
Novel Treatments and Technologies Applied to the Cure of Neuroblastoma

 ,
,  , and
, and 
Abstract
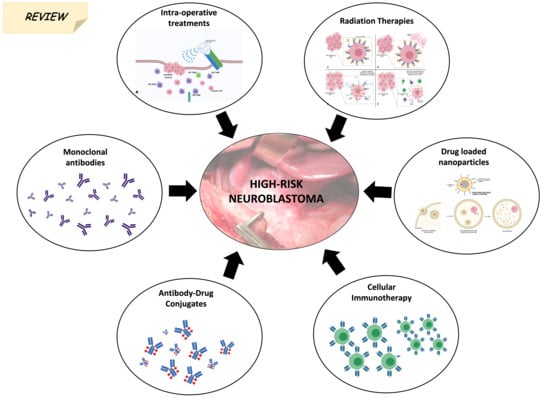
:
1. Introduction
2. Novel Molecules and Nanoparticles
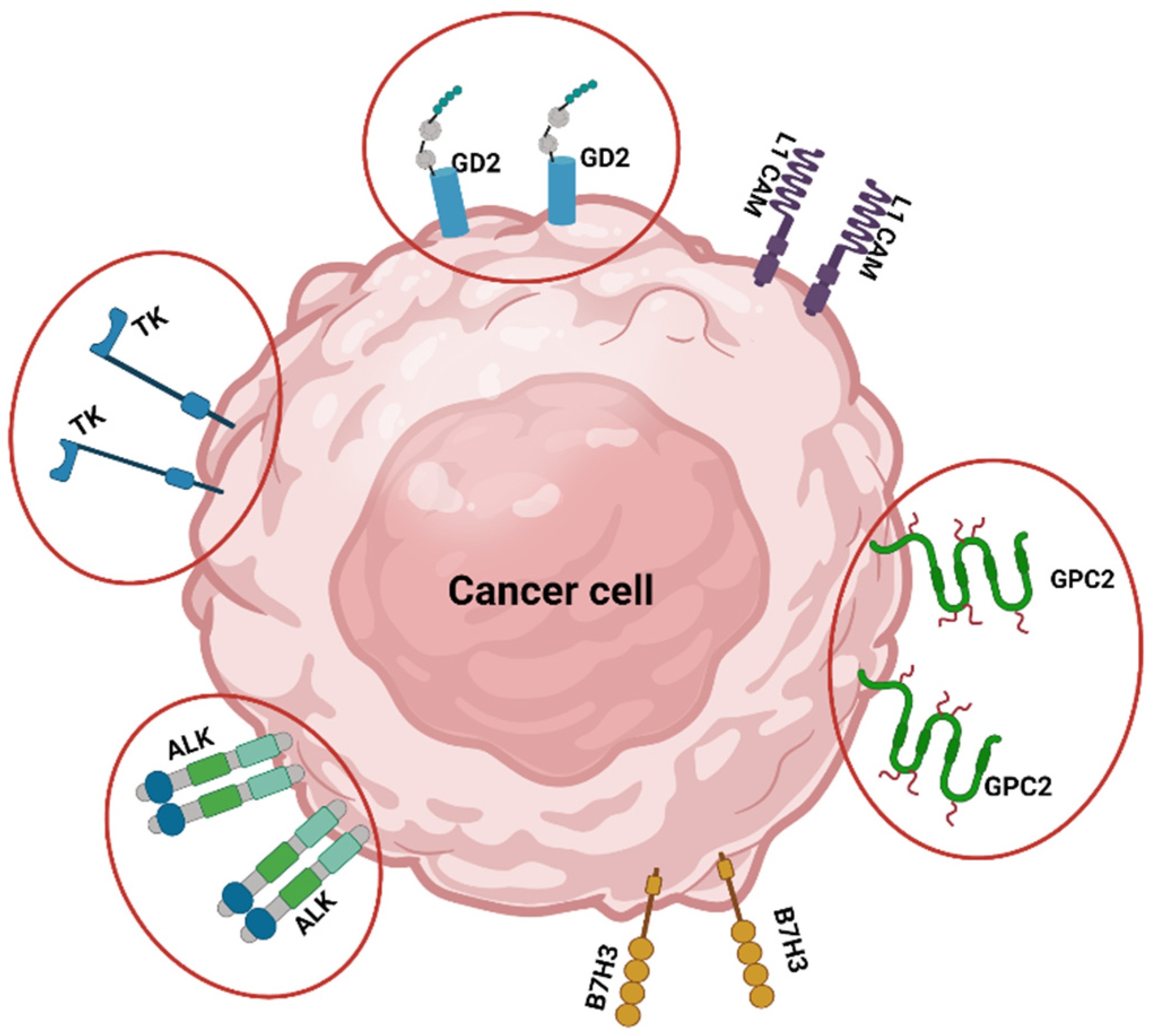
2.1. Monoclonal Antibodies (mAbs)
2.2. Antibody-Drug Conjugates (ADC)s-Based Therapy
2.3. Third-Generation Tyrosine Kinase Inhibitor (TKi)
2.4. Drug-Loaded Nanoparticles
3. Cellular Immunotherapy
4. Tumour Vaccines
5. Radiation Therapy
5.1. Proton Beam Therapy (PBT)
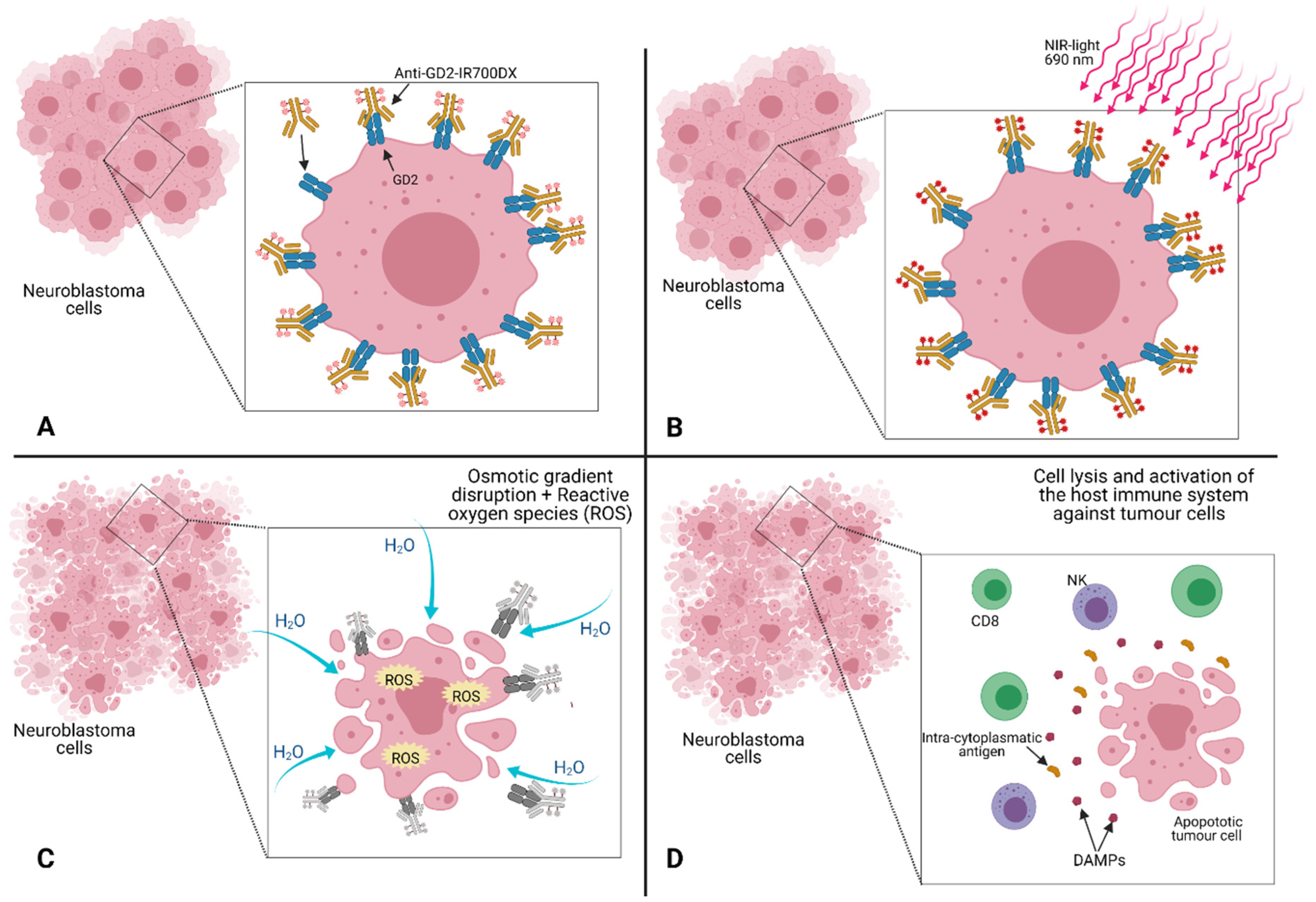
5.2. Near-Infrared Photoimmunotherapy (NIR-PIT)
5.3. Radioisotope Based Radiation
6. Intra-Operative Treatments
6.1. Targeted Probes for Surgery
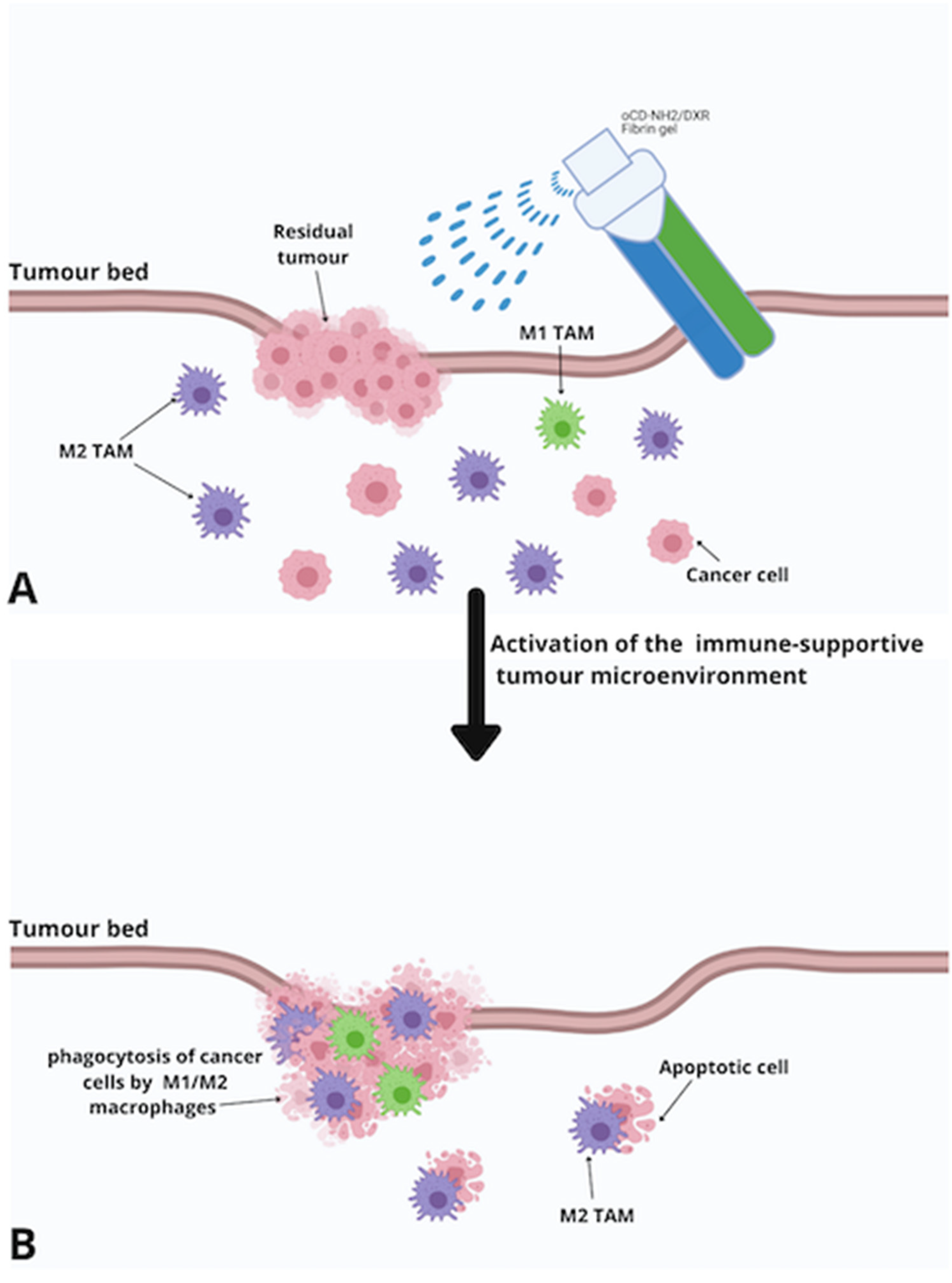
6.2. Fibrin Gels (FBGs)
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Maris, J.M. Recent Advances in Neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef] [Green Version]
- Gatta, G.; Botta, L.; Rossi, S.; Aareleid, T.; Bielska-Lasota, M.; Clavel, J.; Dimitrova, N.; Jakab, Z.; Kaatsch, P.; Lacour, B.; et al. Childhood cancer survival in Europe 1999–2007: Results of EUROCARE-5—A population-based study. Lancet Oncol. 2014, 15, 35–47. [Google Scholar] [CrossRef]
- Tas, M.L.; Reedijk, A.M.J.; Karim-Kos, H.E.; Kremer, L.C.M.; Vand de Ven, C.P.; Dierselhuis, M.P.; Van Eijkelenburg, N.K.A.; Van Grotel, M.; Kraal, K.C.J.M.; Peek, A.M.L.; et al. Neuroblastoma between 1990 and 2014 in the Netherlands: Increased incidence and improved survival of high-risk Neuroblastoma. Eur. J. Cancer 2020, 124, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Siebert, N.; Zumpe, M.; Jüttner, M.; Troschke-Meurer, S.; Lode, H.N. PD-1 blockade augments anti-neuroblastoma immune response induced by anti-GD2 antibody ch14.18/CHO. Oncoimmunology 2017, 6, e1343775. [Google Scholar] [CrossRef] [Green Version]
- Tran, H.C.; Wan, Z.; Sheard, M.A.; Sheard, M.A.; Sun, J.; Jackson, J.R.; Malvar, J.; Xu, Y.; Wang, L.; Sposto, R.; et al. TGFβR1 Blockade with Galunisertib (LY2157299) Enhances Anti-Neuroblastoma Activity of the Anti-GD2 Antibody Dinutuximab (ch14.18) with Natural Killer Cells. Clin. Cancer Res. 2017, 23, 804–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.; Fest, S.; Kunert, R.; Katinger, H.; Pistoia, V.; Michon, J.; Lewis, G.; Ladenstein, R.; Lode, H.N. Anti-neuroblastoma effect of ch14.18 antibody produced in CHO cells is mediated by NK-cells in mice. Mol. Immunol. 2005, 42, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Croce, M.; Corrias, M.V.; Orengo, A.M.; Brizzolara, A.; Carlini, B.; Borghi, M.; Rigo, V.; Pistoia, V.; Ferrini, S. Transient depletion of CD4+ T cells augments IL-21-based immunotherapy of disseminated Neuroblastoma in syngeneic mice. Int. J. Cancer 2009, 127, 1141–1150. [Google Scholar] [CrossRef]
- Rigo, V.; Emionite, L.; Daga, A.; Astigiano, S.; Corrias, M.V.; Quintarelli, C.; Locatelli, F.; Ferrini, S.; Croce, M. Combined immunotherapy with anti-PDL-1/PD-1 and anti-CD4 antibodies cures syngeneic disseminated Neuroblastoma. Sci. Rep. 2017, 7, 14049. [Google Scholar] [CrossRef] [Green Version]
