
Enhanced Enzymatic Production of Antioxidant Peptides from Carya cathayensis Cake Using an Enzymatic Membrane-Coupled Reactor

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Pecan Cake Protein
2.3. Batch Enzymatic Hydrolysis Method of Enzymatic Membrane Reactor
2.4. Optimization of Batch Enzymatic Hydrolysis Conditions in an Enzymatic Membrane Reactor
2.5. Constant Feeding Enzymatic Membrane Coupling Test
2.6. Yield of Peptide
2.7. Peptide Production
2.8. Unit Enzyme Peptide Production
2.9. Membrane Flux
2.10. Antioxidant Activity
2.10.1. 1,1-Diphenyl-2-picrylhydrazyl (DPPH) Free Radical Scavenging Activity
2.10.2. 2,2-Azinobis (3-ethylbenzthiazoline-6-sulfonate) (ABTS) Free Radical Scavenging Activity
2.10.3. Ferrous Ion Chelating Ability
2.11. Analysis of Amino Acid Composition via High-Performance Liquid Chromatography
2.12. Statistical Analysis
3. Results and Discussion
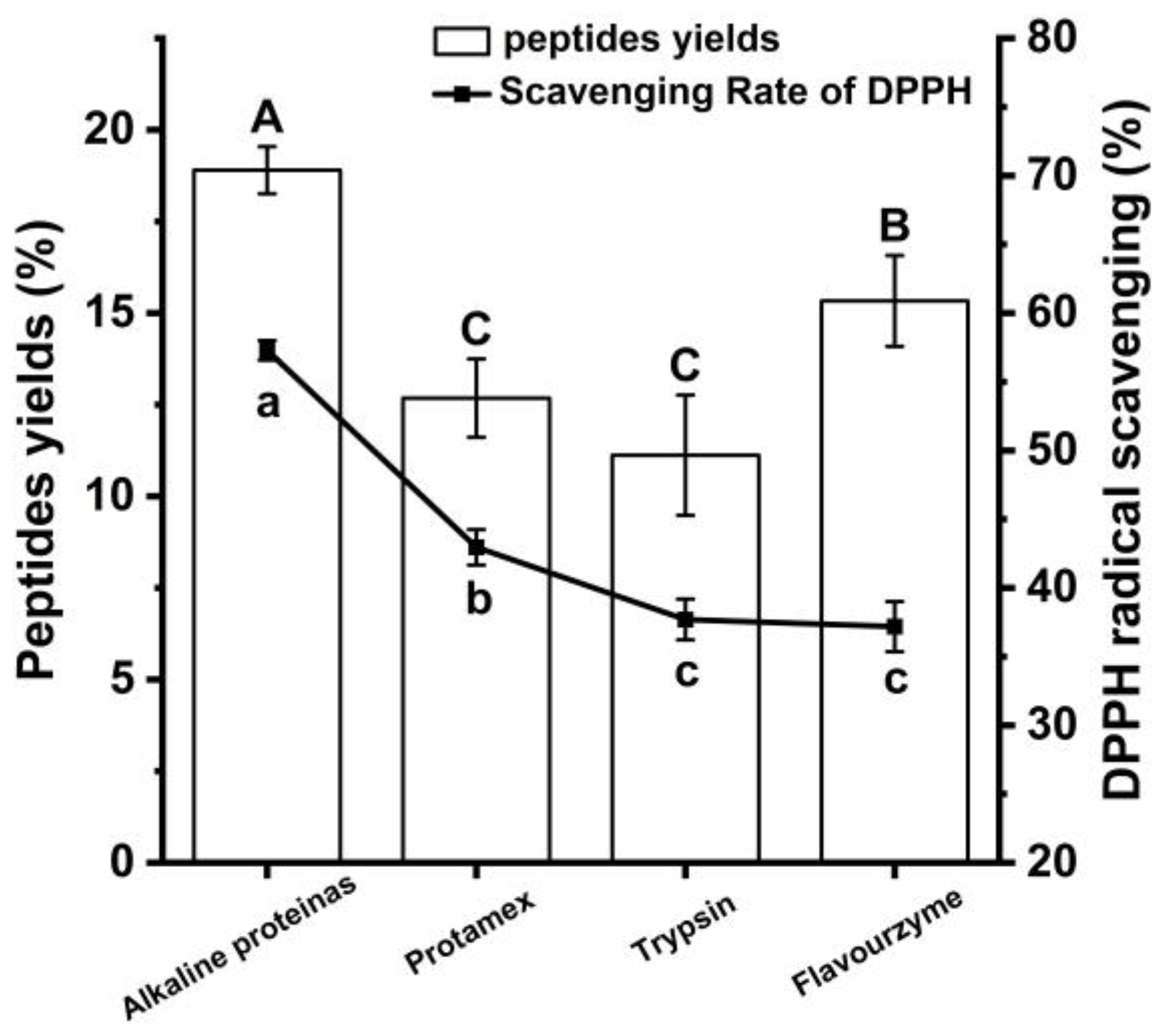
3.1. Establishment of Enzymatic Hydrolysis Method for Chinese Pecan Antioxidant Peptides Enzyme Selection
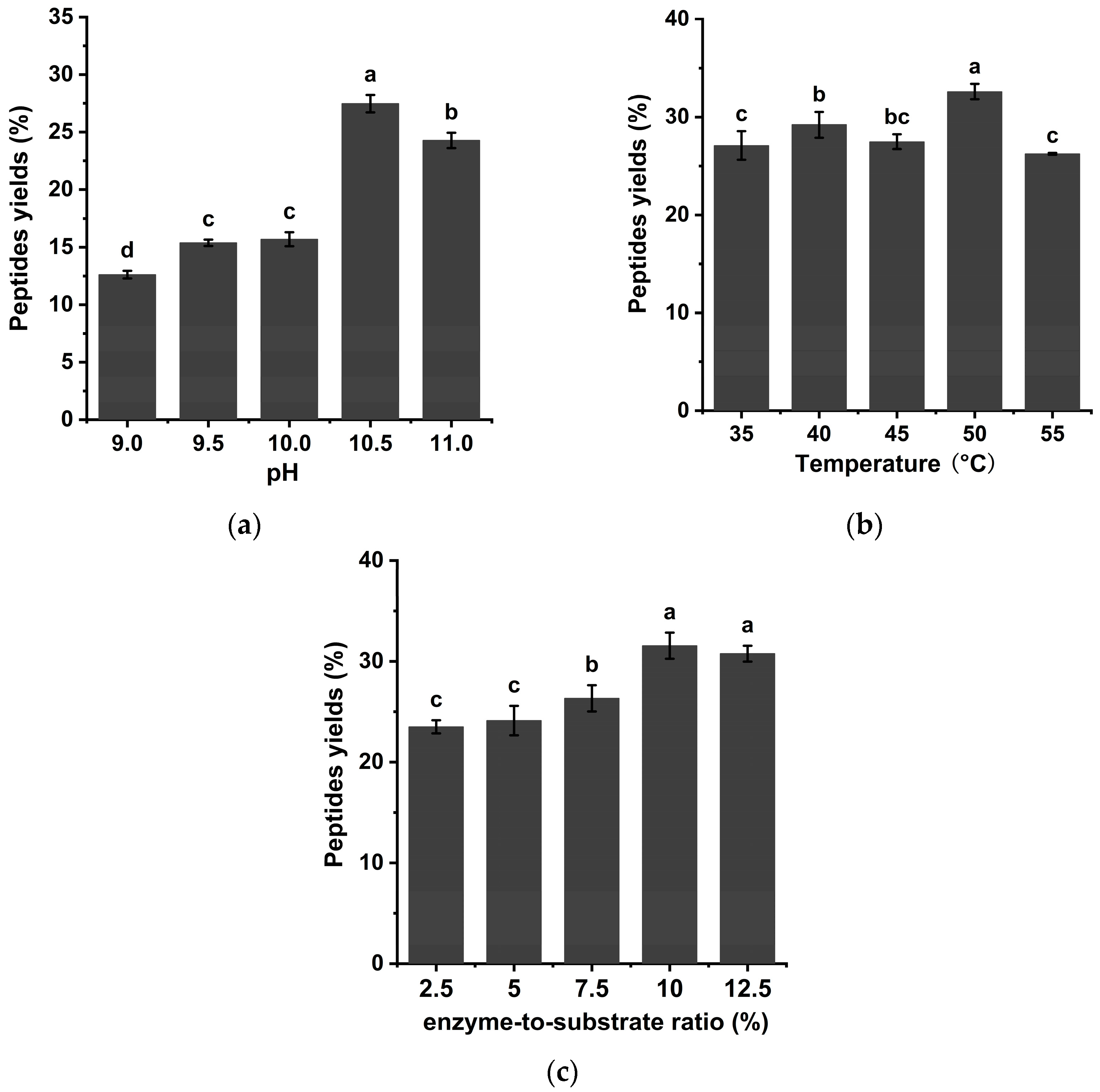
3.2. Impact of Enzymatic Hydrolysis pH on the Yield of Chinese Pecan Cake Peptides in a Batch Enzymatic Process
3.3. Impact of Enzymatic Hydrolysis Temperature on the Yield of Chinese Pecan Cake Peptides in Batch Enzymatic Process
3.4. Impact of Enzyme-to-Substrate Ratio on the Yield of Chinese Pecan Cake Peptides in Batch Enzymatic Process
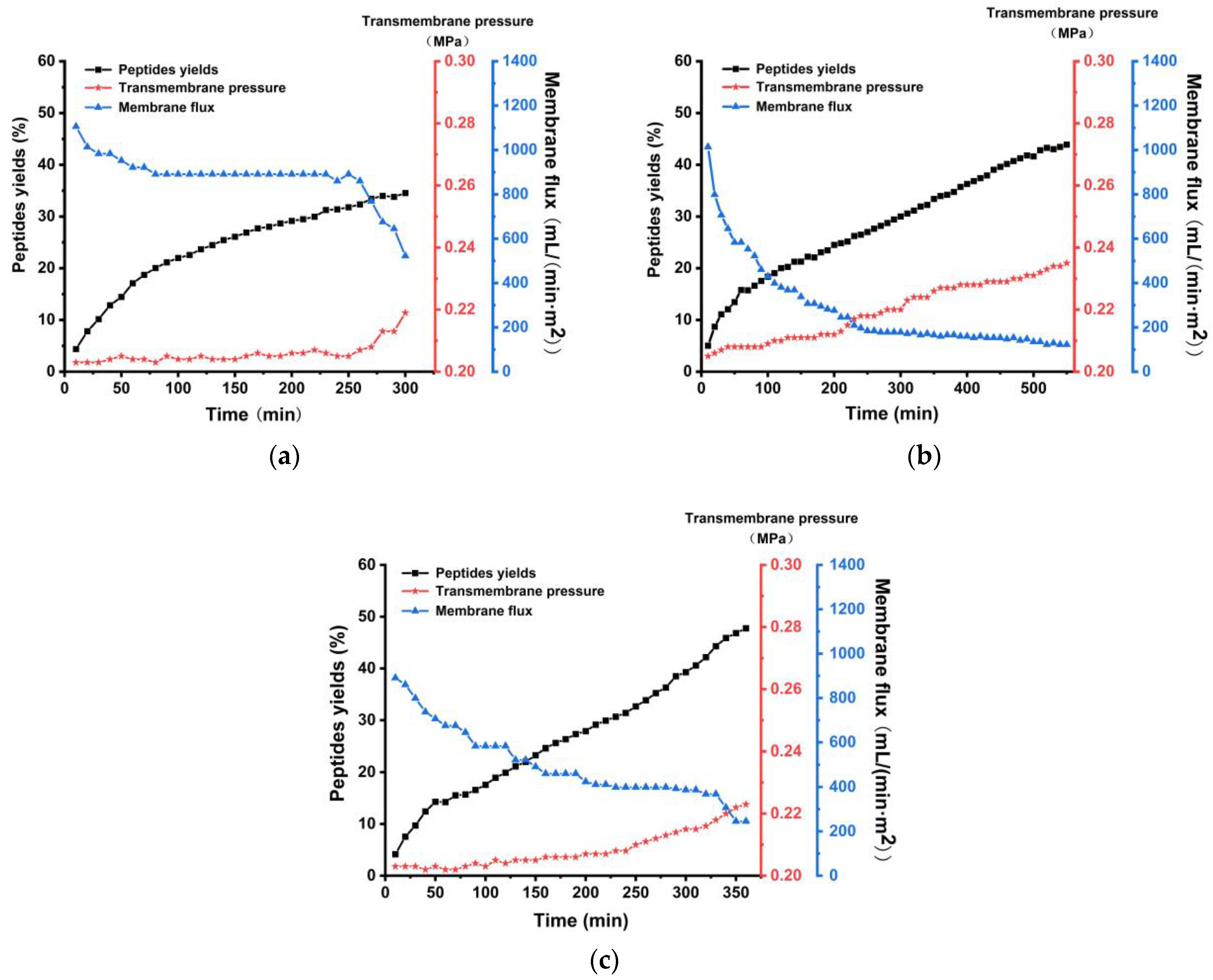
3.5. Comparative Analysis of Enzyme Membrane Coupling Production Processes for Chinese Pecan Cake Peptides Employing Different Supplementary Feeding Modes
