
Both Full-Length and Protease-Cleaved Products of Osteopontin Are Elevated in Infectious Diseases

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. OPN Proteolysis and Bioactivities of Cleaved OPNs
2.1. Binding Specificities and Cellular Functions of Thrombin-Cleaved OPN
2.2. Binding Specificities and Functions of MMP-Cleaved OPN
2.3. Binding Specificities and Functions of Caspase-Cleaved OPN
2.4. Binding Specificities and Functions of Plasmin and Cathepsin D-Cleaved OPN
2.5. Gradual Proteolysis of OPN Regulating Cellular Functions
2.6. Different Expression and Localization between FL-OPN and Its Cleaved OPNs
2.7. The Roles of Thrombin-Cleaved OPN in the Pathogenesis of Inflammatory Diseases
3. Immunological Activities of OPN in Infectious Diseases
4. OPN in Infectious Diseases
4.1. ATL
4.2. Tuberculosis (TB)
4.3. Acquired Immune Deficiency Sndrome (AIDS)
4.4. Hepatitis Virus Infection
4.5. OPN in Tropical Neglected Diseases
4.5.1. Dengue

4.5.2. Leptospirosis
4.5.3. Melioidosis
4.5.4. Parasite Infection
Human African Trypanosomiasis (HAT)
Schistosomiasis
4.6. Coronavirus Disease 2019 (COVID-19)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Senger, D.R.; Wirth, D.F.; Hynes, R.O. Transformed mammalian cells secrete specific proteins and phosphoproteins. Cell 1979, 16, 885–893. [Google Scholar] [CrossRef]
- Weber, G.F.; Cantor, H. The immunology of Eta-1/osteopontin. Cytokine Growth Factor Rev. 1996, 7, 241–248. [Google Scholar] [CrossRef]
- Bornstein, P. Matricellular proteins: An overview. J. Cell Commun. Signal. 2009, 3, 163–165. [Google Scholar] [CrossRef] [Green Version]
- Chiodoni, C.; Colombo, M.P.; Sangaletti, S. Matricellular proteins: From homeostasis to inflammation, cancer, and metastasis. Cancer Metastasis Rev. 2010, 29, 295–307. [Google Scholar] [CrossRef]
- Kon, S.; Maeda, M.; Segawa, T.; Hagiwara, Y.; Horikoshi, Y.; Chikuma, S.; Tanaka, K.; Rashid, M.M.; Inobe, M.; Chambers, A.F.; et al. Antibodies to different peptides in osteopontin reveal complexities in the various secreted forms. J. Cell Biochem. 2000, 77, 487–498. [Google Scholar] [CrossRef]
- Uede, T.; Katagiri, Y.; Iizuka, J.; Murakami, M. Osteopontin, a coordinator of host defense system: A cytokine or an extracellular adhesive protein? Microbiol. Immunol. 1997, 41, 641–648. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Noda, M.; O’Regan, A.W.; Pavlin, D.; Berman, J.S. Osteopontin as a means to cope with environmental insults: Regulation of inflammation, tissue remodeling, and cell survival. J. Clin. Investig. 2001, 107, 1055–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, L.L.; Cheung, H.K.; Ling, L.E.; Chen, J.; Sheppard, D.; Pytela, R.; Giachelli, C.M. Osteopontin N-terminal domain contains a cryptic adhesive sequence recognized by alpha9beta1 integrin. J. Biol. Chem. 1996, 271, 28485–28491. [Google Scholar] [CrossRef] [Green Version]
- Kon, S.; Yokosaki, Y.; Maeda, M.; Segawa, T.; Horikoshi, Y.; Tsukagoshi, H.; Rashid, M.M.; Morimoto, J.; Inobe, M.; Shijubo, N.; et al. Mapping of functional epitopes of osteopontin by monoclonal antibodies raised against defined internal sequences. J. Cell Biochem. 2002, 84, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Kurata, M.; Okura, T.; Kumon, Y.; Tagawa, M.; Watanabe, H.; Nakahara, T.; Miyazaki, T.; Higaki, J.; Nose, M. Plasma thrombin-cleaved osteopontin elevation after carotid artery stenting in symptomatic ischemic stroke patients. Hypertens Res. 2012, 35, 207–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagan-Yasutan, H.; Lacuesta, T.L.; Ndhlovu, L.C.; Oguma, S.; Leano, P.S.; Telan, E.F.; Kubo, T.; Morita, K.; Uede, T.; Dimaano, E.M.; et al. Elevated levels of full-length and thrombin-cleaved osteopontin during acute dengue virus infection are associated with coagulation abnormalities. Thromb Res. 2014, 134, 449–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, N.; Ohashi, T.; Chagan-Yasutan, H.; Hattori, T.; Takahashi, Y.; Harigae, H.; Hasegawa, H.; Yamada, Y.; Fujii, M.; Maenaka, K.; et al. Osteopontin-integrin interaction as a novel molecular target for antibody-mediated immunotherapy in adult T-cell leukemia. Retrovirology 2015, 12, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, M.; Kimura, C.; Uede, T.; Kon, S. Neutralizing antibody against osteopontin attenuates non-alcoholic steatohepatitis in mice. J. Cell Commun. Signal. 2020, 14, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.; Matsuba, T.; Kikuchi, H.; Chagan-Yasutan, H.; Motoda, H.; Ozuru, R.; Yamada, O.; Oshima, Y.; Hattori, T. Inhibition of inflammatory-molecule synthesis in THP-1 cells stimulated with phorbol 12-myristate 13-acetate by brefelamide derivatives. Int. Immunopharm. 2019, 75, 105831. [Google Scholar] [CrossRef]
- Senger, D.R.; Perruzzi, C.A.; Gracey, C.F.; Papadopoulos, A.; Tenen, D.G. Secreted phosphoproteins associated with neoplastic transformation: Close homology with plasma proteins cleaved during blood coagulation. Cancer Res. 1988, 48, 5770–5774. [Google Scholar]
- Senger, D.R.; Perruzzi, C.A.; Papadopoulos, A. Elevated expression of secreted phosphoprotein I (osteopontin, 2ar) as a consequence of neoplastic transformation. Anticancer Res. 1989, 9, 1291–1299. [Google Scholar]
- Senger, D.R.; Perruzzi, C.A.; Papadopoulos, A.; Tenen, D.G. Purification of a human milk protein closely similar to tumor-secreted phosphoproteins and osteopontin. Biochim. Biophys. Acta 1989, 996, 43–48. [Google Scholar] [CrossRef]
- Agnihotri, R.; Crawford, H.C.; Haro, H.; Matrisian, L.M.; Havrda, M.C.; Liaw, L. Osteopontin, a novel substrate for matrix metalloproteinase-3 (stromelysin-1) and matrix metalloproteinase-7 (matrilysin). J. Biol. Chem. 2001, 276, 28261–28267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.A.; Overall, C.M. Proteomics discovery of metalloproteinase substrates in the cellular context by iTRAQ labeling reveals a diverse MMP-2 substrate degradome. Mol. Cell Proteom. 2007, 6, 611–623. [Google Scholar] [CrossRef] [Green Version]
- Takafuji, V.; Forgues, M.; Unsworth, E.; Goldsmith, P.; Wang, X.W. An osteopontin fragment is essential for tumor cell invasion in hepatocellular carcinoma. Oncogene 2007, 26, 6361–6371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Lee, H.J.; Jun, J.I.; Oh, Y.; Choi, S.G.; Kim, H.; Chung, C.W.; Kim, I.K.; Park, I.S.; Chae, H.J.; et al. Intracellular cleavage of osteopontin by caspase-8 modulates hypoxia/reoxygenation cell death through p53. Proc. Natl. Acad. Sci. USA 2009, 106, 15326–15331. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.; Schack, L.; Kläning, E.; Sørensen, E.S. Osteopontin is cleaved at multiple sites close to its integrin-binding motifs in milk and is a novel substrate for plasmin and cathepsin D. J. Biol. Chem. 2010, 285, 7929–7937. [Google Scholar] [CrossRef] [Green Version]
