Genome-Scale In Silico Analysis for Enhanced Production of Succinic Acid in Zymomonas mobilis



Abstract
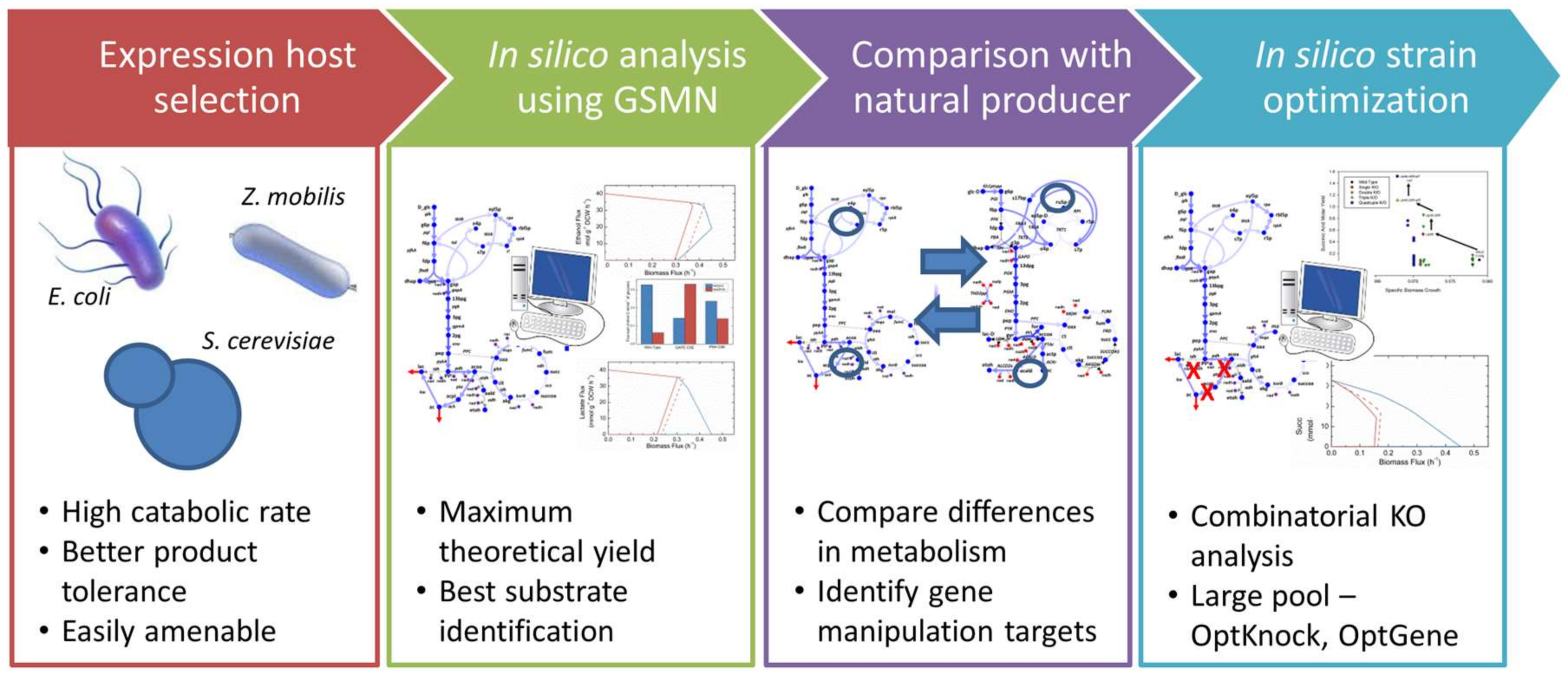
:1. Introduction
2. Materials and Methods
2.1. Genome-Scale Metabolic Model of Z. mobilis
2.2. Constraints-Based Flux Analysis
2.3. Combinatorial Knockout Simulation
2.4. Flux-Sum Quantification
3. Results and Discussion
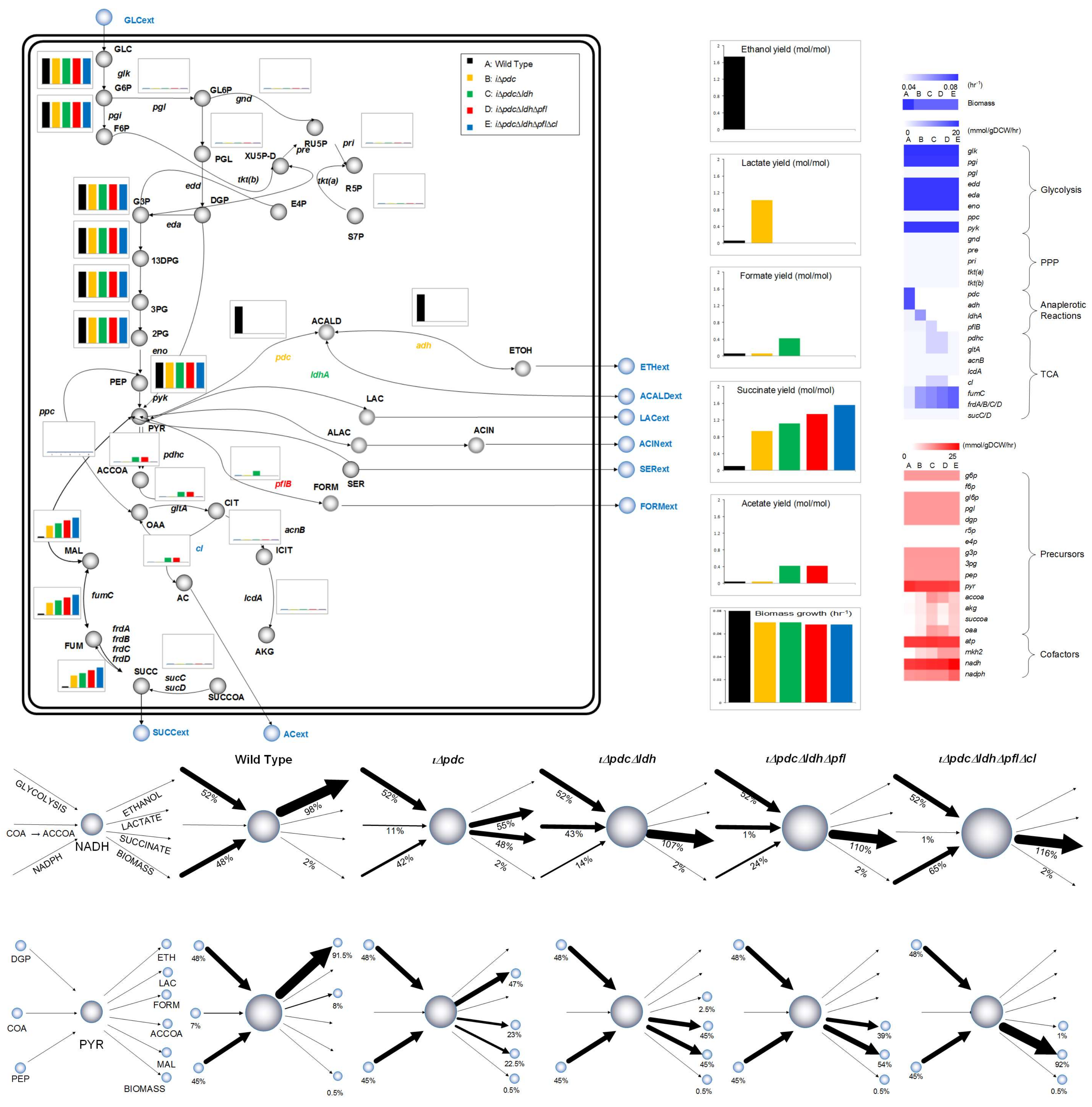
3.1. Exploring Metabolic Capabilities for Succinic Acid Production in Z. mobilis
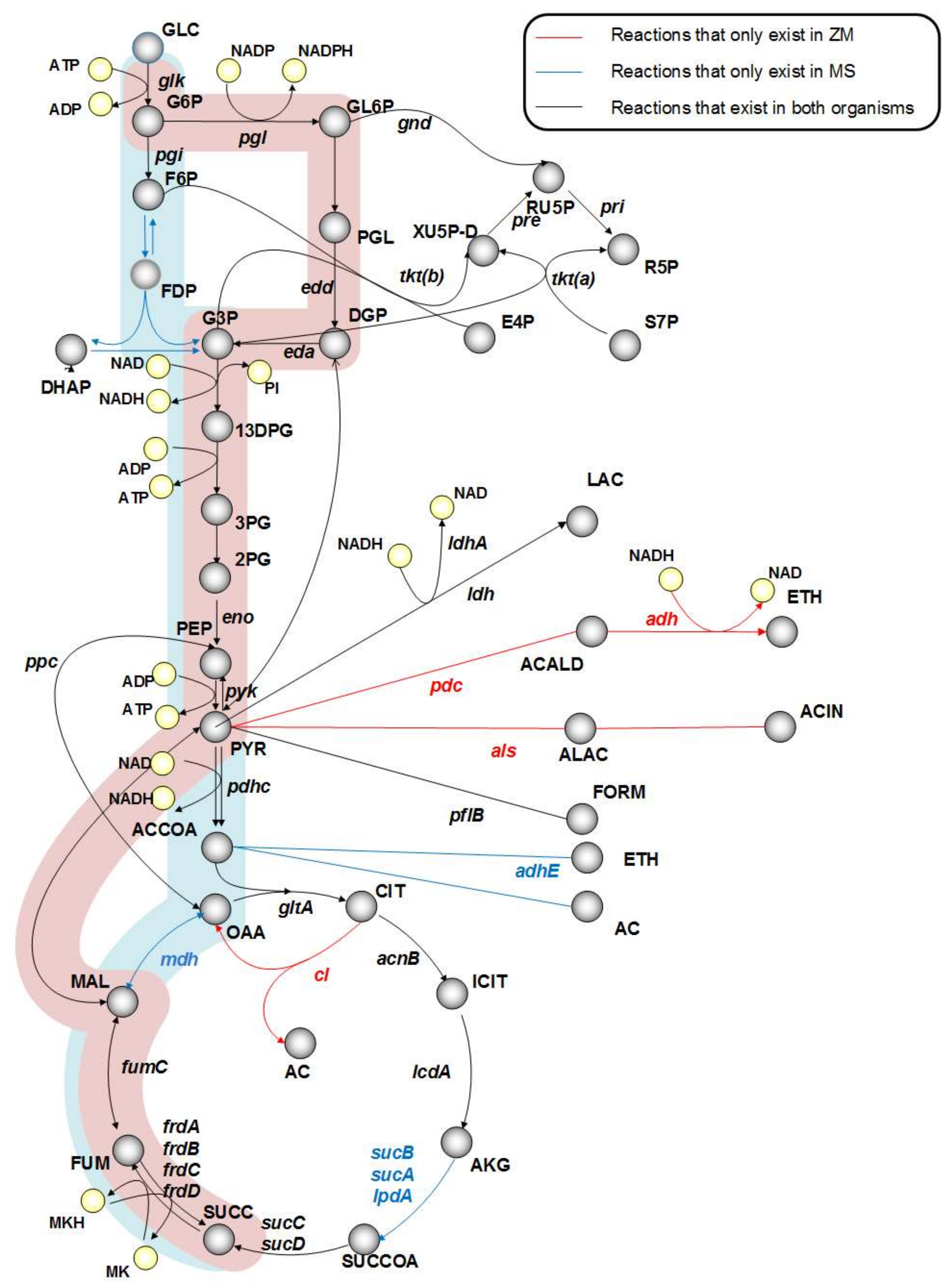
3.2. Central Metabolism Comparison with SA Producer to Identify Gene Candidates in Z. mobilis
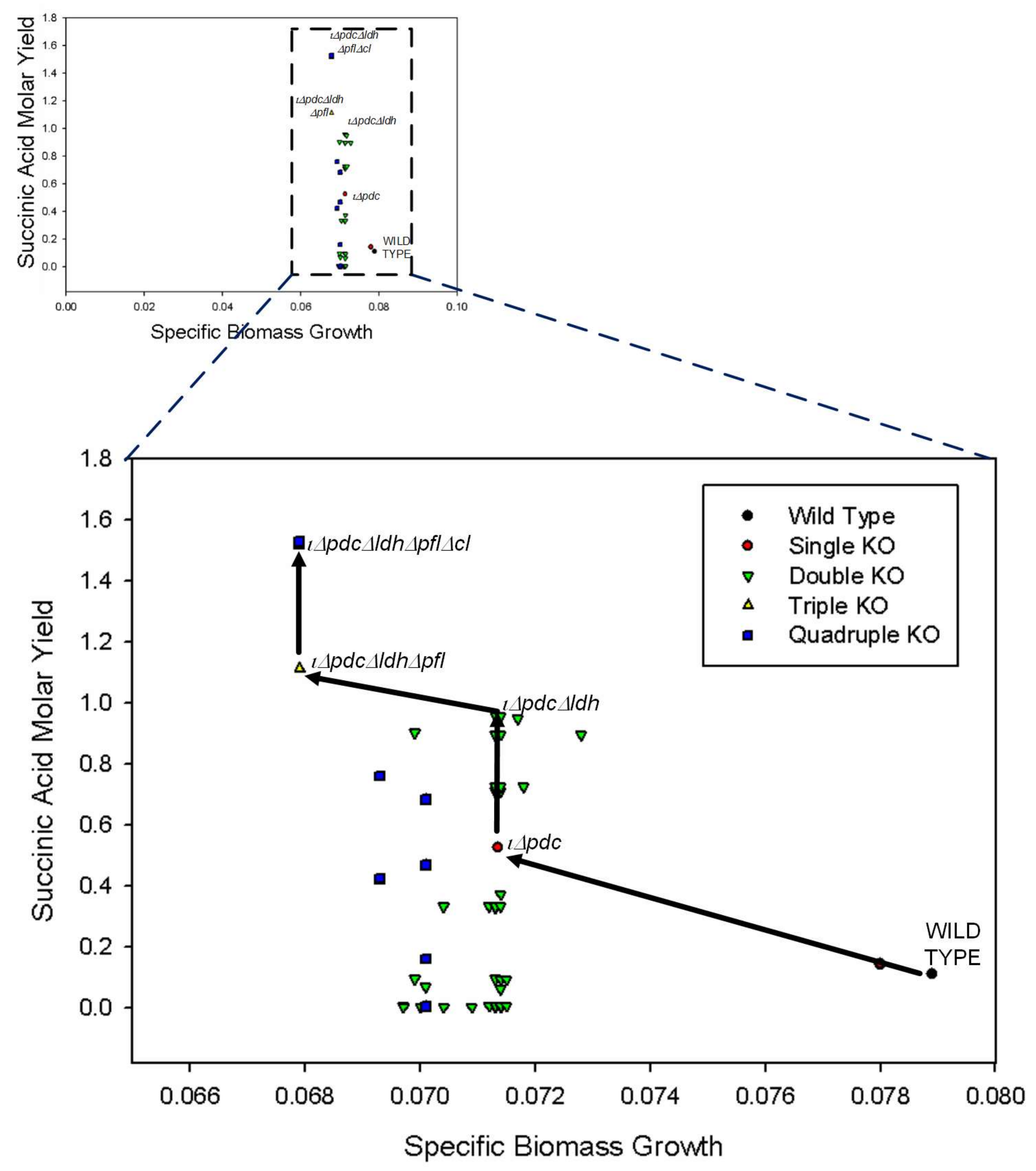
3.3. Combinatorial Knockout Analysis
3.4. Model-Driven Systematic Framework for Strain Optimization
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cukalovic, A.; Stevens, C.V. Feasibility of production methods for succinic acid derivatives: A marriage of renewable resources and chemical technology. Biofuels Bioprod. Biorefining 2008, 2, 505–529. [Google Scholar] [CrossRef]
- Beauprez, J.J.; De Mey, M.; Soetaert, W.K. Microbial succinic acid production: Natural versus metabolic engineered producers. Process Biochem. 2010, 45, 1103–1114. [Google Scholar] [CrossRef]
- Tan, T.; Liu, C.; Liu, L.; Zhang, K.; Zou, S.; Hong, J.; Zhang, M. Hydrogen sulfide formation as well as ethanol production in different media by cysND- and/or cysIJ-Inactivated mutant strains of Zymomonas mobilis ZM4. Bioprocess Biosyst. Eng. 2013, 36, 1363–1373. [Google Scholar] [CrossRef] [PubMed]
- Jantama, K.; Haupt, M.J.; Svoronos, S.A.; Zhang, X.; Moore, J.C.; Shanmugam, K.T.; Ingram, L.O. Combining metabolic engineering and metabolic evolution to develop nonrecombinant strains of Escherichia coli C that produce succinate and malate. Biotechnol. Bioeng. 