3.5.1. Operational Performance Comparison in Different Supplementary Feeding Modes for Enzyme Membrane Coupling Process
3.5.2. Comparative Analysis of Production Efficiency in Enzyme Membrane Coupling Process with Various Supplementary Feeding Modes
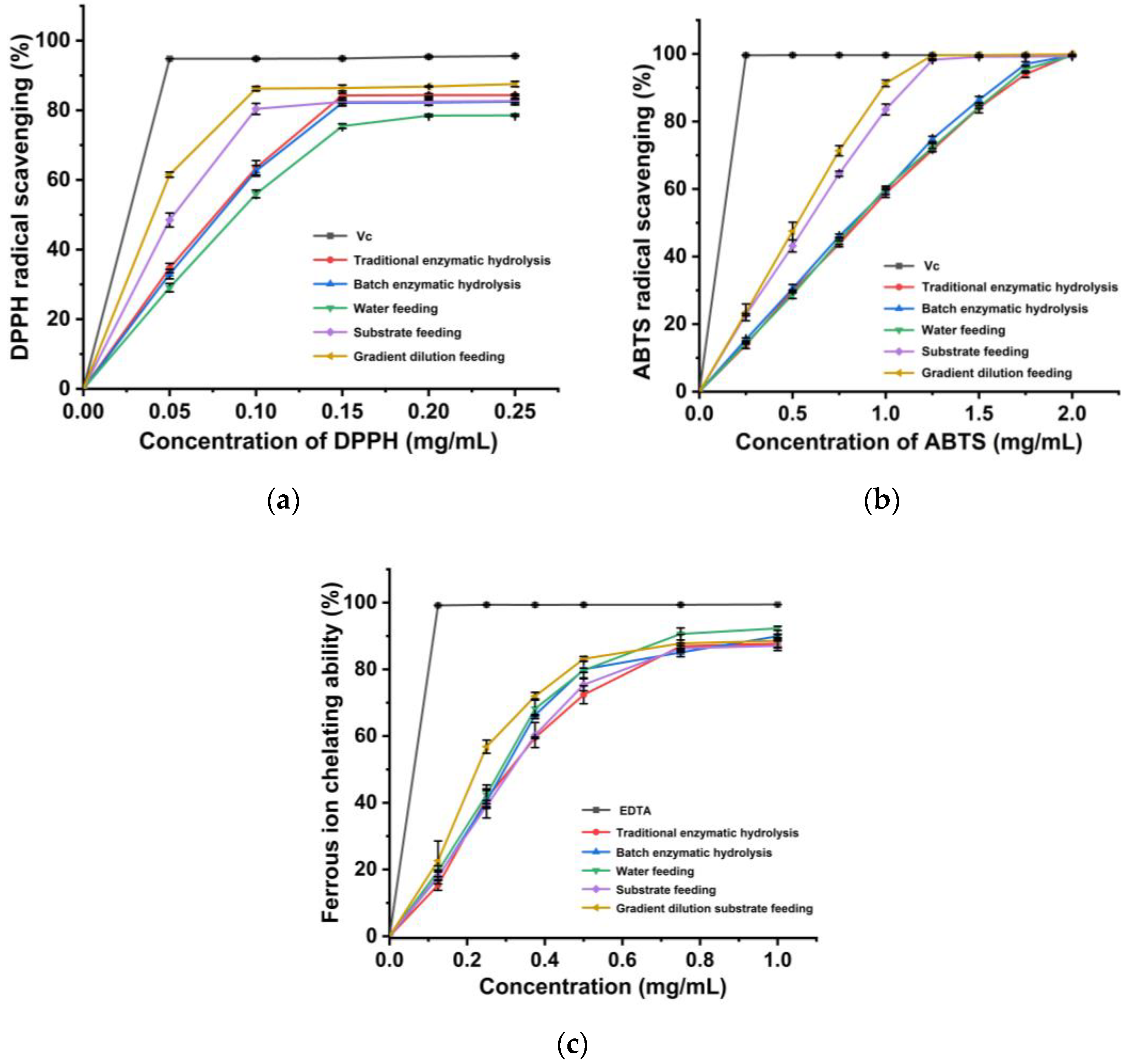
3.6. Comparison of Antioxidant Activity
3.7. Comparison of Amino Acid Composition
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mora, L.; Toldrá, F. Advanced enzymatic hydrolysis of food proteins for the production of bioactive peptides. Curr. Opin. Food Sci. 2023, 49, 100973. [Google Scholar] [CrossRef]
- Huang, S.; Gong, Y.; Li, Y.; Ruan, S.; Ma, H. Preparation of ACE-inhibitory peptides from milk protein in continuous enzyme membrane reactor with gradient dilution feeding substrate. Process Biochem. 2020, 92, 130–137. [Google Scholar] [CrossRef]
- Wang, W.Q.; Han, X.; Yi, H.; Zhang, L.W. The ultrafiltration efficiency and mechanism of transglutaminase enzymatic membrane reactor (EMR) for protein recovery from cheese whey. Int. Dairy J. 2018, 80, 52–61. [Google Scholar] [CrossRef]
- Rodriguez-Nogales, J.M.; Ortega, N.; Perez-Mateos, M.; Busto, M.D. Pectin hydrolysis in a free enzyme membrane reactor: An approach to the wine and juice clarification. Food Chem. 2008, 107, 112–119. [Google Scholar] [CrossRef]
- Heyse, A.; Plikat, C.; Ansorge-Schumacher, M.; Drews, A. Continuous two-phase biocatalysis using water-in-oil Pickering emulsions in a membrane reactor: Evaluation of different nanoparticles. Catal. Today 2019, 331, 60–67. [Google Scholar] [CrossRef]
- Sun, L.; Xing, Z.; Liao, G.; Hou, X.; He, R.; Ma, H. Study on the Preparation of Antioxidant Peptides from Sunflower Seeds Meal by Enzyme Membrane Coupling. Food Ind. 2018, 39, 221–225. [Google Scholar]
