
Photorespiration and Rate Synchronization in a Phototroph-Heterotroph Microbial Consortium




Abstract
:1. Introduction
2. Materials and Methods
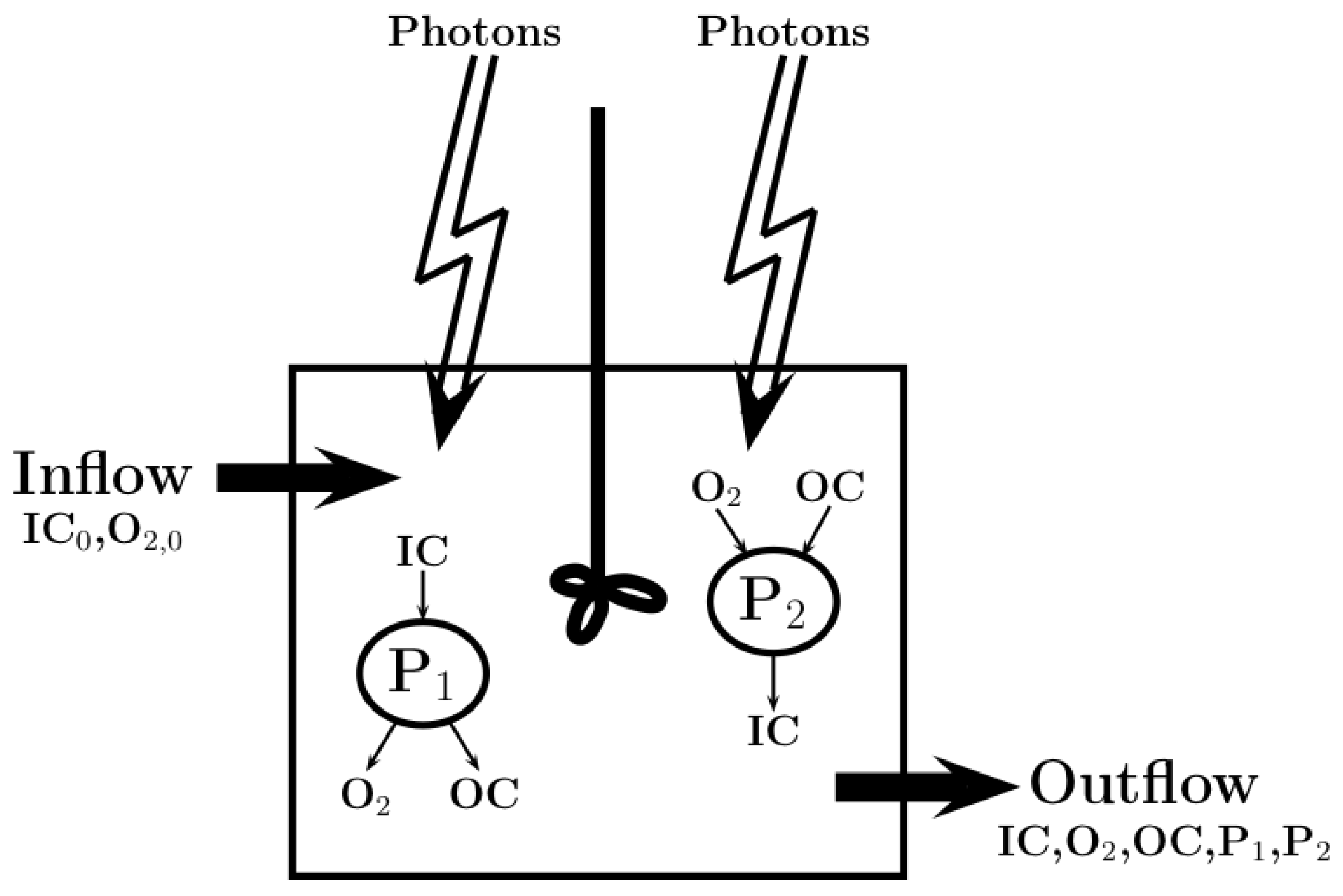
2.1. Model Description
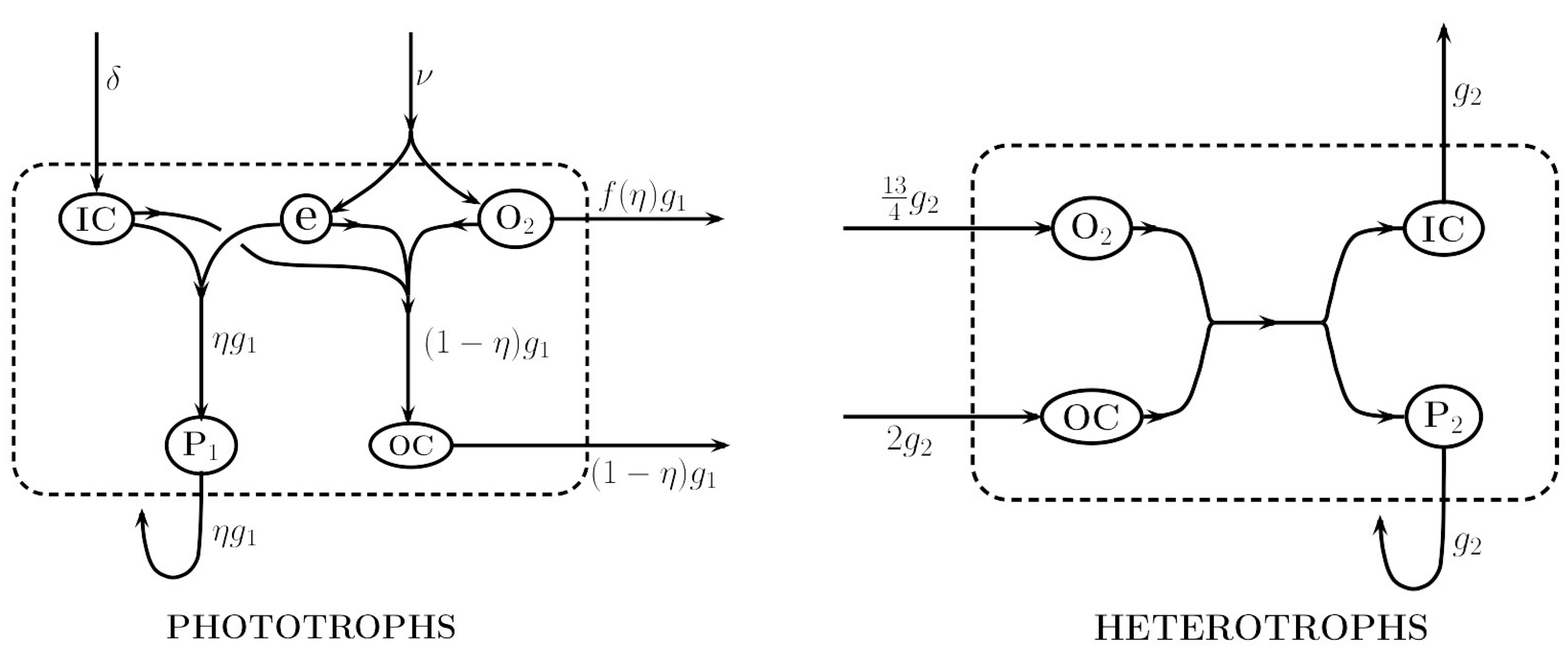
2.2. Metabolic Pathways
Relation to Pathway Analysis
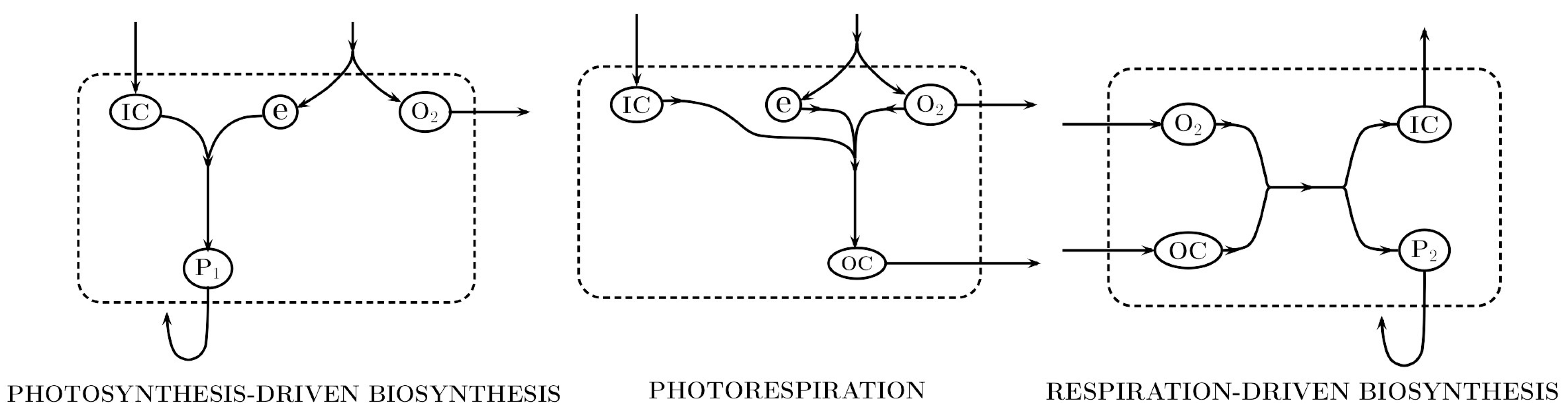
2.3. Photosynthesis
