
Translation of ABCE1 Is Tightly Regulated by Upstream Open Reading Frames in Human Colorectal Cells

Abstract
:1. Introduction
2. Experimental Section
2.1. Reporter Vectors
2.2. Cell Culture
2.3. Cell Transfections and Treatments
2.4. Luminometry Assays
2.5. Western Blotting
2.6. RNA Extraction and cDNA Synthesis
2.7. RT-qPCR
2.8. Semi-Quantitative RT-PCR
2.9. Statistical Analysis
3. Results
3.1. The Human ABCE1 5′-Leader Sequence Contains Five AUG and Five Non-AUG uORFs
3.2. The ABCE1 5′-Leader Sequence Has Translation-Inhibitory AUG uORFs
3.3. The ABCE1 AUG uORFs Have the Potential to Be Translated
3.4. ABCE1 uORF3 and uORF5 Are Competent Translational Repressors, Greatly Regulated by uORF1 and to a Less Extent by uORF2
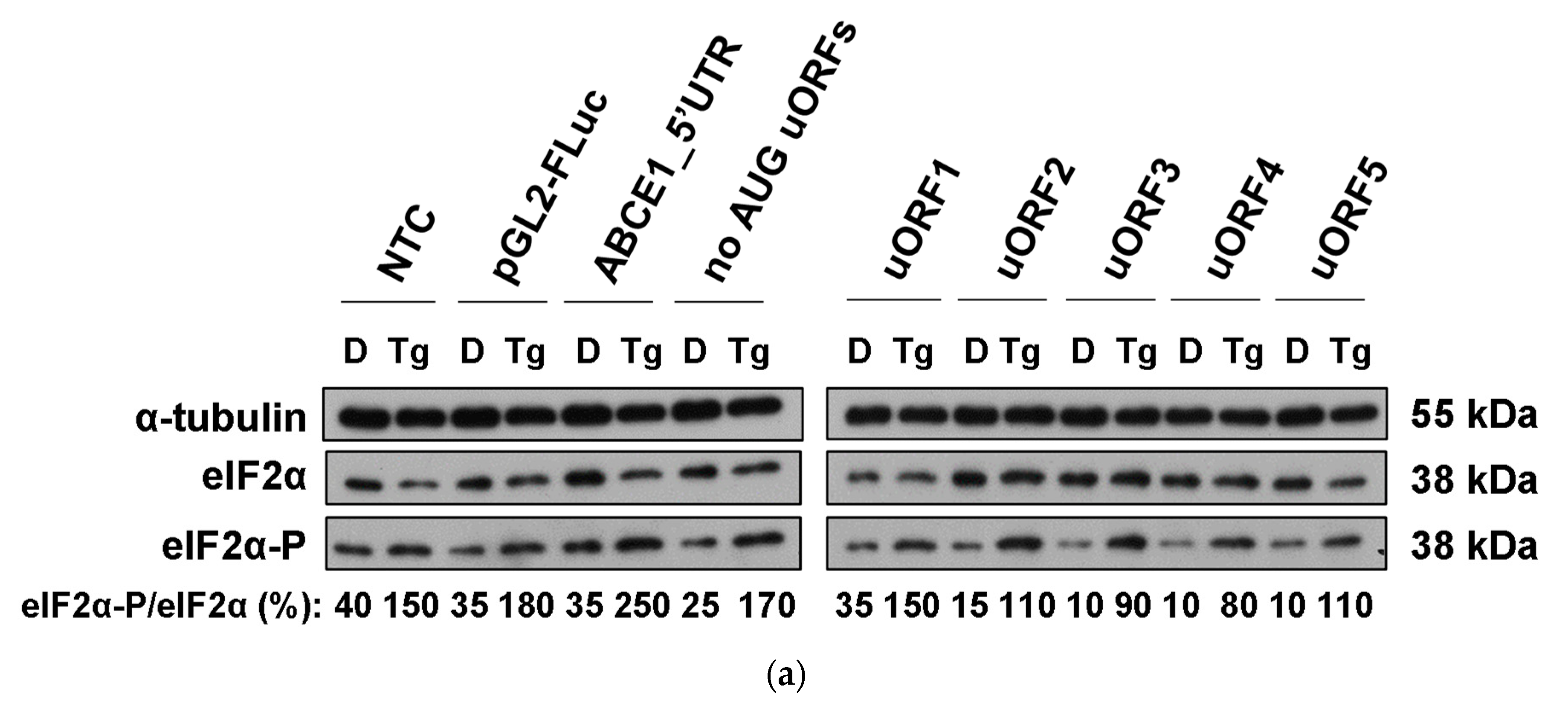
3.5. Although ABCE1 AUG uORFs Mediate Efficient Translational Repression during Thapsigargin-Induced ER Stress, Some Translational Derepression Occurs
3.6. The ABCE1 uORF-Mediated Translational Regulation Does Not Seem to Have an Oncogenic Role in Colorectal Cancer Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Young, S.K.; Wek, R.C. Upstream open reading frames differentially regulate gene-specific translation in the integrated stress response. J. Biol. Chem. 2016, 291, 16927–16935. [Google Scholar] [CrossRef] [Green Version]
- Sonenberg, N.; Hinnebusch, A.G. Regulation of translation initiation in eukaryotes: Mechanisms and biological targets. Cell 2009, 136, 731–745. [Google Scholar] [CrossRef] [Green Version]
- Roux, P.P.; Topisirovic, I. Signaling pathways involved in the regulation of mRNA translation. Mol. Cell. Biol. 2018, 38, e00070-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, C.; Peixeiro, I.; Romão, L. Gene expression regulation by upstream open reading frames and human disease. PLoS Genet. 2013, 9, e1003529. [Google Scholar] [CrossRef] [Green Version]
- Leppek, K.; Das, R.; Barna, M. Functional 5′ UTR mRNA structures in eukaryotic translation regulation and how to find them. Nat. Rev. Mol. Cell Biol. 2018, 19, 158–174. [Google Scholar] [CrossRef] [PubMed]
- Ingolia, N.T.; Ghaemmaghami, S.; Newman, J.R.S.; Weissman, J.S. Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling. Science 2009, 324, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Ingolia, N.T.; Lareau, L.F.; Weissman, J.S. Ribosome profiling of mouse embryonic stem cells reveals the complexity of mammalian proteomes. Cell 2011, 147, 789–802. [Google Scholar] [CrossRef] [Green Version]
- McGillivray, P.; Ault, R.; Pawashe, M.; Kitchen, R.; Balasubramanian, S.; Gerstein, M. A comprehensive catalog of predicted functional upstream open reading frames in humans. Nucleic Acids Res. 2018, 46, 3326–3338. [Google Scholar] [CrossRef]
