
Protective Effect of Ultrasound-Processed Amazonian Sapota-do-Solimões (Quararibea cordata) Juice on Artemia salina Nauplii

 , , ,
, , , 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Raw Material
2.2. Ultrasound Processing of Sapota-do-Solimões Juice
2.3. Sapota-do-Solimões Juice Lyophilization
2.4. Protective Effects of Sapota-do-Solimões Juice against Hydrogen Peroxide-Induced Toxicity in Artemia salina Nauplii
2.5. Determination of Antioxidant Enzymes on Artemia salina Nauplii after Protective Effect Test
2.5.1. Extract Preparation of Artemia salina Nauplii after Protective Effect Test
2.5.2. Protein Determination of Artemia salina Nauplii after Protective Effect Test
2.5.3. Catalase Activity (CAT) of Artemia salina Nauplii after Protective Effect Test
2.5.4. Superoxide Dismutase Activity (SOD) of Artemia salina Nauplii after Protective Effect Test
2.6. Determination of Lipoperoxidation (LPO) of Artemia salina Nauplii after Protective Effect Test
2.7. Morphological Analysis under Light Microscopy of Artemia salina Nauplii
2.8. Morphological Analysis under Scanning Electron Microscopy of Artemia salina Nauplii
2.9. Statistic
3. Results and Discussion
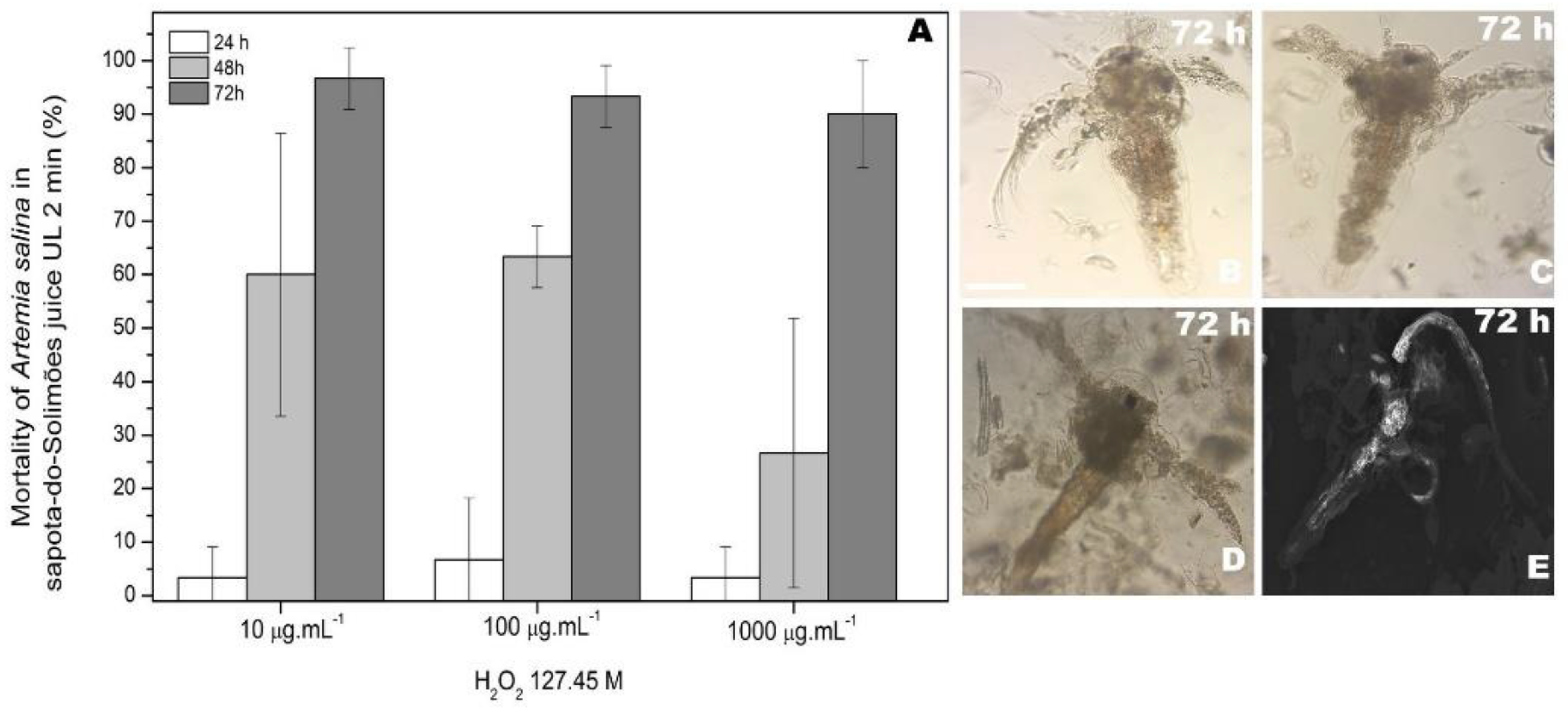
3.1. Toxicity Assay of Ultrasound Processed Sapota-do-Solimões Juice
3.2. Morphological Analysis of Artemia salina Nauplii after Toxicity Assay
3.2.1. Morphological Analysis of Artemia salina Nauplii after Toxicity Assay under an Optical Microscope
3.2.2. Morphological Analysis of A. salina Nauplii after Toxicity Assay under an Electronic Scanning Microscope
3.3. Protective Effect of Unprocessed and Ultrasound-Processed Sapota-do-Solimões Juice against H2O2 Induced Toxicity on Artemia salina
3.4. Evaluation of the Stress Biomarkers on Artemia salina after Protective Effect Assay of Sapota-do-Solimões Juice
3.4.1. Protein Concentration of Artemia salina after Protective Effect Assay of Sapota-do-Solimões Juice
3.4.2. Catalase Activity on Artemia salina Nauplii after Protective Effect Assay of Sapota-do-Solimões Juice
3.4.3. Superoxide Dismutase Activity on Artemia salina Nauplii after Protective Effect Assay of Sapota-do-Solimões Juice
3.4.4. Lipid Peroxidation Level on Artemia salina Nauplii after Protective Effect Assay of Sapota-do-Solimões Juice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alves Filho, E.G.; Sousa, V.M.; Rodrigues, S.; de Brito, E.S.; Fernandes, F.A.N. Green Ultrasound-Assisted Extraction of Chlorogenic Acids from Sweet Potato Peels and Sonochemical Hydrolysis of Caffeoylquinic Acids Derivatives. Ultrason. Sonochem. 2020, 63, 104911. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Sánchez, C.; Lozano-Sánchez, J.; Segura-Carretero, A.; Fernández-Gutiérrez, A. Alternatives to Conventional Thermal Treatments in Fruit-Juice Processing. Part 2: Effect on Composition, Phytochemical Content, and Physicochemical, Rheological, and Organoleptic Properties of Fruit Juices. Crit. Rev. Food Sci. Nutr. 2017, 57, 637–652. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, M.; Yang, C. hui Application of Ultrasound Technology in Processing of Ready-to-Eat Fresh Food: A Review. Ultrason. Sonochem. 2020, 63, 104953. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, Y.; Xie, F.; Gu, X.; Wu, J.; Wang, Z. High Pressure Homogenization versus Ultrasound Treatment of Tomato Juice: Effects on Stability and in Vitro Bioaccessibility of Carotenoids. LWT 2019, 116, 108597. [Google Scholar] [CrossRef]
- Chacha, J.S.; Zhang, L.; Ofoedu, C.E.; Suleiman, R.A.; Dotto, J.M.; Roobab, U.; Agunbiade, A.O.; Duguma, H.T.; Mkojera, B.T.; Hossaini, S.M.; et al. Revisiting Non-Thermal Food Processing and Preservation Methods—Action Mechanisms, Pros and Cons: A Technological Update (2016–2021). Foods 2021, 10, 1430. [Google Scholar] [CrossRef] [PubMed]
- Sanches, M.A.R.; Colombo Silva, P.M.O.; Barretto, T.L.; Darros-Barbosa, R.; da Silva-Barretto, A.C.; Telis-Romero, J. Technological and Diffusion Properties in the Wet Salting of Beef Assisted by Ultrasound. LWT 2021, 149, 112036. [Google Scholar] [CrossRef]
- Campoli, S.S.; Rojas, M.L.; do Amaral, J.E.P.G.; Canniatti-Brazaca, S.G.; Augusto, P.E.D. Ultrasound Processing of Guava Juice: Effect on Structure, Physical Properties and Lycopene in Vitro Accessibility. Food Chem. 2018, 268, 594–601. [Google Scholar] [CrossRef]
- Alves Filho, E.G.; Silva, L.M.A.; de Brito, E.S.; Wurlitzer, N.J.; Fernandes, F.A.N.; Rabelo, M.C.; Fonteles, T.V.; Rodrigues, S. Evaluation of Thermal and Non-Thermal Processing Effect on Non-Prebiotic and Prebiotic Acerola Juices Using 1H QNMR and GC–MS Coupled to Chemometrics. Food Chem. 2018, 265, 23–31. [Google Scholar] [CrossRef]
- de Linhares, M.F.D.; Alves Filho, E.G.; Silva, L.M.A.; Fonteles, T.V.; Wurlitzer, N.J.; de Brito, E.S.; Fernandes, F.A.N.; Rodrigues, S. Thermal and Non-Thermal Processing Effect on Açai Juice Composition. Food Res. Int. 2020, 136, 109506. [Google Scholar] [CrossRef]
- Yıkmış, S.; Erdal, B.; Bozgeyik, E.; Levent, O.; Yinanç, A. Evaluation of Purple Onion Waste from the Perspective of Sustainability in Gastronomy: Ultrasound-Treated Vinegar. Int. J. Gastron. Food Sci. 2022, 29, 100574. [Google Scholar] [CrossRef]