2008, 99, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Thakker, C.; Martínez, I.; San, K.-Y.; Bennett, G.N. Succinate production in Escherichia coli. Biotechnol. J. 2012, 7, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.W.; Xia, S.T.; Wei, L.N.; Li, H.M.; Yuan, Z.P.; Tang, Y.J. Enhancing succinic acid biosynthesis in Escherichia coli by engineering its global transcription factor, catabolite repressor/activator (Cra). Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Guan, Z.; Wang, Y.; Ma, J.; Wu, H.; Jiang, M. Efficient succinic acid production by engineered Escherichia coli using ammonia as neutralizer. J. Chem. Technol. Biotechnol. 2016, 91, 2412–2418. [Google Scholar] [CrossRef]
- Mienda Faezah Mohd, B.S. Bio-succinic acid production: Escherichia coli strains design from genome-scale perspectives. AIMS Bioeng. 2017, 4, 418–430. [Google Scholar] [CrossRef]
- Lee, S.; Lee, D.; Kim, T.; Kim, B. Metabolic engineering of Escherichia coli for enhanced production of succinic acid, based on genome comparison and in silico gene knockout simulation. Appl. Environ. Microbiol. 2005, 71, 7880–7887. [Google Scholar] [CrossRef] [PubMed]
- Millard, C.S.; Chao, Y.P.; Liao, J.C.; Donnelly, M.I. Enhanced production of succinic acid by overexpression of phosphoenolpyruvate carboxylase in Escherichia coli. Appl. Environ. Microbiol. 1996, 62, 1808–1810. [Google Scholar] [PubMed]
- Wang, X.; Gong, C.S.; Tsao, G.T. Bioconversion of fumaric acid to succinic acid by recombinant E. coli. Appl. Biochem. Biotechnol. 1998, 70–72, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Effects of growth mode and pyruvate carboxylase on succinic acid production by metabolically engineered strains of Escherichia coli. Appl. Environ. Microbiol. 2002, 68, 1715–1727. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Li, Q.; Mao, Y.; Xing, J.; Su, Z. High-level succinic acid production and yield by lactose-induced expression of phosphoenolpyruvate carboxylase in ptsG mutant Escherichia coli. Appl. Microbiol. Biotechnol. 2010, 87, 2025–2035. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Kim, J.S.; Lee, S.Y.; In, Y.H.; Choi, S.S.; Rih, J.K.; Kim, C.H.; Jeong, H.; Hur, C.G.; Kim, J.J. The genome sequence of the capnophilic rumen bacterium Mannheimia succiniciproducens. Nat. Biotechnol. 2004, 22, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Stols, L.; Donnelly, M.I. Production of succinic acid through overexpression of NAD+-dependent malic enzyme in an Escherichia coli mutant. Appl. Environ. Microbiol. 1997, 63, 2695–2701. [Google Scholar] [PubMed]
- Lin, H.; Bennett, G.N.; San, K.Y. Fed-batch culture of a metabolically engineered Escherichia coli strain designed for high-level succinate production and yield under aerobic conditions. Biotechnol. Bioeng. 2005, 90, 775–779. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Bennett, G.N.; San, K.Y. Genetic reconstruction of the aerobic central metabolism in Escherichia coli for the absolute aerobic production of succinate. Biotechnol. Bioeng. 2005, 89, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Bennett, G.N.; San, K.Y. Metabolic engineering of aerobic succinate production systems in Escherichia coli to improve process productivity and achieve the maximum theoretical succinate yield. Metab. Eng. 2005, 7, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Chayabutra, C.; Wu, J.; Ju, L.K. Succinic acid production with reduced by-product formation in the fermentation of Anaerobiospirillum succiniciproducens using glycerol as a carbon source. Biotechnol. Bioeng. 2001, 72, 41–48. [Google Scholar] [CrossRef]