- Calvo, S.E.; Pagliarini, D.J.; Mootha, V.K. Upstream open reading frames cause widespread reduction of protein expression and are polymorphic among humans. Proc. Natl. Acad. Sci. USA 2009, 106, 7507–7512. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Liang, Y.; Yu, Q.; Hu, L.; Li, H.; Zhang, Z.; Xu, X. Analysis of human upstream open reading frames and impact on gene expression. Hum. Genet. 2015, 134, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.R.; Geballe, A.P. Upstream open reading frames as regulators of mRNA translation. Mol. Cell. Biol. 2000, 20, 8635–8642. [Google Scholar] [CrossRef] [Green Version]
- Sajjanar, B.; Deb, R.; Raina, S.K.; Pawar, S.; Brahmane, M.P.; Nirmale, A.V.; Kurade, N.P.; Manjunathareddy, G.B.; Bal, S.K.; Singh, N.P. Untranslated regions (UTRs) orchestrate translation reprogramming in cellular stress responses. J. Therm. Biol. 2017, 65, 69–75. [Google Scholar] [CrossRef]
- Wethmar, K. The regulatory potential of upstream open reading frames in eukaryotic gene expression. Wiley Interdiscip. Rev. 2014, 5, 765–768. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tarn, W. uORF-mediated translational control: Recently elucidated mechanisms and implications in cancer. RNA Biol. 2019, 16, 1327–1338. [Google Scholar] [CrossRef]
- Mendell, J.T.; Sharifi, N.A.; Meyers, J.L.; Martinez-Murillo, F.; Dietz, H.C. Nonsense surveillance regulates expression of diverse classes of mammalian transcripts and mutes genomic noise. Nat. Genet. 2004, 36, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Gardner, L.B. Hypoxic inhibition of nonsense-mediated RNA decay regulates gene expression and the integrated stress response. Mol. Cell Biol. 2008, 28, 3729–3741. [Google Scholar] [CrossRef] [Green Version]
- Rebbapragada, I.; Lykke-Andersen, J. Execution of nonsense-mediated mRNA decay: What defines a substrate? Curr. Opin. Cell Biol. 2009, 21, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Somers, J.; Pöyry, T.; Willis, A.E. A perspective on mammalian upstream open reading frame function. Int. J. Biochem. Cell Biol. 2013, 45, 1690–1700. [Google Scholar] [CrossRef] [Green Version]
- Andrews, S.J.; Rothnagel, J.A. Emerging evidence for functional peptides encoded by short open reading frames. Nat. Rev. Genet. 2014, 15, 193–204. [Google Scholar] [CrossRef]
- Raney, A.; Law, G.L.; Mize, G.J.; Morris, D.R. Regulated translation termination at the upstream open reading frame in s-adenosylmethionine decarboxylase mRNA. J. Biol. Chem. 2002, 277, 5988–5994. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, L.C.; Goodwin, B.L.; Solomonson, L.P.; Eichler, D.C. Regulation of endothelial argininosuccinate synthase expression and NO production by an upstream open reading frame. J. Biol. Chem. 2005, 280, 24252–24260. [Google Scholar] [CrossRef] [Green Version]
- Kozak, M. Constraints on reinitiation of translation in mammals. Nucleic Acids Res. 2001, 29, 5226–5232. [Google Scholar] [CrossRef] [Green Version]