2.3.1. Carbon Fixation
2.3.2. Photoinhibition and Oxidative Stress
2.3.3. Photorespiration
2.3.4. Fixation Stability
2.4. Heterotrophic Biosynthesis
2.5. Equations
3. Results
3.1. Single Species Chemostat Community
3.1.1. Steady States
3.1.2. Stability of Steady States
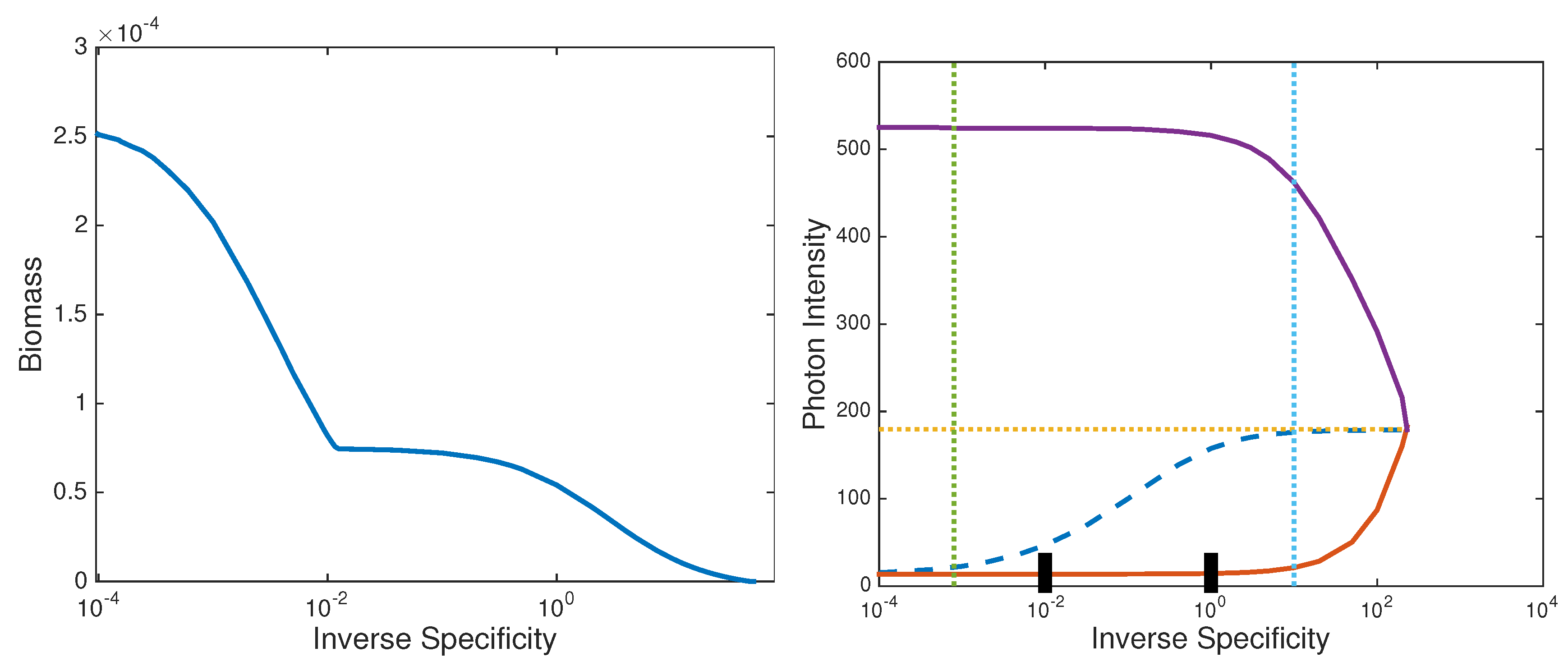
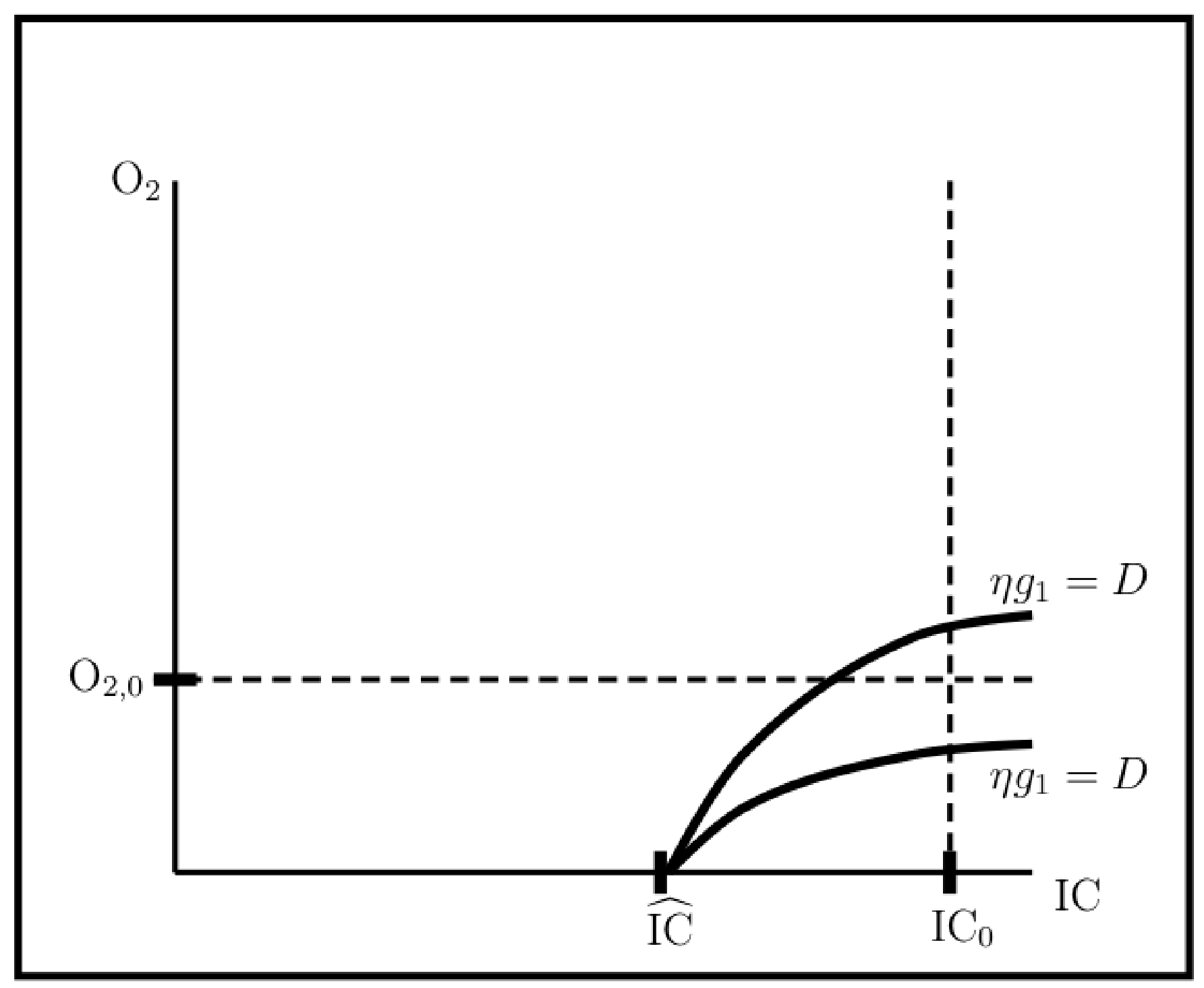