- Giachelli, C.M.; Liaw, L.; Murry, C.E.; Schwartz, S.M.; Almeida, M. Osteopontin expression in cardiovascular diseases. Ann. N. Y. Acad. Sci. 1995, 760, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, H.F. Tumors: Wounds that do not heal. Similarities between tumor stroma generation and wound healing. N. Engl. J. Med. 1986, 315, 1650–1659. [Google Scholar] [CrossRef]
- Oldberg, A.; Franzén, A.; Heinegård, D. Cloning and sequence analysis of rat bone sialoprotein (osteopontin) cDNA reveals an Arg-Gly-Asp cell-binding sequence. Proc. Natl. Acad. Sci. USA 1986, 83, 8819–8823. [Google Scholar] [CrossRef] [Green Version]
- Senger, D.R.; Perruzzi, C.A.; Papadopoulos-Sergiou, A.; Van de Water, L. Adhesive properties of osteopontin: Regulation by a naturally occurring thrombin-cleavage in close proximity to the GRGDS cell-binding domain. Mol. Biol. Cell 1994, 5, 565–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, D.D.; Lin, E.C.; Kovach, N.L.; Hoyer, J.R.; Smith, J.W. A biochemical characterization of the binding of osteopontin to integrins alpha v beta 1 and alpha v beta 5. J. Biol. Chem. 1995, 270, 26232–26238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyauchi, A.; Alvarez, J.; Greenfield, E.M.; Teti, A.; Grano, M.; Colucci, S.; Zambonin-Zallone, A.; Ross, F.P.; Teitelbaum, S.L.; Cheresh, D.; et al. Recognition of osteopontin and related peptides by an alpha v beta 3 integrin stimulates immediate cell signals in osteoclasts. J. Biol. Chem. 1991, 266, 20369–20374. [Google Scholar] [CrossRef]
- Ross, F.P.; Chappel, J.; Alvarez, J.I.; Sander, D.; Butler, W.T.; Farach-Carson, M.C.; Mintz, K.A.; Robey, P.G.; Teitelbaum, S.L.; Cheresh, D.A. Interactions between the bone matrix proteins osteopontin and bone sialoprotein and the osteoclast integrin alpha v beta 3 potentiate bone resorption. J. Biol. Chem. 1993, 268, 9901–9907. [Google Scholar] [CrossRef]
- Liaw, L.; Almeida, M.; Hart, C.E.; Schwartz, S.M.; Giachelli, C.M. Osteopontin promotes vascular cell adhesion and spreading and is chemotactic for smooth muscle cells in vitro. Circ. Res. 1994, 74, 214–224. [Google Scholar] [CrossRef] [Green Version]
- Liaw, L.; Skinner, M.P.; Raines, E.W.; Ross, R.; Cheresh, D.A.; Schwartz, S.M.; Giachelli, C.M. The adhesive and migratory effects of osteopontin are mediated via distinct cell surface integrins. Role of alpha v beta 3 in smooth muscle cell migration to osteopontin in vitro. J. Clin. Investig. 1995, 95, 713–724. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.D.; Hoyer, J.R.; Smith, J.W. Ca2+ suppresses cell adhesion to osteopontin by attenuating binding affinity for integrin alpha v beta 3. J. Biol. Chem. 1995, 270, 9917–9925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokosaki, Y.; Tanaka, K.; Higashikawa, F.; Yamashita, K.; Eboshida, A. Distinct structural requirements for binding of the integrins alphavbeta6, alphavbeta3, alphavbeta5, alpha5beta1 and alpha9beta1 to osteopontin. Matrix Biol. 2005, 24, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Barry, S.T.; Ludbrook, S.B.; Murrison, E.; Horgan, C.M. A regulated interaction between alpha5beta1 integrin and osteopontin. Biochem. Biophys. Res. Commun. 2000, 267, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Denda, S.; Reichardt, L.F.; Müller, U. Identification of osteopontin as a novel ligand for the integrin alpha8 beta1 and potential roles for this integrin-ligand interaction in kidney morphogenesis. Mol. Biol. Cell 1998, 9, 1425–1435. [Google Scholar] [CrossRef] [PubMed]
- Yokosaki, Y.; Kido, M.; Nagata, N.; Nikaido, Y.; Tsuda, T.; Miyake, J.; Manabe, H. Hypoglycemia associated with localized fibrous mesothelioma of the pleura. J. UOEH 1995, 17, 191–197. [Google Scholar] [CrossRef] [Green Version]
- Green, P.M.; Ludbrook, S.B.; Miller, D.D.; Horgan, C.M.; Barry, S.T. Structural elements of the osteopontin SVVYGLR motif important for the interaction with alpha(4) integrins. FEBS Lett. 2001, 503, 75–79. [Google Scholar] [CrossRef] [Green Version]
- Ito, N.; Obata, H.; Saito, S. Spinal microglial expression and mechanical hypersensitivity in a postoperative pain model: Comparison with a neuropathic pain model. Anesthesiology 2009, 111, 640–648. [Google Scholar] [CrossRef] [Green Version]
- Bayless, K.J.; Davis, G.E. Identification of dual alpha 4beta1 integrin binding sites within a 38 amino acid domain in the N-terminal thrombin fragment of human osteopontin. J. Biol. Chem. 2001, 276, 13483–13489. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.L.; Giachelli, C.M. Structural requirements for alpha 9 beta 1-mediated adhesion and migration to thrombin-cleaved osteopontin. Exp. Cell Res. 1998, 242, 351–360. [Google Scholar] [CrossRef]
- Taooka, Y.; Chen, J.; Yednock, T.; Sheppard, D. The integrin alpha9beta1 mediates adhesion to activated endothelial cells and transendothelial neutrophil migration through interaction with vascular cell adhesion molecule-1. J. Cell Biol. 1999, 145, 413–420. [Google Scholar] [CrossRef]
- Yokosaki, Y.; Matsuura, N.; Sasaki, T.; Murakami, I.; Schneider, H.; Higashiyama, S.; Saitoh, Y.; Yamakido, M.; Taooka, Y.; Sheppard, D. The integrin alpha(9)beta(1) binds to a novel recognition sequence (SVVYGLR) in the thrombin-cleaved amino-terminal fragment of osteopontin. J. Biol. Chem. 1999, 274, 36328–36334. [Google Scholar] [CrossRef] [Green Version]
- Barry, S.T.; Ludbrook, S.B.; Murrison, E.; Horgan, C.M. Analysis of the alpha4beta1 integrin-osteopontin interaction. Exp. Cell Res. 2000, 258, 342–351. [Google Scholar] [CrossRef]
- Kon, S.; Ikesue, M.; Kimura, C.; Aoki, M.; Nakayama, Y.; Saito, Y.; Kurotaki, D.; Diao, H.; Matsui, Y.; Segawa, T.; et al. Syndecan-4 protects against osteopontin-mediated acute hepatic injury by masking functional domains of osteopontin. J. Exp. Med. 2008, 205, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, J.; Kon, S.; Matsui, Y.; Uede, T. Osteopontin; as a target molecule for the treatment of inflammatory diseases. Curr. Drug Targets 2010, 11, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Grassinger, J.; Haylock, D.N.; Storan, M.J.; Haines, G.O.; Williams, B.; Whitty, G.A.; Vinson, A.R.; Be, C.L.; Li, S.; Sørensen, E.S.; et al. Thrombin-cleaved osteopontin regulates hemopoietic stem and progenitor cell functions through interactions with alpha9beta1 and alpha4beta1 integrins. Blood 2009, 114, 49–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokasaki, Y.; Sheppard, D. Mapping of the cryptic integrin-binding site in osteopontin suggests a new mechanism by which thrombin can regulate inflammation and tissue repair. Trends Cardiovasc. Med. 2000, 10, 155–159. [Google Scholar] [CrossRef]