- Child, S.J.; Miller, M.K.; Geballe, A.P. Translational control by an upstream open reading frame in the HER-2/neu transcript. J. Biol. Chem. 1999, 274, 24335–24341. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, R.; Romão, L. Translational regulation by upstream open reading frames and its relevance to human genetic disease. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; Volume 1. [Google Scholar] [CrossRef]
- Chew, G.; Pauli, A.; Schier, A.F. Conservation of uORF repressiveness and sequence features in mouse, human and zebrafish. Nat. Commun. 2016, 7, 11663. [Google Scholar] [CrossRef] [PubMed]
- Kozak, M. Downstream secondary structure facilitates recognition of initiator codons by eukaryotic ribosomes. Proc. Natl. Acad. Sci. USA 1990, 87, 8301–8305. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wan, J.; Shu, X.E.; Mao, Y.; Liu, X.; Yuan, X.; Zhang, X.; Hess, M.E.; Brüning, J.C.; Qian, S. N6-methyladenosine guides mRNA alternative translation during integrated stress response. Mol. Cell 2018, 69, 636–647. [Google Scholar] [CrossRef] [Green Version]
- Hir, H.; Saulière, J.; Wang, Z. The exon junction complex as a node of post-transcriptional networks. Nat. Rev. Mol. Cell Biol. 2016, 17, 41–54. [Google Scholar] [CrossRef]
- Kochetov, A.V.; Prayaga, P.D.; Volkova, O.A.; Sankararamakrishnan, R. Hidden coding potential of eukaryotic genomes: nonAUG started ORFs. J. Biomol. Struct. Dyn. 2013, 31, 103–114. [Google Scholar] [CrossRef]
- Schwab, S.R.; Shugart, J.A.; Horng, T.; Malarkannan, S.; Shastri, N. Unanticipated antigens: Translation initiation at CUG with leucine. PLoS Biol. 2004, 2, e366. [Google Scholar] [CrossRef]
- Sendoel, A.; Dunn, J.G.; Rodriguez, E.H.; Naik, S.; Gomez, N.C.; Hurwitz, B.; Levorse, J.; Dill, B.D.; Schramek, D.; Molina, H.; et al. Translation from unconventional 5′ start sites drives tumour initiation. Nature 2017, 541, 494–499. [Google Scholar] [CrossRef]
- Donnelly, N.; Gorman, A.M.; Gupta, S.; Samali, A. The eIF2α kinases: Their structures and functions. Cell. Mol. Life Sci. 2013, 70, 3493–3511. [Google Scholar] [CrossRef]
- Novoa, I.; Zeng, H.; Harding, H.P.; Ron, D. Feedback inhibition of the unfolded protein response by GADD34-mediated dephosphorylation of eIF2α. J. Cell Biol. 2001, 5, 1011–1021. [Google Scholar] [CrossRef] [Green Version]
- Spilka, R.; Ernst, C.; Mehta, A.K.; Haybaeck, J. Eukaryotic translation initiation factors in cancer development and progression. Cancer Lett. 2013, 340, 9–21. [Google Scholar] [CrossRef]
- Palam, L.R.; Baird, T.D.; Wek, R.C. Phosphorylation of eIF2 facilitates ribosomal bypass of an inhibitory upstream ORF to enhance CHOP translation. J. Biol. Chem. 2011, 286, 10939–10949. [Google Scholar] [CrossRef] [Green Version]
- Andreev, D.E.; O’Connor, P.B.F.; Fahey, C.; Kenny, E.M.; Terenin, I.M.; Dmitriev, S.E.; Cormican, P.; Morris, D.W.; Shatsky, I.N.; Baranov, P.V. Translation of 5’ leaders is pervasive in genes resistant to eIF2 repression. eLife 2015, 4, e03971. [Google Scholar] [CrossRef]