- Maciel da Silva, R.; Henrique Campelo, P.; Rodrigues, S. In Vitro Viability of L. Casei B-442 and Fructooligosaccharides Integrity in Amazonian Sapota-Do-Solimões Functional Juice. Food Res. Int. 2022, 154, 111036. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, S.S.; Ribeiro, S.R.; Soquetta, M.B.; Pires, F.J.; Wagner, R.; da Rosa, C.S. Evaluation of the Chemical, Sensory and Volatile Composition of Sapota-Do-Solimões Pulp at Different Ripening Stages. Food Res. Int. 2018, 109, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, S.S.; Karnopp, G.; Michelon, N.; Arantes, A.C.N.R.; Monego, M.A.; Kipper, D.K.; Soquetta, M.B.; Wagner, R.; da Rosa, C.S. Influence of Preservation by Heat and Cold on the Physicochemical and Microbiological Characteristics, Bioactive Compounds of Pulp from Sapota-Do-Solimões (Quararibea Cordata). CyTA J. Food 2018, 16, 85–95. [Google Scholar] [CrossRef]
- Berto, A.; da Silva, A.F.; Visentainer, J.V.; Matsushita, M.; de Souza, N.E. Proximate Compositions, Mineral Contents and Fatty Acid Compositions of Native Amazonian Fruits. Food Res. Int. 2015, 77, 441–449. [Google Scholar] [CrossRef]
- Berto, A.; Ribeiro, A.B.; de Souza, N.E.; Fernandes, E.; Chisté, R.C. Bioactive Compounds and Scavenging Capacity of Pulp, Peel and Seed Extracts of the Amazonian Fruit Quararibea Cordata against ROS and RNS. Food Res. Int. 2015, 77, 236–243. [Google Scholar] [CrossRef]
- Silva, R.M.; Filho, E.G.A.; Campelo, P.H.; Silva, F.E.F.; Zampieri, D.S.; Gramosa, N.V.; Fernandes, F.A.N.; Rodrigues, S. NMR Spectroscopy and Chemometrics to Evaluate the Effect of Different Non-Thermal Plasma Processing on Sapota-Do-Solimões (Quararibea Cordata Vischer) Juice Quality and Composition. Food Bioprocess Technol. 2022, 15, 875–890. [Google Scholar] [CrossRef]
- Zhu, B.; Zhu, S.; Li, J.; Hui, X.; Wang, G.X. The Developmental Toxicity, Bioaccumulation and Distribution of Oxidized Single Walled Carbon Nanotubes in Artemia Salina. Toxicol. Res. 2018, 7, 897–906. [Google Scholar] [CrossRef]
- Les, F.; Prieto, J.M.; Arbonés-Mainar, J.M.; Valero, M.S.; López, V. Bioactive Properties of Commercialised Pomegranate (Punica Granatum) Juice: Antioxidant, Antiproliferative and Enzyme Inhibiting Activities. Food Funct. 2015, 6, 2049–2057. [Google Scholar] [CrossRef]
- Sousa, E.C.; Uchôa-Thomaz, A.M.A.; Carioca, J.O.B.; de Morais, S.M.; de Lima, A.; Martins, C.G.; Alexandrino, C.D.; Ferreira, P.A.T.; Rodrigues, A.L.M.; Rodrigues, S.P.; et al. Chemical Composition and Bioactive Compounds of Grape Pomace (Vitis vinifera L.), Benitaka Variety, Grown in the Semiarid Region of Northeast Brazil. Food Sci. Technol. 2014, 34, 135–142. [Google Scholar] [CrossRef]
- Nam Pung, S. Effect of Ozone Microbubbles and Ultrasonic Irradiation on Pesticide Detoxification in Tangerine Cv. Int. Food Res. J. 2017, 24, 1135–1139. [Google Scholar]
- Angelini, P.; Flores, G.A.; Piccirilli, A.; Venanzoni, R.; Acquaviva, A.; Di Simone, S.C.; Libero, M.L.; Tirillini, B.; Zengin, G.; Chiavaroli, A.; et al. Polyphenolic Composition and Antimicrobial Activity of Extracts Obtained from Grape Processing By-Products: Between Green Biotechnology and Nutraceutical. Process Biochem. 2022, 118, 84–91. [Google Scholar] [CrossRef]