- Li, Y.; Shao, Y.; Zhou, W.; Liu, L.; Tang, S.; Liu, T. Preparation of Collagen Polypeptide with Homogeneous Molecular Weight by Enzyme Membrane Coupled Method. Chem. Bioeng. 2017, 34, 25–28+32. [Google Scholar]
- Guo, Y.; Wang, K.; Wu, B.; Wu, P.; Duan, Y.; Ma, H. Production of ACE inhibitory peptides from corn germ meal by an enzymatic membrane reactor with a novel gradient diafiltration feeding working-mode and in vivo evaluation of antihypertensive effect. J. Funct. Foods 2020, 64, 103584. [Google Scholar] [CrossRef]
- O’Halloran, J.; O’Sullivan, M.; Casey, E. Production of Whey-Derived DPP-IV Inhibitory Peptides Using an Enzymatic Membrane Reactor. Food Bioprocess Technol. 2019, 12, 799–808. [Google Scholar] [CrossRef]
- Yang, J.; Zhou, F.; Xiong, L.; Mao, S.; Hu, Y.; Lu, B. Comparison of phenolic compounds, tocopherols, phytosterols and antioxidant potential in Zhejiang pecan [Carya cathayensis] at different stir-frying steps. LWT-Food Sci. Technol. 2015, 62, 541–548. [Google Scholar] [CrossRef]
- Maciel, L.G.; Ribeiro, F.L.; Teixeira, G.L.; Molognoni, L.; Block, J.M. The potential of the pecan nut cake as an ingredient for the food industry. Food Res. Int. 2019, 127, 108718. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; He, Z.; Wang, Q.; Wu, F.; Liu, X. Response surface optimization of enzymatic hydrolysis of peptides of chinese pecan (Carya cathayensis) and analysis of their antioxidant capacities and structures. Int. J. Pept. Res. Ther. 2021, 27, 1239–1251. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, S.; Wang, Q.; Yuan, Q.; Li, Y.; Reboredo-Rodríguez, P.; Varela-López, A.; He, Z.; Wu, F.; Hu, H. Oxidative stress amelioration of novel peptides extracted from enzymatic hydrolysates of Chinese pecan cake. Int. J. Mol. Sci. 2022, 23, 12086. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Wang, Q.; Teng, J.; Wu, F.; He, Z. Preparation process optimization and evaluation of bioactive peptides from Carya cathayensis Sarg meal. Curr. Res. Food Sci. 2023, 6, 100408. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Hao, L.; Chen, F.; Yang, C. Study on Extraction of Peanut Protein and Oil Bodies by Aqueous Enzymatic Extraction and Characterization of Protein. J. Chem. 2020, 2020, 5148967. [Google Scholar] [CrossRef]
- Moaveni, S.; Salami, M.; Khodadadi, M.; McDougall, M.; Emam-Djomeh, Z. Investigation of S. limacinum microalgae digestibility and production of antioxidant bioactive peptides. LWT 2022, 154, 112468. [Google Scholar] [CrossRef]
- Hu, X.; Pan, C.; Cai, M.; Li, L.; Yang, X.; Xiang, H.; Chen, S. Novel antioxidant peptides from Grateloupia livida hydrolysates: Purification and identification. Foods 2022, 11, 1498. [Google Scholar] [CrossRef] [PubMed]
- Abeynayake, R.; Zhang, S.; Yang, W.; Chen, L. Development of antioxidant peptides from brewers’ spent grain proteins. LWT 2022, 158, 113162. [Google Scholar] [CrossRef]
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.-H.; Tavano, O.; Berenguer-Murcia, A.; Fernandez-Lafuente, R. Use of Alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196. [Google Scholar] [CrossRef]
- Jiang, H.; Pan, J.; Hu, X.; Zhu, M.; Gong, D.; Zhang, G. A combination of alkaline pH-shifting/acidic pH and thermal treatments improves the solubility and emulsification properties of wheat glutenin. Food Chem. 2022, 393, 133358. [Google Scholar] [CrossRef]
