface 2013, 16, 125–141. [Google Scholar] [CrossRef]
- Martins, L.; Rezende, M.; Loguercio, A.; Bortoluzzi, M.; Reis, A. Analgesic efficacy of ketorolac associated with a tramadol/acetaminophen combination after third molar surgery - a randomized, triple-blind clinical trial. Medicina Oral Patología Oral Cirugia Bucal 2018, 24, e96–e102. [Google Scholar] [CrossRef]
- Ullah, S.; Edrees, M.; Alhabtar, A.M. Baclofen Unusual Response In Spinal Cord Injury Patients. J. Ayub Med. Coll. Abbottabad 2019, 31, 269–271. [Google Scholar]
- O’Connor, A.B.; Dworkin, R.H. Treatment of Neuropathic Pain: An Overview of Recent Guidelines. Am. J. Med. 2009, 122, S22–S32. [Google Scholar] [CrossRef]
- Orhurhu, V.J.; Roberts, J.S.; Ly, N.; Cohen, S.P. Ketamine in Acute and Chronic Pain Management; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2021. [Google Scholar]
- Kirkpatrick, A.F.; Saghafi, A.; Yang, K.; Qiu, P.; Alexander, J.; Bavry, E.; Schwartzman, R. Optimizing the Treatment of CRPS With Ketamine. Clin. J. Pain 2020, 36, 516–523. [Google Scholar] [CrossRef]
- Coderre, T.J.; Xanthos, D.N.; Francis, L.; Bennett, G.J. Chronic post-ischemia pain (CPIP): A novel animal model of complex regional pain syndrome-Type I (CRPS-I; reflex sympathetic dystrophy) produced by prolonged hindpaw ischemia and reperfusion in the rat. Pain 2004, 112, 94–105. [Google Scholar] [CrossRef]
- Millecamps, M.; Laferrière, A.; Ragavendran, V.J.; Stone, L.S.; Coderre, T.J. Role of peripheral endothelin receptors in an animal model of complex regional pain syndrome type 1 (CRPS-I). Pain 2010, 151, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Cha, M.; Chae, Y.; Bai, S.J.; Lee, B.H. Spatiotemporal changes of optical signals in the somatosensory cortex of neuropathic rats after electroacupuncture stimulation. BMC Complement. Altern. Med. 2017, 17, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Lee, J.M.; Kim, Y.H.; Choi, J.H.; Jeon, S.H.; Kim, D.K.; Jeong, H.D.; Lee, Y.J.; Park, H.J. Antiallodynic Effects of Bee Venom in an Animal Model of Complex Regional Pain Syndrome Type 1 (CRPS-I). Toxins 2017, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Mor, M.; Rivara, S.; Lodola, A.; Plazzi, P.V.; Tarzia, G.; Duranti, A.; Tontini, A.; Piersanti, G.; Kathuria, A.S.; Piomelli, D. Cyclohexylcarbamic Acid 3‘- or 4‘-Substituted Biphenyl-3-yl Esters as Fatty Acid Amide Hydrolase Inhibitors: Synthesis, Quantitative Structure−Activity Relationships, and Molecular Modeling Studies. J. Med. Chem. 2004, 47, 4998–5008. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.P.; Cravatt, B.F. Mechanism of Carbamate Inactivation of FAAH: Implications for the Design of Covalent Inhibitors and In Vivo Functional Probes for Enzymes. Chem. Biol. 2005, 12, 1179–1187. [Google Scholar] [CrossRef] [Green Version]
- Mitrirattanakul, S.; Ramakul, N.; Guerrero, A.V.; Matsuka, Y.; Ono, T.; Iwase, H.; Mackie, K.; Faull, K.F.; Spigelman, I. Site-specific increases in peripheral cannabinoid receptors and their endogenous ligands in a model of neuropathic pain. Pain 2006, 126, 102–114. [Google Scholar] [CrossRef] [Green Version]
- Malek, N.; Mrugala, M.; Makuch, W.; Kolosowska, N.; Przewlocka, B.; Binkowski, M.; Czaja, M.; Morera, E.; Di Marzo, V.; Starowicz, K. A multi-target approach for pain treatment. Pain 2015, 156, 890–903. [Google Scholar] [CrossRef]
- Nasirinezhad, F.; Jergová, S.; Pearson, J.P.; Sagen, J. Attenuation of persistent pain-related behavior by fatty acid amide hydrolase (FAAH) inhibitors in a rat model of HIV sensory neuropathy. Neuropharmacology 2015, 95, 100–9. [Google Scholar] [CrossRef] [Green Version]