- Lee, P.C.; Lee, W.G.; Kwon, S.; Lee, S.Y.; Chang, H.N. Succinic acid production by Anaerobiospirillum succiniciproducens: Effects of the H2/CO2 supply and glucose concentration. Enzyme Microb. Technol. 1999, 24, 549–554. [Google Scholar] [CrossRef]
- McKinlay, J.B.; Laivenieks, M.; Schindler, B.D.; McKinlay, A.A.; Siddaramappa, S.; Challacombe, J.F.; Lowry, S.R.; Clum, A.; Lapidus, A.L.; Burkhart, K.B.; et al. A genomic perspective on the potential of Actinobacillus succinogenes for industrial succinate production. BMC Genom. 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Park, D.H.; Zeikus, J.G. Utilization of electrically reduced neutral red by Actinobacillus succinogenes: Physiological function of neutral red in membrane-driven fumarate reduction and energy conservation. J. Bacteriol. 1999, 181, 2403–2410. [Google Scholar] [PubMed]
- Bradfield, M.F.A.; Mohagheghi, A.; Salvachúa, D.; Smith, H.; Black, B.A.; Dowe, N.; Beckham, G.T.; Nicol, W. Continuous succinic acid production by Actinobacillus succinogenes on xylose-enriched hydrolysate. Biotechnol. Biofuels 2015, 8, 181. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Song, H.; Lee, S.Y. Genome-based metabolic engineering of Mannheimia succiniciproducens for succinic acid production. Appl. Environ. Microbiol. 2006, 72, 1939–1948. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Yi, J.; Kim, T.Y.; Choi, S.; Ahn, J.H.; Song, H.; Lee, M.-H.; Lee, S.Y. Homo-succinic acid production by metabolically engineered Mannheimia succiniciproducens. Metab. Eng. 2016, 38, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Johns, M.R.; Greenfield, P.F.; Doelle, H.W. Byproducts from Zymomonas mobilis. In Bioreactor Systems and Effects; Springer: Berlin/Heidelberg, Germany, 1991; pp. 97–121. ISBN 978-3-540-47400-5. [Google Scholar]
