
Different Within-Host Viral Evolution Dynamics in Severely Immunosuppressed Cases with Persistent SARS-CoV-2

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diagnostic RT-PCR
2.2. Whole Genome Sequencing
2.3. Short Tandem Repeat Analysis
2.4. Cell Cultures
2.5. Subgenomic RNA
3. Results
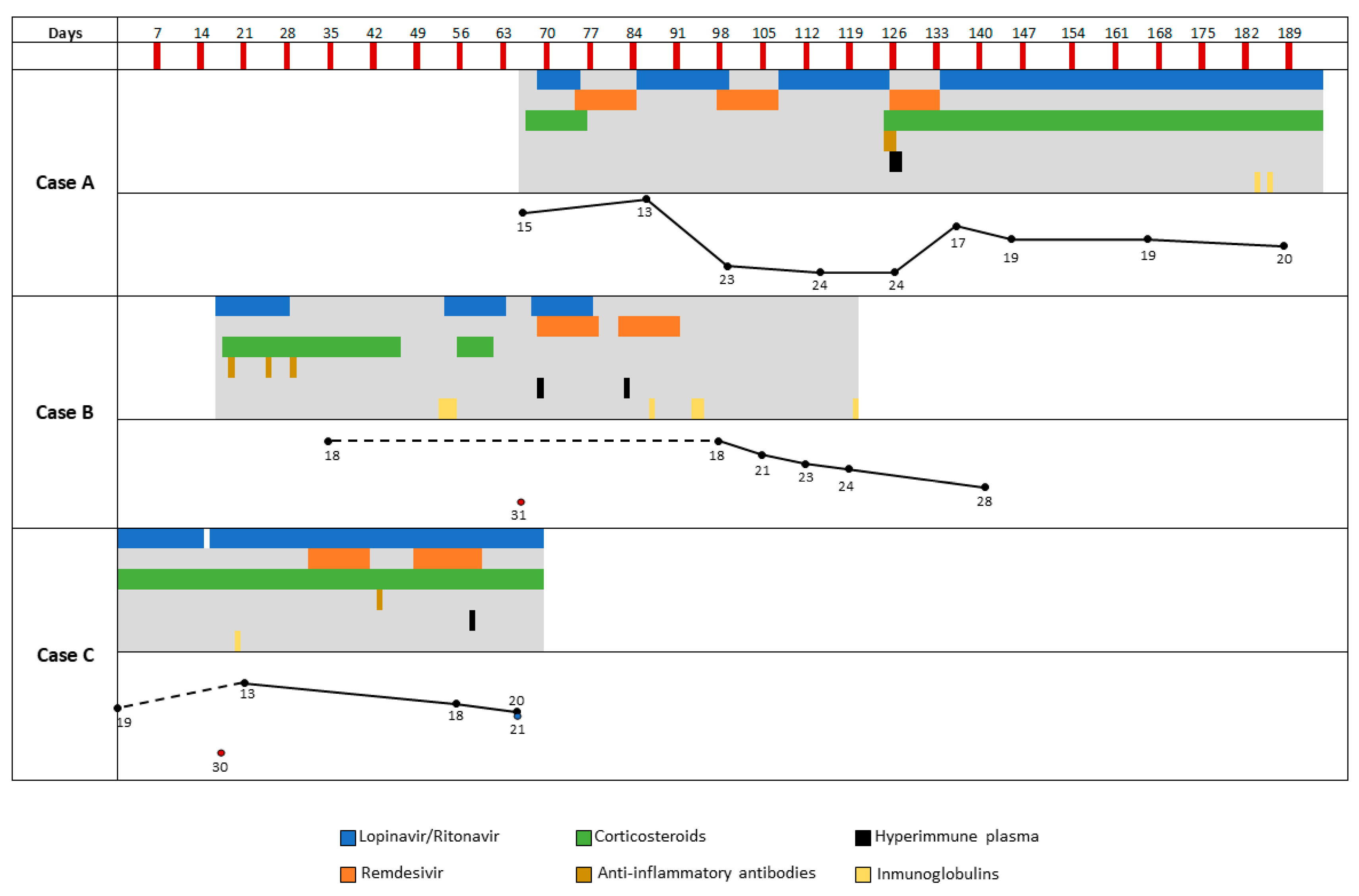
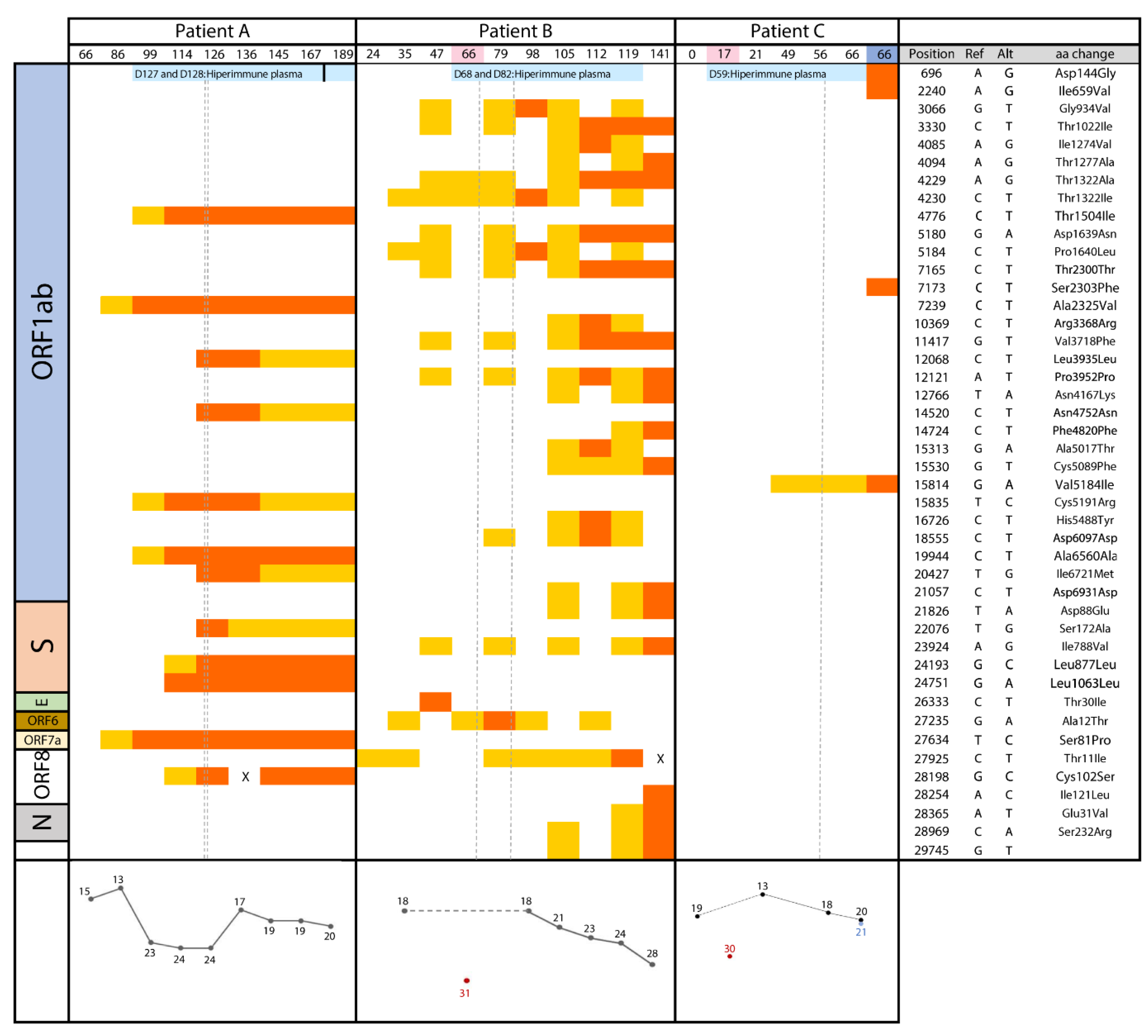
3.1. Case A
3.1.1. Clinical Case
3.1.2. Microbiological Analysis
3.2. Case B
3.2.1. Clinical Case
3.2.2. Microbiological Analysis
3.3. Case C
3.3.1. Clinical Case
3.3.2. Microbiological Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cevik, M.; Tate, M.; Lloyd, O.; Maraolo, A.E.; Schafers, J.; Ho, A. SARS-CoV-2, SARS-CoV, and MERS-CoV viral load dynamics, duration of viral shedding, and infectiousness: A systematic review and meta-analysis. Lancet Microbe 2020, 20, 30172–30175. [Google Scholar] [CrossRef]
- Rodriguez-Grande, C.; Adan-Jimenez, J.; Catalan, P.; Alcala, L.; Estevez, A.; Munoz, P.; Perez-Lago, L.; de Viedma, D.G. Inference of active viral replication in cases with sustained positive RT-PCRs for SARS-CoV-2. J. Clin. Microbiol. 2020. [Google Scholar] [CrossRef]
- Alm, E.; Broberg, E.K.; Connor, T.; Hodcroft, E.B.; Komissarov, A.B.; Maurer-Stroh, S.; Melidou, A.; Neher, R.A.; O’Toole, A.; Pereyaslov, D. Geographical and temporal distribution of SARS-CoV-2 clades in the WHO European Region, January to June 2020. Eurosurveillance 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Hodcroft, E.B.; Zuber, M.; Nadeau, S.; Crawford, K.H.D.; Bloom, J.D.; Veesler, D.; Vaughan, T.G.; Comas, I.; Candelas, F.G.; Stadler, T.; et al. Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020. Medrxiv Prepr. Serv. Health Sci. 2020. [Google Scholar] [CrossRef]
- Lucey, M.; Macori, G.; Mullane, N.; Sutton-Fitzpatrick, U.; Gonzalez, G.; Coughlan, S.; Purcell, A.; Fenelon, L.; Fanning, S.; Schaffer, K. Whole-genome sequencing to track SARS-CoV-2 transmission in nosocomial outbreaks. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Mulder, M.; van der Vegt, D.; Oude Munnink, B.B.; GeurtsvanKessel, C.H.; van de Bovenkamp, J.; Sikkema, R.S.; Jacobs, E.M.G.; Koopmans, M.P.G.; Wegdam-Blans, M.C.A. Reinfection of SARS-CoV-2 in an immunocompromised patient: A case report. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Aydillo, T.; Gonzalez-Reiche, A.S.; Aslam, S.; van de Guchte, A.; Khan, Z.; Obla, A.; Dutta, J.; van Bakel, H.; Aberg, J.; Garcia-Sastre, A.; et al. Shedding of viable SARS-CoV-2 after immunosuppressive therapy for cancer. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Baang, J.H.; Smith, C.; Mirabelli, C.; Valesano, A.L.; Manthei, D.M.; Bachman, M.; Wobus, C.E.; Adams, M.; Washer, L.; Martin, E.T.; et al. Prolonged SARS-CoV-2 replication in an immunocompromised patient. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Choi, B.; Choudhary, M.C.; Regan, J.; Sparks, J.A.; Padera, R.F.; Qiu, X.; Solomon, I.H.; Kuo, H.H.; Boucau, J.; Bowman, K.; et al. Persistence and evolution of SARS-CoV-2 in an immunocompromised host. N. Engl. J. Med. 2020, 383, 2291–2293. [Google Scholar] [CrossRef] [PubMed]
- Threat Assessment Brief. Rapid increase of a SARS-CoV-2 variant with multiple spike protein mutations observed in the United Kingdom. Epidemiology 2020, 7, 1–13. [Google Scholar]
- Wolfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Muller, M.A.; Niemeyer, D.; Jones, T.C.; Vollmar, P.; Rothe, C.; et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 581, 465–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Loman, N.; Pybus, O.; Barclay, W.; Barrett, J.; Carabelli, A.; Connor, T.; Peacock, T.; Robertson, D.L.; Volz, E.; et al. Preliminary genomic characterisation of an emergent SARS-CoV-2 lineage in the UK defined by a novel set of spike mutations. Virological. Org. 2020. [Google Scholar]
- Hung, I.; To, K.; Lee, C.; Lee, K.; Chan, K.; Yan, W.; Liu, R.; Watt, C.; Chan, W.; Lai, K.; et al. Convalescent plasma treatment reduced mortality in patients with severe pandemic influenza A (H1N1) 2009 virus infection. Clin. Infect. Dis. 2011, 52, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.D.; Datir, R.; Gayed, S.; Jahun, A.; Hosmillo, M.; Ferreira, I.A.T.M.; Rees-Spear, C.; Mlcochova, P.; Lumb, I.U. Neutralising antibodies drive Spike mediated SARS-CoV-2 evasion. MedRxiv 2020. [Google Scholar] [CrossRef]