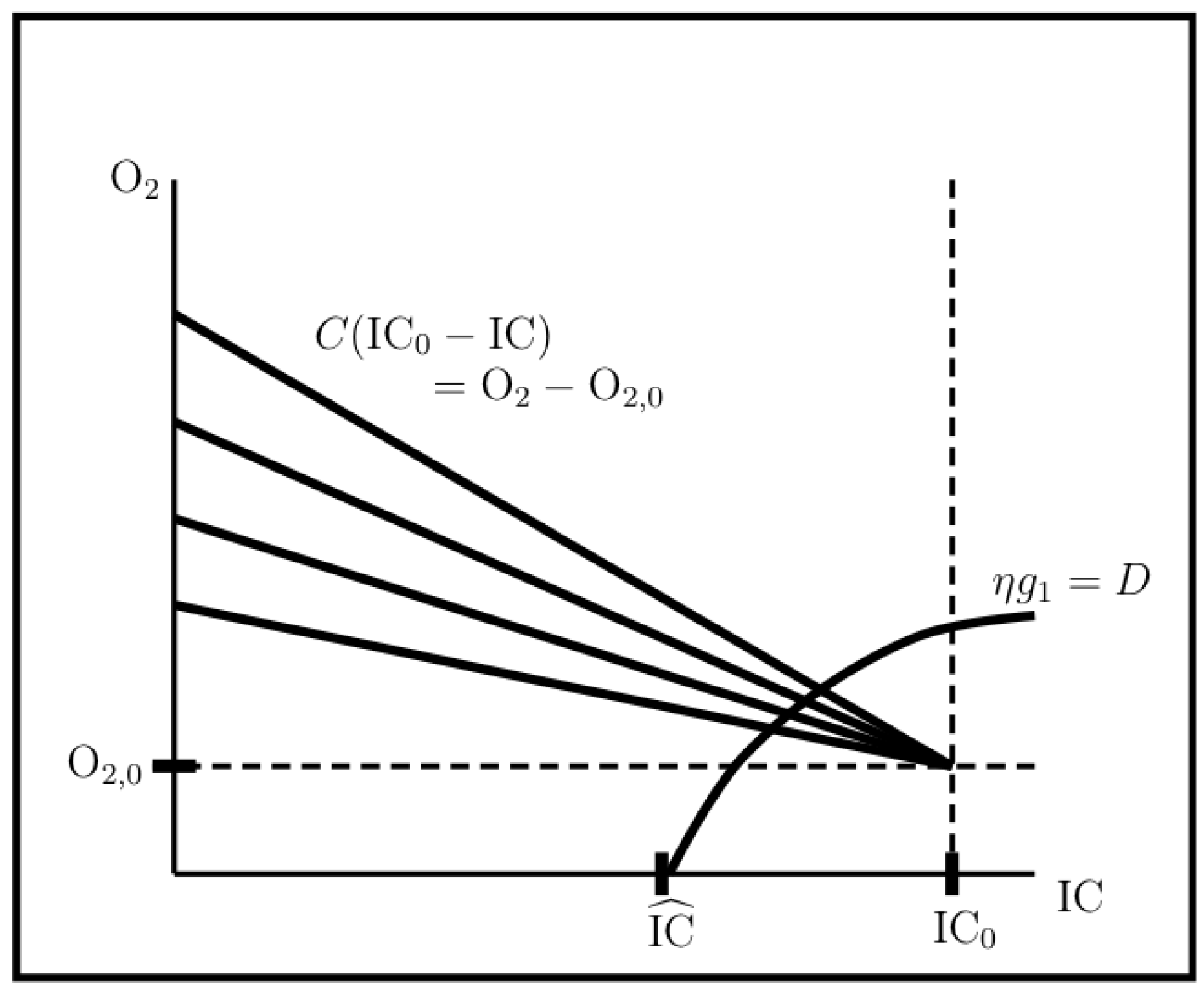
3.1.3. Viability and Light-Limited Ranges
- Setting , i.e., turning photorespiration off entirely, results in only a single light intensity with a viable, non carbon-limited steady state population. However, at ambient O and IC concentration levels, competition for RuBisCO binding is insignificant for inverse specificities . Hence, from the point of view of population viability at least, there is no penalty for allowing RuBisCO oxygenase activity over this inverse specificity range.
- On the other hand, inverse specificities such that result in significantly enlarged light-limited intensity range, so that large enough inverse specificities may have some advantage.
