



Wound Healing Activity of the Flavonoid-Enriched Fraction of Selaginella bryopteris Linn. against Streptozocin-Induced Diabetes in Rats

 ,
,  , , ,
, , ,
Abstract
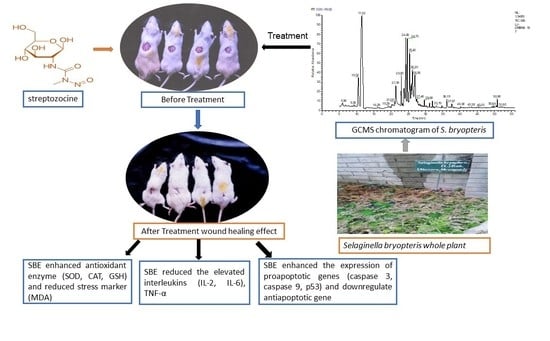
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Material and Methods
2.1. Reagents
2.2. Extract Preparation and Phytochemical Profiling
2.3. In Vitro Wound Healing Studies
2.3.1. Cell Culture
2.3.2. MTT Assay
2.3.3. Assessment of Nitric Oxide (NO)
2.3.4. Assessment of Cell Organelles Health Parameters
Nuclear Morphology Analysis Using DAPI Staining
Mitochondrial Membrane Potential Analysis Using JC-1 Staining
Morphological Evaluation for Apoptosis
