
Integration of Vestibular and Auditory Information in Ontogenesis

Abstract
:1. Introduction
2. Materials and Methods
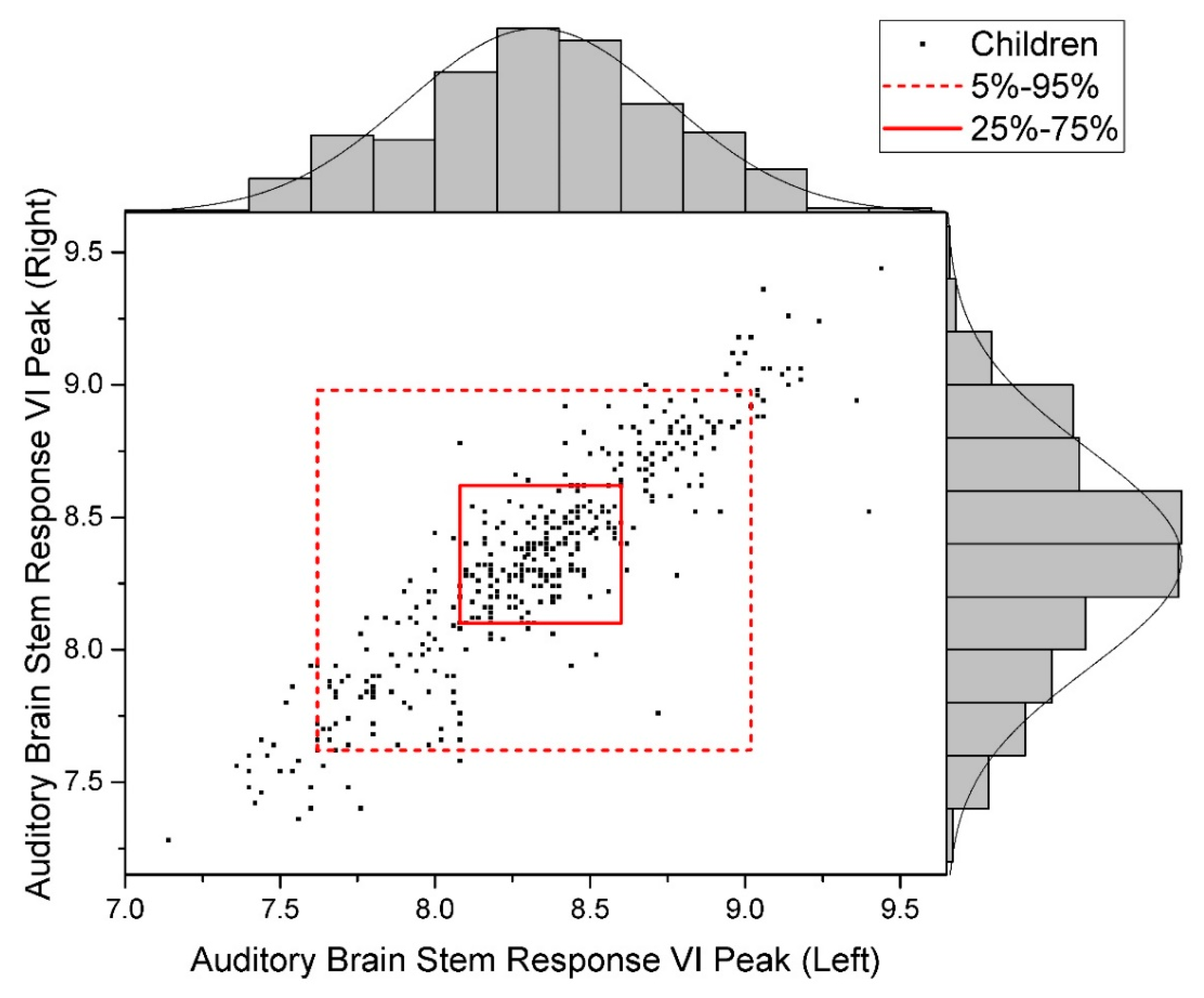