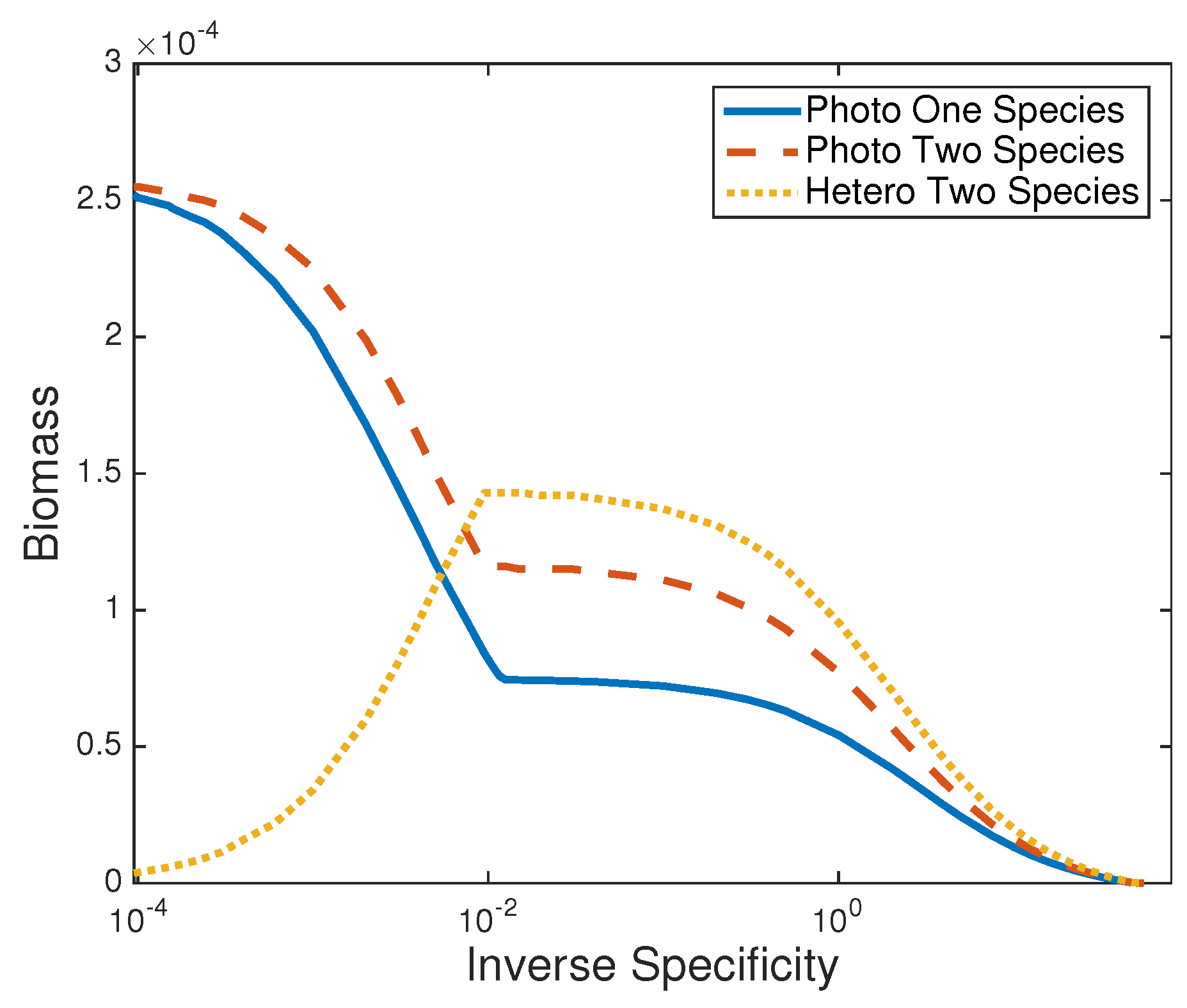
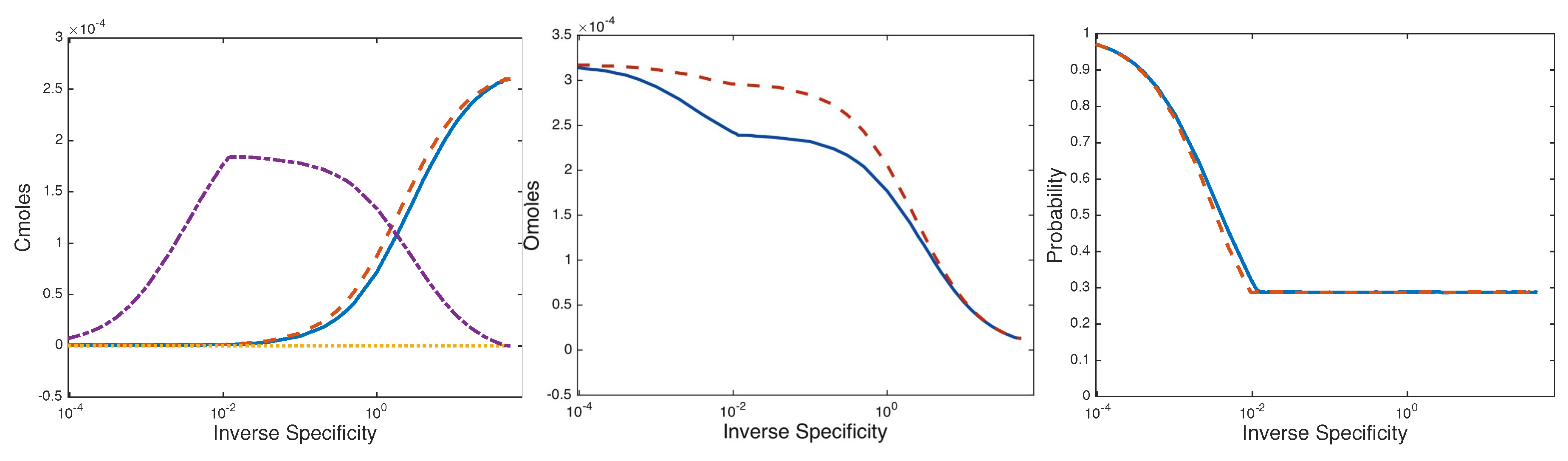
3.2. Two Species Community
3.2.1. Steady States and Stability
3.2.2. Two Species Consortium Steady State
4. Discussion
- decrease in population biomass,
- increase in population light tolerance,
- and decrease in oxygen concentration.
- biomass increase,
- reduction in dissolved organic carbon,
- and oxygen concentration increase.
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A. Degree of Reduction
Appendix B. Parameter Estimation
| Symbol | Name | Unit | Value | Reference |
| Inverse specificity | Cmol·Omol | 0.01–1 | [11,26,27] | |
| Excess elec. rate capacity | s | – | – | |
| Maximal transport rate | s | Measured | ||
| Maximal transport rate | s | [32] | ||
| Half saturation | Cmol·L | [29] | ||
| Half saturation | Omol L | [35,36] | ||
| Half saturation | Cmol·L | [37] | ||
| ν | Photon flux | E·m·s | 0–2000 | [38] |
| α | Efficiency | - | 1 | – |
| IC | Inflow IC concentration | Cmol·L | – | |
| OC | Inflow OC concentration | Cmol·L | 0 | – |
| O | Inflow O concentration | Omol·L | – | |
| D | Chemostat turnover rate | s | various | – |
| Yield | Cmol·cell | z | ||
| Yield | Cmol·cell | z | ||
| Yield | Omol·ph | |||
| Yield | Omol·cell | |||
| Yield | Omol·cell | |||
| Yield | Omol·electron | |||
| Yield | Electron·ph |
Appendix C. Existence and Uniqueness of Single Species Viable State Solutions
- η and are smooth.