- Qu, W.; Ma, H.; Li, W.; Pan, Z.; Owusu, J.; Venkitasamy, C. Performance of coupled enzymatic hydrolysis and membrane separation bioreactor for antihypertensive peptides production from Porphyra yezoensis protein. Process Biochem. 2015, 50, 245–252. [Google Scholar] [CrossRef]
- Gao, R.; Shen, Y.; Shu, W.; Bai, F.; Yuan, L. Optimization of Enzymatic Conditions of Sturgeon Muscles and Their Anti-Inflammatory Potential. J. Food Qual. 2020, 2020, 9698134. [Google Scholar] [CrossRef]
- Zhang, J.; Feng, C.; Xu, H.; Tan, X.; Hagedoorn, P.-L.; Ding, S. Enhanced hypericin extraction from Hypericum perforatum L. by coupling microwave with enzyme-assisted strategy. Ind. Crops Prod. 2019, 137, 231–238. [Google Scholar] [CrossRef]
- Jiao, S.; Li, Y.; Wang, Z.; Sun-Waterhouse, D.; Waterhouse, G.I.; Liu, C.; Wang, X. Optimization of enzyme-assisted extraction of bioactive-rich juice from Chaenomeles sinensis (Thouin) Koehne by response surface methodology. J. Food Process. Preserv. 2020, 44, e14638. [Google Scholar] [CrossRef]
- Liu, M.C.; Yang, S.J.; Hong, D.; Yang, J.P.; Wang, C.J. A simple and convenient method for the preparation of antioxidant peptides from walnut (Juglans regia L.) protein hydrolysates. Chem. Cent. J. 2016, 10, 39. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Feng, Y.; Duan, Y.; Ma, H.; Zhang, H. Purification and identification of novel antioxidant peptides from watermelon seed protein hydrolysates and their cytoprotective effects on H2O2-induced oxidative stress. Food Chem. 2020, 327, 127059. [Google Scholar] [CrossRef]