- Bosse, K.R.; Raman, P.; Zhu, Z.; Lane, M.; Martinez, D.; Heitzeneder, S.; Rathi, K.S.; Kendersky, N.M.; Randall, M.; Donovan, L.; et al. Identification of GPC2 as an Oncoprotein and Candidate Immunotherapeutic Target in High-Risk Neuroblastoma. Cancer Cell 2017, 32, 295–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, R.; Krytska, K.; Larmour, C.E.; Raman, P.; Martinez, D.; Ligon, G.F.; Lillquist, J.S.; Cucchi, U.; Orsini, P.; Rizzi, S.; et al. An antibody-drug conjugate directed to the ALK receptor demonstrates efficacy in preclinical models of Neuroblastoma. Sci. Transl. Med. 2019, 11, eaau9732. [Google Scholar] [CrossRef] [PubMed]
- Capone, E.; Lamolinara, A.; Pastorino, F.; Gentile, R.; Ponziani, S.; Di Vittorio, G.; D’Agostino, D.; Bibbò, S.; Rossi, C.; Piccolo, E.; et al. Targeting Vesicular LGALS3BP by an Antibody-Drug Conjugate as Novel Therapeutic Strategy for Neuroblastoma. Cancers 2020, 12, 2989. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.; Chen, Z.; Lu, J.; Pan, J.; Yu, Y.; Zhao, Y.; Zhang, H.; Hu, T.; Liu, Q.; et al. Novel multiple tyrosine kinase inhibitor ponatinib inhibits bFGF-activated signaling in neuroblastoma cells and suppresses neuroblastoma growth in vivo. Oncotarget 2017, 8, 5874–5884. [Google Scholar] [CrossRef]
- Whittle, S.B.; Patel, K.; Zhang, L.; Woodfield, S.E.; Du, M.; Smith, V.; Zage, P.E. The novel kinase inhibitor ponatinib is an effective anti-angiogenic agent against Neuroblastoma. Investig. New Drugs 2016, 34, 685–692. [Google Scholar] [CrossRef]
- Corallo, D.; Pastorino, F.; Pantile, M.; Mariotto, E.; Caicci, F.; Viola, G.; Ponzoni, M.; Tonini, G.P.; Aveic, S. Autophagic flux inhibition enhances cytotoxicity of the receptor tyrosine kinase inhibitor ponatinib. J. Exp. Clin. Cancer Res. 2020, 39, 195. [Google Scholar] [CrossRef] [PubMed]
- Yadav, D.; Sandeep, K.; Pandey, D.; Dutta, R.K. Liposomes for Drug Delivery. J. Biotechnol. Biomater. 2017, 7. [Google Scholar] [CrossRef]
- Pastorino, F.; Brignole, C.; Di Paolo, D.; Perri, P.; Curnis, F.; Corti, A.; Ponzoni, M. Overcoming Biological Barriers in Neuroblastoma Therapy: The Vascular Targeting Approach with Liposomal Drug Nanocarriers. Small 2019, 15, e1804591. [Google Scholar] [CrossRef]
- Pastorino, F.; Di Paolo, D.; Piccardi, F.; Nico, B.; Ribatti, D.; Daga, A.; Baio, G.; Neumaier, C.E.; Brignole, C.; Loi, M.; et al. Enhanced antitumor efficacy of clinical-grade vasculature-targeted liposomal doxorubicin. Clin. Cancer Res. 2008, 14, 7320–7329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Paolo, D.; Pastorino, F.; Brignole, C.; Corrias, M.V.; Emionite, L.; Cilli, M.; Tamma, R.; Priddy, L.; Amaro, A.; Ferrari, D.; et al. Combined Replenishment of miR-34a and let-7b by Targeted Nanoparticles Inhibits Tumor Growth in Neuroblastoma Preclinical Models. Small 2020, 16, 1906426. [Google Scholar] [CrossRef] [PubMed]