2.4. In Vivo Wound Healing Study
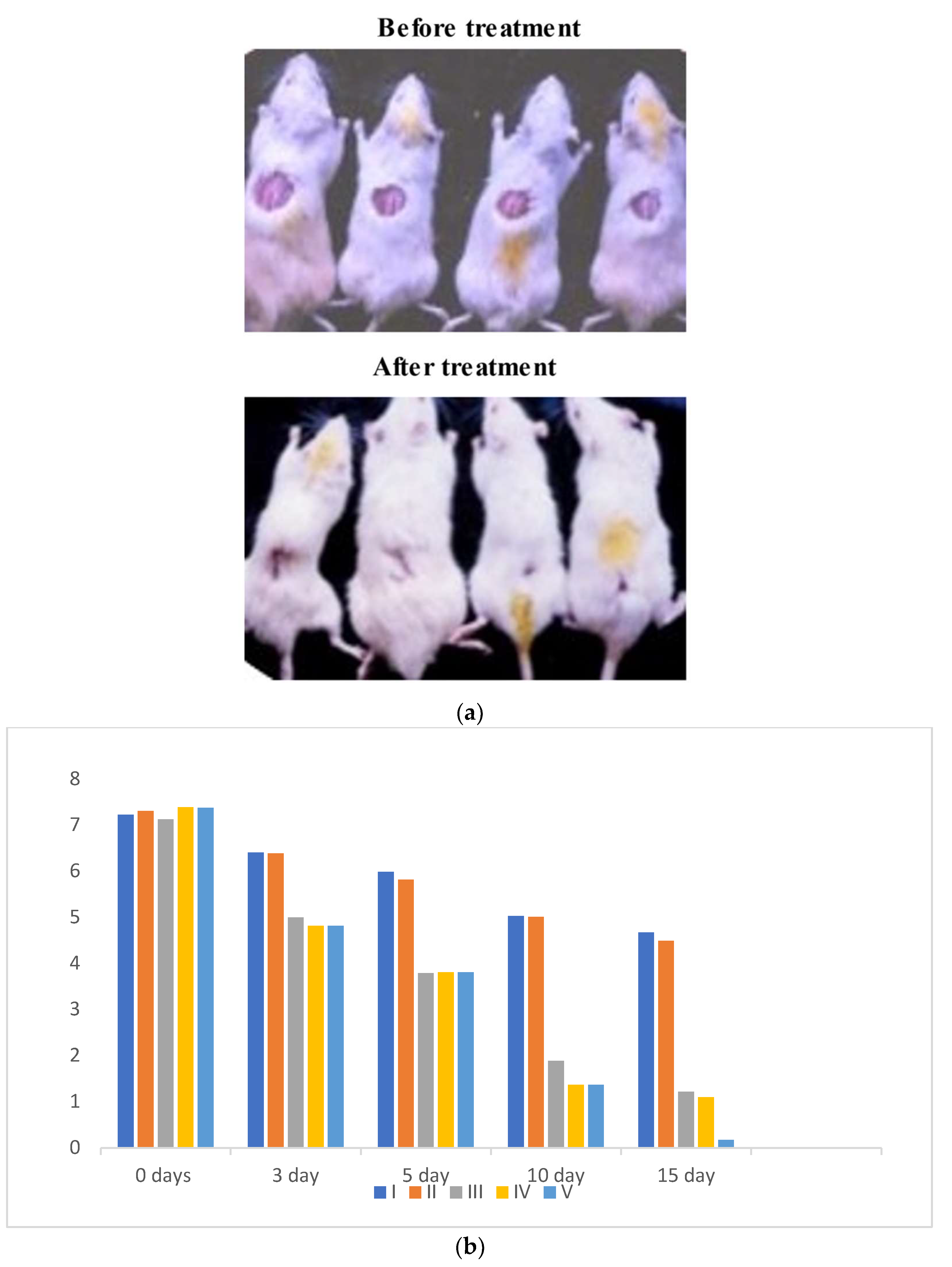
2.4.1. Experimental Animals and Protocol
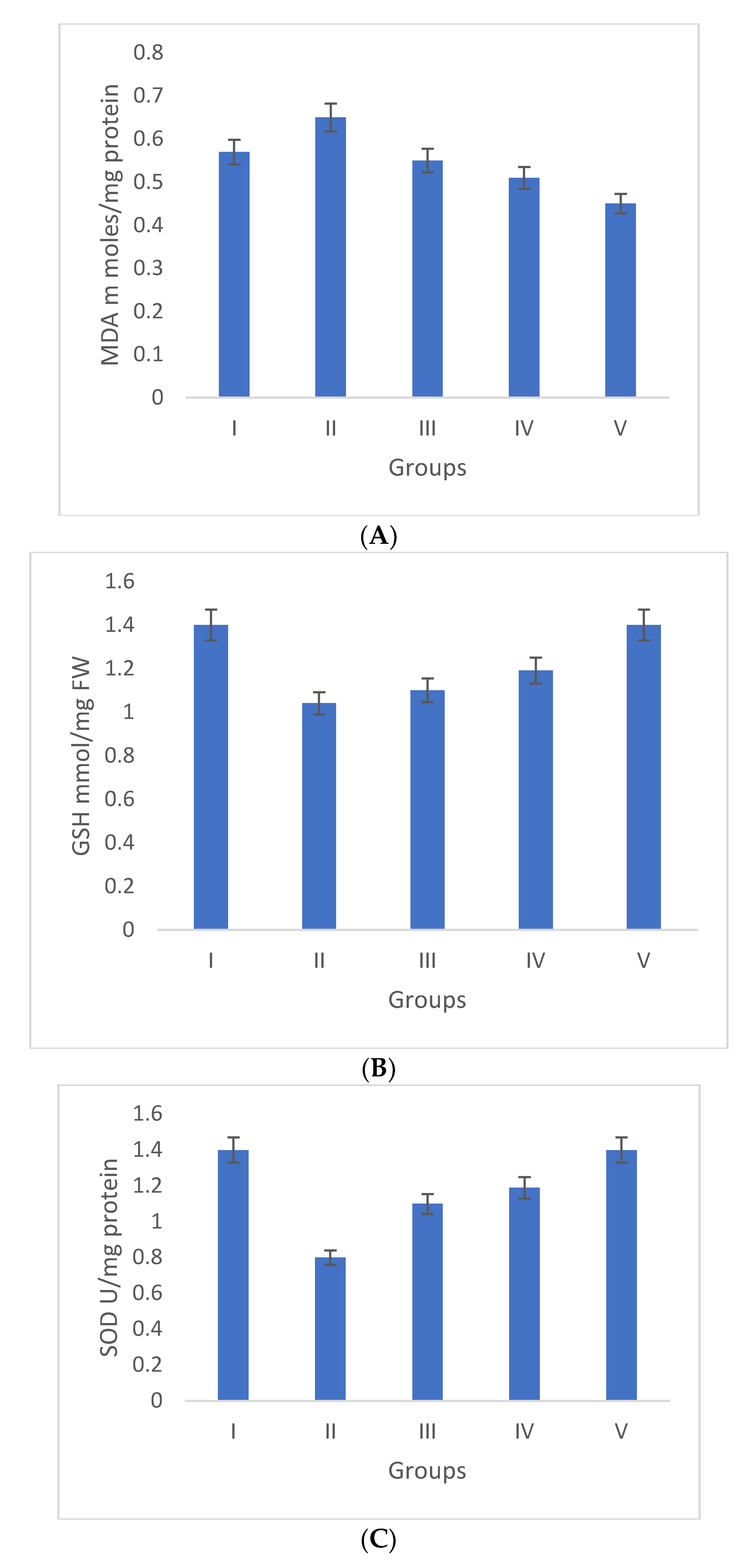
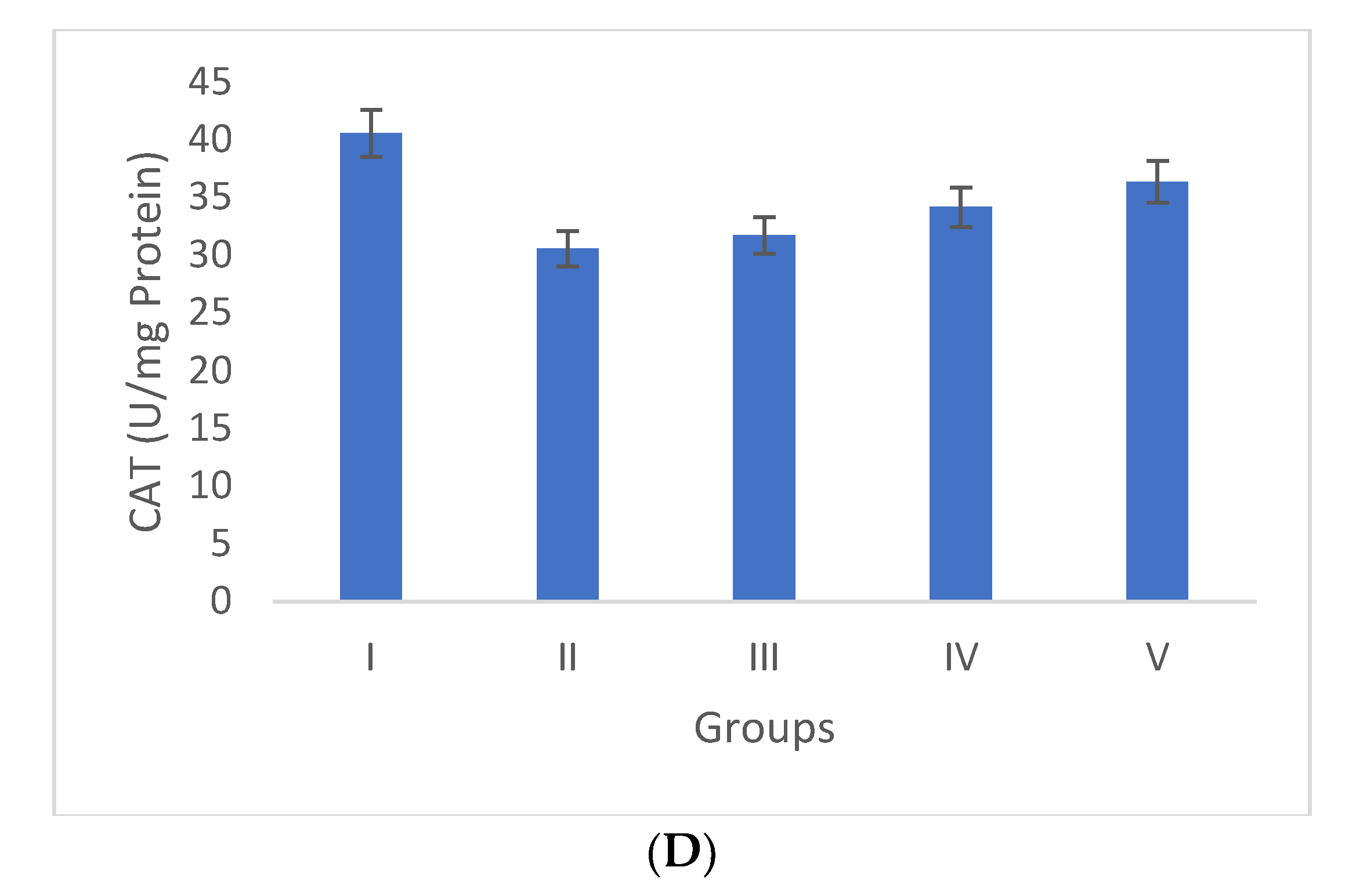