- Sun, C.; Wu, W.; Yin, Z.; Fan, L.; Ma, Y.; Lai, F.; Wu, H. Effects of simulated gastrointestinal digestion on the physicochemical properties, erythrocyte haemolysis inhibitory ability and chemical antioxidant activity of mulberry leaf protein and its hydrolysates. Int. J. Food Sci. Technol. 2017, 53, 282–295. [Google Scholar] [CrossRef]
- Xie, Z.; Huang, J.; Xu, X.; Jin, Z. Antioxidant activity of peptides isolated from alfalfa leaf protein hydrolysate. Food Chem. 2008, 111, 370–376. [Google Scholar] [CrossRef]
- Ketnawa, S.; Wickramathilaka, M.; Liceaga, A.M. Changes on antioxidant activity of microwave-treated protein hydrolysates after simulated gastrointestinal digestion: Purification and identification. Food Chem. 2018, 254, 36–46. [Google Scholar] [CrossRef]
- Zou, T.-B.; He, T.-P.; Li, H.-B.; Tang, H.-W.; Xia, E.-Q. The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef]
- Phongthai, S.; D’Amico, S.; Schoenlechner, R.; Homthawornchoo, W.; Rawdkuen, S. Fractionation and antioxidant properties of rice bran protein hydrolysates stimulated by in vitro gastrointestinal digestion. Food Chem. 2018, 240, 156–164. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Traditional Enzymatic Hydrolysis | Enzyme Membrane Coupling | |||
|---|---|---|---|---|---|
| Batch Enzymatic Hydrolysis | Water Feeding | Substrate Feeding | Gradient Dilution Feeding | ||
| Peptides yield (%) | 33.97 ± 0.60 c | 31.12 ± 1.70 d | 34.61 ± 0.06 c | 43.83 ± 0.98 b | 48.33 ± 1.19 a |
| Peptides production (%) | 21.27 ± 0.95 c | 20.38 ± 0.63 c | 22.12 ± 1.90 c | 25.20 ± 0.96 b | 32.63 ± 0.31 a |
| Unit enzyme peptides production (g/g) | 2.13 ± 0.10 c | 2.04 ± 0.06 c | 2.21 ± 0.19 c | 6.30 ± 0.24 b | 7.14 ± 0.07 a |