- Baird, T.D.; Palam, L.R.; Fusakio, M.E.; Willy, J.A.; Davis, C.M.; McClintick, J.N.; Anthony, T.G.; Wek, R.C. Selective mRNA translation during eIF2 phosphorylation induces expression of IBTKα. Mol. Biol. Cell 2014, 25, 1686–1697. [Google Scholar] [CrossRef]
- Silva, J.; Fernandes, R.; Romão, L. Translational regulation by upstream open reading frames and human diseases. In The MRNA Metabolism in Human Disease; Romão, L., Ed.; Springer: Cham, Germany, 2019; Volume 1157, pp. 99–116. [Google Scholar] [CrossRef]
- Schulz, J.; Mah, N.; Neuenschwander, M.; Kischka, T.; Ratei, R.; Schlag, P.M.; Castaños-Vélez, E.; Fichtner, I.; Tunn, P.; Denkert, C.; et al. Loss-of-function uORF mutations in human malignancies. Sci. Rep. 2018, 8, 2395. [Google Scholar] [CrossRef] [Green Version]
- Dean, M.; Rzhetsky, A.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Genome Res. 2001, 11, 1156–1166. [Google Scholar] [CrossRef]
- Tian, Y.; Han, X.; Tian, D. The biological regulation of ABCE1. IUBMB Life 2012, 64, 795–800. [Google Scholar] [CrossRef]
- Dean, M.; Annilo, T. Evolution of the ATP-binding cassette (ABC) transporter superfamily in vertebrates. Annu. Rev. Genomics Hum. Genet. 2005, 6, 123–142. [Google Scholar] [CrossRef]
- Bisbal, C.; Martinand, C.; Silhol, M.; Lebleu, B.; Salehzada, T. Cloning and characterization of a RNase L inhibitor. A new component of the interferon-regulated 2-5A pathway. J. Biol. Chem. 1995, 270, 13308–13317. [Google Scholar] [CrossRef] [Green Version]
- Toompuu, M.; Kärblane, K.; Pata, P.; Truve, E.; Sarmiento, C. ABCE1 is essential for S phase progression in human cells. Cell Cycle 2016, 15, 1234–1247. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Blackford, J.A.; Hassel, B.A. RNase L mediates the antiviral effect of interferon through a selective reduction in viral RNA during encephalomyocarditis virus infection. J. Virol. 1998, 72, 2752–2759. [Google Scholar] [CrossRef] [Green Version]
- Liang, S.; Quirk, D.; Zhou, A. RNase L: Its biological roles and regulation. IUBMB Life 2006, 58, 508–514. [Google Scholar] [CrossRef]
- Chen, Z.; Dong, J.; Ishimura, A.; Daar, I.; Hinnebusch, A.G.; Dean, M. The essential vertebrate ABCE1 protein interacts with eukaryotic initiation factors. J. Biol. Chem. 2006, 281, 7452–7457. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Lai, R.; Nielsen, K.; Fekete, C.A.; Qiu, H.; Hinnebusch, A.G. The essential ATP-binding cassette protein RLI1 functions in translation by promoting preinitiation complex assembly. J. Biol. Chem. 2004, 279, 42157–42168. [Google Scholar] [CrossRef] [Green Version]
- Beißel, C.; Neumann, B.; Uhse, S.; Hampe, I.; Karki, P.; Krebber, H. Translation termination depends on the sequential ribosomal entry of eRF1 and eRF3. Nucleic Acids Res. 2019, 47, 4798–4813. [Google Scholar] [CrossRef]
- Pisarev, A.V.; Skabkin, M.A.; Pisareva, V.P.; Skabkina, O.V.; Rakotondrafara, A.M.; Hentze, M.W.; Hellen, C.U.T.; Pestova, T.V. The role of ABCE1 in eukaryotic post-termination ribosomal recycling. Mol. Cell 2010, 37, 196–210. [Google Scholar] [CrossRef] [Green Version]
