
Assessment of Platelet Mitochondrial Respiration in a Pediatric Population: A Pilot Study in Healthy Children and Children with Acute Lymphoblastic Leukemia

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Sample Acquisition and Platelet Preparation
2.3. Characteristics of the Patients with ALL
2.4. High-Resolution Respirometry Studies
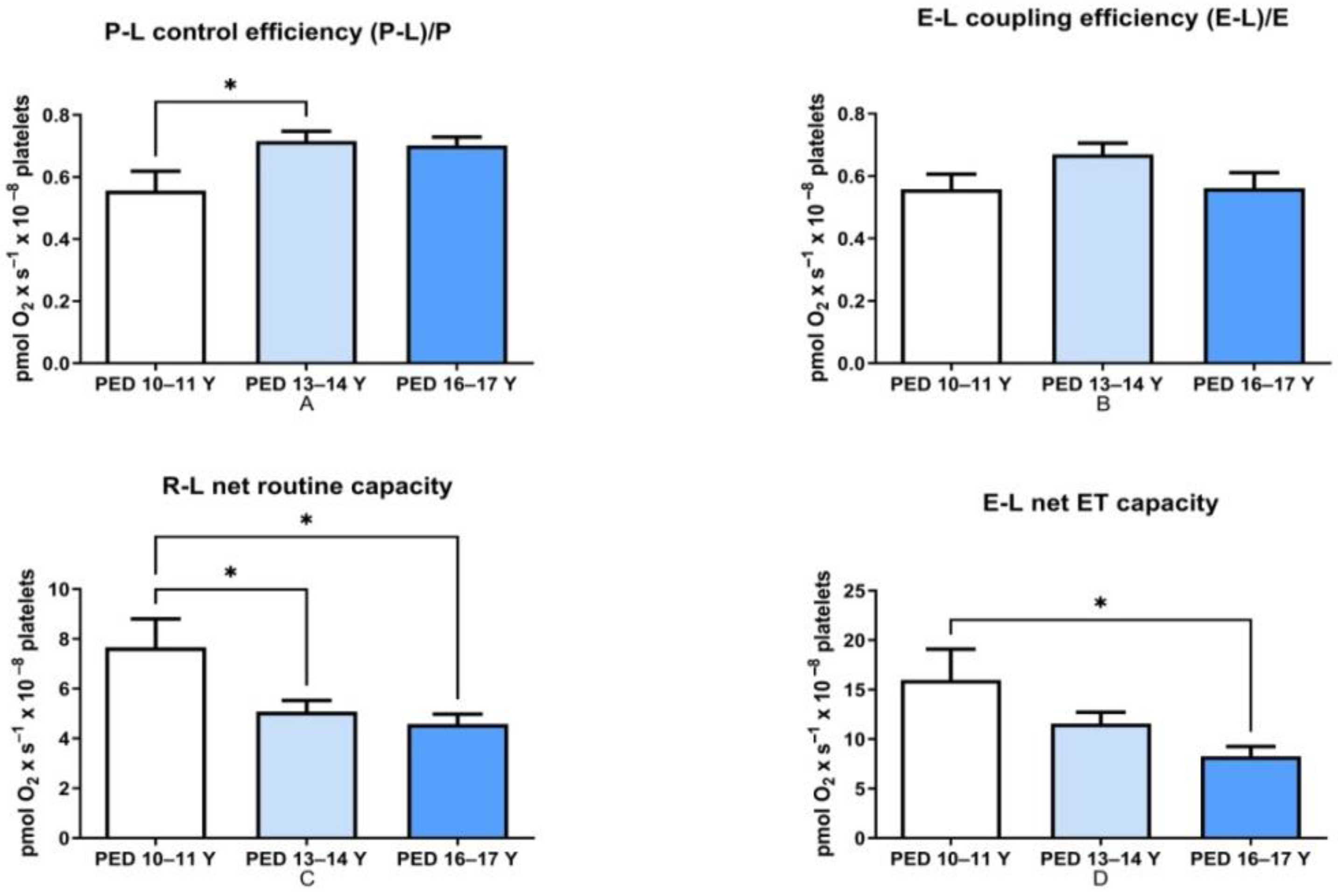
- P-L control efficiency—evaluates the efficiency of ATP generation and was calculated by subtracting LEAK respiration from OXPHOS capacity and then dividing the result by the OXPHOS capacity (P-L/P).