2.1. Assessment of the Conduct of Auditory Information
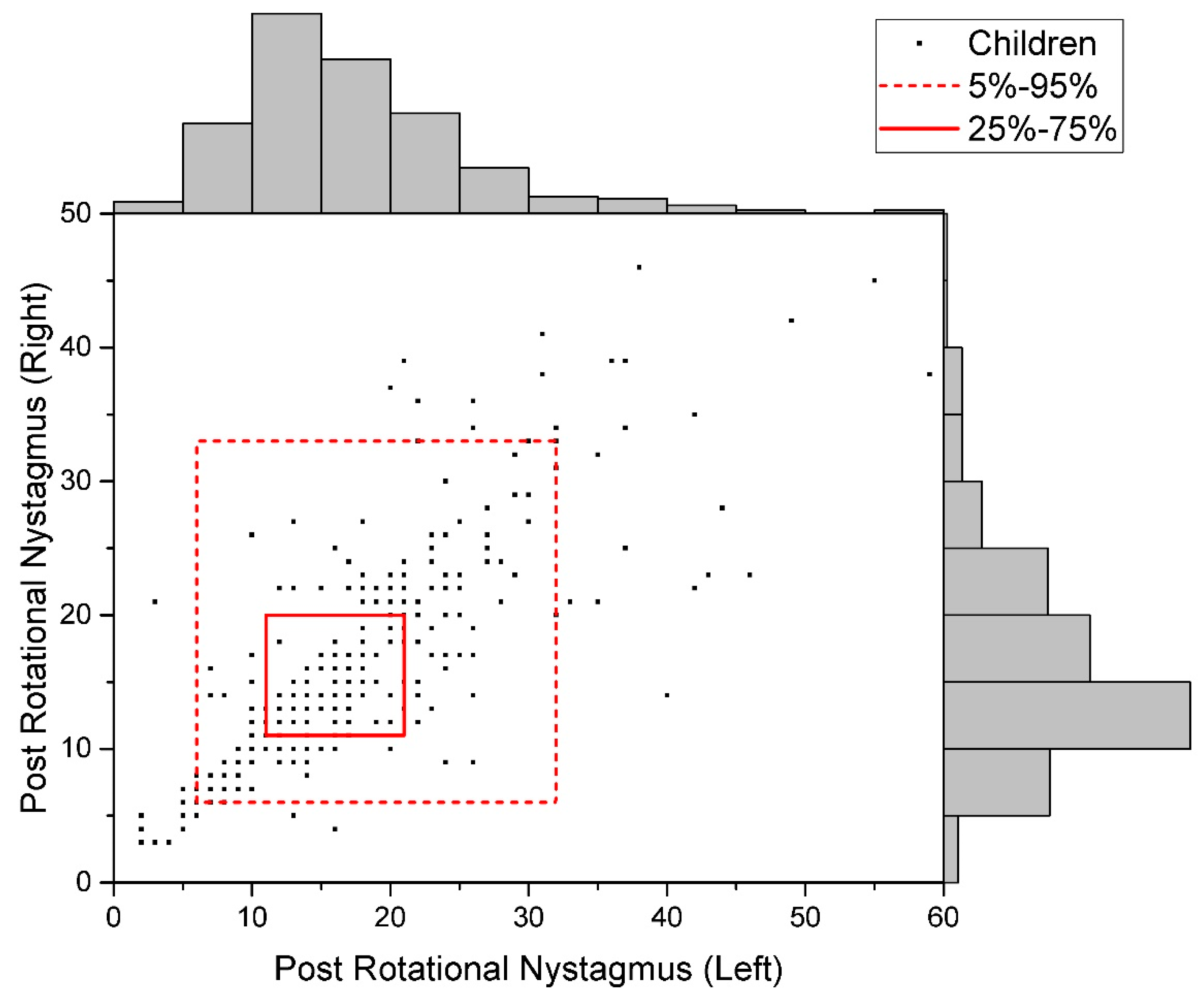
2.2. Post-Rotational Nystagmus with the Registration of the Electrooculogram