- Monotonicity in O: for fixed value of IC, is monotonically non-increasing in O with values decreasing from to 0 as O varies from 0 to ∞, and is monotonically decreasing in O with values decreasing from 1 to 0 as O varies from 0 to ∞. Roughly speaking, increasing oxygen concentration if anything inhibits photosynthesis and always shifts photosynthetic product from biosynthesis to photorespiration.
- Monotonicity in IC: for fixed value of O, is monotonically non-decreasing in IC with values increasing from 0 to as IC varies from 0 to IC, and is monotonically increasing in IC with values increasing from 0 to as IC varies from 0 to IC. (In fact, η should tend to 1 as ). Roughly speaking, increasing inorganic carbon concentration if anything promotes photosynthesis and always shifts photosynthetic product from photorespiration to biosynthesis.
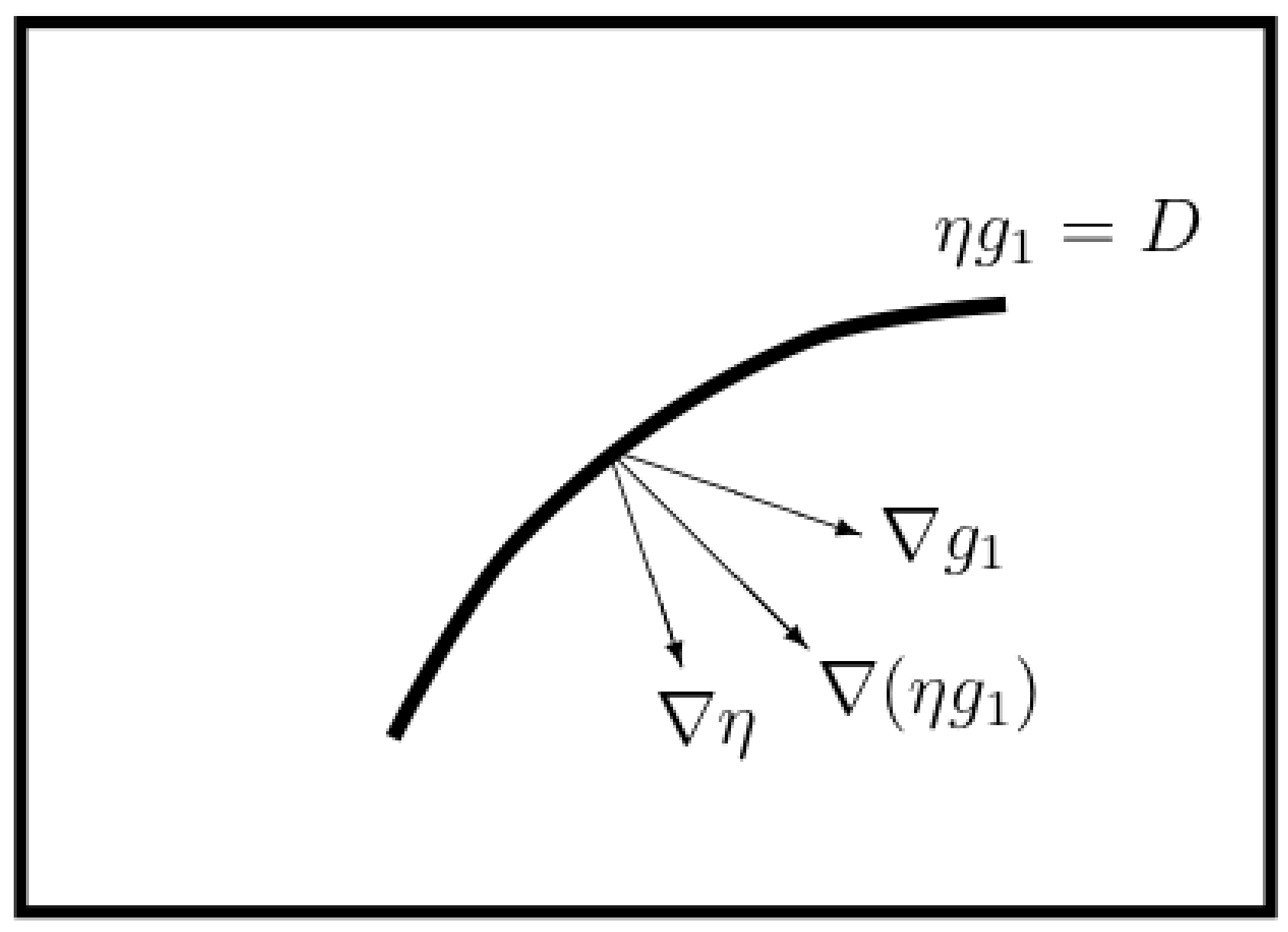
- Fixation stability: we assume that condition (17), namely , holds.
- Note as well that the function is necessarily a linear function with parameterization determined by stoichiometry and degree of reduction values. In fact, for the particular choices we use, , however we here need only suppose that for some .