- E-L coupling efficiency—measures the degree of coupling and was calculated by subtracting LEAK respiration from the ETS capacity and then dividing the result by the ETS capacity (E-L/E).
- R-L net routine capacity—is the respiratory capacity available for phosphorylation of ADP to ATP and was calculated as the difference between Routine respiration and LEAK respiration.
- E-L net ET capacity—is the respiratory capacity potentially available for ion transport and phosphorylation of ADP to ATP and was calculated as the difference between ETS capacity and LEAK respiration.
- Routine control ratio—evaluates how close routine respiration (R) operates to the ETS maximal capacity and was calculated as the ratio between Routine and ETS capacity (R/E).
2.5. Data Analysis
2.6. Reagents
3. Results
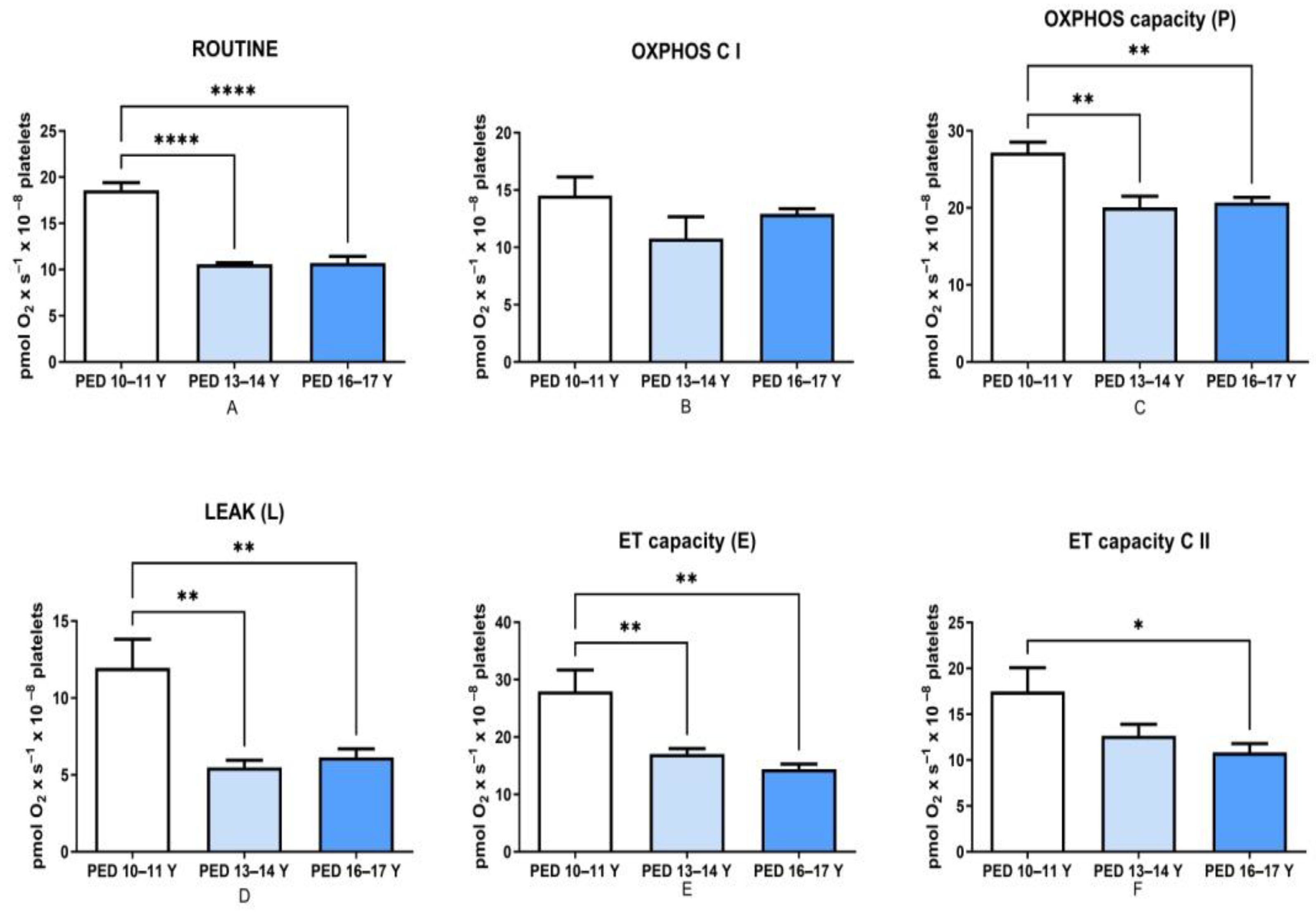
3.1. Mitochondrial Respiration Decreased with Age in Non-HM Children
3.2. Platelet Respiration Is Modified at the Onset of ALL
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petrus, A.T.; Lighezan, L.D.; Danila, M.D.; Duicu, O.M.; Sturza, A.; Muntean, D.M.; Ionita, I. Assessment of platelets respiration as emerging biomarker of disease. Physiol. Res. 2019, 68, 347–363. [Google Scholar] [CrossRef]
- Sjövall, F.; Ehinger, J.K.H.; Marelsson, S.E.; Morota, S.; Åsander Frostner, E.; Uchino, H.; Lundgren, J.; Arnbjörnsson, E.; Hansson, M.J.; Fellman, V.; et al. Mitochondrial respiration in human viable platelets—Methodology and influence of gender, age and storage. Mitochondrion 2013, 13, 7–14. [Google Scholar] [CrossRef]
- Chacko, B.K.; Kramer, P.A.; Ravi, S.; Johnson, M.S.; Hardy, R.W.; Ballinger, S.W.; Darley-Usmar, V.M. Methods for defining distinct bioenergetic profiles in platelets, lymphocytes, monocytes, and neutrophils, and the oxidative burst from human blood. Lab. Investig. 2013, 93, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Gvozdjakova, A.; Kucharska, J.; Chladekova, A.; Rausova, Z.; Vancova, O.; Komlosi, M.; Ulicna, O.; Mojto, V. Platelet mitochondrial bioenergetic analysis in patients with nephropathies and non-communicable diseases: A new method. Bratisl. Med. J. 2019, 120, 630–635. [Google Scholar] [CrossRef] [Green Version]