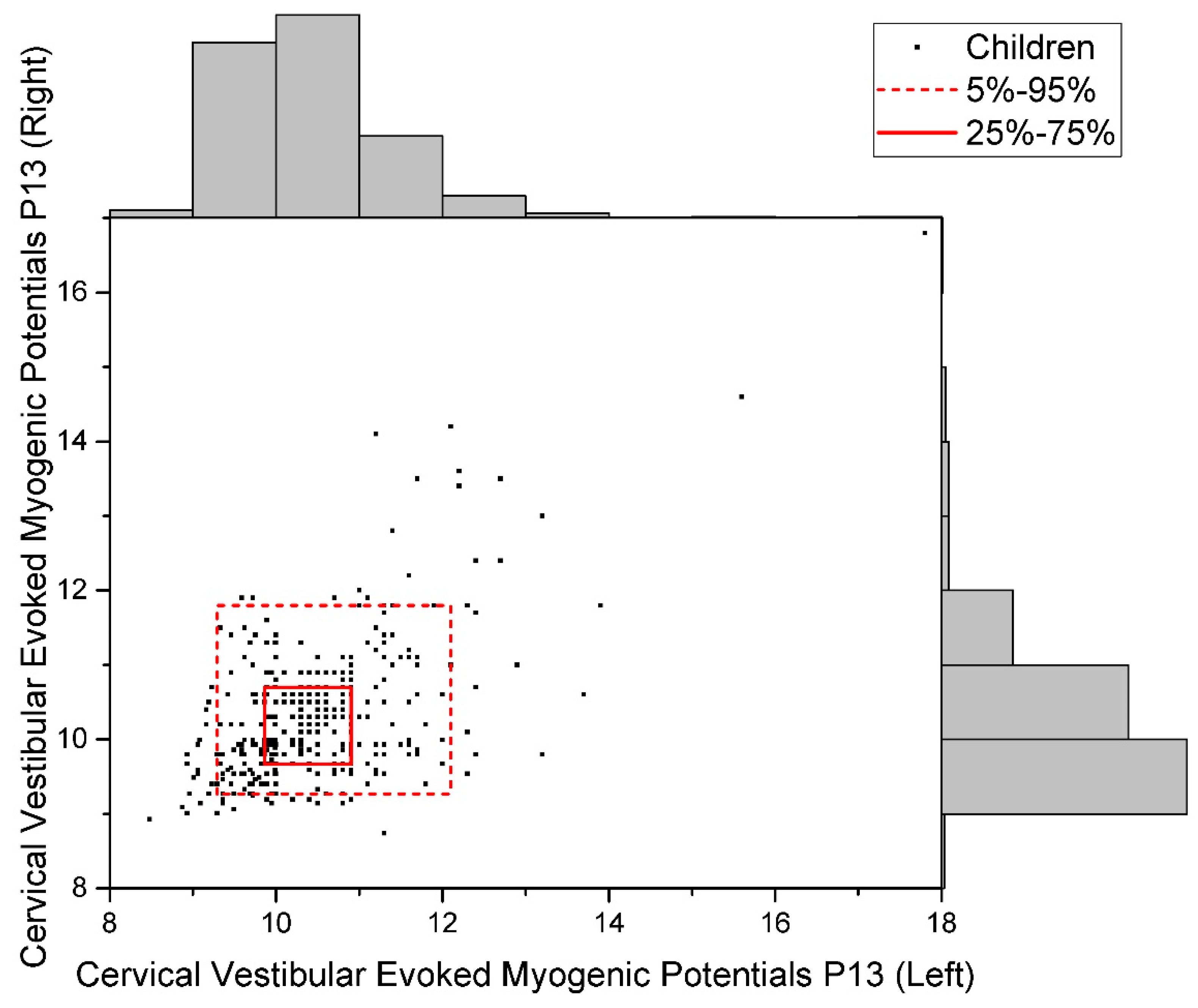
2.3. Cervical Vestibular Evoked Myogenic Potentials (cVEMP)
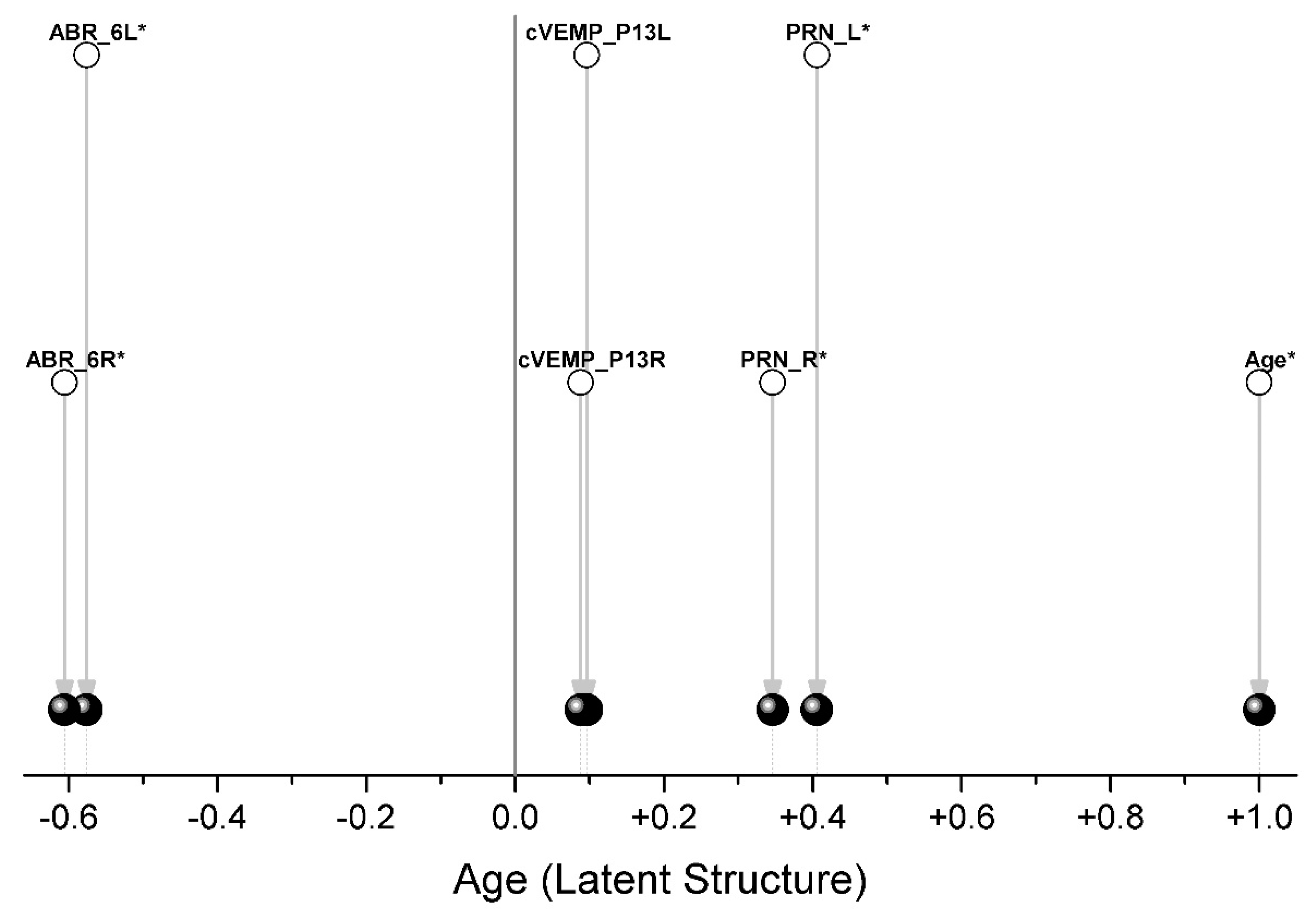