2.4.2. Effect of Selaginella bryopteris on Antioxidant Parameters
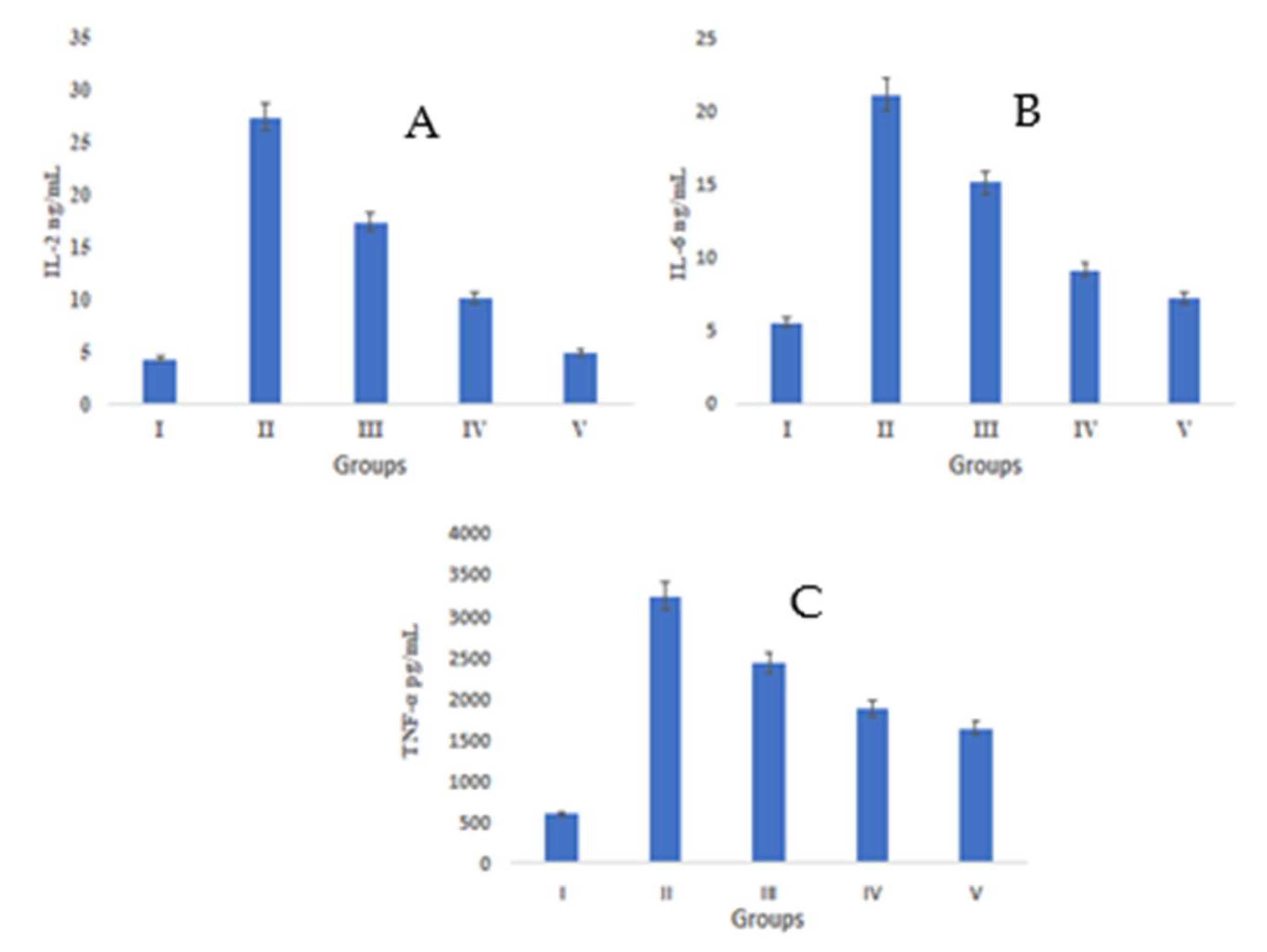
2.4.3. Assessment of Cytokine Levels in Wounded Tissue
2.5. Histological Analysis
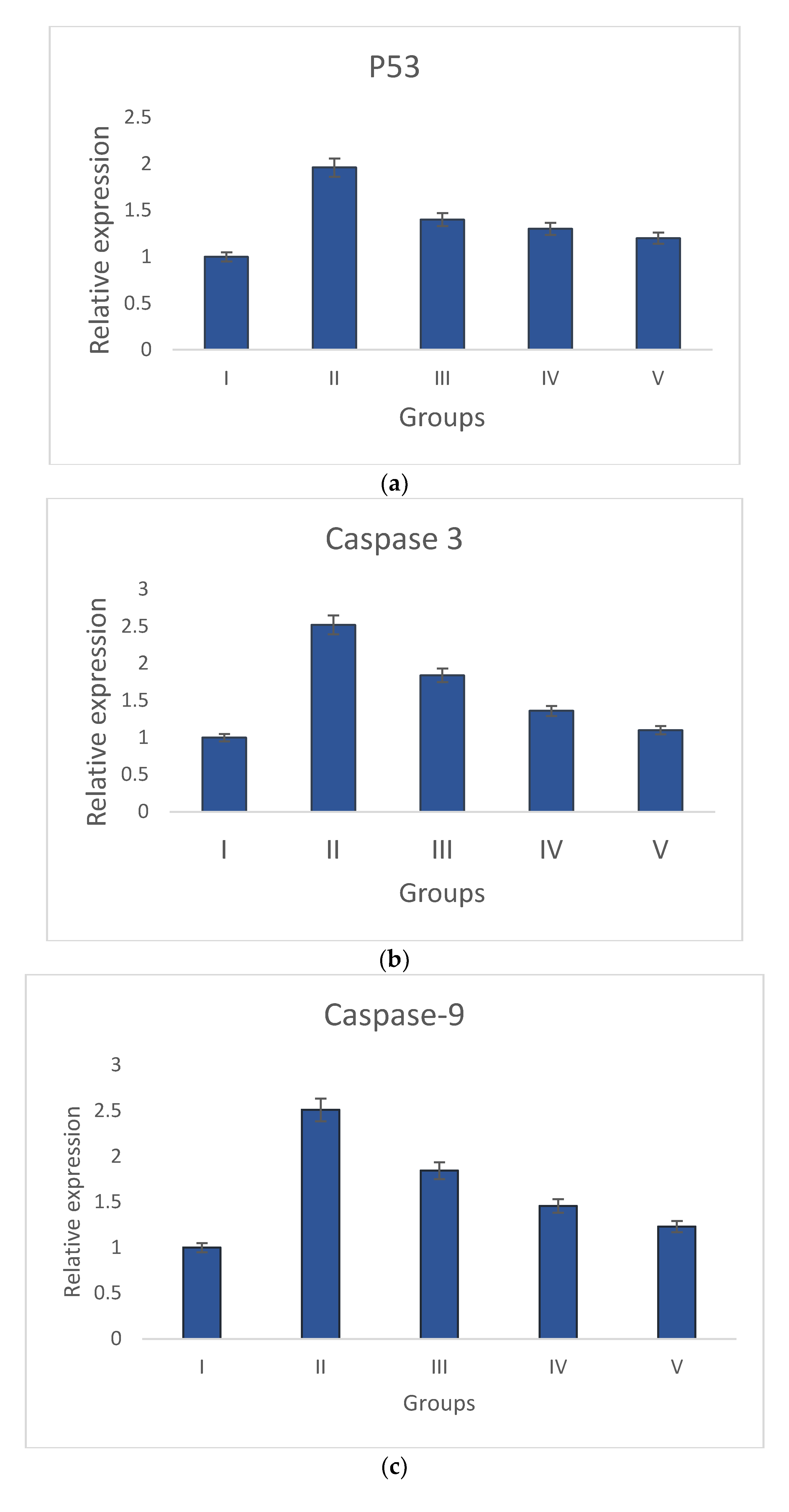