- Chacko, B.K.; Kramer, P.A.; Ravi, S.; Benavides, G.A.; Mitchell, T.; Dranka, B.P.; Ferrick, D.; Singal, A.K.; Ballinger, S.W.; Bailey, S.M.; et al. The Bioenergetic Health Index: A new concept in mitochondrial translational research. Clin. Sci. 2014, 127, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.T.; Corken, A.; Ware, J. Platelets at the interface of thrombosis, inflammation, and cancer. Blood 2015, 126, 582–588. [Google Scholar] [CrossRef] [Green Version]
- Cordero, M.D.; De Miguel, M.; Moreno Fernández, A.M.; Carmona López, I.M.; Garrido Maraver, J.; Cotán, D.; Gómez Izquierdo, L.; Bonal, P.; Campa, F.; Bullon, P.; et al. Mitochondrial dysfunction and mitophagy activation in blood mononuclear cells of fibromyalgia patients: Implications in the pathogenesis of the disease. Arthritis Res. 2010, 12, R17. [Google Scholar] [CrossRef] [Green Version]
- Japiassú, A.M.; Santiago, A.P.S.A.; d’Avila, J.d.C.P.; Garcia-Souza, L.F.; Galina, A.; Castro Faria-Neto, H.C.; Bozza, F.A.; Oliveira, M.F. Bioenergetic failure of human peripheral blood monocytes in patients with septic shock is mediated by reduced F1Fo adenosine-5′-triphosphate synthase activity. Crit. Care Med. 2011, 39, 1056–1063. [Google Scholar] [CrossRef]
- Sjövall, F.; Morota, S.; Hansson, M.J.; Friberg, H.; Gnaiger, E.; Elmér, E. Temporal increase of platelet mitochondrial respiration is negatively associated with clinical outcome in patients with sepsis. Crit. Care 2010, 14, R214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila, C.; Huang, R.J.; Stevens, M.V.; Aponte, A.M.; Tripodi, D.; Kim, K.Y.; Sack, M.N. Platelet mitochondrial dysfunction is evident in type 2 diabetes in association with modifications of mitochondrial anti-oxidant stress proteins. Exp. Clin. Endocrinol. Diabetes 2012, 120, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Gnaiger, E.; Kuznetsov, A.V.; Schneeberger, S.; Seiler, R.; Brandacher, G.; Steurer, W.; Margreiter, R. Mitochondria in the Cold. In Life in the Cold; Heldmaier, G., Klingenspor, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2000. [Google Scholar] [CrossRef]