- Miguel, T.B.A.R.; Porto, E.C.M.; de Paiva Pinheiro, S.K.; de Castro Miguel, E.; Fernandes, F.A.N.; Rodrigues, S. Protective Effect of Natural and Processed Coconut Water by Non-Thermal Technologies Against Oxidative Stress in Brine Shrimp (Artemia salina). Food Bioprocess Technol. 2021, 14, 702–716. [Google Scholar] [CrossRef]
- Tsai, F.-H.; Lien, J.-C.; Lin, L.-W.; Chen, H.-Y.; Ching, H.; Wu, C.-R. Protective Effect of Broussonetia Papyrifera against Hydrogen Peroxide-Induced Oxidative Stress in SH-SY5Y Cells. Biosci. Biotechnol. Biochem. 2009, 73, 90080–90081. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, B.; Maharajan, K.; Hemalatha, D.; Rangasamy, B.; Arul, N.; Ramesh, M. Comparative Toxicity of UV-Filter Octyl Methoxycinnamate and Its Photoproducts on Zebrafish Development. Sci. Total Environ. 2020, 718, 134546. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.; Barros, L.; Ferreira, I.C.F.R. In Vivo Antioxidant Activity of Phenolic Compounds: Facts and Gaps. Trends Food Sci. Technol. 2016, 48, 1–12. [Google Scholar] [CrossRef]
- Demarchi, C.A.; da Silva, L.M.; Niedźwiecka, A.; Ślawska-Waniewska, A.; Lewińska, S.; Dal Magro, J.; Fossá Calisto, J.F.; Martello, R.; Rodrigues, C.A. Nanoecotoxicology Study of the Response of Magnetic O-Carboxymethylchitosan Loaded Silver Nanoparticles on Artemia Salina. Environ. Toxicol. Pharmacol. 2020, 74, 103298. [Google Scholar] [CrossRef]
- Thangal, S.H.; Nivetha, M.; Muttharasi, C.; Anandhan, K.; Muralisankar, T. Effects of Acidified Seawater on Biological and Physiological Responses of Artemia Franciscana. Mar. Pollut. Bull. 2021, 169, 112476. [Google Scholar] [CrossRef]
- Ñañez Pacheco, G.K.; Sanabio Maldonado, N.S.; Pastrana Alta, R.Y.; Aguilar Vitorino, H. Short Exposure of Artemia salina to Group-12 Metals: Comparing Hatchability, Mortality, Lipid Peroxidation, and Swimming Speed. Ecotoxicol. Environ. Saf. 2021, 213, 112052. [Google Scholar] [CrossRef]
- Thiagarajan, V.; Seenivasan, N.; Jenkins, D.; Chandrasekaran, N.; Mukherjee, A. Combined Effects of Nano-TiO2 and Hexavalent Chromium towards Marine Crustacean Artemia Salina. Aquat. Toxicol. 2020, 225, 105541. [Google Scholar] [CrossRef]
- Noguera, N.H.; Lima, D.C.; Filho, E.G.A.; Fonteles, T.V.; Rodrigues, S. Influence of Different Non-Thermal Processing on Guava, Orange, and Tangerine Juices and the Food Matrix Effects. Food Bioprocess Technol. 2021, 14, 1662–1672. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. [13] Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Kono, Y. Generation of Superoxide Radical during Autoxidation of Hydroxylamine and an Assay for Superoxide Dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Hermes-Lima, M.; Willmore, W.G.; Storey, K.B. Quantification of Lipid Peroxidation in Tissue Extracts Based on Fe(III)Xylenol Orange Complex Formation. Free Radic. Biol. Med. 1995, 19, 271–280. [Google Scholar] [CrossRef]