| Amino Acid | Proportion (%) | ||||
|---|---|---|---|---|---|
| Traditional Enzymatic Hydrolysis | Batch Enzymatic Hydrolysis | Water Feeding | Substrate Feeding | Gradient Dilution Feeding | |
| Asp | 7.45 ± 0.51 b | 7.37 ± 0.66 b | 8.64 ± 0.30 a | 8.00 ± 0.15 ab | 7.81 ± 0.40 b |
| Glu | 20.08 ± 0.20 a | 20.47 ± 0.56 a | 20.63 ± 0.39 a | 20.15 ± 0.13 a | 20.7 ± 0.37 a |
| Ser | 5.29 ± 0.08 a | 5.51 ± 0.02 a | 5.11 ± 0.17 ab | 4.91 ± 0.01 bc | 4.66 ± 0.03 c |
| Gly | 4.66 ± 0.06 b | 4.51 ± 0.02 c | 4.66 ± 0.08 b | 4.89 ± 0.05 a | 4.29 ± 0.13 d |
| His | 1.97 ± 0.03 a | 1.82 ± 0.01 b | 1.72 ± 0.06 c | 1.74 ± 0.08 c | 1.94 ± 0.01 a |
| The | 2.89 ± 0.01 a | 3.06 ± 0.01 a | 2.61 ± 0.44 a | 2.96 ± 0.05 a | 2.89 ± 0.02 a |
| Ala | 4.77 ± 0.15 a | 4.51 ± 0.01 b | 4.28 ± 0.07 c | 4.16 ± 0.01 d | 4.06 ± 0.05 d |
| Arg | 11.54 ± 0.22 b | 11.43 ± 0.03 ab | 11.46 ± 0.01 b | 11.52 ± 0.03 b | 11.98 ± 0.02 a |
| Pro | 8.35 ± 0.07 c | 8.72 ± 0.06 b | 8.99 ± 0.11 a | 8.68 ± 0.01 b | 9.06 ± 0.01 a |
| Tyr | 2.73 ± 0.08 c | 2.75 ± 0.02 bc | 2.79 ± 0.01 b | 2.85 ± 0.01 a | 2.72 ± 0.01 c |
| Val | 5.99 ± 0.01 a | 5.76 ± 0.01 b | 5.92 ± 0.05 ab | 6.06 ± 0.01 a | 6.00 ± 0.02 a |
| Met | 1.59 ± 0.01 bc | 1.84 ± 0.02 a | 1.51 ± 0.02 cd | 1.69 ± 0.04 ab | 1.41 ± 0.01 d |
| Cys | 0.79 ± 0.01 b | 0.99 ± 0.01 a | 0.77 ± 0.01 d | 0.79 ± 0.01 b | 0.78 ± 0.01 c |
| Ile | 4.39 ± 0.03 d | 4.49 ± 0.03 ab | 4.50 ± 0.04 a | 4.45 ± 0.01 bc | 4.41 ± 0.01 cd |
| Leu | 8.56 ± 0.13 a | 7.93 ± 0.06 b | 7.56 ± 0.04 d | 7.74 ± 0.01 c | 7.79 ± 0.04 c |
| Phe | 5.37 ± 0.06 ab | 5.29 ± 0.04 b | 5.28 ± 0.02 ab | 5.32 ± 0.01 ab | 5.46 ± 0.02 a |