- Senger, D.R.; Perruzzi, C.A. Cell migration promoted by a potent GRGDS-containing thrombin-cleavage fragment of osteopontin. Biochim. Biophys. Acta 1996, 1314, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Senger, D.R.; Ledbetter, S.R.; Claffey, K.P.; Papadopoulos-Sergiou, A.; Peruzzi, C.A.; Detmar, M. Stimulation of endothelial cell migration by vascular permeability factor/vascular endothelial growth factor through cooperative mechanisms involving the alphavbeta3 integrin, osteopontin, and thrombin. Am. J. Pathol. 1996, 149, 293–305. [Google Scholar]
- Desai, B.; Rogers, M.J.; Chellaiah, M.A. Mechanisms of osteopontin and CD44 as metastatic principles in prostate cancer cells. Mol. Cancer 2007, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Iczkowski, K.A. Cell adhesion molecule CD44: Its functional roles in prostate cancer. Am. J. Transl. Res. 2010, 3, 1–7. [Google Scholar]
- Weber, G.F.; Zawaideh, S.; Hikita, S.; Kumar, V.A.; Cantor, H.; Ashkar, S. Phosphorylation-dependent interaction of osteopontin with its receptors regulates macrophage migration and activation. J. Leukoc. Biol. 2002, 72, 752–761. [Google Scholar]
- Katagiri, Y.U.; Sleeman, J.; Fujii, H.; Herrlich, P.; Hotta, H.; Tanaka, K.; Chikuma, S.; Yagita, H.; Okumura, K.; Murakami, M.; et al. CD44 variants but not CD44s cooperate with beta1-containing integrins to permit cells to bind to osteopontin independently of arginine-glycine-aspartic acid, thereby stimulating cell motility and chemotaxis. Cancer Res. 1999, 59, 219–226. [Google Scholar]
- Shao, Z.; Morser, J.; Leung, L.L.K. Thrombin cleavage of osteopontin disrupts a pro-chemotactic sequence for dendritic cells, which is compensated by the release of its pro-chemotactic C-terminal fragment. J. Biol. Chem. 2014, 289, 27146–27158. [Google Scholar] [CrossRef] [Green Version]
- Myles, T.; Nishimura, T.; Yun, T.H.; Nagashima, M.; Morser, J.; Patterson, A.J.; Pearl, R.G.; Leung, L.L. Thrombin activatable fibrinolysis inhibitor, a potential regulator of vascular inflammation. J. Biol. Chem. 2003, 278, 51059–51067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, S.A.; Du, X.; Myles, T.; Song, J.J.; Price, E.; Lee, D.M.; Goodman, S.B.; Nagashima, M.; Morser, J.; Robinson, W.H.; et al. Thrombin-activatable carboxypeptidase B cleavage of osteopontin regulates neutrophil survival and synoviocyte binding in rheumatoid arthritis. Arthritis Rheum. 2009, 60, 2902–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCawley, L.J.; Matrisian, L.M. Matrix metalloproteinases: Multifunctional contributors to tumor progression. Mol. Med. Today 2000, 6, 149–156. [Google Scholar] [CrossRef]
- Crawford, H.C.; Matrisian, L.M.; Liaw, L. Distinct roles of osteopontin in host defense activity and tumor survival during squamous cell carcinoma progression in vivo. Cancer Res. 1998, 58, 5206–5215. [Google Scholar]
- Wright, J.H.; McDonnell, S.; Portella, G.; Bowden, G.T.; Balmain, A.; Matrisian, L.M. A switch from stromal to tumor cell expression of stromelysin-1 mRNA associated with the conversion of squamous to spindle carcinomas during mouse skin tumor progression. Mol. Carcinog. 1994, 10, 207–215. [Google Scholar] [CrossRef]
- Kon, S.; Nakayama, Y.; Matsumoto, N.; Ito, K.; Kanayama, M.; Kimura, C.; Kouro, H.; Ashitomi, D.; Matsuda, T.; Uede, T. A novel cryptic binding motif, LRSKSRSFQVSDEQY, in the C-terminal fragment of MMP-3/7-cleaved osteopontin as a novel ligand for α9β1 integrin is involved in the anti-type II collagen antibody-induced arthritis. PLoS ONE 2014, 9, e116210. [Google Scholar] [CrossRef]
- Liaw, L.; Birk, D.E.; Ballas, C.B.; Whitsitt, J.S.; Davidson, J.M.; Hogan, B.L. Altered wound healing in mice lacking a functional osteopontin gene (spp1). J. Clin. Investig. 1998, 101, 1468–1478. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.L.; Heppner, K.J.; Rudolph, L.A.; Matrisian, L.M. The metalloproteinase matrilysin is preferentially expressed by epithelial cells in a tissue-restricted pattern in the mouse. Mol. Biol. Cell 1995, 6, 851–869. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Li, D.; Lv, X.H.; Hua, S.C.; Han, J.C.; Xu, F.; Li, X.D. Roles of osteopontin and matrix metalloproteinase-7 in occurrence, progression, and prognosis of nonsmall cell lung cancer. J. Res. Med. Sci. 2015, 20, 1138–1146. [Google Scholar] [CrossRef]
- Leitner, L.; Schuch, K.; Jürets, A.; Itariu, B.K.; Keck, M.; Grablowitz, V.; Aszmann, O.C.; Prager, G.; Staffler, G.; Zeyda, M.; et al. Immunological blockade of adipocyte inflammation caused by increased matrix metalloproteinase-cleaved osteopontin in obesity. Obesity 2015, 23, 779–785. [Google Scholar] [CrossRef]
- Slovacek, H.; Khanna, R.; Poredos, P.; Jezovnik, M.; Hoppensteadt, D.; Fareed, J.; Hopkinson, W. Interrelationship of Osteopontin, MMP-9 and ADAMTS4 in Patients With Osteoarthritis Undergoing Total Joint Arthroplasty. Clin. Appl. Thromb. Hemost. 2020, 26, 1076029620964864. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, M.L.; Zouein, F.A.; Tian, Y.; Padmanabhan Iyer, R.; de Castro Brás, L.E. Osteopontin is proteolytically processed by matrix metalloproteinase 9. Can. J. Physiol. Pharmcol. 2015, 93, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves DaSilva, A.; Liaw, L.; Yong, V.W. Cleavage of osteopontin by matrix metalloproteinase-12 modulates experimental autoimmune encephalomyelitis disease in C57BL/6 mice. Am. J. Pathol. 2010, 177, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.M.; Wilson, P.D.; Laskin, J.D.; Denhardt, D.T. Age and development-related changes in osteopontin and nitric oxide synthase mRNA levels in human kidney proximal tubule epithelial cells: Contrasting responses to hypoxia and reoxygenation. J. Cell Physiol. 1994, 160, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, M.W.; Cao, Y.; Moeller, B. Cycling hypoxia and free radicals regulate angiogenesis and radiotherapy response. Nat. Rev. Cancer 2008, 8, 425–437. [Google Scholar] [CrossRef] [Green Version]
- Christensen, B.; Kläning, E.; Nielsen, M.S.; Andersen, M.H.; Sørensen, E.S. C-terminal modification of osteopontin inhibits interaction with the αVβ3-integrin. J. Biol. Chem. 2012, 287, 3788–3797. [Google Scholar] [CrossRef] [Green Version]