- Vitorino, H.A.; Mantovanelli, L.; Zanotto, F.P.; Espósito, B.P. Iron Metallodrugs: Stability, Redox Activity and Toxicity against Artemia Salina. PLoS ONE 2015, 10, e0121997. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, S.; Oliviero, M.; Li, J.; Manzo, S. Testing ZnO Nanoparticle Ecotoxicity: Linking Time Variable Exposure to Effects on Different Marine Model Organisms. Environ. Sci. Pollut. Res. 2018, 25, 4871–4880. [Google Scholar] [CrossRef] [PubMed]
- Arulvasu, C.; Jennifer, S.M.; Prabhu, D.; Chandhirasekar, D. Toxicity Effect of Silver Nanoparticles in Brine Shrimp Artemia. Sci. World J. 2014, 2014, 256919. [Google Scholar] [CrossRef]
- Silva, L.S.L.F.R.; da Gomes, A.S.; Castro, D.R.G.; Souza, F.d.C.d.A.; Mar, J.M.; Silva, L.S.L.F.R.; Sanches, E.A.; Bezerra, J.d.A.; Bakry, A.M.; Campelo, P.H. Ultrasound-assisted Homogenization and Gum Arabic Combined to Physicochemical Quality of Cupuaçu Juice. J. Food Process. Preserv. 2019, 43, e14072. [Google Scholar] [CrossRef]
- Zhu, S.; Luo, F.; Tu, X.; Chen, W.C.; Zhu, B.; Wang, G.X. Developmental Toxicity of Oxidized Multi-Walled Carbon Nanotubes on Artemia Salina Cysts and Larvae: Uptake, Accumulation, Excretion and Toxic Responses. Environ. Pollut. 2017, 229, 679–687. [Google Scholar] [CrossRef]
- Bergami, E.; Bocci, E.; Vannuccini, M.L.; Monopoli, M.; Salvati, A.; Dawson, K.A.; Corsi, I. Nano-Sized Polystyrene Affects Feeding, Behavior and Physiology of Brine Shrimp Artemia Franciscana Larvae. Ecotoxicol. Environ. Saf. 2016, 123, 18–25. [Google Scholar] [CrossRef]
- Defoirdt, T.; Halet, D.; Sorgeloos, P.; Bossier, P.; Verstraete, W. Short-Chain Fatty Acids Protect Gnotobiotic Artemia Franciscana from Pathogenic Vibrio Campbellii. Aquaculture 2006, 261, 804–808. [Google Scholar] [CrossRef]
- Bhuvaneshwari, M.; Sagar, B.; Doshi, S.; Chandrasekaran, N.; Mukherjee, A. Comparative Study on Toxicity of ZnO and TiO2 Nanoparticles on Artemia Salina: Effect of Pre-UV-A and Visible Light Irradiation. Environ. Sci. Pollut. Res. 2017, 24, 5633–5646. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.Y.; Ryu, B.M.; Yang, H.W.; Kim, E.A.; Lee, J.S.; Jeon, Y.J. Protective Effects of Ecklonia Cava Extract on the Toxicity and Oxidative Stress Induced by Hair Dye in In-Vitro and in-Vivo Models. J. Oceanol. Limnol. 2019, 37, 909–917. [Google Scholar] [CrossRef]