- Pesta, D.; Gnaiger, E. High-Resolution Respirometry: OXPHOS Protocols for Human Cells and Permeabilized Fibers from Small Biopsies of Human Muscle. In Mitochondrial Bioenergetics: Methods and Protocols; Palmeira, C.M., Moreno, A.J., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 25–58. [Google Scholar]
- Doerrier, C.; Sumbalova, Z.; Krumschnabel, G.; Hiller, E.; Gnaiger, E. SUIT reference protocol for OXPHOS analysis by high-resolution respirometry. Mitochondr. Physiol. Netw. 2016, 21, 1–12. [Google Scholar]
- Sumbalova, Z.; Garcia-Souza, L.F.; Velika, B.; Volani, C.; Gnaiger, E. Respirometric Analysis of Mitochondrial Function in Human Blood Cells. In:Recent Advances in Mitochondrial Medicine and Coenzyme Q10; Gvozdjáková, A., Cornelissen, G., Singh, R.B., Eds.; Nova Sciences: Hauppauge, NY, USA, 2018; pp. 255–268. [Google Scholar]
- Baaten, C.C.F.M.J.; Moenen, F.C.J.I.; Henskens, Y.M.C.; Swieringa, F.; Wetzels, R.J.H.; van Oerle, R.; Heijnen, H.F.G.; Ten Cate, H.; Holloway, G.P.; Beckers, E.A.M.; et al. Impaired mitochondrial activity explains platelet dysfunction in thrombocytopenic cancer patients undergoing chemotherapy. Haematologica 2018, 103, 1557–1567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avram, V.F.; Bîna, A.M.; Sima, A.; Aburel, O.M.; Sturza, A.; Burlacu, O.; Timar, R.Z.; Muntean, D.M.; Elmér, E.; Crețu, O.M. Improvement of Platelet Respiration by Cell–Permeable Succinate in Diabetic Patients Treated with Statins. Life 2021, 11, 288. [Google Scholar] [CrossRef]
- ALL IC BFM 2009—A Randomized Trial of the I BFM-SG for the Management of Childhood Non-B Acute Lymphoblastic Leukemia. 2009. Available online: https://www.bialaczka.org/wp-content/uploads/2016/10/ALLIC_BFM_2009.pdf (accessed on 5 December 2021).