- Kemp, S.A.; Datir, R.P.; Collier, D.A.; Ferreira, I.A.T.M.; Carabelli, A.; Harvey, W.; Robertson, D.L.; Gupta, R.K. Recurrent emergence and transmission of a SARS-CoV-2 Spike deletion ΔH69/ΔV70. BioRxiv 2020. [Google Scholar] [CrossRef]
- Martin, M.A.; VanInsberghe, D.; Koelle, K. Insights from SARS-CoV-2 sequences. Science 2021, 371, 466–467. [Google Scholar] [CrossRef]
- Baric, R.S. Emergence of a highly fit SARS-CoV-2 variant. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Rueca, M.; Bartolini, B.; Maria Gruber, C.; Piralla, A.; Baldanti, F.; Giombini, E.; Messina, F.; Marchioni, L.; Ippolito, G.; Di Caro, A.; et al. Compartmentalized replication of SARS-Cov-2 in upper vs. lower respiratory tract assessed by whole genome quasispecies analysis. Microroganisms 2020, 8, 1302. [Google Scholar] [CrossRef] [PubMed]
- Du, P.; Song, C.; Li, R.; Song, Y.; Li, J.Z.; Ding, N.; Zhang, J.; Song, R.; Han, J.; Gao, G.; et al. Specific redistribution of severe acute respiratory syndrome coronavirus 2 variants in the respiratory system and intestinal tract. Clin. Infect. Dis. 2020, in press. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Case A | 27/May–15/June | 15/June–30/June | 01/July–15/July | 16/July–30/July | |
| Lymphocytes (×103/µL) | 300 | 700 | 1100 | 600 | |
| D-dimer (ng/mL) | 309 | 286 | 233 | 378 | |
| LDH (U/L) | 262 | 280 | 303 | 418 | |
| C- reactive protein (mg/dL) | 3.2 | 8.3 | 15.9 | 22.4 | |
| Ferritin (µg/L) | 4797 | 3683 | 7504 | 7433 | |
| IL-6 (pg/mL) | 20 | 20.5 | 56.3 | 335 | |
| CD4 (cells/µL) | 86 (23%) | 175 | |||
| IgG (mg/dL) | 545 | 517 | 429 | ||
| SARS-CoV-2 Serology | Negative | Negative | Negative | Negative | |
| Case B | 8/April | 14/May | 12/August | 20/October | |
| Lymphocytes (×103/µL) | 100 | 100 | 400 | 500 | |
| D-dimer (ng/mL) | 581 | 168 | |||
| LDH (U/L) | 374 | 326 | 403 | ||
| C- reactive protein (mg/dL) | 4.1 | 10.3 | 0.5 | 0.3 | |
| Ferritin (µg/L) | 4559 | 2723 | 575 | 50 | |
| IL-6 (pg/mL) | 44.1 | 88.5 | |||
| CD4 (cells/µL) | 17 ( 16%) | 9 (27.3%) | 106(25.7%) | 109(20.8%) | |
| IgG (mg/dL) | 578 | 353 | 1070 | 760 | |
| SARS-CoV-2 Serology | Negative | Negative | Negative | ||