- Pule, M.A.; Savoldo, B.; Myers, G.D.; Rossig, C.; Russel, H.V.; Dotti, G.; Huls, M.H.; Liu, E.; Gee, A.P.; Mei, Z.; et al. Virus-specific T cells engineered to coexpress tumor-specific receptors: Persistence and antitumor activity in individuals with Neuroblastoma. Nat. Med. 2008, 14, 1264–1270. [Google Scholar] [CrossRef]
- Louis, C.U.; Savoldo, B.; Dotti, G.; Pule, M.; Yvon, E.; Myers, G.D.; Rossig, C.; Russel, H.V.; Diouf, O.; Liu, E.; et al. Antitumor activity and long-term fate of chimeric antigen receptor–positive T cells in patients with Neuroblastoma. Blood 2011, 118, 6050–6056. [Google Scholar] [CrossRef]
- Heczey, A.; Louis, C.U.; Savoldo, B.; Dakhova, O.; Durett, A.; Grilley, B.; Liu, H.; Wu, M.F.; Mei, Z.; Gee, A.; et al. CAR T Cells Administered in Combination with Lymphodepletion and PD-1 Inhibition to Patients with Neuroblastoma. Mol. Ther. 2017, 25, 2214–2224. [Google Scholar] [CrossRef] [Green Version]
- Straathof, K.; Flutter, B.; Wallace, R.; Jain, N.; Loka, T.; Depani, S.; Wright, G.; Thomas, S.; Cheung, G.W.K.; Gileadi, T.; et al. Antitumor activity without on-target off-tumor toxicity of GD2–chimeric antigen receptor T cells in patients with Neuroblastoma. Sci. Transl. Med. 2020, 12, eabd6169. [Google Scholar] [CrossRef]
- Bocca, P.; Di Carlo, E.; Caruana, I.; Emionite, L.; Cilli, M.; De Angelis, B.; Quintarelli, C.; Pezzolo, A.; Raffaghello, L.; Morandi, F.; et al. Bevacizumab-mediated tumor vasculature remodelling improves tumor infiltration and antitumor efficacy of GD2-CAR T cells in a human Neuroblastoma preclinical model. Oncoimmunology 2018, 7, e1378843. [Google Scholar] [CrossRef]
- Di Carlo, E.; Bocca, P.; Emionite, L.; Cilli, M.; Cipollone, G.; Morandi, F.; Raffaghello, L.; Pistoia, V.; Prigione, I. Mechanisms of the Antitumor Activity of Human Vγ9Vδ2 T Cells in Combination with Zoledronic Acid in a Preclinical Model of Neuroblastoma. Mol. Ther. 2013, 21, 1034–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, J.P.H.; Flutter, B.; Wesemann, F.; Frosch, J.; Rossig, C.; Gustafsson, K.; Anderson, J. Effective combination treatment of GD2-expressing Neuroblastoma and Ewing’s sarcoma using anti-GD2 ch14.18/CHO antibody with Vγ9Vδ2+ γδT cells. Oncoimmunology 2016, 5, e1025194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spel, L.; Boelens, J.-J.; van der Steen, D.M.; Blokland, N.J.G.; Van Noesel, M.M.; Molenaar, J.J.; Heemskerk, M.H.M.; Boes, M.; Nierkens, S. Natural killer cells facilitate PRAME-specific T-cell reactivity against Neuroblastoma. Oncotarget 2015, 6, 35770–35781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castriconi, R.; Dondero, A.; Cilli, M.; Ognio, E.; Pezzolo, A.; De Giovanni, B.; Gambini, C.; Pistoia, V.; Moretta, L.; Moretta, A.; et al. Human NK cell infusions prolong survival of metastatic human Neuroblastoma-bearing NOD/scid mice. Cancer Immunol. Immunother. 2007, 56, 1733–1742. [Google Scholar] [CrossRef]