- Diriong, S.; Salehzada, T.; Bisbal, C.; Martinand, C.; Taviaux, S. Localization of the ribonuclease L inhibitor gene (RNS4I), a new member of the interferon-regulated 2–5A pathway, to 4q31 by fluorescence in situ hybridization. Genomics 1996, 32, 488–490. [Google Scholar] [CrossRef]
- Ren, Y.; Li, Y.; Tian, D. Role of the ABCE1 gene in human lung adenocarcinoma. Oncol. Rep. 2012, 27, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Yu, Q.; Ji, H.; Tian, D. Tip60-siRNA regulates ABCE1 acetylation to suppress lung cancer growth via activation of the apoptotic signaling pathway. Exp. Ther. Med. 2019, 17, 3195–3202. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Gao, Y.; Tian, D.; Zheng, M. A small interfering ABCE1-targeting RNA inhibits the proliferation and invasiveness of small cell lung cancer. Int. J. Mol. Med. 2010, 25, 687–693. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Zhou, H.; Lang, X.; Liu, Z. siRNA-induced ABCE1 silencing inhibits proliferation and invasion of breast cancer cells. Mol. Med. Rep. 2014, 10, 1685–1690. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Gong, X.; Zhou, H.; Xiong, F.; Wang, S. Depleting ABCE1 expression induces apoptosis and inhibits the ability of proliferation and migration of human esophageal carcinoma cells. Int. J. Clin. Exp. Pathol. 2014, 7, 584–592. [Google Scholar]
- Seborova, K.; Vaclavikova, R.; Soucek, P.; Elsnerova, K.; Bartakova, A.; Cernaj, P.; Bouda, J.; Rob, L.; Hruda, M.; Dvorak, P. Association of ABC gene profiles with time to progression and resistance in ovarian cancer revealed by bioinformatics analyses. Cancer Med. 2019, 8, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Chen, X.; Ding, B.; Liu, H.; He, T. Down-regulation of ABCE1 inhibits temozolomide resistance in glioma through the PI3K/Akt/NF-κB signaling pathway. Biosci. Rep. 2018, 38, BSR20181711. [Google Scholar] [CrossRef] [Green Version]
- Shichijo, S.; Ishihara, Y.; Azuma, K.; Komatsu, N.; Higashimoto, N.; Ito, M.; Nakamura, T.; Ueno, T.; Harada, M.; Itoh, K. ABCE1, a member of ATP-binding cassette transporter gene, encodes peptides capable of inducing HLA-A2-restricted and tumor-reactive cytotoxic T lymphocytes in colon cancer patients. Oncol. Rep. 2005, 13, 907–913. [Google Scholar] [CrossRef]
- Hlavata, I.; Mohelnikova-Duchonova, B.; Vaclavikova, R.; Liska, V.; Pitule, P.; Novak, P.; Bruha, J.; Vycital, O.; Holubec, L.; Treska, V.; et al. The role of ABC transporters in progression and clinical outcome of colorectal cancer. Mutagenesis 2012, 27, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crappé, J.; Ndah, E.; Koch, A.; Steyaert, S.; Gawron, D.; De Keulenaer, S.; De Meester, E.; De Meyer, T.; Van Criekinge, W.; Van Damme, P.; et al. PROTEOFORMER: Deep proteome coverage through ribosome profiling and MS integration. Nucleic Acids Res. 2015, 43, e29. [Google Scholar] [CrossRef]