- Bai, G.; Motoda, H.; Ozuru, R.; Chagan-Yasutan, H.; Hattori, T.; Matsuba, T. Synthesis of a Cleaved Form of Osteopontin by THP-1 Cells and Its Alteration by Phorbol 12-Myristate 13-Acetate and BCG Infection. Int. J. Mol. Sci. 2018, 19, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohro, T.; Tanaka, T.; Murakami, T.; Wada, Y.; Aburatani, H.; Hamakubo, T.; Kodama, T. A comparison of differences in the gene expression profiles of phorbol 12-myristate 13-acetate differentiated THP-1 cells and human monocyte-derived macrophage. J. Atheroscler Thromb. 2004, 11, 88–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Zeng, L.; Liu, Z.; Ke, X.; Lei, L.; Li, G. MicroRNA-206 regulates the secretion of inflammatory cytokines and MMP9 expression by targeting TIMP3 in Mycobacterium tuberculosis-infected THP-1 human macrophages. Biochem. Biophys. Res. Commun. 2016, 477, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Marrero, C.A.; Stewart, J.; Shafer, W.M.; Roman, J. The down-regulation of cathepsin G in THP-1 monocytes after infection with Mycobacterium tuberculosis is associated with increased intracellular survival of bacilli. Infect. Immun. 2004, 72, 5712–5721. [Google Scholar] [CrossRef] [Green Version]
- Franchini, M.; Mannucci, P.M. Thrombin and cancer: From molecular basis to therapeutic implications. Semin. Thromb. Hemost. 2012, 38, 95–101. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Shao, Z.; Sharif, S.; Du, X.Y.; Myles, T.; Merchant, M.; Harsh, G.; Glantz, M.; Recht, L.; Morser, J.; et al. Thrombin-cleaved fragments of osteopontin are overexpressed in malignant glial tumors and provide a molecular niche with survival advantage. J. Biol. Chem. 2013, 288, 3097–3111. [Google Scholar] [CrossRef] [Green Version]
- Beausoleil, M.S.; Schulze, E.B.; Goodale, D.; Postenka, C.O.; Allan, A.L. Deletion of the thrombin cleavage domain of osteopontin mediates breast cancer cell adhesion, proteolytic activity, tumorgenicity, and metastasis. BMC Cancer 2011, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, M.; Segawa, T.; Maeda, M.; Yoshida, T.; Sudo, A. Thrombin-cleaved osteopontin levels in synovial fluid correlate with disease severity of knee osteoarthritis. J. Rheumatol. 2011, 38, 129–134. [Google Scholar] [CrossRef]
- Beilin, O.; Karussis, D.M.; Korczyn, A.D.; Gurwitz, D.; Aronovich, R.; Hantai, D.; Grigoriadis, N.; Mizrachi-Kol, R.; Chapman, J. Increased thrombin inhibition in experimental autoimmune encephalomyelitis. J. Neurosci. Res. 2005, 79, 351–359. [Google Scholar] [CrossRef]
- Davalos, D.; Ryu, J.K.; Merlini, M.; Baeten, K.M.; Le Moan, N.; Petersen, M.A.; Deerinck, T.J.; Smirnoff, D.S.; Bedard, C.; Hakozaki, H.; et al. Fibrinogen-induced perivascular microglial clustering is required for the development of axonal damage in neuroinflammation. Nat. Commun. 2012, 3, 1227. [Google Scholar] [CrossRef] [Green Version]
- Davalos, D.; Baeten, K.M.; Whitney, M.A.; Mullins, E.S.; Friedman, B.; Olson, E.S.; Ryu, J.K.; Smirnoff, D.S.; Petersen, M.A.; Bedard, C.; et al. Early detection of thrombin activity in neuroinflammatory disease. Ann. Neurol. 2014, 75, 303–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.H.; Hwang, S.I.; Roy, D.B.; Lundgren, D.H.; Price, J.V.; Ousman, S.S.; Fernald, G.H.; Gerlitz, B.; Robinson, W.H.; Baranzini, S.E.; et al. Proteomic analysis of active multiple sclerosis lesions reveals therapeutic targets. Nature 2008, 451, 1076–1081. [Google Scholar] [CrossRef] [PubMed]
- Boggio, E.; Dianzani, C.; Gigliotti, C.L.; Soluri, M.F.; Clemente, N.; Cappellano, G.; Toth, E.; Raineri, D.; Ferrara, B.; Comi, C.; et al. Thrombin Cleavage of Osteopontin Modulates Its Activities in Human Cells In Vitro and Mouse Experimental Autoimmune Encephalomyelitis In Vivo. J. Immunol. Res. 2016, 2016, 9345495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, G.; Chen, J.; Wu, Z.; Huang, H.; Wang, L.; Liang, Y.; Zeng, P.; Yang, J.; Uede, T.; Diao, H. Thrombin cleavage of osteopontin controls activation of hepatic stellate cells and is essential for liver fibrogenesis. J. Cell Physiol. 2019, 234, 8988–8997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagori, K.; Yoshifuji, H.; Oku, T.; Sasaki, C.; Miyata, H.; Mori, K.P.; Nakajima, T.; Ohmura, K.; Kawabata, D.; Yukawa, N.; et al. Cleaved Form of Osteopontin in Urine as a Clinical Marker of Lupus Nephritis. PLoS ONE 2016, 11, e0167141. [Google Scholar] [CrossRef] [Green Version]
- Ashkar, S.; Weber, G.F.; Panoutsakopoulou, V.; Sanchirico, M.E.; Jansson, M.; Zawaideh, S.; Rittling, S.R.; Denhardt, D.T.; Glimcher, M.J.; Cantor, H. Eta-1 (osteopontin): An early component of type-1 (cell-mediated) immunity. Science 2000, 287, 860–864. [Google Scholar] [CrossRef]
- Koguchi, Y.; Kawakami, K.; Kon, S.; Segawa, T.; Maeda, M.; Uede, T.; Saito, A. Penicillium marneffei causes osteopontin-mediated production of interleukin-12 by peripheral blood mononuclear cells. Infect. Immun. 2002, 70, 1042–1048. [Google Scholar] [CrossRef] [Green Version]
- Renkl, A.C.; Wussler, J.; Ahrens, T.; Thoma, K.; Kon, S.; Uede, T.; Martin, S.F.; Simon, J.C.; Weiss, J.M. Osteopontin functionally activates dendritic cells and induces their differentiation toward a Th1-polarizing phenotype. Blood 2005, 106, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Maeno, Y.; Nakazawa, S.; Yamamoto, N.; Shinzato, M.; Nagashima, S.; Tanaka, K.; Sasaki, J.; Rittling, S.R.; Denhardt, D.T.; Uede, T.; et al. Osteopontin participates in Th1-mediated host resistance against nonlethal malaria parasite Plasmodium chabaudi chabaudi infection in mice. Infect. Immun. 2006, 74, 2423–2427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trinchieri, G. The two faces of interleukin 12: A pro-inflammatory cytokine and a key immunoregulatory molecule produced by antigen-presenting cells. Ciba Found Symp. 1995, 195, 203–214, discussion 214–220. [Google Scholar] [CrossRef]
- Abel, B.; Freigang, S.; Bachmann, M.F.; Boschert, U.; Kopf, M. Osteopontin is not required for the development of Th1 responses and viral immunity. J. Immunol. 2005, 175, 6006–6013. [Google Scholar] [CrossRef] [Green Version]
- Begum, M.D.; Umemura, M.; Kon, S.; Yahagi, A.; Hamada, S.; Oshiro, K.; Gotoh, K.; Nishizono, A.; Uede, T.; Matsuzaki, G. Suppression of the bacterial antigen-specific T cell response and the dendritic cell migration to the lymph nodes by osteopontin. Microbiol. Immunol. 2007, 51, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Santamaría, M.H.; Corral, R.S. Osteopontin-dependent regulation of Th1 and Th17 cytokine responses in Trypanosoma cruzi-infected C57BL/6 mice. Cytokine 2013, 61, 491–498. [Google Scholar] [CrossRef]