| Lys | 3.59 ± 0.04 b | 3.55 ± 0.03 b | 3.57 ± 0.02 b | 4.10 ± 0.01 a | 4.04 ± 0.05 a |
| Aci AAs | 27.53 ± 0.31 e | 27.84 ± 0.33 d | 29.27 ± 0.23 a | 28.15 ± 0.02 c | 28.51 ± 0.03 b |
| BAAs | 17.10 ± 0.16 c | 16.8 ± 0.02 d | 16.76 ± 0.08 d | 17.36 ± 0.05 b | 17.96 ± 0.03 a |
| EAAs | 34.34 ± 0.01 a | 33.74 ± 0.15 a | 32.66 ± 0.20 a | 34.06 ± 0.04 a | 33.94 ± 0.11 a |
| HAAs | 40.17 ± 0.32 a | 39.45 ± 0.14 b | 39.32 ± 0.06 b | 39.25 ± 0.01 b | 39.50 ± 0.14 b |
| Aro AAs | 8.10 ± 0.07 b | 8.04 ± 0.05 c | 8.07 ± 0.03 bc | 8.17 ± 0.01 a | 8.18 ± 0.03 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Chang, Y.; Yuan, Q.; Wu, F.; Hu, H.; Zhang, J.; He, Z. Enhanced Enzymatic Production of Antioxidant Peptides from Carya cathayensis Cake Using an Enzymatic Membrane-Coupled Reactor. Processes 2024, 12, 2. https://doi.org/10.3390/pr12010002
Wang Q, Chang Y, Yuan Q, Wu F, Hu H, Zhang J, He Z. Enhanced Enzymatic Production of Antioxidant Peptides from Carya cathayensis Cake Using an Enzymatic Membrane-Coupled Reactor. Processes. 2024; 12(1):2. https://doi.org/10.3390/pr12010002
Chicago/Turabian StyleWang, Qingqing, Yinzi Chang, Qinjie Yuan, Fenghua Wu, Hao Hu, Jiaojiao Zhang, and Zhiping He. 2024. "Enhanced Enzymatic Production of Antioxidant Peptides from Carya cathayensis Cake Using an Enzymatic Membrane-Coupled Reactor" Processes 12, no. 1: 2. https://doi.org/10.3390/pr12010002
APA StyleWang, Q., Chang, Y., Yuan, Q., Wu, F., Hu, H., Zhang, J., & He, Z. (2024). Enhanced Enzymatic Production of Antioxidant Peptides from Carya cathayensis Cake Using an Enzymatic Membrane-Coupled Reactor. Processes, 12(1), 2. https://doi.org/10.3390/pr12010002