- Seillier, A.; Aguilar, D.D.; Giuffrida, A. The dual FAAH/MAGL inhibitor JZL195 has enhanced effects on endocannabinoid transmission and motor behavior in rats as compared to those of the MAGL inhibitor JZL184. Pharmacol. Biochem. Behav. 2014, 124, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.Z.; Ando, H.; Unno, S.; Kitagawa, J. Targeting Peripherally Restricted Cannabinoid Receptor 1, Cannabinoid Receptor 2, and Endocannabinoid-Degrading Enzymes for the Treatment of Neuropathic Pain Including Neuropathic Orofacial Pain. Int. J. Mol. Sci. 2020, 21, 1423. [Google Scholar] [CrossRef] [Green Version]
- Sasso, O.; Bertorelli, R.; Bandiera, T.; Scarpelli, R.; Colombano, G.; Armirotti, A.; Moreno-Sanz, G.; Reggiani, A.; Piomelli, D. Peripheral FAAH inhibition causes profound antinociception and protects against indomethacin-induced gastric lesions. Pharmacol. Res. 2012, 65, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Sun, R.; Ma, N.; Liu, Q.; Sun, X.; Zi, P.; Wang, J.; Chao, K.; Yu, L. Inhibition of nuclear factor-κB signal by pyrrolidine dithiocarbamate alleviates lipopolysaccharide-induced acute lung injury. Oncotarget 2017, 8, 47296–47304. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Wu, P.; Zhao, L.; Huang, L.; Zhang, Z.; Zhao, S. NF-κB Expression and Outcomes in Solid Tumors. Medicine 2015, 94, e1687. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-κB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Pinho-Ribeiro, F.A.; Fattori, V.; Zarpelon, A.C.; Borghi, S.M.; Staurengo-Ferrari, L.; de Carvalho, T.T.; Alves-Filho, J.C.; Cunha, F.Q.; Cunha, T.; Casagrande, R.; et al. Pyrrolidine dithiocarbamate inhibits superoxide anion-induced pain and inflammation in the paw skin and spinal cord by targeting NF-κB and oxidative stress. Inflammopharmacology 2016, 24, 97–107. [Google Scholar] [CrossRef] [PubMed]
- De Mos, M.; Laferrière, A.; Millecamps, M.; Pilkington, M.; Sturkenboom, M.C.; Huygen, F.J.; Coderre, T.J. Role of NFκB in an Animal Model of Complex Regional Pain Syndrome–type I (CRPS-I). J. Pain 2009, 10, 1161–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kario, K. 26—Central sympathetic agents and direct vasodilators. In Hypertension: A Companion to Braunwald’s Heart Disease, 3rd ed.; Bakris, G.L., Sorrentino, M.J., Eds.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 3, pp. 254–260. [Google Scholar]
- Li, Y.; Hou, D.; Chen, X.; Zhu, J.; Zhang, R.; Sun, W.; Li, P.; Tian, Y.; Kong, X. Hydralazine protects against renal ischemia-reperfusion injury in rats. Eur. J. Pharmacol. 2019, 843, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Stanton-Hicks, M. Complex regional pain syndrome. Anesthesiol. Clin. N. Am. 2003, 21, 733–744. [Google Scholar] [CrossRef]
- Bruehl, S. Complex regional pain syndrome. BMJ 2015, 351, h2730. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cha, M.; Lee, K.H.; Kwon, M.; Lee, B.H. Possible Therapeutic Options for Complex Regional Pain Syndrome. Biomedicines 2021, 9, 596. https://doi.org/10.3390/biomedicines9060596
Cha M, Lee KH, Kwon M, Lee BH. Possible Therapeutic Options for Complex Regional Pain Syndrome. Biomedicines. 2021; 9(6):596. https://doi.org/10.3390/biomedicines9060596
Chicago/Turabian StyleCha, Myeounghoon, Kyung Hee Lee, Minjee Kwon, and Bae Hwan Lee. 2021. "Possible Therapeutic Options for Complex Regional Pain Syndrome" Biomedicines 9, no. 6: 596. https://doi.org/10.3390/biomedicines9060596
APA StyleCha, M., Lee, K. H., Kwon, M., & Lee, B. H. (2021). Possible Therapeutic Options for Complex Regional Pain Syndrome. Biomedicines, 9(6), 596. https://doi.org/10.3390/biomedicines9060596