| Case C | 3/April–30/April | 1/May–14/May | 15/May–31/May | 1/June–15/June | 5/July |
| Lymphocytes (×103/µL) | 200 | 300 | 100 | 200 | 100 |
| D-dimer (ng/mL) | 185 | 1007 | 837 | 384 | |
| LDH (U/L) | 203 | 268 | 350 | 246 | 199 |
| C- reactive protein (mg/dL) | 5 | 3.7 | 5.3 | 4.5 | 14.3 |
| Ferritin (µg/L) | 2349 | 2655 | 2637 | 4940 | |
| IL-6 (pg/mL) | 38.4 | 56.1 | 108.4 | ||
| CD4 (cells/µL) | 45 | ||||
| IgG (mg/dL) | 917 | 605 | |||
| SARS-CoV-2 Serology | Negative | Negative | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Lago, L.; Aldámiz-Echevarría, T.; García-Martínez, R.; Pérez-Latorre, L.; Herranz, M.; Sola-Campoy, P.J.; Suárez-González, J.; Martínez-Laperche, C.; Comas, I.; González-Candelas, F.; et al. Different Within-Host Viral Evolution Dynamics in Severely Immunosuppressed Cases with Persistent SARS-CoV-2. Biomedicines 2021, 9, 808. https://doi.org/10.3390/biomedicines9070808
Pérez-Lago L, Aldámiz-Echevarría T, García-Martínez R, Pérez-Latorre L, Herranz M, Sola-Campoy PJ, Suárez-González J, Martínez-Laperche C, Comas I, González-Candelas F, et al. Different Within-Host Viral Evolution Dynamics in Severely Immunosuppressed Cases with Persistent SARS-CoV-2. Biomedicines. 2021; 9(7):808. https://doi.org/10.3390/biomedicines9070808
Chicago/Turabian StylePérez-Lago, Laura, Teresa Aldámiz-Echevarría, Rita García-Martínez, Leire Pérez-Latorre, Marta Herranz, Pedro J. Sola-Campoy, Julia Suárez-González, Carolina Martínez-Laperche, Iñaki Comas, Fernando González-Candelas, and et al. 2021. "Different Within-Host Viral Evolution Dynamics in Severely Immunosuppressed Cases with Persistent SARS-CoV-2" Biomedicines 9, no. 7: 808. https://doi.org/10.3390/biomedicines9070808
APA StylePérez-Lago, L., Aldámiz-Echevarría, T., García-Martínez, R., Pérez-Latorre, L., Herranz, M., Sola-Campoy, P. J., Suárez-González, J., Martínez-Laperche, C., Comas, I., González-Candelas, F., Catalán, P., Muñoz, P., García de Viedma, D., & on behalf of Gregorio Marañón Microbiology-ID COVID 19 Study Group. (2021). Different Within-Host Viral Evolution Dynamics in Severely Immunosuppressed Cases with Persistent SARS-CoV-2. Biomedicines, 9(7), 808. https://doi.org/10.3390/biomedicines9070808