2.4. The Assessment of the Integration of the Parameters
3. Results
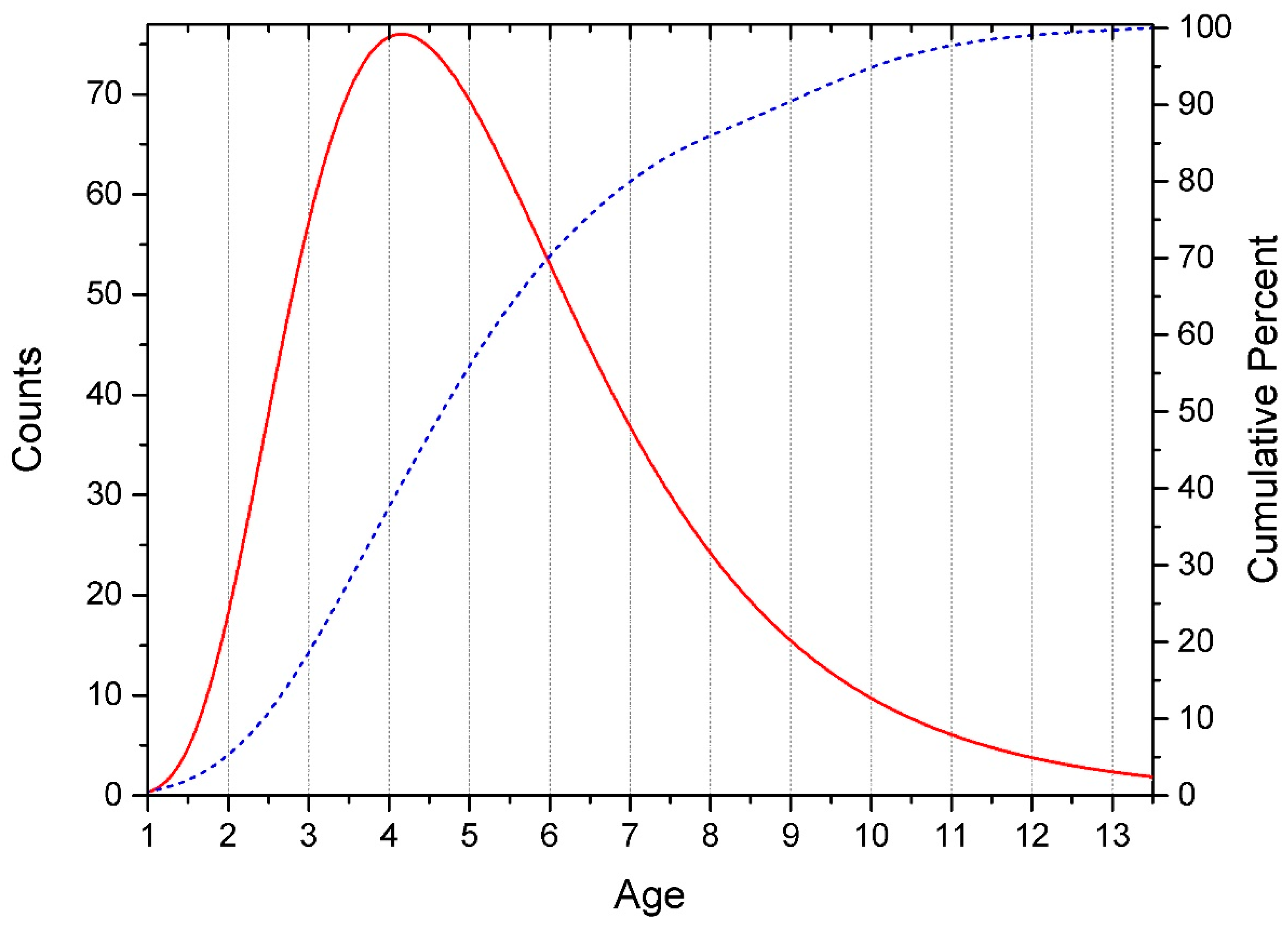
3.1. Data Preprocessing
- -
- for Normal Distribution PKS << 0.01 (Reject Normal) and PAD = 5.7 × 1018 (Reject Normal),
- -
- for Lognormal Distribution PKS > 0.15 (Can’t reject Lognormal) and PAD = 0.090 (Can’t reject Lognormal).