- Brown, A.; Islam, T.; Adams, R.; Nerle, S.; Kamara, M.; Eger, C.; Marder, K.; Cohen, B.; Schifitto, G.; McArthur, J.C.; et al. Osteopontin enhances HIV replication and is increased in the brain and cerebrospinal fluid of HIV-infected individuals. J. Neurovirol. 2011, 17, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, J.; Sarkar-Dutta, M.; McRae, S.; Ramachandran, A.; Kumar, B.; Waris, G. Osteopontin Regulates Hepatitis C Virus (HCV) Replication and Assembly by Interacting with HCV Proteins and Lipid Droplets and by Binding to Receptors αVβ3 and CD44. J. Virol. 2018, 92, e02116-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gessain, A.; Cassar, O. Epidemiological Aspects and World Distribution of HTLV-1 Infection. Front. Microbiol. 2012, 3, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducasa, N.; Grasso, D.; Benencio, P.; Papademetrio, D.L.; Biglione, M.; Kashanchi, F.; Berini, C.; Garcia, M.N. Autophagy in Human T-Cell Leukemia Virus Type 1 (HTLV-1) Induced Leukemia. Front. Oncol. 2021, 11, 641269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yamada, O.; Matsushita, Y.; Chagan-Yasutan, H.; Hattori, T. Transactivation of human osteopontin promoter by human T-cell leukemia virus type 1-encoded Tax protein. Leuk. Res. 2010, 34, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yamada, O.; Kida, S.; Matsushita, Y.; Yamaoka, S.; Chagan-Yasutan, H.; Hattori, T. Identification of CD44 as a downstream target of noncanonical NF-κB pathway activated by human T-cell leukemia virus type 1-encoded Tax protein. Virology 2011, 413, 244–252. [Google Scholar] [CrossRef]
- Chagan-Yasutan, H.; Tsukasaki, K.; Takahashi, Y.; Oguma, S.; Harigae, H.; Ishii, N.; Zhang, J.; Fukumoto, M.; Hattori, T. Involvement of osteopontin and its signaling molecule CD44 in clinicopathological features of adult T cell leukemia. Leuk. Res. 2011, 35, 1484–1490. [Google Scholar] [CrossRef]
- Liersch, R.; Gerss, J.; Schliemann, C.; Bayer, M.; Schwöppe, C.; Biermann, C.; Appelmann, I.; Kessler, T.; Löwenberg, B.; Büchner, T.; et al. Osteopontin is a prognostic factor for survival of acute myeloid leukemia patients. Blood 2012, 119, 5215–5220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moorman, H.R.; Poschel, D.; Klement, J.D.; Lu, C.; Redd, P.S.; Liu, K. Osteopontin: A Key Regulator of Tumor Progression and Immunomodulation. Cancers 2020, 12, 3379. [Google Scholar] [CrossRef]
- Nakachi, S.; Nakazato, T.; Ishikawa, C.; Kimura, R.; Mann, D.A.; Senba, M.; Masuzaki, H.; Mori, N. Human T-cell leukemia virus type 1 tax transactivates the matrix metalloproteinase 7 gene via JunD/AP-1 signaling. Biochim. Biophys. Acta 2011, 1813, 731–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koguchi, Y.; Kawakami, K.; Uezu, K.; Fukushima, K.; Kon, S.; Maeda, M.; Nakamoto, A.; Owan, I.; Kuba, M.; Kudeken, N.; et al. High plasma osteopontin level and its relationship with interleukin-12-mediated type 1 T helper cell response in tuberculosis. Am. J. Respir. Crit. Care Med. 2003, 167, 1355–1359. [Google Scholar] [CrossRef]
- Shiratori, B.; Leano, S.; Nakajima, C.; Chagan-Yasutan, H.; Niki, T.; Ashino, Y.; Suzuki, Y.; Telan, E.; Hattori, T. Elevated OPN, IP-10, and neutrophilia in loop-mediated isothermal amplification confirmed tuberculosis patients. Mediat. Inflamm. 2014, 2014, 513263. [Google Scholar] [CrossRef] [PubMed]
- Padilla, S.T.; Niki, T.; Furushima, D.; Bai, G.; Chagan-Yasutan, H.; Telan, E.F.; Tactacan-Abrenica, R.J.; Maeda, Y.; Solante, R.; Hattori, T. Plasma Levels of a Cleaved Form of Galectin-9 Are the Most Sensitive Biomarkers of Acquired Immune Deficiency Syndrome and Tuberculosis Coinfection. Biomolecules 2020, 10, 1495. [Google Scholar] [CrossRef] [PubMed]
- Diao, H.; Liu, X.; Wu, Z.; Kang, L.; Cui, G.; Morimoto, J.; Denhardt, D.T.; Rittling, S.; Iwakura, Y.; Uede, T.; et al. Osteopontin regulates interleukin-17 production in hepatitis. Cytokine 2012, 60, 129–137. [Google Scholar] [CrossRef]
- Liu, L.; Lu, J.; Ye, C.; Lin, L.; Zheng, S.; Zhang, H.; Lan, Q.; Xue, Y. Serum osteopontin is a predictor of prognosis for HBV-associated acute-on-chronic liver failure. Biomed. Rep. 2018, 8, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Li, T.; Wang, Y.; Pan, Y.; Ning, H.; Hui, X.; Xie, H.; Wang, J.; Han, Y.; Liu, Z.; et al. Elevated plasma osteopontin level is predictive of cirrhosis in patients with hepatitis B infection. Int. J. Clin. Pract. 2008, 62, 1056–1062. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Salles, T.; Misra, S.; Stepien, M.; Plymoth, A.; Muller, D.; Overvad, K.; Olsen, A.; Tjonneland, A.; Baglietto, L.; Severi, G.; et al. Circulating Osteopontin and Prediction of Hepatocellular Carcinoma Development in a Large European Population. Cancer Prev. Res. 2016, 9, 758–765. [Google Scholar] [CrossRef] [Green Version]
- Chagan-Yasutan, H.; Hanan, F.; Niki, T.; Bai, G.; Ashino, Y.; Egawa, S.; Telan, E.F.O.; Hattori, T. Plasma Osteopontin Levels is Associated with Biochemical Markers of Kidney Injury in Patients with Leptospirosis. Diagnostics 2020, 10, 439. [Google Scholar] [CrossRef]
- Van der Windt, G.J.; Wiersinga, W.J.; Wieland, C.W.; Tjia, I.C.; Day, N.P.; Peacock, S.J.; Florquin, S.; van der Poll, T. Osteopontin impairs host defense during established gram-negative sepsis caused by Burkholderia pseudomallei (melioidosis). PLoS Negl. Trop. Dis. 2010, 4, e806. [Google Scholar] [CrossRef]
- Pereira, T.A.; Syn, W.K.; Amancio, F.F.; Cunha, P.H.; Caporali, J.F.; Trindade, G.V.; Santos, E.T.; Souza, M.M.; Andrade, Z.A.; Witek, R.P.; et al. Osteopontin Is Upregulated in Human and Murine Acute Schistosomiasis Mansoni. PLoS Negl. Trop. Dis. 2016, 10, e0005057. [Google Scholar] [CrossRef]
- Shete, A.; Bichare, S.; Pujari, V.; Virkar, R.; Thakar, M.; Ghate, M.; Patil, S.; Vyakarnam, A.; Gangakhedkar, R.; Bai, G.; et al. Elevated Levels of Galectin-9 but Not Osteopontin in HIV and Tuberculosis Infections Indicate Their Roles in Detecting MTB Infection in HIV Infected Individuals. Front. Microbiol. 2020, 11, 1685. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.; Furushima, D.; Niki, T.; Matsuba, T.; Maeda, Y.; Takahashi, A.; Hattori, T.; Ashino, Y. High Levels of the Cleaved Form of Galectin-9 and Osteopontin in the Plasma Are Associated with Inflammatory Markers Tha. Int. J. Mol. Sci. 2021, 22, 4978. [Google Scholar] [CrossRef] [PubMed]