- Swings, J.; De Ley, J. The biology of Zymomonas. Bacteriol. Rev. 1977, 41, 1–46. [Google Scholar] [PubMed]
- Beer, L.L.; Boyd, E.S.; Peters, J.W.; Posewitz, M.C. Engineering algae for biohydrogen and biofuel production. Curr. Opin. Biotechnol. 2009, 20, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Martín, H.G.; Ivanova, N.; Kunin, V.; Warnecke, F.; Barry, K.W.; McHardy, A.C.; Yeates, C.; He, S.; Salamov, A.A.; Szeto, E.; et al. Metagenomic analysis of two enhanced biological phosphorus removal (EBPR) sludge communities. Nat. Biotechnol. 2006, 24, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, S.Y. Towards systems metabolic engineering of microorganisms for amino acid production. Curr. Opin. Biotechnol. 2008, 19, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Rossouw, D.; Næs, T.; Bauer, F.F. Linking gene regulation and the exo-metabolome: A comparative transcriptomics approach to identify genes that impact on the production of volatile aroma compounds in yeast. BMC Genom. 2008, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.Z.; Wu, C.Y.; Chen, T.; Chen, X.; Zhao, X.M. Integrating metabolomics into systems biology framework to exploit metabolic complexity: Strategies and applications in microorganisms. Appl. Microbiol. Biotechnol. 2006, 70, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Mienda, B.S.; Shamsir, M.S.; Illias, M.R. Model-assisted formate dehydrogenase-O (fdoH) gene knockout for enhanced succinate production in Escherichia coli from glucose and glycerol carbon sources. J. Biomol. Struct. Dyn. 2016, 34, 2305–2316. [Google Scholar] [CrossRef] [PubMed]