3.2. Data Processing
4. Discussion
5. Conclusions
Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, L.; Stepchenkova, S.; Kirilenko, A.P. Will the present younger adults become future orbital space tourists? Tour. Recreat. Res. 2021, 46, 109–123. [Google Scholar] [CrossRef]
- Pal, N.; Goswami, S.; Singh, R.; Yadav, T.; Singh, R.P. Precautions & Possible Therapeutic Approaches of Health Hazards of Astronauts in Microgravity. Int. J. Aerosp. Psychol. 2021, 31, 149–161. [Google Scholar] [CrossRef]
- Efimova, V.L.; Nikolaeva, E.I. The relationship of language and intelligence development to the maturity of the subcortical structures in children with specific language disorders. Psychol. Russ. State Art 2019, 12, 79–88. [Google Scholar] [CrossRef]
- Nikolaeva, E.I.; Efimova, V.L. The role of the vestibular system in the development of specific language disorders in children. Hum. Physiol. 2020, 46, 306–311. [Google Scholar] [CrossRef]
- Bense, S.; Stephan, T.; Yousry, T.A.; Brandt, T.; Dieterich, M. Multisensory cortical signal increases and decreases during vestibular galvanic stimulation (fMRI). J. Neurophysiol. 2001, 85, 886–899. [Google Scholar] [CrossRef] [PubMed]
- Klingner, C.M.; Volk, G.F.; Flatz, C.; Brodoehl, S.; Dieterich, M.; Witte, O.W.; Guntinas-Lichius, O. Components of vestibular cortical function. Behav. Brain Res. 2013, 236, 194–199. [Google Scholar] [CrossRef]
- Sherwood, C.C.; Bauernfeind, A.L.; Bianchi, S.; Raghanti, M.A.; Hof, P.R. Human Brain Evolution. Writ Large and Small. Prog. Brain Res. 2012, 195, 237–254. [Google Scholar] [CrossRef]
- Ferrè, R.A.; Haggard, P. Vestibular cognition: State-of-the-art and future directions. Cogn. Neuropsychol. 2020, 37, 413–420. [Google Scholar] [CrossRef]
- Raphan, T.; Cohen, B. The vestibulo-ocular reflex in three dimensions. Exp. Brain Res. 2002, 145, 1–27. [Google Scholar] [CrossRef]
- Dichgans, J.; Diener, H.C. The contribution of vestibulo-spinal mechanisms to the maintenance of human upright posture. Acta Oto-Laryngol. 1989, 107, 338–345. [Google Scholar] [CrossRef]
- Gu, Y. Vestibular signals in primate cortex for self-motion perception. Curr. Opin. Neurobiol. 2018, 52, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.; Blanke, O.; Mast, F.W. The human vestibular cortex revealed by coordinate-based activation likelihood estimation meta-analysis. Neuroscience 2012, 212, 159–179. [Google Scholar] [CrossRef] [PubMed]
- zu Eulenburg, P.; Caspers, S.; Roski, C.; Eickhoff, S.B. Meta-analytical definition and functional connectivity of the human vestibular cortex. Neuroimage 2012, 60, 162–169. [Google Scholar] [CrossRef]
- Bense, S.; Bartenstein, P.; Lutz, S.; Stephan, T.; Schwaiger, M.; Brandt, T.; Dieterich, M. Three determinants of vestibular hemispheric dominance during caloric stimulation: A positron emission tomography study. Ann. N. Y. Acad. Sci. 2003, 1004, 440–445. [Google Scholar] [CrossRef]
- Dieterich, M.; Kirsch, V.; Brandt, T. Right-sided dominance of the bilateral vestibular system in the upper brainstem and thalamus. J. Neurol. 2017, 264, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, M.; Bense, S.; Lutz, S.; Drzezga, A.; Stephan, T.; Bartenstein, P.; Brandt, T. Dominance for vestibular cortical function in the non dominant hemisphere. Cerebr. Cortex 2003, 13, 994–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dieterich, M.; Brandt, T. The bilateral central vestibular system: Its pathways, functions, and disorders. Ann. N. Y. Acad. Sci. 2015, 1343, 10–26. [Google Scholar] [CrossRef]
- Olson, E.C.; Miller, R.L. Morphological Integration; University of Chicago Press: Chicago, IL, USA, 1958. [Google Scholar]
- Klingenberg, C.P. Morphological integration and developmental modularity. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Klingenberg, C.P. Studying morphological integration and modularity at multiple levels: Concepts and analysis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130249. [Google Scholar] [CrossRef]