- Taib Mimi Iryani, M.; Lv, A.; Sun, J.; Anirudhan, A.; Pau Tan, M.; Danish-Daniel, M.; Lian Wong, L.; Baruah, K.; Sorgeloos, P.; Sifzizul Tengku Muhammad, T.; et al. Effects of Heat Shock Protein 70 Knockdown on the Tolerance of the Brine Shrimp Artemia Franciscana to Aquaculture-Related Stressors: Implications for Aquatic Animal Health and Production Liqing. Aquaculture 2021, 550, 737872. [Google Scholar] [CrossRef]
- Madhav, M.R.; David, S.E.M.; Kumar, R.S.S.; Swathy, J.S.; Bhuvaneshwari, M.; Mukherjee, A.; Chandrasekaran, N. Toxicity and Accumulation of Copper Oxide (CuO) Nanoparticles in Different Life Stages of Artemia Salina. Environ. Toxicol. Pharmacol. 2017, 52, 227–238. [Google Scholar] [CrossRef]
- Ekonomou, G.; Lolas, A.; Castritsi-Catharios, J.; Neofitou, C.; Zouganelis, G.D.; Tsiropoulos, N.; Exadactylos, A. Mortality and Effect on Growth of Artemia Franciscana Exposed to Two Common Organic Pollutants. Water 2019, 11, 1614. [Google Scholar] [CrossRef]
- Najla, M.; AbuShaala, N.; Syaizwan, Z.; Ismail, A.; Noor, M.; Azmai, A.; Mohamat-Yusuff, F. Selected Morphological Changes in Nauplii of Brine Shrimp (Artemia salina) after Tributyltin Chloride (TBTCL) Exposure. World Appl. Sci. J. 2015, 33, 1334–1340. [Google Scholar] [CrossRef]
- Baruah, K.; Huy, T.T.; Norouzitallab, P.; Niu, Y.; Gupta, S.K.; De Schryver, P.; Bossier, P. Probing the Protective Mechanism of Poly-β-Hydroxybutyrate against Vibriosis by Using Gnotobiotic Artemia Franciscana and Vibrio Campbellii as Host-Pathogen Model. Sci. Rep. 2015, 5, 9427. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, R.M.; Miguel, T.B.A.R.; de Castro Miguel, E.; Campelo, P.H.; Fernandes, F.A.N.; Rodrigues, S. Protective Effect of Ultrasound-Processed Amazonian Sapota-do-Solimões (Quararibea cordata) Juice on Artemia salina Nauplii. Processes 2022, 10, 1880. https://doi.org/10.3390/pr10091880
da Silva RM, Miguel TBAR, de Castro Miguel E, Campelo PH, Fernandes FAN, Rodrigues S. Protective Effect of Ultrasound-Processed Amazonian Sapota-do-Solimões (Quararibea cordata) Juice on Artemia salina Nauplii. Processes. 2022; 10(9):1880. https://doi.org/10.3390/pr10091880
Chicago/Turabian Styleda Silva, Rhonyele Maciel, Thaiz Batista Azevedo Rangel Miguel, Emilio de Castro Miguel, Pedro Henrique Campelo, Fabiano A. N. Fernandes, and Sueli Rodrigues. 2022. "Protective Effect of Ultrasound-Processed Amazonian Sapota-do-Solimões (Quararibea cordata) Juice on Artemia salina Nauplii" Processes 10, no. 9: 1880. https://doi.org/10.3390/pr10091880
APA Styleda Silva, R. M., Miguel, T. B. A. R., de Castro Miguel, E., Campelo, P. H., Fernandes, F. A. N., & Rodrigues, S. (2022). Protective Effect of Ultrasound-Processed Amazonian Sapota-do-Solimões (Quararibea cordata) Juice on Artemia salina Nauplii. Processes, 10(9), 1880. https://doi.org/10.3390/pr10091880