- Varım, C.; Demirci, T.; Cengiz, H.; Hacıbekiroğlu, İ.; Tuncer, F.B.; Çokluk, E.; Toptan, H.; Karabay, O.; Yıldırım, İ. Relationship between serum osteopontin levels and the severity of COVID-19 infection. Wien. Klin. Wochenschr. 2021, 133, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Tiberti, N.; Hainard, A.; Lejon, V.; Robin, X.; Ngoyi, D.M.; Turck, N.; Matovu, E.; Enyaru, J.; Ndung’u, J.M.; Scherl, A.; et al. Discovery and verification of osteopontin and Beta-2-microglobulin as promising markers for staging human African trypanosomiasis. Mol. Cell Proteom. 2010, 9, 2783–2795. [Google Scholar] [CrossRef] [Green Version]
- Hasibuan, F.M.; Shiratori, B.; Senoputra, M.A.; Chagan-Yasutan, H.; Koesoemadinata, R.C.; Apriani, L.; Takahashi, Y.; Niki, T.; Alisjahbana, B.; Hattori, T. Evaluation of matricellular proteins in systemic and local immune response to Mycobacterium tuberculosis infection. Microbiol. Immunol. 2015, 59, 623–632. [Google Scholar] [CrossRef]
- Nau, G.J.; Liaw, L.; Chupp, G.L.; Berman, J.S.; Hogan, B.L.; Young, R.A. Attenuated host resistance against Mycobacterium bovis BCG infection in mice lacking osteopontin. Infect. Immun. 1999, 67, 4223–4230. [Google Scholar] [CrossRef] [Green Version]
- Van der Windt, G.J.; Wieland, C.W.; Wiersinga, W.J.; Florquin, S.; van der Poll, T. Osteopontin is not crucial to protective immunity during murine tuberculosis. Immunology 2009, 128, e766–e776. [Google Scholar] [CrossRef]
- Ridruechai, C.; Sakurada, S.; Yanai, H.; Yamada, N.; Kantipong, P.; Piyaworawong, S.; Dhepakson, P.; Khusmith, S.; Keicho, N. Association between circulating full-length osteopontin and IFN-gamma with disease status of tuberculosis and response to successful treatment. Southeast Asian J. Trop. Med. Public Health 2011, 42, 876–889. [Google Scholar] [PubMed]
- Shiratori, B.; Zhao, J.; Okumura, M.; Chagan-Yasutan, H.; Yanai, H.; Mizuno, K.; Yoshiyama, T.; Idei, T.; Ashino, Y.; Nakajima, C.; et al. Immunological Roles of Elevated Plasma Levels of Matricellular Proteins in Japanese Patients with Pulmonary Tuberculosis. Int. J. Mol. Sci. 2016, 18, 19. [Google Scholar] [CrossRef]
- Ragno, S.; Romano, M.; Howell, S.; Pappin, D.J.; Jenner, P.J.; Colston, M.J. Changes in gene expression in macrophages infected with Mycobacterium tuberculosis: A combined transcriptomic and proteomic approach. Immunology 2001, 104, 99–108. [Google Scholar] [CrossRef] [Green Version]
- Kathamuthu, G.R.; Kumar, N.P.; Moideen, K.; Nair, D.; Banurekha, V.V.; Sridhar, R.; Baskaran, D.; Babu, S. Matrix Metalloproteinases and Tissue Inhibitors of Metalloproteinases Are Potential Biomarkers of Pulmonary and Extra-Pulmonary Tuberculosis. Front. Immunol. 2020, 11, 419. [Google Scholar] [CrossRef] [Green Version]
- Le, Y.; Cao, W.; Zhou, L.; Fan, X.; Liu, Q.; Liu, F.; Gai, X.; Chang, C.; Xiong, J.; Rao, Y.; et al. Infection of Mycobacterium tuberculosis Promotes Both M1/M2 Polarization and MMP Production in Cigarette Smoke-Exposed Macrophages. Front. Immunol. 2020, 11, 1902. [Google Scholar] [CrossRef]
- Burdo, T.H.; Ellis, R.J.; Fox, H.S. Osteopontin is increased in HIV-associated dementia. J. Infect. Dis. 2008, 198, 715–722. [Google Scholar] [CrossRef]
- Chagan-Yasutan, H.; Saitoh, H.; Ashino, Y.; Arikawa, T.; Hirashima, M.; Li, S.; Usuzawa, M.; Oguma, S.; EF, O.T.; Obi, C.L.; et al. Persistent elevation of plasma osteopontin levels in HIV patients despite highly active antiretroviral therapy. Tohoku J. Exp. Med. 2009, 218, 285–292. [Google Scholar] [CrossRef] [Green Version]
- Richert, Q.; Trajtman, A.; Arroyave, L.; Toews, J.; Becker, M.; Kasper, K.; McLaren, P.; Rueda, Z.; Keynan, Y. Systemic inflammation before and after antiretroviral therapy initiation as a predictor of immune response among HIV-infected individuals in Manitoba. Cytokine 2017, 91, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Shepherd, N.; Lan, J.; Li, W.; Rane, S.; Gupta, S.K.; Zhang, S.; Dong, J.; Yu, Q. MMPs/TIMPs imbalances in the peripheral blood and cerebrospinal fluid are associated with the pathogenesis of HIV-1-associated neurocognitive disorders. Brain Behav. Immun. 2017, 65, 161–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozzelli, P.L.; Caccavano, A.; Avdoshina, V.; Mocchetti, I.; Wu, J.Y.; Conant, K. Increased matrix metalloproteinase levels and perineuronal net proteolysis in the HIV-infected brain; relevance to altered neuronal population dynamics. Exp. Neurol. 2020, 323, 113077. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, F.J.; Boucher, T.; Liang, S.; Brown, A.M. Osteopontin and Integrin Mediated Modulation of Post-Synapses in HIV Envelope Glycoprotein Exposed Hippocampal Neurons. Brain Sci. 2020, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Vo Quang, E.; Shimakawa, Y.; Nahon, P. Epidemiological projections of viral-induced hepatocellular carcinoma in the perspective of WHO global hepatitis elimination. Liver Int. 2021, 41, 915–927. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pan, X.; Huang, Z.; Weber, G.F.; Zhang, G. Osteopontin enhances the expression and activity of MMP-2 via the SDF-1/CXCR4 axis in hepatocellular carcinoma cell lines. PLoS ONE 2011, 6, e23831. [Google Scholar] [CrossRef]
- Yang, S.; Wang, L.; Pan, W.; Bayer, W.; Thoens, C.; Heim, K.; Dittmer, U.; Timm, J.; Wang, Q.; Yu, Q.; et al. MMP2/MMP9-mediated CD100 shedding is crucial for inducing intrahepatic anti-HBV CD8 T cell responses and HBV clearance. J. Hepatol. 2019, 71, 685–698. [Google Scholar] [CrossRef]
- Sumi, A.; Telan, E.F.; Chagan-Yasutan, H.; Piolo, M.B.; Hattori, T.; Kobayashi, N. Effect of temperature, relative humidity and rainfall on dengue fever and leptospirosis infections in Manila, the Philippines. Epidemiol. Infect. 2017, 145, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Excler, J.L.; Saville, M.; Berkley, S.; Kim, J.H. Vaccine development for emerging infectious diseases. Nat. Med. 2021, 27, 591–600. [Google Scholar] [CrossRef] [PubMed]
- WHO. Dengue: Guidelines for Diagnosis, Treatment, Prevention and Control; WHO: Geneva, Switzerland, 2009. [Google Scholar]
- WHO. Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control, 2nd ed.; WHO: Geneva, Switzerland, 1997. [Google Scholar]
- Kubelka, C.F.; Azeredo, E.L.; Gandini, M.; Oliveira-Pinto, L.M.; Barbosa, L.S.; Damasco, P.V.; Avila, C.A.; Motta-Castro, A.R.; Cunha, R.V.; Cruz, O.G. Metalloproteinases are produced during dengue fever and MMP9 is associated with severity. J. Infect. 2010, 61, 501–505. [Google Scholar] [CrossRef]
- Her, Z.; Kam, Y.W.; Gan, V.C.; Lee, B.; Thein, T.L.; Tan, J.J.; Lee, L.K.; Fink, K.; Lye, D.C.; Rénia, L.; et al. Severity of Plasma Leakage Is Associated With High Levels of Interferon γ-Inducible Protein 10, Hepatocyte Growth Factor, Matrix Metalloproteinase 2 (MMP-2), and MMP-9 During Dengue Virus Infection. J. Infect. Dis. 2017, 215, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Niranjan, R.; Sumitha, M.K.; Sankari, T.; Muthukumaravel, S.; Jambulingam, P. Nonstructural protein-1 (NS1) of dengue virus type-2 differentially stimulate expressions of matrix metalloproteinases in monocytes: Protective effect of paracetamol. Int. Immunopharmacol. 2019, 73, 270–279. [Google Scholar] [CrossRef]