- Cheverud, J.M. Phenotypic, genetic, and environmental morphological integration in the cranium. Evolution 1982, 36, 499–516. [Google Scholar] [CrossRef]
- Willmore, K.E.; Klingenberg, C.P.; Hallgrimsson, B. The relationship between fluctuating asymmetry and environmental variance in rhesus macaque skulls. Evolution 2005, 59, 898–909. [Google Scholar] [CrossRef] [PubMed]
- Debat, V.; Alibert, P.; David, P.; Paradis, E.; Auffray, J.C. Independence between developmental stability and canalization in the skull of the house mouse. Proc. Biol. Sci. 2000, 267, 423–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingenberg, C.P.; Zaklan, S.D. Morphological integration between developmental compartments in the Drosophila wing. Evolution 2000, 54, 1273–1285. [Google Scholar] [CrossRef]
- Klingenberg, C.P.; Badyaev, A.V.; Sowry, S.M.; Beckwith, N.J. Inferring developmental modularity from morphological integration: Analysis of individual variation and asymmetry in bumblebee wings. Am. Nat. 2001, 157, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Ivanovic, A.; Kalezic, M.L. Testing the hypothesis of morphological integration on a skull of a vertebrate with a biphasic life cycle: A case study of the alpine newt. J. Exp. Zool. B Mol. Dev. Evol. 2010, 314, 527–538. [Google Scholar] [CrossRef]
- Jojic, V.; Blagojevic, J.; Vujosevic, M. B chromosomes and cranial variability in yellow-necked field mice (Apodemus flavicollis). J. Mammal. 2011, 92, 396–406. [Google Scholar] [CrossRef] [Green Version]
- Wan, G.; Corfas, G. Transient auditory nerve demyelination as a new mechanism for hidden hearing loss. Nat. Commun. 2017, 8, 14487. [Google Scholar] [CrossRef]
- Hashimoto, I.; Ishiyama, Y.; Yoshimoto, T.; Nemoto, S. Brain-stem auditory-evoked potentials recorded directly from human brain-stem and thalamus. Brain 1981, 104 Pt 4, 841–859. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, I. Auditory evoked potentials recorded directly from the human VIIIth nerve and brain stem: Origins of their fast and slow components. Electroencephalogr. Clin. Neurophysiol. Suppl. 1982, 36, 305–314. [Google Scholar]
- Efimov, O.I.; Efimova, V.L.; Rozhkov, V.P. Violation of the speed of the auditory information in the structures of the brain stem in children with speech disorders and learning difficulties. Sens. Syst. 2014, 28, 36–44. (In Russian) [Google Scholar]
- Chiappa, K.H. Evoked Potentials in Clinical Medicine; Raven Press: New York, NY, USA, 1990; Volume XVII, 647p. [Google Scholar]
- Lotfi, Y.; Rezazadeh, N.; Moossavi, A.; Haghgoo, H.A.; Rostami, R.; Bakhshi, E.; Badfar, F.; Moghadam, S.F.; Sadeghi-Firoozabadi, V.; Khodabandelou, Y. Rotational and Collic Vestibular-Evoked Myogenic Potential Testing in Normal Developing Children and Children with Combined Attention Deficit/Hyperactivity Disorder. Ear Hear. 2017, 38, e352–e358. [Google Scholar] [CrossRef] [PubMed]
- Rine, R.M.; Wiener-Vacher, S. Evaluation and treatment of vestibular dysfunction in children. NeuroRehabilitation 2013, 32, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Dargie, J.; Dornan, B.; Whittemore, K. Clinical uses of cervical vestibular-evoked myogenic potential testing in pediatric patients. Medicine 2014, 93, e37. [Google Scholar] [CrossRef] [PubMed]
- Young, Y.-H.; Chen, C.-N.; Hsieh, W.-S.; Wang, S.-J. Development of vestibular evoked myogenic potentials in early life. Eur. J. Paediatr. Neurol. 2009, 13, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Wiener-Vacher, S.R.; Quarez, J.; Le Priol, A. Epidemiology of vestibular impairments in a pediatric population. Semin. Hear. 2018, 39, 229–242. [Google Scholar] [CrossRef] [Green Version]
- Polunin, D.; Shtaiger, I.; Efimov, V. JACOBI4 software for multivariate analysis of biological data. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.J.; Corti, M. The use of two-block partial least-squares to study covariation in shape. Syst. Biol. 2000, 49, 740–753. [Google Scholar] [CrossRef] [Green Version]