- Siemann, D.W.; Chaplin, D.J.; Horsman, M.R. Vascular-targeting therapies for treatment of malignant disease. Cancer 2004, 100, 2491–2499. [Google Scholar] [CrossRef]
- Chaplin, D.J.; Horsman, M.R.; Siemann, D.W. Current development status of small-molecule vascular disrupting agents. Curr. Opin. Investig. Drugs 2006, 7, 522–528. [Google Scholar]
- Loi, M.; Marchiò, S.; Becherini, P.; Di Paolo, D.; Soster, M.; Curnis, F.; Brignole, C.; Pagna, G.; Perri, P.; Caffa, I. Combined targeting of perivascular and endothelial tumor cells enhances anti-tumor efficacy of liposomal chemotherapy in Neuroblastoma. J. Control. Release 2010, 145, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Marchiò, S.; Lahdenranta, J.; Schlingemann, R.O.; Valdembri, D.; Wesseling, P.; Arap, M.A.; Hajitou, A.; Ozawa, M.G.; Trepel, M.; Giordano, R.J.; et al. Aminopeptidase A is a functional target in angiogenic blood vessels. Cancer Cell 2004, 5, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Schlingemann, R.O.; Oosterwijk, E.; Wesseling, P.; Rietveld, F.J.; Ruiter, D.J. Aminopeptidase a is a constituent of activated pericytes in angiogenesis. J. Pathol. 1996, 179, 436–442. [Google Scholar] [CrossRef]
- Pastorino, F.; Brignole, C.; Marimpietri, D.; Sapra, P.; Moase, E.H.; Allen, T.M.; Ponzoni, M. Doxorubicin-loaded Fab’ Fragments of Anti-disialoganglioside Immunoliposomes Selectively Inhibit the Growth and Dissemination of Human Neuroblastoma in Nude Mice. Cancer Res. 2003, 63, 86. [Google Scholar]
- Zuccari, G.; Milelli, A.; Pastorino, F.; Loi, M.; Petretto, A.; Parise, A.; Marchetti, C.; Minarini, A.; Cilli, M.; Emionite, L.; et al. Tumor vascular targeted liposomal-bortezomib minimises side effects and increases therapeutic activity in human Neuroblastoma. J. Control. Release 2015, 211, 44–52. [Google Scholar] [CrossRef]
- Bauer, D.F.; Pereboeva, L.; Gillespie, G.Y.; Cloud, G.A.; Elzafarany, O.; Langford, C.; Market, J.M.; Lawrence, S.L. Effect of HSV-IL12 Loaded Tumor Cell-Based Vaccination in a Mouse Model of High-Grade Neuroblastoma. J. Immunol. Res. 2016, 2016, 2568125. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, L.; Morgan, C.; Sandler, A.D.; Bai, X. Combination of Id2 Knockdown Whole Tumor Cells and Checkpoint Blockade: A Potent Vaccine Strategy in a Mouse Neuroblastoma Model. PLoS ONE 2015, 10, e0129237. [Google Scholar] [CrossRef] [PubMed]
- Berger, E.; Soldati, R.; Huebener, N.; Hohn, O.; Stermann, A.; Durmus, T.; Lobitz, S.; Zenclussen, A.C.; Christiansen, H.; Lode, H.N.; et al. Salmonella SL7207 application is the most effective DNA vaccine delivery method for successful tumor eradication in a murine model for Neuroblastoma. Cancer Lett. 2013, 331, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Fest, S.; Huebener, N.; Bleeke, M.; Durmus, T.; Stermann, A.; Woehler, A.; Baykan, B.; Zenclussen, A.C.; Michalsky, E.; Jaeger, I.S.; et al. Survivin minigene DNA vaccination is effective against Neuroblastoma. Int. J. Cancer 2009, 125, 104–114. [Google Scholar] [CrossRef]