- Khodayari, A.; Chowdhury, A.; Maranas, C.D. Succinate Overproduction: A Case Study of Computational Strain Design Using a Comprehensive Escherichia coli Kinetic Model. Front. Bioeng. Biotechnol. 2015, 2. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Zeng, B.; Qian, X. Adaptive bi-level programming for optimal gene knockouts for targeted overproduction under phenotypic constraints. BMC Bioinform. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, K.; Yang, L.; Cluett, W.R.; Mahadevan, R. Dynamic strain scanning optimization: An efficient strain design strategy for balanced yield, titer, and productivity. DySScO strategy for strain design. BMC Biotechnol. 2013, 13. [Google Scholar] [CrossRef] [PubMed]
- Patil, K.R.; Åkesson, M.; Nielsen, J. Use of genome-scale microbial models for metabolic engineering. Curr. Opin. Biotechnol. 2004, 15, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Blazeck, J.; Alper, H. Systems metabolic engineering: Genome-scale models and beyond. Biotechnol. J. 2010, 5, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Teusink, B.; Van Enckevort, F.H.J.; Francke, C.; Wiersma, A.; Wegkamp, A.; Smid, E.J.; Siezen, R.J. In silico reconstruction of the metabolic pathways of Lactobacillus plantarum: Comparing predictions of nutrient requirements with those from growth experiments. Appl. Environ. Microbiol. 2005, 71, 7253–7262. [Google Scholar] [CrossRef] [PubMed]
- Burgard, A.P.; Pharkya, P.; Maranas, C.D. OptKnock: A Bilevel Programming Framework for Identifying Gene Knockout Strategies for Microbial Strain Optimization. Biotechnol. Bioeng. 2003, 84, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Pharkya, P.; Burgard, A.P.; Maranas, C.D. OptStrain: A computational framework for redesign of microbial production systems. Genome Res. 2004, 14, 2367–2376. [Google Scholar] [CrossRef] [PubMed]
- Patil, K.R.; Rocha, I.; Förster, J.; Nielsen, J. Evolutionary programming as a platform for in silico metabolic engineering. BMC Bioinform. 2005, 6. [Google Scholar] [CrossRef] [Green Version]