Appendix D. Linearization and Stability
Appendix E. Optimization in the Single Species Chemostat With Respect to Affinity
Appendix F. Invasion Eigenvector
References
- Villa, F.; Pitts, B.; Lauchnor, E.G.; Cappitelli, F.; Stewart, P.S. Development of a laboratory model of a phototroph-heterotroph mixed-species biofilm at the stone/air interface. Front. Microbiol. 2015, 6, 1251. [Google Scholar] [CrossRef] [PubMed]
- Croft, M.T.; Lawrence, A.D.; Raux-Deery, E.; Warren, M.J.; Smith, A.G. Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature 2005, 438, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Palsson, B.Ø. Systems Biology: Simulation of Dynamics Network States; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Klamt, S.; Stelling, J. Two approaches for metabolic pathway analysis? Trends Biotechnol. 2003, 21, 64–69. [Google Scholar] [CrossRef]
- Schilling, C.H.; Edwards, J.S.; Letscher, D.; Palsson, B.O. Combining pathway analysis with flux balance analysis for the comprehensive study of metabolic systems. Biotechnol. Bioeng. 2001, 71, 286–306. [Google Scholar] [CrossRef]
- Schuster, S.; Hilgetag, C. On elementary flux modes in biochemical reaction systems at steady state. J. Biol. Syst. 1994, 2, 165–182. [Google Scholar] [CrossRef]
- Carlson, R.P. Decomposition of complex microbial behaviors into resource-based stresses. Bioinformatics 2009, 25, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Taffs, R.; Aston, J.E.; Brileya, K.; Jay, Z.; Klatt, C.G.; McGlynn, S.; Mallette, N.; Montross, S.; Gerlach, R.; Inskeep, W.P.; et al. In silico approaches to study mass and energy flows in microbial consortia: A syntrophic case study. BMC Syst. Biol. 2009, 3, 114. [Google Scholar] [CrossRef] [PubMed]
- Phalak, P.; Chen, J.; Carlson, R.P.; Henson, M.A. Metabolic modeling of a chronic wound biofilm consortium predicts spatial partitioning of bacterial species. BMC Syst. Biol. 2016, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L.; Waltman, P. The Theory of the Chemostat: Dynamics of Microbial Competition; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Beck, A.E.; Bernstein, H.C.; Carlson, R.P. Stoichiometric network analysis of cyanobacterial acclimation to photosynthesis-associated stresses identifies heterotrophic niches. Processes 2017. submitted. [Google Scholar]
- Knoop, H.; Grundel, M.; Zilliges, Y.; Lehmann, R.; Hoffmann, S.; Lockau, W.; Steuer, R. Flux balance analysis of cyanobacterial metabolism: The metabolic network of Synechocystis sp. PCC 6803. PLoS Comput. Biol. 2013, 9, e1003081. [Google Scholar] [CrossRef] [PubMed]
- Nogales, J.; Gudmundsson, S.; Knight, E.M.; Palsson, B.O.; Thiele, I. Detailing the optimality of photosynthesis in cyanobacteria through systems biology analysis. Proc. Natl. Acad. Sci. USA 2012, 109, 2678–2683. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.T.; Stolyar, S.M.; Pinchuk, G.E.; Hill, E.A.; Kucek, L.A.; Brown, R.N.; Lipton, M.S.; Osterman, A.; Fredrickson, J.K.; Konopka, A.E.; et al. Genome-scale modeling of light-driven reductant partitioning and carbon fluxes in diazotrophic unicellular cyanobacterium Cyanothece sp. ATCC 51142. PloS Comput. Biol. 2012, 8, e1002460. [Google Scholar] [CrossRef] [PubMed]
- Henry, C.S.; Bernstein, H.C.; Weisenhorn, P.; Taylor, R.C.; Lee, J.Y.; Zucker, J.; Song, H.S. Microbial community metabolic modeling: A community data-driven network reconstruction. J. Cell. Physiol. 2016, 231, 2339–2345. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, O.; Lear, G.; Singhal, N. Metabolic network modeling of microbial interactions in natural and engineered environmental systems. Front. Microbiol. 2016, 7, 673. [Google Scholar] [CrossRef] [PubMed]
- Roels, J.A. Application of macroscopic principles to microbial metabolism. Biotechnol. Bioeng. 1980, 22, 2457–2514. [Google Scholar] [CrossRef]
- White, D. The Physiology and Biochemistry of Prokaryotes, 3rd ed.; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Mangam, N.M.; Brenner, M.P. Systems analysis of the CO2 concentrating mechanism in cyanobacteria. eLIFE 2014, 3, e02043. [Google Scholar]
- Rae, B.D.; Long, B.M.; Whitehead, L.F.; Forster, B.; Badger, M.R.; Price, G.D. Cyanobacterial carboxysomes: Microcompartments that facilitate CO2 fixation. J. Mol. Microbiol. Biotechnol. 2013, 23, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. The water-water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plan Mol. Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef] [PubMed]
- Eilers, P.H.C.; Peeters, J.C.H. A model for the relationship between Light- Intensity and the rate of photosynthesis in Phytoplankton. Ecol. Model. 1988, 42, 199–215. [Google Scholar] [CrossRef]
- Wolf, G.; Picioreanu, C.; van Loosdrecht, M.C.M. Kinetic modeling of phototrophic biofilms—The PHOBIA model. Biotechnol. Bioeng. 2007, 97, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.; Grossman, A. Photoprotection in cyanobacteria: Regulation of light harvesting. Photochem. Photobiol. 2008, 84, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Gubernator, B.; Bartoszewski, R.; Kroliczewski, J.; Wildner, G.; Szczepaniak, A. Ribulose-1,5-bisphosphate carboxylase/oxygenase from thermophilic cyanobacterium Thermosynechococcus elongatus. Photosynth. Res. 2008, 95, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Jordan, D.B.; Ogren, W.L. Species variation in the specificity of ribulose biphosphate carboxylase/oxygenase. Nature 1981, 291, 513–515. [Google Scholar] [CrossRef]