- Chagan-Yasutan, H.; Ndhlovu, L.C.; Lacuesta, T.L.; Kubo, T.; Leano, P.S.; Niki, T.; Oguma, S.; Morita, K.; Chew, G.M.; Barbour, J.D.; et al. Galectin-9 plasma levels reflect adverse hematological and immunological features in acute dengue virus infection. J. Clin. Virol. 2013, 58, 635–640. [Google Scholar] [CrossRef] [Green Version]
- WHO. Human Leptospirosis: Guidance for Diagnosis, Surveillance and Control; WHO: Geneva, Switzerland, 2003. [Google Scholar]
- Torgerson, P.R.; Hagan, J.E.; Costa, F.; Calcagno, J.; Kane, M.; Martinez-Silveira, M.S.; Goris, M.G.; Stein, C.; Ko, A.I.; Abela-Ridder, B. Global Burden of Leptospirosis: Estimated in Terms of Disability Adjusted Life Years. PLoS Negl. Trop. Dis. 2015, 9, e0004122. [Google Scholar] [CrossRef] [Green Version]
- Chagan-Yasutan, H.; Chen, Y.; Lacuesta, T.L.; Leano, P.S.; Iwasaki, H.; Hanan, F.; Taurustiati, D.; Ohmoto, Y.; Ashino, Y.; Saitoh, H.; et al. Urine Levels of Defensin α1 Reflect Kidney Injury in Leptospirosis Patients. Int. J. Mol. Sci. 2016, 17, 1637. [Google Scholar] [CrossRef] [Green Version]
- Vijayachari, P.; Sugunan, A.P.; Shriram, A.N. Leptospirosis: An emerging global public health problem. J. Biosci. 2008, 33, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Becquart, P.; Wauquier, N.; Nkoghe, D.; Ndjoyi-Mbiguino, A.; Padilla, C.; Souris, M.; Leroy, E.M. Acute dengue virus 2 infection in Gabonese patients is associated with an early innate immune response, including strong interferon alpha production. BMC Infect. Dis. 2010, 10, 356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki-Hozumi, H.; Chagan-Yasutan, H.; Ashino, Y.; Hattori, T. Blood Levels of Galectin-9, an Immuno-Regulating Molecule, Reflect the Severity for the Acute and Chronic Infectious Diseases. Biomolecules 2021, 11, 430. [Google Scholar] [CrossRef] [PubMed]
- Cagliero, J.; Villanueva, S.; Matsui, M. Leptospirosis Pathophysiology: Into the Storm of Cytokines. Front. Cell Infect. Microbiol. 2018, 8, 204. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.C.; Currie, B.J. Melioidosis: Epidemiology, pathophysiology, and management. Clin. Microbiol. Rev. 2005, 18, 383–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gassiep, I.; Armstrong, M.; Norton, R. Human Melioidosis. Clin. Microbiol. Rev. 2020, 33, e00006-19. [Google Scholar] [CrossRef] [PubMed]
- Conejero, L.; Potempa, K.; Graham, C.M.; Spink, N.; Blankley, S.; Salguero, F.J.; Pankla-Sranujit, R.; Khaenam, P.; Banchereau, J.F.; Pascual, V.; et al. The Blood Transcriptome of Experimental Melioidosis Reflects Disease Severity and Shows Considerable Similarity with the Human Disease. J. Immunol. 2015, 195, 3248–3261. [Google Scholar] [CrossRef]
- Kasozi, K.I.; Zirintunda, G.; Ssempijja, F.; Buyinza, B.; Alzahrani, K.J.; Matama, K.; Nakimbugwe, H.N.; Alkazmi, L.; Onanyang, D.; Bogere, P.; et al. Epidemiology of Trypanosomiasis in Wildlife-Implications for Humans at the Wildlife Interface in Africa. Front. Vet. Sci. 2021, 8, 621699. [Google Scholar] [CrossRef]
- Hainard, A.; Tiberti, N.; Robin, X.; Ngoyi, D.M.; Matovu, E.; Enyaru, J.C.; Müller, M.; Turck, N.; Ndung’u, J.M.; Lejon, V.; et al. Matrix metalloproteinase-9 and intercellular adhesion molecule 1 are powerful staging markers for human African trypanosomiasis. Trop. Med. Int. Health 2011, 16, 119–126. [Google Scholar] [CrossRef]
- Chitsulo, L.; Engels, D.; Montresor, A.; Savioli, L. The global status of schistosomiasis and its control. Acta Trop. 2000, 77, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Tsuji, M. From discovery to eradication of schistosomiasis in Japan: 1847–1996. Int. J. Parasitol. 1997, 27, 1465–1480. [Google Scholar] [CrossRef]
- Pereira, T.A.; Syn, W.K.; Machado, M.V.; Vidigal, P.V.; Resende, V.; Voieta, I.; Xie, G.; Otoni, A.; Souza, M.M.; Santos, E.T.; et al. Schistosome-induced cholangiocyte proliferation and osteopontin secretion correlate with fibrosis and portal hypertension in human and murine schistosomiasis mansoni. Clin. Sci. 2015, 129, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Chuah, C.; Jones, M.K.; Burke, M.L.; McManus, D.P.; Owen, H.C.; Gobert, G.N. Defining a pro-inflammatory neutrophil phenotype in response to schistosome eggs. Cell Microbiol. 2014, 16, 1666–1677. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19): A Review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef]
- Yan, G.; Lee, C.K.; Lam, L.T.M.; Yan, B.; Chua, Y.X.; Lim, A.Y.N.; Phang, K.F.; Kew, G.S.; Teng, H.; Ngai, C.H.; et al. Covert COVID-19 and false-positive dengue serology in Singapore. Lancet Infect. Dis. 2020, 20, 536. [Google Scholar] [CrossRef] [Green Version]
- Silvestre, O.M.; Costa, L.R.; Lopes, B.V.R.; Barbosa, M.R.; Botelho, K.K.P.; Albuquerque, K.L.C.; Souza, A.G.S.; Coelho, L.A.; de Oliveira, A.J.; Barantini, C.B.; et al. Previous dengue infection and mortality in COVID-19. Clin. Infect. Dis. 2020, ciaa1895. [Google Scholar] [CrossRef]
- Nicolete, V.C.; Rodrigues, P.T.; Johansen, I.C.; Corder, R.M.; Tonini, J.; Cardoso, M.A.; de Jesus, J.G.; Claro, I.M.; Faria, N.R.; Sabino, E.C.; et al. Interacting Epidemics in Amazonian Brazil: Prior Dengue Infection Associated with Increased COVID-19 Risk in a Population-Based Cohort Study. Clin. Infect. Dis. 2021, ciab410. [Google Scholar] [CrossRef]
- Niriella, M.A.; Ediriweera, D.S.; De Silva, A.P.; Premarathna, B.H.R.; Jayasinghe, S.; de Silva, H.J. Dengue and leptospirosis infection during the coronavirus 2019 outbreak in Sri Lanka. Trans. R. Soc. Trop. Med. Hyg. 2021, trab058. [Google Scholar] [CrossRef]
- Joubert, A.; Andry, F.; Bertolotti, A.; Accot, F.; Koumar, Y.; Legrand, F.; Poubeau, P.; Manaquin, R.; Gerardin, P.; Levin, C. Distinguishing non severe cases of dengue from COVID-19 in the context of co-epidemics: A cohort study in a SARS-CoV-2 testing center on Reunion island. PLoS Negl. Trop. Dis. 2021, 15, e0008879. [Google Scholar] [CrossRef]
- Adu-Agyeiwaah, Y.; Grant, M.B.; Obukhov, A.G. The Potential Role of Osteopontin and Furin in Worsening Disease Outcomes in COVID-19 Patients with Pre-Existing Diabetes. Cells 2020, 9, 2528. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Rosas, I.O.; Bräu, N.; Waters, M.; Go, R.C.; Hunter, B.D.; Bhagani, S.; Skiest, D.; Aziz, M.S.; Cooper, N.; Douglas, I.S.; et al. Tocilizumab in Hospitalized Patients with Severe Covid-19 Pneumonia. N. Engl. J. Med. 2021, 384, 1503–1516. [Google Scholar] [CrossRef] [PubMed]