- Huebener, N.; Fest, S.; Hilt, K.; Schramm, A.; Eggert, A.; Durmus, T.; Woehler, A.; Stermann, A.; Bleeke, M.; Baykan, B.; et al. Xenogeneic immunisation with human tyrosine hydroxylase DNA vaccines suppresses growth of established Neuroblastoma. Mol. Cancer Ther. 2009, 8, 2392–2401. [Google Scholar] [CrossRef] [Green Version]
- Gil, M.; Bieniasz, M.; Wierzbicki, A.; Bambach, B.J.; Rokita, H.; Kozbor, D. Targeting a mimotope vaccine to activating Fcgamma receptors empowers dendritic cells to prime specific CD8+ T cell responses in tumor-bearing mice. J. Immunol. 2009, 183, 6808–6818. [Google Scholar] [CrossRef] [Green Version]
- Cheung, I.Y.; Cheung, N.-K.V.; Modak, S.; Mauguen, A.; Feng, Y.; Basu, E.; Roberts, S.S.; Ragupathi, G.; Kushner, B.H. Survival Impact of Anti-GD2 Antibody Response in a Phase II Ganglioside Vaccine Trial Among Patients with High-Risk Neuroblastoma with Prior Disease Progression. J. Clin. Oncol. 2021, 39, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, J.; Krieg, A.M. Immunotherapeutic applications of CpG oligodeoxynucleotide TLR9 agonists. Adv. Drug Deliv. Rev. 2009, 61, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Brignole, C.; Pastorino, F.; Marimpietri, D.; Pagnan, G.; Pistorio, A.; Allen, T.M.; Pistoia, V.; Ponzoni, M. Immune cell-mediated antitumor activities of GD2-targeted liposomal c-myb antisense oligonucleotides containing CpG motifs. J. Natl. Cancer Inst. 2004, 96, 1171–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brignole, C.; Marimpietri, D.; Pastorino, F.; Di Paolo, D.; Pagnan, G.; Loi, M.; Piccardi, F.; Cilli, M.; Tradori-Cappai, A.; Arrigoni, G.; et al. Anti-IL-10R antibody improves the therapeutic efficacy of targeted liposomal oligonucleotides. J. Control. Release 2009, 138, 122–127. [Google Scholar] [CrossRef]
- Zhao, Q.; Liu, Y.; Zhang, Y.; Meng, L.; Wei, J.; Wang, B.; Wang, H.; Xin, Y.; Dong, L.; Jiang, X. Role and toxicity of radiation therapy in neuroblastoma patients: A literature review. Crit. Rev. Oncol. Hematol. 2020, 149, 102924. [Google Scholar] [CrossRef]
- Hill-Kayser, C.E.; Tochner, Z.; Li, Y.; Kurtz, G.; Lustig, R.A.; James, P.; Balamuth, N.; Womer, R.; Mattei, P.; Grupp, S.; et al. Outcomes After Proton Therapy for Treatment of Pediatric High-Risk Neuroblastoma. Int. J. Radiat. Oncol. Biol. Phys. 2019, 104, 401–408. [Google Scholar] [CrossRef]
- Bagley, A.F.; Grosshans, D.R.; Philip, N.V.; Foster, J.; McAleer, M.F.; McGovern, S.L.; Lassen-Ramshad, Y.; Mahajan, A.; Paulino, A.C. Efficacy of proton therapy in children with high-risk and locally recurrent neuroblastoma. Pediatr. Blood Cancer 2019, 66. [Google Scholar] [CrossRef] [PubMed]