- Wurth, L.; Papasaikas, P.; Olmeda, D.; Bley, N.; Calvo, G.T.; Guerrero, S.; Cerezo-Wallis, D.; Martinez-Useros, J.; García-Fernández, M.; Hüttelmaier, S.; et al. UNR/CSDE1 Drives a post-transcriptional program to promote melanoma invasion and metastasis. Cancer Cell 2016, 30, 694–707. [Google Scholar] [CrossRef] [Green Version]
- Vanderperre, B.; Lucier, J.; Bissonnette, C.; Motard, J.; Tremblay, G.; Vanderperre, S.; Wisztorski, M.; Salzet, M.; Boisvert, F.; Roucou, X. Direct detection of alternative open reading frames translation products in human significantly expands the proteome. PLoS ONE 2013, 8, e70698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, C.; Romão, L. Translation of the human erythropoietin transcript is regulated by an upstream open reading frame in response to hypoxia. RNA 2014, 20, 594–608. [Google Scholar] [CrossRef] [Green Version]
- Kozak, M. Point mutations define a sequence flanking the AUG initiator codon that modulates translation by eukaryotic ribosomes. Cell 1986, 44, 283–292. [Google Scholar] [CrossRef]
- Kozak, M. At least six nucleotides preceding the AUG initiator codon enhance translation in mammalian cells. J. Mol. Biol. 1987, 196, 947–950. [Google Scholar] [CrossRef]
- Kozak, M. Context Effects and inefficient initiation at non-AUG codons in eucaryotic cell-free translation systems. Mol. Cell Biol. 1989, 9, 5073–5080. [Google Scholar] [CrossRef] [Green Version]
- Iacono, M.; Mignone, F.; Pesole, G. uAUG and uORFs in human and rodent 5′untranslated mRNAs. Gene 2005, 349, 97–105. [Google Scholar] [CrossRef]
- Johnstone, T.G.; Bazzini, A.A.; Giraldez, A.J. Upstream ORFs are prevalent translational repressors in vertebrates. EMBO J. 2016, 35, 706–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onofre, C.; Tomé, F.; Barbosa, C.; Silva, A.L.; Romão, L. Expression of human hemojuvelin (HJV) is tightly regulated by two upstream open reading frames in HJV mRNA that respond to iron overload in hepatic cells. Mol. Cell Biol. 2015, 35, 1376–1389. [Google Scholar] [CrossRef] [Green Version]
- Young, S.K.; Baird, T.D.; Wek, R.C. Translation regulation of the glutamyl-prolyl-tRNA synthetase gene EPRS through bypass of upstream open reading frames with noncanonical initiation codons. J. Biol. Chem. 2016, 291, 10824–10835. [Google Scholar] [CrossRef] [Green Version]
- Wethmar, K.; Smink, J.J.; Leutz, A. Upstream open reading frames: Molecular switches in (patho)physiology. Bioessays 2010, 32, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Hinnebusch, A.G. Translational regulation of GCN4 and the general amino acid control of yeast. Annu. Rev. Microbiol. 2005, 59, 407–450. [Google Scholar] [CrossRef] [PubMed]