- Klement, J.D.; Paschall, A.V.; Redd, P.S.; Ibrahim, M.L.; Lu, C.; Yang, D.; Celis, E.; Abrams, S.I.; Ozato, K.; Liu, K. An osteopontin/CD44 immune checkpoint controls CD8+ T cell activation and tumor immune evasion. J. Clin. Investig. 2018, 128, 5549–5560. [Google Scholar] [CrossRef]
- Ueland, T.; Holter, J.C.; Holten, A.R.; Müller, K.E.; Lind, A.; Bekken, G.K.; Dudman, S.; Aukrust, P.; Dyrhol-Riise, A.M.; Heggelund, L. Distinct and early increase in circulating MMP-9 in COVID-19 patients with respiratory failure. J. Infect. 2020, 81, e41–e43. [Google Scholar] [CrossRef] [PubMed]
- Abers, M.S.; Delmonte, O.M.; Ricotta, E.E.; Fintzi, J.; Fink, D.L.; de Jesus, A.A.A.; Zarember, K.A.; Alehashemi, S.; Oikonomou, V.; Desai, J.V.; et al. An immune-based biomarker signature is associated with mortality in COVID-19 patients. JCI Insight 2021, 6, e144455. [Google Scholar] [CrossRef]
- Stukalov, A.; Girault, V.; Grass, V.; Karayel, O.; Bergant, V.; Urban, C.; Haas, D.A.; Huang, Y.; Oubraham, L.; Wang, A.; et al. Multilevel proteomics reveals host perturbations by SARS-CoV-2 and SARS-CoV. Nature 2021, 594, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Ranucci, M.; Sitzia, C.; Baryshnikova, E.; Di Dedda, U.; Cardani, R.; Martelli, F.; Corsi Romanelli, M. Covid-19-Associated Coagulopathy: Biomarkers of Thrombin Generation and Fibrinolysis Leading the Outcome. J. Clin. Med. 2020, 9, 3487. [Google Scholar] [CrossRef] [PubMed]
- Aliter, K.F.; Al-Horani, R.A. Thrombin Inhibition by Argatroban: Potential Therapeutic Benefits in COVID-19. Cardiovasc. Drugs Ther. 2021, 35, 195–203. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
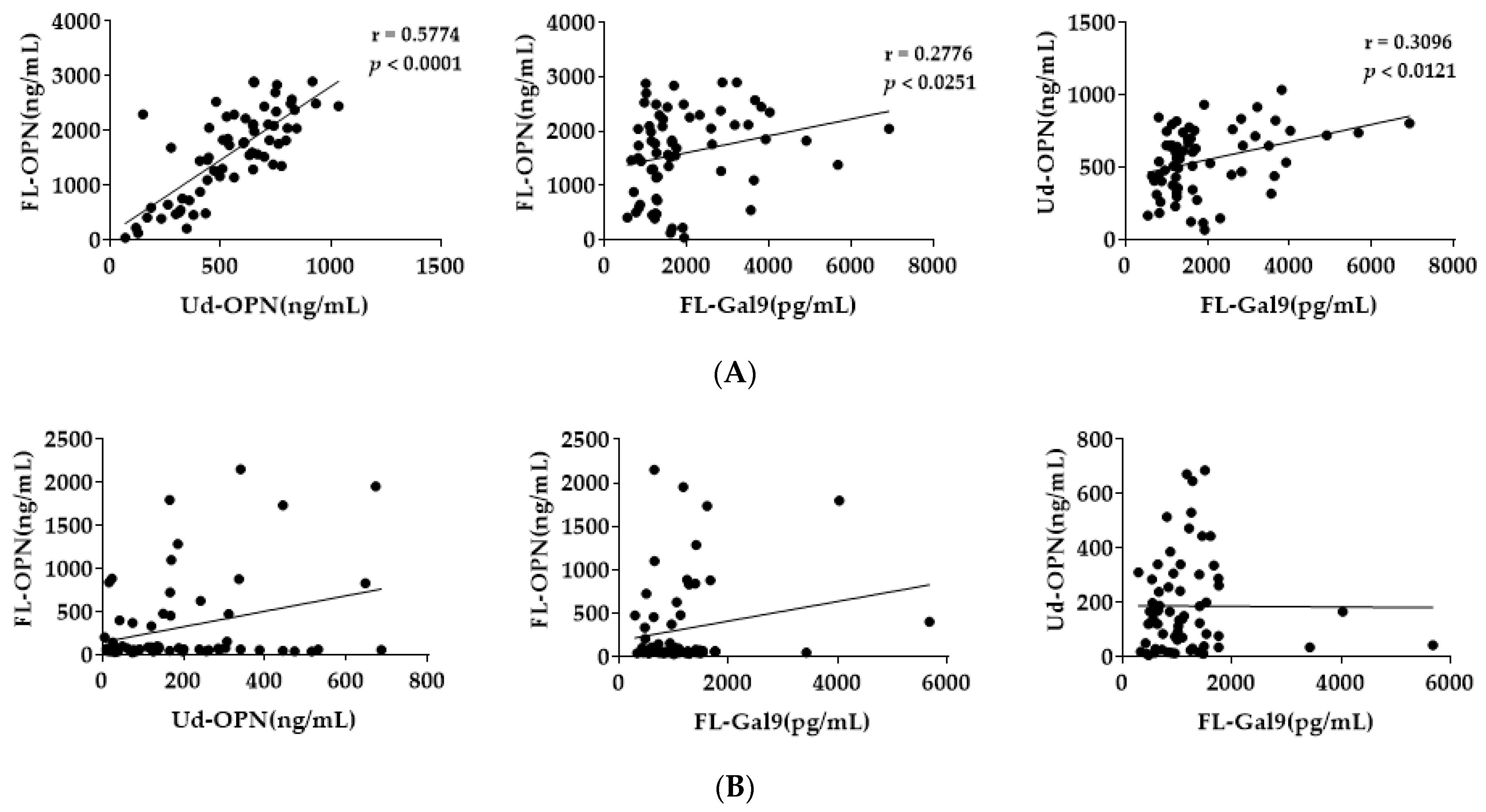{kind=link}
{kind=link}
| Disease | OPN [ng/mL] | p Value | Statistic | Statistical Analysis | Sample Type | ELISA | Ref. | ||
|---|---|---|---|---|---|---|---|---|---|
| Patients (n) | Control (n) | ||||||||
| ATL | Acute | 843 (n = 13) | <396 (n = 30) | <0.0001 | Median | Kruskal–Wallis test and two tail Dunn’s post test | Plasma | IBL | [100] |
| Lymphoma | 800 (n = 3) | <0.05 | |||||||
| Chronic | 318 (n = 9) | - | |||||||
| Smoldering | 259 (n = 2) | - | |||||||
| TB | 600 (n = 48) | 250 (n =34) | <0.0001 | Median | Fisher’s test | Plasma | IBL | [104] | |
| 159 (n = 37) | 69 (n =30) | <0.0001 | Median | Mann–Whitney U-test | Plasma | RDS | [105] | ||
| 666 (n = 49) | 129 (n = 30) | <0.0001 | Median | Kruskal–Wallis test and Mann–Whitney U-test | Plasma | IBL | [106] | ||
| 65.1 (n = 49) | 8.34 (n = 30) | <0.0001 | RDS | ||||||
| AIDS/TB | 810 (n = 33) | 129 (n = 30) | <0.0001 | Median | Kruskal–Wallis test and Mann–Whitney U-test | Plasma | IBL | [106] | |
| 103 (n = 49) | 8.34 (n = 30) | <0.0001 | RDS | ||||||
| AIDS | 661 (n = 24) | 129 (n = 30) | <0.0001 | Median | Kruskal–Wallis test and Mann–Whitney U-test | Plasma | IBL | [106] | |
| 123 (n = 24) | 8.34 (n = 30) | <0.0001 | RDS | ||||||
| Hepatitis virus infection | HBV | 560 * (n = 10) | 490 * (n = 20) | <0.01 | Mean | Student’s t-test | Serum | IBL | [107] |
| ACLF | 8491 (n = 54) | 3880 (n = 20) | 0.015 | Mean | Student’s t-test | Serum | RDS | [108] | |
| LC | 4.52 (n = 39) | 3.23 (n = 14) | <0.001 | Median | Mann–Whitney U-test | Plasma | RDS | [109] | |
| HCC | 67.4 (n = 100) | 53.7 (n = 194) | <0.001 | Median | Student’s paired t-test | Plasma | RDS | [110] | |
| Dengue | DF (n =53) | 25.951 | 2.814 (n = 30) | <0.0001 | Median | Dunn’s multiple comparison test | Plasma | IBL | [11] |
| 540 | 68 (n = 30) | <0.0001 | RDS | ||||||
| DHF (n = 12) | 27.55 | 2.814 (n = 30) | <0.0001 | IBL | |||||
| 692 | 68 (n = 30) | <0.0001 | RDS | ||||||
| Leptospirosis | 442 (n = 112) | 187.23 (n = 30) | <0.0001 | Median | Mann–Whitney U-test | Plasma | IBL | [111] | |
| Melioidosis | 7100 * (n = 33) | 100 * (n = 31) | <0.001 | Mean | Mann–Whitney U-test | Plasma | RDS | [112] | |
| Schistosomiasis | Acute | 80 * (n = 28) | 5 ¶* (n = 21) | <0.0001 | Median | Mann–Whitney U-test | Serum | RDS | [113] |
| Disease | OPN [ng/mL] | p Value | Statistic | Statistical Analysis | Sample Type | ELISA | Ref. | |
|---|---|---|---|---|---|---|---|---|
| ATL | Acute 843 (n = 13) | Chronic 318 (n = 9) | 0.0002 | Median | Mann–Whitney U-test | Plasma | IBL | [100] |
| Acute & Lymphoma 843 (n = 16) | Chronic & Smoldering 318 (n = 11) | <0.0001 | ||||||
| TB | Severe PTB 192 (n = 10) | Non-severe PTB 114 (n = 10) | <0.0001 | Median | Mann–Whitney U-test | Plasma | RDS | [114] |
| COVID-19 | CP 425 (n = 25) | CV 330 (n = 23) | <0.0146 | Median | Mann–Whitney U-test | Plasma | IBL | [115] |
| CP 63.3 (n = 13) | CV 42.3 (n = 9) | 0.0001 | RDS | |||||
| Clitically ill 13.75 (n = 40) | Non-critically ill 9.85 (n = 48) | 0.002 | Median | Mann–Whitney U-test | Serum | Elabscience | [116] | |
| Melioidosis | Non-survivors 8750 * (n = 14) | Survivors 6200 * (n = 19) | <0.05 | Mean | Mann–Whitney U-test | Plasma | RDS | [112] |
| Trypanosomiasis | Moderate neurological singns 550 * (n = 31) | Absence of neurological signs 100 * (n = 21) | <0.01 | Median | Mann–Whitney U-test | CSF | RDS | [117] |
| Severe neurological sings 950 * (n = 5) | <0.05 | Kruskal–Wallis test | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hattori, T.; Iwasaki-Hozumi, H.; Bai, G.; Chagan-Yasutan, H.; Shete, A.; Telan, E.F.; Takahashi, A.; Ashino, Y.; Matsuba, T. Both Full-Length and Protease-Cleaved Products of Osteopontin Are Elevated in Infectious Diseases. Biomedicines 2021, 9, 1006. https://doi.org/10.3390/biomedicines9081006
Hattori T, Iwasaki-Hozumi H, Bai G, Chagan-Yasutan H, Shete A, Telan EF, Takahashi A, Ashino Y, Matsuba T. Both Full-Length and Protease-Cleaved Products of Osteopontin Are Elevated in Infectious Diseases. Biomedicines. 2021; 9(8):1006. https://doi.org/10.3390/biomedicines9081006
Chicago/Turabian StyleHattori, Toshio, Hiroko Iwasaki-Hozumi, Gaowa Bai, Haorile Chagan-Yasutan, Ashwnini Shete, Elizabeth Freda Telan, Atsushi Takahashi, Yugo Ashino, and Takashi Matsuba. 2021. "Both Full-Length and Protease-Cleaved Products of Osteopontin Are Elevated in Infectious Diseases" Biomedicines 9, no. 8: 1006. https://doi.org/10.3390/biomedicines9081006
APA StyleHattori, T., Iwasaki-Hozumi, H., Bai, G., Chagan-Yasutan, H., Shete, A., Telan, E. F., Takahashi, A., Ashino, Y., & Matsuba, T. (2021). Both Full-Length and Protease-Cleaved Products of Osteopontin Are Elevated in Infectious Diseases. Biomedicines, 9(8), 1006. https://doi.org/10.3390/biomedicines9081006