- Ranganathan, S.; Suthers, P.F.; Maranas, C.D. OptForce: An optimization procedure for identifying all genetic manipulations leading to targeted overproductions. PLoS Comput. Biol. 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.K.S.; Lee, D.Y. Flux-sum analysis: A metabolite-centric approach for understanding the metabolic network. BMC Syst. Biol. 2009, 3, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, Z.A.; Feist, A.M. Optimizing Cofactor Specificity of Oxidoreductase Enzymes for the Generation of Microbial Production Strains—OptSwap. Ind. Biotechnol. 2013, 9, 236–246. [Google Scholar] [CrossRef]
- Lakshmanan, M.; Chung, B.K.-S.; Liu, C.; Kim, S.-W.; Lee, D.-Y. Cofactor modification analysis: A computational framework to identify cofactor specificity engineering targets for strain improvement. J. Bioinform. Comput. Biol. 2013, 11, 1343006. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, M.; Lee, N.-R.; Lee, D.-Y. Genome-Scale Metabolic Modeling and In silico Strain Design of Escherichia coli. In Systems Biology; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2017; pp. 109–137. ISBN 9783527696130. [Google Scholar]
- Widiastuti, H.; Kim, J.Y.; Selvarasu, S.; Karimi, I.A.; Kim, H.; Seo, J.S.; Lee, D.Y. Genome-scale modeling and in silico analysis of ethanologenic bacteria Zymomonas mobilis. Biotechnol. Bioeng. 2011, 108, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Yun, H.; Park, S.; Lee, S.Y. MetaFluxNet: The management of metabolic reaction information and quantitative metabolic flux analysis. Bioinformatics 2003, 19, 2144–2146. [Google Scholar] [CrossRef] [PubMed]
- McCarl, B.A.; Meeraus, A.; van der Eijk, P.; Bussieck, M.; Dirkse, S.; Steacy, P. McCarl GAMS User Guide (2008). Available online: http//gams.com/docs/document.htm (accessed on 14 February 2004).