- Jazmati, D.; Butzer, S.; Hero, B.; Ahmad Khalil, D.; Merta, J.; Baumer, C.; Plum, G.; Fuchs, J.; Koerber, F.; Steinmeier, T.; et al. Proton Beam Therapy for Children with Neuroblastoma: Experiences From the Prospective KiProReg Registry. Front. Oncol. 2021, 10, 617506. [Google Scholar] [CrossRef]
- Nouso, H.; Tazawa, H.; Tanimoto, T.; Tani, M.; Oyama, T.; Sato, H.; Noma, K.; Kagawa, S.; Kobayashi, H.; Noda, T.; et al. Abstract 3831: Development of near-infrared photoimmunotherapy targeting GD2-positive Neuroblastoma. Cancer Res. 2018, 3831. [Google Scholar] [CrossRef]
- Maruoka, Y.; Furusawa, A.; Okada, R.; Inagaki, F.; Wakiyama, H.; Kato, T.; Nagaya, T.; Choyke, P.L.; Kobayashi, H. Interleukin-15 after Near-Infrared Photoimmunotherapy (NIR-PIT) Enhances T Cell Response against Syngeneic Mouse Tumors. Cancers 2020, 12, 2575. [Google Scholar] [CrossRef]
- Gaze, M.N.; Gains, J.E.; Walker, C.; Bomanji, J.B. Optimisation of molecular radiotherapy with [131I]-meta Iodobenzylguanidine for high-risk Neuroblastoma. Q. J. Nucl. Med. Mol. Imaging 2013, 57, 66–78. [Google Scholar]
- Wilson, J.S.; Gains, J.E.; Moroz, V.; Wheatley, K.; Gaze, M.N. A systematic review of 131I-meta iodobenzylguanidine molecular radiotherapy for Neuroblastoma. Eur. J. Cancer 2014, 50, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Corroyer-Dulmont, A.; Falzone, N.; Kersemans, V.; Thompson, J.; Allen, D.P.; Able, S.; Kartsonaki, C.; Malcolm, J.; Kinchesh, P.; Hill, M.A.; et al. Improved outcome of 131 I-mIBG treatment through combination with external beam radiotherapy in the SK-N-SH mouse model of Neuroblastoma. Radiother. Oncol. 2017, 124, 488–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Chen, J.; Wang, N.; Liu, Z.; Li, F.; Zhou, J.; Boyuan, T. Impact of extent of resection on survival in high-risk Neuroblastoma: A systematic review and meta-analysis. J. Pediatr. Surg. 2019, 54, 1487–1494. [Google Scholar] [CrossRef]
- Holmes, K.; Pötschger, U.; Pearson, A.D.J.; Sarnacki, S.; Cecchetto, G.; Gomez-Chacon, J.; Squire, R.; Freud, E.; Bysiek, A.; Matthyssens, L.F.; et al. Influence of Surgical Excision on the Survival of Patients with Stage 4 High-Risk Neuroblastoma: A Report From the HR-NBL1/SIOPEN Study. J. Clin. Oncol. 2020, 38, 2902–2915. [Google Scholar] [CrossRef]
- Wellens, L.M.; Deken, M.M.; Sier, C.F.M.; Johnson, H.R.; De La Jara Ortiz, F.; Bhairosingh, S.S.; Houvast, R.D.; Kholosy, W.M.; Baart, V.M.; Pieters, A.M.M.J.; et al. Anti-GD2-IRDye800CW as a targeted probe for fluorescence-guided surgery in Neuroblastoma. Sci. Rep. 2020, 10, 17667. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Li, Y.; Yang, X.; Tian, J. Neuroblastoma-targeting triangular gadolinium oxide nanoplates for precise excision of cancer. Acta Biomater. 2019, 87, 223–234. [Google Scholar] [CrossRef]
- Chen, Q.; Ci, T.; Gu, Z. Sprayable gel for postsurgical immunotherapy. Immuno-Oncol. Technol. 2019, 2, 11–13. [Google Scholar] [CrossRef] [Green Version]