- Spriggs, K.A.; Bushell, M.; Willis, A.E. Translational regulation of gene expression during conditions of cell stress. Mol. Cell 2010, 40, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Vattem, K.M.; Wek, R.C. Reinitiation involving upstream ORFs regulates ATF4 mRNA translation in mammalian cells. Proc. Natl. Acad. Sci. USA 2004, 101, 11269–11274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, S.K.; Willy, J.A.; Wu, C.; Sachs, M.S.; Wek, R.C. Ribosome reinitiation directs gene-specific translation and regulates the integrated stress response. J. Biol. Chem. 2015, 290, 28257–28271. [Google Scholar] [CrossRef] [PubMed] [Green Version]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| uORF ID | uORF Nucleotide Sequence | uORF Length (nts) | Intercistronic Distance (nts) | uORF-Peptide Length (aa) | Kozak Sequence Context |
|---|---|---|---|---|---|
| uORF1 | ATGACTAAGGCTCCACTCCTGACCCACCGGCCTTGA | 36 | 426 | 11 | GTTAGAATGA |
| uORF2 | ATGCATCTGTTTACGCTAGGACCACGCTCGACGTCGGAGAAAAGCCCACACACTCACAGTTTCCAGACCTGGCTGCTTGCCGAAACTCAGATTCTCGGCACCTCCAGCAGCTGGCTTCGCCAACGGCGTTGA | 132 | 252 | 43 | AGGCGAATGC |
| uORF3 | ATGACGTCATATCTCCCTACCTACCTCCAGGGTTCCGCCTCACGCTCTATGTCGCGCGCGCGCACTACGTCCTATGGCTTGCGCGTGCGGCGGCTGGGCACCGCCATTTTGGCCGGTGGCCGTGAGAACACGCTGTGTGGCTGA | 144 | 91 | 47 | AGCTCAATGA |
| uORF4 | ATGTCGCGCGCGCGCACTACGTCCTATGGCTTGCGCGTGCGGCGGCTGGGCACCGCCATTTTGGCCGGTGGCCGTGAGAACACGCTGTGTGGCTGA | 96 | 91 | 31 | CGCTCTATGT |
| uORF5 | ATGGCTTGCGCGTGCGGCGGCTGGGCACCGCCATTTTGGCCGGTGGCCGTGAGAACACGCTGTGTGGCTGAAAAGTGA | 78 | 84 | 25 | CGTCCTATGG |
| uORF6 | GTGCGGCGGCTGGGCACCGCCATTTTGGCCGGTGGCCGTGAGAACACGCTGTGTGGCTGA | 60 | 91 | 29 | TTGCGCGTGC |
| uORF7 | GTGGCCGTGAGAACACGCTGTGTGGCTGAAAAGTGA | 36 | 84 | 11 | TGGCCGGTGG |
| uORF8 | GTGAAGGCAAGAGCTGATTTGGCCTCTGTGCTCCCCTCCGCAAGGGGATCGTTTTCTCCAGAAGAGCTGGATATTCTTTCGCCCAGTTATGGCAGACAAGTTAACGAGAATTGCTATTGTCAACCATGA | 129 | (overlapped) | 42 | TGAAAAGTGA |
| uORF9 | GTGCTCCCCTCCGCAAGGGGATCGTTTTCTCCAGAAGAGCTGGATATTCTTTCGCCCAGTTATGGCAGACAAGTTAACGAGAATTGCTATTGTCAACCATGA | 102 | (overlapped) | 33 | GCCTCTGTGC |
| uORF10 | CTGGATATTCTTTCGCCCAGTTATGGCAGACAAGTTAACGAGAATTGCTATTGTCAACCATGA | 63 | (overlapped) | 20 | GAAGAGCTGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, J.; Nina, P.; Romão, L. Translation of ABCE1 Is Tightly Regulated by Upstream Open Reading Frames in Human Colorectal Cells. Biomedicines 2021, 9, 911. https://doi.org/10.3390/biomedicines9080911
Silva J, Nina P, Romão L. Translation of ABCE1 Is Tightly Regulated by Upstream Open Reading Frames in Human Colorectal Cells. Biomedicines. 2021; 9(8):911. https://doi.org/10.3390/biomedicines9080911
Chicago/Turabian StyleSilva, Joana, Pedro Nina, and Luísa Romão. 2021. "Translation of ABCE1 Is Tightly Regulated by Upstream Open Reading Frames in Human Colorectal Cells" Biomedicines 9, no. 8: 911. https://doi.org/10.3390/biomedicines9080911
APA StyleSilva, J., Nina, P., & Romão, L. (2021). Translation of ABCE1 Is Tightly Regulated by Upstream Open Reading Frames in Human Colorectal Cells. Biomedicines, 9(8), 911. https://doi.org/10.3390/biomedicines9080911