- Sang, Y.L.; Lee, D.Y.; Tae, Y.K. Systems biotechnology for strain improvement. Trends Biotechnol. 2005, 23, 349–358. [Google Scholar]
- Seo, J.S.; Chong, H.; Park, H.S.; Yoon, K.O.; Jung, C.; Kim, J.J.; Hong, J.H.; Kim, H.; Kim, J.H.; Kil, J.; et al. The genome sequence of the ethanologenic bacterium Zymomonas mobil is ZM4. Nat. Biotechnol. 2005, 23, 63–68. [Google Scholar] [CrossRef] [PubMed]
- DiMarco, A.A.; Romano, A.H. D-Glucose transport system of Zymomonas mobilis. Appl. Environ. Microbiol. 1985, 49, 151–157. [Google Scholar] [PubMed]
- Lee, K.Y.; Park, J.M.; Kim, T.Y.; Yun, H.; Lee, S.Y. The genome-scale metabolic network analysis of Zymomonas mobilis ZM4 explains physiological features and suggests ethanol and succinic acid production strategies. Microb. Cell Fact. 2010, 9. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.-S.; Chong, H.-Y.; Kim, J.H.; Kim, J.-Y. Method for Mass Production of Primary Metabolites, Strain for Mass Production of Primary Metabolites, and Method for Preparation Thereof. U.S. Patent Application 20090162910 A1, 25 June 2009. [Google Scholar]
- Hong, S.H.; Lee, S.Y. Importance of redox balance on the production of succinic acid by metabolically engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2002, 58, 286–290. [Google Scholar] [CrossRef] [PubMed]
- Meijer, S.; Nielsen, M.L.; Olsson, L.; Nielsen, J. Gene deletion of cytosolic ATP: Citrate lyase leads to altered organic acid production in aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1275–1280. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In Silico Strains | Growth Rate (1/h) | Molar Yield (mol Metabolite/mol Glucose) | ||||
|---|---|---|---|---|---|---|
| Succinic Acid | Ethanol | Lactic Acid | Formic Acid | Acetic Acid | ||
| Wild-Type ZM4 | 0.08 | 0.06 | 1.76 | 0 | 0 | 0 |
| Metabolism Comparison | ||||||
| Δpdc | 0.07 | 0.53 | 0 | 0.49 | 0.28 | 0.42 |
| Δals | 0 | 0 | 0 | 0 | 0 | 0 |
| Δcl | 0.08 | 0.06 | 1.76 | 0 | 0 | 0 |
| ΔpdcΔcl | 0.07 | 0.6 | 0 | 0.71 | 0.28 | 0 |
| Combinatorial Knockout | ||||||
| Δpdc | 0.07 | 0.53 | 0 | 0.49 | 0.28 | 0.42 |
| ΔpdcΔldh | 0.07 | 0.93 | 0 | 0 | 1.11 | 0.56 |
| ΔpdcΔpfl | 0.07 | 0.67 | 0 | 1.12 | 0 | 0 |
| ΔpdcΔcl | 0.07 | 0.6 | 0 | 0.71 | 0.28 | 0 |
| ΔpdcΔldhΔpfl | 0.07 | 1.14 | 0 | 0 | 0 | 0.75 |
| ΔpdcΔpflΔcl | 0.07 | 0.47 | 0 | 1.42 | 0 | 0 |
| ΔpdcΔldhΔcl | 0.07 | 1.09 | 0 | 0 | 1.64 | 0 |
| ΔpdcΔldhΔpflΔcl | 0.07 | 1.52 | 0 | 0 | 0 | 0 |
| OptKnock/OptGene | ||||||
| Δpdc | 0.07/0.07 | 0.53/0.66 | 0/0 | 0.59/0.44 | 0.59/0.94 | 0 |
| ΔpdcΔldh | 0.07/0.07 | 0.94/0.92 | 0/0 | 0.42/0.45 | 0/0 | 0.38/0.44 |
| ΔpdcΔldhΔpfl | 0.07/0.07 | 1.17/1.17 | 0/0 | 0/0 | 0/0 | 0.58/0.45 |
| ΔpdcΔldhΔpflΔcl | 0.07/0.07 | 1.42/1.42 | 0/0 | 0/0 | 0/0 | 0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widiastuti, H.; Lee, N.-R.; Karimi, I.A.; Lee, D.-Y. Genome-Scale In Silico Analysis for Enhanced Production of Succinic Acid in Zymomonas mobilis. Processes 2018, 6, 30. https://doi.org/10.3390/pr6040030
Widiastuti H, Lee N-R, Karimi IA, Lee D-Y. Genome-Scale In Silico Analysis for Enhanced Production of Succinic Acid in Zymomonas mobilis. Processes. 2018; 6(4):30. https://doi.org/10.3390/pr6040030
Chicago/Turabian StyleWidiastuti, Hanifah, Na-Rae Lee, Iftekhar A. Karimi, and Dong-Yup Lee. 2018. "Genome-Scale In Silico Analysis for Enhanced Production of Succinic Acid in Zymomonas mobilis" Processes 6, no. 4: 30. https://doi.org/10.3390/pr6040030
APA StyleWidiastuti, H., Lee, N. -R., Karimi, I. A., & Lee, D. -Y. (2018). Genome-Scale In Silico Analysis for Enhanced Production of Succinic Acid in Zymomonas mobilis. Processes, 6(4), 30. https://doi.org/10.3390/pr6040030