- Viale, M.; Vecchio, G.; Maric, I.; Cilli, M.; Aprile, A.; Ponzoni, M.; Fontana, V.; Priori, E.C.; Bertone, V.; Rocco, M. Fibrin gels entrapment of a doxorubicin-containing targeted polycyclodextrin: Evaluation of in vivo antitumor activity in orthotopic models of human Neuroblastoma. Toxicol. Appl. Pharmacol. 2019, 385, 114811. [Google Scholar] [CrossRef]
- Viale, M.; Bertone, V.; Maric, I.; Cilli, M.; Emionite, L.; Bocchini, V.; Ponzoni, M.; Fontana, V.; De Luca, F.; Rocco, M. Enhanced therapeutic index of liposomal doxorubicin Myocet locally delivered by fibrin gels in immunodeficient mice bearing human Neuroblastoma. Pharmacol. Res. 2020, 105294. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author, Year | Title | Investigated Treatment | Results |
|---|---|---|---|
| Monoclonal Antibodies | |||
| Siebert N et al. [4], 2017 | PD-1 blockade augments anti-NB immune response induced by anti-GD2 antibody ch14.18/CHO | ch14.18/CHO + anti-PD-1 Ab | ch14.18/CHO + anti-PD-1 Ab results in synergistic treatment effects in mice, representing a new effective treatment strategy against GD2-positive NBs. |
| Croce M et al. [7], 2009 | Transient depletion of CD4+ T cells augments IL-21-based immunotherapy of disseminated NB in syngeneic mice | anti-CD4 Ab | Anti-CD4 Ab potentiated IL-21-based immunotherapy by removing Treg cells, their precursors and other CD4+ cell subsets. This allows the development of an IL-21-driven CD8+ Tcell response, which mediates NB rejection. |
| Rigo V et al. [8], 2017 | Combined immunotherapy withanti-PDL-1/PD-1 and anti-CD4 antibodies cure syngeneic disseminated NB | anti-PD-1/PD-L1 Ab | The combined use of anti-PD-1+ anti-CD4 Ab mediated a potent, CD8-dependent, synergistic effect leading to the elongation of mice’s tumour-free survival, complete tumour regression, and durable anti-NB immunity. |
| Tran et al. [5], 2017 | TGFβR1 Blockade with Galunisertib (LY2157299) Enhances Anti-NB Activity of Anti-GD2 Antibody Dinutuximab (ch14.18) with Natural Killer Cells | ch14.18 + TGFβR1 inhibitor (Galunisertib) | Galunisertib suppresses the activation of SMAD2 in NBs and aNK cells, restores NK cytotoxic mechanisms, and increases the efficacy of ch14.18 with aNK cells against NBs. |
| Antibody-Drug Conjugates (Adc)S-Based Therapy | |||
| Bosse KR et al. [9], 2017 | Identification of GPC2 as an oncoprotein and candidate immunotherapeutic target in high-risk NB | GPC2 targeting ADC | A GPC2 directed ADC proved to be cytotoxic to GPC2-expressing NB cells in vitro and in vivo. |
| Sano R et al. [10], 2019 | An antibody-drug conjugate directed to the ALK receptor demonstrates efficacy in preclinical models of NB | ALK targeting ADC (CDX-0125-TEI) | CDX-0125-TEI exhibited efficient antigen binding, internalisation and cytotoxicity in cells with different ALK expression. In vivo studies showed that CDX-0125-TEI is effective against ALK wild-type and mutant patient-derived xenograft models. |