- Ränner, S.; Lindgren, F.; Geladi, P.; Wold, S. A PLS kernel algorithm for data sets with many variables and fewer objects. J. Chemom. 1994, 8, 111–125. [Google Scholar] [CrossRef]
- Kovaleva, V.Y.; Pozdnyakov, A.A.; Litvinov, Y.N.; Efimov, V.M. Assessment of the conjugation of morphogenetic and molecular genetic moduli of variation in the common vole Microtus s.l. in gradient environmental conditions. Ecol. Genet. 2019, 17, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Salmaso, D.; Longoni, A.M. Problems in the assessment of hand preference. Cortex 1985, 21, 533–549. [Google Scholar] [CrossRef]
- Kirsch, V.; Boegle, R.; Keeser, D.; Kierig, E.; Ertl-Wagner, B.; Brandt, T.; Dieterich, M. Handedness-dependent functional organizational patterns within the bilateral vestibular cortical network revealed by fMRI connectivity based parcellation. NeuroImage 2018, 178, 224–237. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.M.; Baumann, O.; Mattingley, J.B.; Greenlee, M.W. Vestibular and visual responses in human posterior insular cortex. J. Neurophysiol. 2014, 112, 2481–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Preuss, T.M. The human brain: Rewired and running hot. Ann. N. Y. Acad. Sci. 2011, 1255, E182–E191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, T.; Dieterich, M. The dizzy patient: Don’t forget disorders of the central vestibular system. Nat. Rev. Neurol. 2017, 13, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Newman, N.; Latifi, A. Vygotsky, education, and teacher education. J. Educ. Teach. 2021, 47, 4–17. [Google Scholar] [CrossRef]
- Efimova, V.; Efimov, O.; Dudetskaia, T.; Rozhkov, V. Sensory motor disorders of children with language impairments. In Neurobiology of Speech and Language; Skifiya-Print: Saint-Petersburg, Russia, 2018; p. 53. (In Russian) [Google Scholar]
- Siegel, D.J. The Developing Mind. How Relationships and Brain Interact to Shape Who We Are; Guilford Press: New York, NY, USA, 2012. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Integrative Functions | Left vs. Right |
|---|---|
| Auditory Brain Stem Response, VI Peak | r = +0.907; R2 = 0.82 |
| Post-Rotational Nystagmus | r = +0.803; R2 = 0.64 |
| Cervical Vestibular Evoked Myogenic Potentials, P13 | r = +0.573; R2 = 0.33 |
| Code | Variable | Block |
|---|---|---|
| ABR_6L | Auditory Brain Stem Response, VI Peak, Left | #1 |
| ABR_6R | Auditory Brain Stem Response, VI Peak, Right | #1 |
| PRN_L | Post-Rotational Nystagmus, Left | #1 |
| PRN_R | Post-Rotational Nystagmus, Right | #1 |
| cVEMP_P13L | Cervical Vestibular Evoked Myogenic Potentials, P13, Left | #1 |
| cVEMP_P13R | Cervical Vestibular Evoked Myogenic Potentials, P13, Right | #1 |
| Age | Age | #2 |
| Indicators | Left vs. Right | PRN | cVEMP_P13 |
|---|---|---|---|
| Auditory Brain Stem Response, VI Peak | not found | p < 0.05 | p < 0.05 |
| Post-Rotational Nystagmus | not found | p < 0.05 | |
| Cervical Vestibular Evoked Myogenic Potentials, P13 | not found |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nikolaeva, E.I.; Efimova, V.L.; Vergunov, E.G. Integration of Vestibular and Auditory Information in Ontogenesis. Children 2022, 9, 401. https://doi.org/10.3390/children9030401
Nikolaeva EI, Efimova VL, Vergunov EG. Integration of Vestibular and Auditory Information in Ontogenesis. Children. 2022; 9(3):401. https://doi.org/10.3390/children9030401
Chicago/Turabian StyleNikolaeva, Elena I., Victoria L. Efimova, and Eugeny G. Vergunov. 2022. "Integration of Vestibular and Auditory Information in Ontogenesis" Children 9, no. 3: 401. https://doi.org/10.3390/children9030401
APA StyleNikolaeva, E. I., Efimova, V. L., & Vergunov, E. G. (2022). Integration of Vestibular and Auditory Information in Ontogenesis. Children, 9(3), 401. https://doi.org/10.3390/children9030401