- Brand, M.D.; Nicholls, D.G. Assessing mitochondrial dysfunction in cells. Biochem. J. 2011, 435, 297–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnaiger, E.; Asander Frostner, E.; Abdul, K.N.; Abdel-Rahman, E.A.; Abumrad, N.A.; Acuna-Castroviejo, D.; Adiele, R.C.; Ahn, B.; Alencar, M.B.; Ali, S.S.; et al. Mitochondrial physiology. Bioenerg. Commun. 2020, 1, 44. [Google Scholar] [CrossRef]
- Amiel, S.A.; Caprio, S.; Sherwin, R.S.; Plewe, G.; Haymond, M.W.; Tamborlane, W.V. Insulin Resistance of Puberty: A Defect Restricted to Peripheral Glucose Metabolism. J. Clin. Endocrinol. Metab. 1991, 72, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Arslanian, S.; Suprasongsin, C. Insulin sensitivity, lipids, and body composition in childhood: Is “syndrome X” present? J. Clin. Endocrinol. Metab. 1996, 81, 1058–1062. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.S.; Hoffman, R.P.; Stene, M.A.; Hansen, J.R. Effects of maturational stage on insulin sensitivity during puberty. J. Clin. Endocrinol. Metab. 1993, 77, 725–730. [Google Scholar] [CrossRef]
- Travers, S.H.; Jeffers, B.W.; Bloch, C.A.; Hill, J.O.; Eckel, R.H. Gender and Tanner stage differences in body composition and insulin sensitivity in early pubertal children. J. Clin. Endocrinol. Metab. 1995, 80, 172–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, A.; Jacobs, D.R.; Steinberger, J.; Hong, C.P.; Prineas, R.; Luepker, R.; Sinaiko, A.R. Insulin resistance during puberty: Results from clamp studies in 357 children. Diabetes 1999, 48, 2039. [Google Scholar] [CrossRef]
- Fleischman, A.; Kron, M.; Systrom, D.M.; Hrovat, M.; Grinspoon, S.K. Mitochondrial Function and Insulin Resistance in Overweight and Normal-Weight Children. J. Clin. Endocrinol. Metab. 2009, 94, 4923–4930. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, C.-P.; Hoppel, C. Analyzing mitochondrial function in human peripheral blood mononuclear cells. Anal. Biochem. 2018, 549, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Schimmer, A.D.; Škrtić, M. Therapeutic potential of mitochondrial translation inhibition for treatment of acute myeloid leukemia. Expert Rev. Hematol. 2012, 5, 117–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panina, S.B.; Pei, J.; Kirienko, N.V. Mitochondrial metabolism as a target for acute myeloid leukemia treatment. Cancer Metab. 2021, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Basak, N.P.; Banerjee, S. Mitochondrial dependency in progression of acute myeloid leukemia. Mitochondrion 2015, 21, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Farge, T.; Saland, E.; de Toni, F.; Aroua, N.; Hosseini, M.; Perry, R.; Bosc, C.; Sugita, M.; Stuani, L.; Fraisse, M.; et al. Chemotherapy-Resistant Human Acute Myeloid Leukemia Cells Are Not Enriched for Leukemic Stem Cells but Require Oxidative Metabolism. Cancer Discov. 2017, 7, 716–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashton, T.M.; McKenna, W.G.; Kunz-Schughart, L.A.; Higgins, G.S. Oxidative Phosphorylation as an Emerging Target in Cancer Therapy. Clin. Cancer Res. 2018, 24, 2482. [Google Scholar] [CrossRef] [Green Version]
- Baccelli, I.; Gareau, Y.; Lehnertz, B.; Gingras, S.; Spinella, J.-F.; Corneau, S.; Mayotte, N.; Girard, S.; Frechette, M.; Blouin-Chagnon, V.; et al. Mubritinib Targets the Electron Transport Chain Complex I and Reveals the Landscape of OXPHOS Dependency in Acute Myeloid Leukemia. Cancer Cell 2019, 36, 84–99.e88. [Google Scholar] [CrossRef]