| Capone E et al. [11], 2020 | Targeting vesicular LGALS3BP by an antibody-drug conjugate as a novel therapeutic strategy for NB | LGALS3BP targeting ADC (1959-sss/DM3) | LGALS3BP targeting ADC can cure mice with established NB tumours in pseudometastatic, orthotopic and PDX models. |
| Third-Generation Tyrosine Kinase Inhibitor (Tki) | |||
| Li et al. [12], 2017 | Novel multiple tyrosine kinase inhibitor ponatinib inhibits bFGF- activated signalling in NB cells and suppresses NB growth in vivo | Ponatinib | Ponatinib can inhibit tumour growth as a single agent or combined with other therapeutic agents, such as doxorubicin. |
| Whittle et al. [13], 2016 | The novel kinase inhibitor ponatinib is an effective anti-angiogenic agent against NB. | Ponatinib | Ponatinib reduces NB cell viability in vitro and reduces tumour growth and vascularity in vivo. |
| Corallo et al. [14], 2020 | Autophagic flux inhibition enhances cytotoxicity of the receptor tyrosine kinase inhibitor ponatinib. | Ponatinib | Inhibition of autophagic flux by CQ restores the cytotoxic potential of PON. In vivo, the use of CQ as adjuvant therapy significantly improves the anti-tumour effects obtained by ponatinib, leading to ulterior reduction of tumour sizes. |
| Drug-Loaded Nanoparticles | |||
| Pastorino F et al. [17], 2008 | Enhanced anti-tumour efficacy of clinical-grade vasculature-targeted liposomal doxorubicin. | CD13-targeted liposomal doxorubicin | TVT-DOX proves to be effective in reducing cell proliferation, blood vessel density, and microvessel area, showing increased cell apoptosis. |
| Di Paolo et al. [18], 2020 | Combined Replenishment of miR-34a and let-7b by targeted nanoparticles inhibits tumour growth in NB preclinical models. | GD2-targeted liposomes entrapping miR-34a and let-7b | The replenishment of miR-34a and let-7b by NB-targeted nanoparticles, individually and more powerfully in combination, significantly reduces cell division, proliferation, neoangiogenesis, and tumour growth, induces apoptosis in orthotopic xenografts, and improves mice survival in pseudometastatic models. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paraboschi, I.; Privitera, L.; Kramer-Marek, G.; Anderson, J.; Giuliani, S. Novel Treatments and Technologies Applied to the Cure of Neuroblastoma. Children 2021, 8, 482. https://doi.org/10.3390/children8060482
Paraboschi I, Privitera L, Kramer-Marek G, Anderson J, Giuliani S. Novel Treatments and Technologies Applied to the Cure of Neuroblastoma. Children. 2021; 8(6):482. https://doi.org/10.3390/children8060482
Chicago/Turabian StyleParaboschi, Irene, Laura Privitera, Gabriela Kramer-Marek, John Anderson, and Stefano Giuliani. 2021. "Novel Treatments and Technologies Applied to the Cure of Neuroblastoma" Children 8, no. 6: 482. https://doi.org/10.3390/children8060482
APA StyleParaboschi, I., Privitera, L., Kramer-Marek, G., Anderson, J., & Giuliani, S. (2021). Novel Treatments and Technologies Applied to the Cure of Neuroblastoma. Children, 8(6), 482. https://doi.org/10.3390/children8060482