- Eisenhut, M.; Ruth, W.; Haimovich, M.; Bauwe, H.; Kaplan, A.; Hagemann, M. The photorespiratory glycolate metabolism is essential for cyanobacteria and might have been conveyed endosymbiontically to plants. Proc. Natl. Acad. Sci. USA 2008, 105, 17199–17204. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.G.; Turpin, D.H.; Canvin, D.T. Growth and photosynthesis of the cyanobacterium Synechococcus leopoliensis in -limited chemostats. Plant Physiol. 1984, 75, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Rohr, R.P.; Saavedra, S.; Bascompte, J. On the structural stability of mutualistic systems. Nature 2014, 345, 1253497. [Google Scholar] [CrossRef] [PubMed]
- Mayberry, W.R.; Prochacka, G.J.; Payne, W.J. Factors derived from studies of aerobic growth in minimal media. J. Bacteriol. 1968, 96, 1424–1426. [Google Scholar] [PubMed]
- Benschop, J.J.; Badger, M.R.; Price, G.D. Characterisation of CO2 and uptake in the cyanobacterium Synechocystis sp. PCC6803. Photosynth. Res. 2003, 77, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Badger, M.R.; Hanson, D.; Price, G.D. Evolution and diversity of CO2 concentrating mechanisms in cyanobacteria. Funct. Plant Biol. 2002, 29, 161–173. [Google Scholar] [CrossRef]
- Al-Najjar, M.A.A.; de Beer, D.; Jørgensen, B.B.; Kühl, M.; Polerecky, L. Conversion and conservation of light energy in a photosynthetic microbial mat ecosystem. ISME J. 2010, 4, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tannahill, A.L.; Shuler, M.L. Design of a system for the control of low dissolved oxygen concentrations: critical oxygen concentrations for Azotobacter vinelandii and Escherichia coli. Biotechnol. Bioeng. 1985, 27, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Shaler, T.A.; Klecka, G.M. Effects of dissolved oxygen concentration on biodegradation of 2,4-dichlorophenoxyacetic acid. Appl. Environ. Microbiol. 1986, 51, 950–955. [Google Scholar] [PubMed]
- Füchslin, H.P.; Schneider, C.; Egli, T. In glucose-limited continuous culture the minimum substrate concentration for growth, Smin, is crucial in the competition between the enterobacterium Escherichia coli and Chelatobacter heintzii, an environmentally abundant bacterium. ISME J. 2012, 6, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Nowack, S.; Olsen, M.T.; Schaible, G.A.; Becraft, E.D.; Shen, G.; Klapper, I.; Bryant, D.A.; Ward, D.M. The molecular dimension of microbial species: 2. Synechococcus strains representative of putative ecotypes inhabiting different depths in the Mushroom Spring microbial mat exhibit different adaptive and acclimative responses to light. Front. Microbiol. 2015, 6, 626. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State Quantities | Key Environmental Parameters | ||||
|---|---|---|---|---|---|
| Symbol | Description | Units | Symbol | Description | Units |
| P | Phototroph Concentration | Cmol·L | D | Dilution Rate | s |
| P | Heterotroph Concentration | Cmol·L | ν | Photon Flux | E m·s |
| IC | Inorganic Carbon Concentration | Cmol·L | IC | Inflow IC Conc. | Cmol·L |
| OC | Organic Carbon Concentration | Cmol·L | |||
| O | Oxygen Concentration | Omol·L | O | Inflow O Conc. | Omol·L |
| Rate Functions | Yield Parameters | ||||
|---|---|---|---|---|---|
| Symbol | Description | Units | Symbol | Description | Reference |
| η | Photorespiration Branching Function | – | |||
| Photosynthesis Rate | s | Phototroph Yields α per β | see Section 2.3.1 | ||
| Photobiosynthesis Rate | |||||
| Photorespiration rate | |||||
| Heterotroph Biosynthesis Rate | Heterotroph Yields α per β | see Section 2.4 | |||
| Symbol | Description | Units | Definition |
|---|---|---|---|
| A | Average cell cross-sectional area | m | Appendix B |
| c | Carbon moles per cell | Cmole/cell | Appendix B |
| e | Electron production rate by the light reaction | emole/Cmole·s | Equation (12) |
| f | Net oxygen per photosynthetically fixed carbon | Omole/Cmole | Equation (10) |
| I | Photoinhibition function | – | Equation (15) |
| α | photosynthesis efficiency factor | – | Equation (12) |
| inverse specificity factor | Cmole/Omole | Equation (16) | |
| excess electron capacity | s/Omole | Equation (15) | |
| ϵ | Maximum electron consumption rate | 1/s | Equation (13) |
| η | RuBisCO inorganic carbon binding probability | – | Equation (16) |
| ν | environmental photon flux | E/ms | – |
| ω | Electron demand: emoles needed to fix a cmole | emole/Cmole | Equation (9) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Moustaid, F.; Carlson, R.P.; Villa, F.; Klapper, I. Photorespiration and Rate Synchronization in a Phototroph-Heterotroph Microbial Consortium. Processes 2017, 5, 11. https://doi.org/10.3390/pr5010011
El Moustaid F, Carlson RP, Villa F, Klapper I. Photorespiration and Rate Synchronization in a Phototroph-Heterotroph Microbial Consortium. Processes. 2017; 5(1):11. https://doi.org/10.3390/pr5010011
Chicago/Turabian StyleEl Moustaid, Fadoua, Ross P. Carlson, Federica Villa, and Isaac Klapper. 2017. "Photorespiration and Rate Synchronization in a Phototroph-Heterotroph Microbial Consortium" Processes 5, no. 1: 11. https://doi.org/10.3390/pr5010011
APA StyleEl Moustaid, F., Carlson, R. P., Villa, F., & Klapper, I. (2017). Photorespiration and Rate Synchronization in a Phototroph-Heterotroph Microbial Consortium. Processes, 5(1), 11. https://doi.org/10.3390/pr5010011