- Eberhart, K.; Rainer, J.; Bindreither, D.; Ritter, I.; Gnaiger, E.; Kofler, R.; Oefner, P.J.; Renner, K. Glucocorticoid-induced alterations in mitochondrial membrane properties and respiration in childhood acute lymphoblastic leukemia. Biochim. Biophys. Acta (BBA)-Bioenerg. 2011, 1807, 719–725. [Google Scholar] [CrossRef] [Green Version]
- Beesley, A.H.; Firth, M.J.; Ford, J.; Weller, R.E.; Freitas, J.R.; Perera, K.U.; Kees, U.R. Glucocorticoid resistance in T-lineage acute lymphoblastic leukaemia is associated with a proliferative metabolism. Br. J. Cancer 2009, 100, 1926–1936. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Liu, W.; Huang, Q.; Wang, Y.; Li, H.; Xiong, Y. Targeting mitochondrial respiration selectively sensitizes pediatric acute lymphoblastic leukemia cell lines and patient samples to standard chemotherapy. Am. J. Cancer Res. 2017, 7, 2395–2405. [Google Scholar]
- Hlozkova, K.; Pecinova, A.; Alquezar-Artieda, N.; Pajuelo-Reguera, D.; Simcikova, M.; Hovorkova, L.; Rejlova, K.; Zaliova, M.; Mracek, T.; Kolenova, A.; et al. Metabolic profile of leukemia cells influences treatment efficacy of L-asparaginase. BMC Cancer 2020, 20, 526. [Google Scholar] [CrossRef] [PubMed]
- Barbato, A.; Scandura, G.; Puglisi, F.; Cambria, D.; La Spina, E.; Palumbo, G.A.; Lazzarino, G.; Tibullo, D.; Di Raimondo, F.; Giallongo, C.; et al. Mitochondrial Bioenergetics at the Onset of Drug Resistance in Hematological Malignancies: An Overview. Front. Oncol. 2020, 10, 604143. [Google Scholar] [CrossRef]
- Caro, P.; Kishan, A.U.; Norberg, E.; Stanley, I.A.; Chapuy, B.; Ficarro, S.B.; Polak, K.; Tondera, D.; Gounarides, J.; Yin, H.; et al. Metabolic signatures uncover distinct targets in molecular subsets of diffuse large B cell lymphoma. Cancer Cell 2012, 22, 547–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M. Generation, function and diagnostic value of mitochondrial DNA copy number alterations in human cancers. Life Sci. 2011, 89, 65–71. [Google Scholar] [CrossRef]
- Roy Chowdhury, S.; Bouchard, E.D.J.; Saleh, R.; Nugent, Z.; Peltier, C.; Mejia, E.; Hou, S.; McFall, C.; Squires, M.; Hewitt, D.; et al. Mitochondrial Respiration Correlates with Prognostic Markers in Chronic Lymphocytic Leukemia and Is Normalized by Ibrutinib Treatment. Cancers 2020, 12, 650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.; Wen-jun, L. Platelet Changes in Acute Leukemia. Cell Biochem. Biophys. 2013, 67, 1473–1479. [Google Scholar] [CrossRef]
- Strauß, G.; Vollert, C.; von Stackelberg, A.; Weimann, A.; Gaedicke, G.; Schulze, H. Immature platelet count: A simple parameter for distinguishing thrombocytopenia in pediatric acute lymphocytic leukemia from immune thrombocytopenia. Pediatric Blood Cancer 2011, 57, 641–647. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, J.; Wu, P.; Feng, K.; Li, J.; Xie, Z.; Xue, P.; Cai, T.; Cui, Z.; Chen, X.; et al. Discovery and identification of potential biomarkers of pediatric acute lymphoblastic leukemia. Proteome Sci. 2009, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Noh, J.-Y. Megakaryopoiesis and Platelet Biology: Roles of Transcription Factors and Emerging Clinical Implications. Int. J. Mol. Sci. 2021, 22, 9615. [Google Scholar] [CrossRef]
- Sun, C.; Chang, L.; Zhu, X. Pathogenesis of ETV6/RUNX1-positive childhood acute lymphoblastic leukemia and mechanisms underlying its relapse. Oncotarget 2017, 8, 35445–35459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez, J.; Enciso, L.J.; Suarez, M.; Fiegl, M.; Grismaldo, A.; López, C.; Barreto, A.; Cardozo, C.; Palacios, P.; Morales, L.; et al. Platelets Promote Mitochondrial Uncoupling and Resistance to Apoptosis in Leukemia Cells: A Novel Paradigm for the Bone Marrow Microenvironment. Cancer Microenviron. 2014, 7, 79–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horacek, J.M.; Kupsa, T.; Vanek, J.; Jebavy, L.; Zak, P. Serum Levels of Selected Cytokines in B-Cell Precursor Acute Lymphoblastic Leukemia and Their Association with Prognostic Factors and Survival. Blood 2019, 134, 5078. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Immuno- Phenotype | Risk Group | Karyotype/Genomic Characteristics | |
|---|---|---|---|
| Patient 1 | Mature T-ALL (Egil stage IV) | High risk (HR) | t(1;14)(p32q11) |
| Patient 2 | Common B ALL | Medium risk (MR) | TEL-AML1 (ETV6-RUNX1) positive |
| Patient 3 | Common B ALL | Medium risk (MR) | Normal Karyotype; major BCR-ABL1(b2a2-b3a2-Fusion)-negative; minor BCR-ABL1 (e1a2 Fusion)-negative; KMT2A-AFF1 (MLL1-AF4)-negative; TCF3-PBX1 (E2A-PBX1)-negative; ETV6-RUNX1 (TEL-AML1)-negative |
| Patient 4 | Pre-B ALL | Medium risk (MR) | Normal Karyotype; major BCR-ABL1 (b2a2-b3a2-Fusion)-negative; minor BCR-ABL1 (e1a2 Fusion)-negative; KMT2A-AFF1 (MLL1-AF4)-negative; TCF3-PBX1 (E2A-PBX1)-negative; ETV6-RUNX1 (TEL-AML1)-negative |
| Respiratory Parameters | ALL Group | Age-Matched Non-HM Group |
|---|---|---|
| Routine | 10.8 ± 0.67 **** | 18.6 ± 0.8 |
| OXPHOS C I | 19.3 ± 0.95 * | 14.5 ± 1.61 |
| Maximal OXPHOS (C I and II) | 27.0 ± 1.42 | 27.2 ± 1.36 |
| LEAK (L) | 7.79 ± 1.25 | 12 ± 1.85 |
| ET capacity (E) | 20.8 ± 1.42 * | 28 ± 3.73 |
| ET capacity C II | 12.2 ± 0.79 * | 17.5 ± 2.59 |
| P-L control efficiency (P-L/P) | 19.2 ± 1.67 | 15.2 ± 1.95 |
| ET coupling efficiency (E-L/E) | 0.65 ± 0.054 | 0.558 ± 0.048 |
| R/E | 0.545 ± 0.05 * | 0.737 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lelcu, T.; Bînă, A.M.; Dănilă, M.D.; Popoiu, C.M.; Aburel, O.M.; Arghirescu, S.T.; Borza, C.; Muntean, D.M. Assessment of Platelet Mitochondrial Respiration in a Pediatric Population: A Pilot Study in Healthy Children and Children with Acute Lymphoblastic Leukemia. Children 2021, 8, 1196. https://doi.org/10.3390/children8121196
Lelcu T, Bînă AM, Dănilă MD, Popoiu CM, Aburel OM, Arghirescu ST, Borza C, Muntean DM. Assessment of Platelet Mitochondrial Respiration in a Pediatric Population: A Pilot Study in Healthy Children and Children with Acute Lymphoblastic Leukemia. Children. 2021; 8(12):1196. https://doi.org/10.3390/children8121196
Chicago/Turabian StyleLelcu, Theia, Anca M. Bînă, Maria D. Dănilă, Călin M. Popoiu, Oana M. Aburel, Smaranda T. Arghirescu, Claudia Borza, and Danina M. Muntean. 2021. "Assessment of Platelet Mitochondrial Respiration in a Pediatric Population: A Pilot Study in Healthy Children and Children with Acute Lymphoblastic Leukemia" Children 8, no. 12: 1196. https://doi.org/10.3390/children8121196
APA StyleLelcu, T., Bînă, A. M., Dănilă, M. D., Popoiu, C. M., Aburel, O. M., Arghirescu, S. T., Borza, C., & Muntean, D. M. (2021). Assessment of Platelet Mitochondrial Respiration in a Pediatric Population: A Pilot Study in Healthy Children and Children with Acute Lymphoblastic Leukemia. Children, 8(12), 1196. https://doi.org/10.3390/children8121196