2.6. Gene Expression Study via Quantitative Real-Time PCR
2.7. Statistical Analysis
3. Results and Discussion
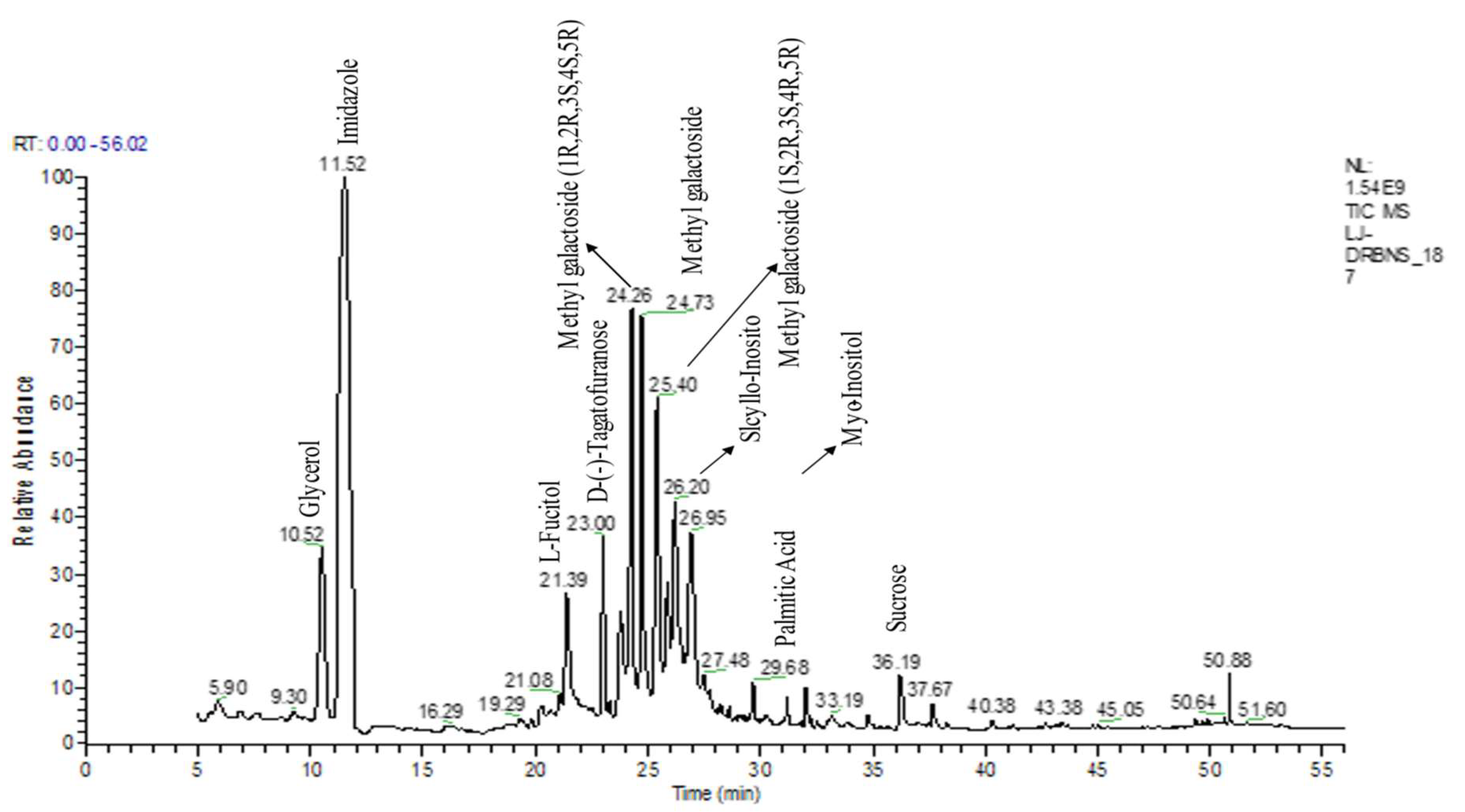
3.1. Phytochemical Profiling Selaginella bryopteris
3.2. In Vitro Wound Healing Study
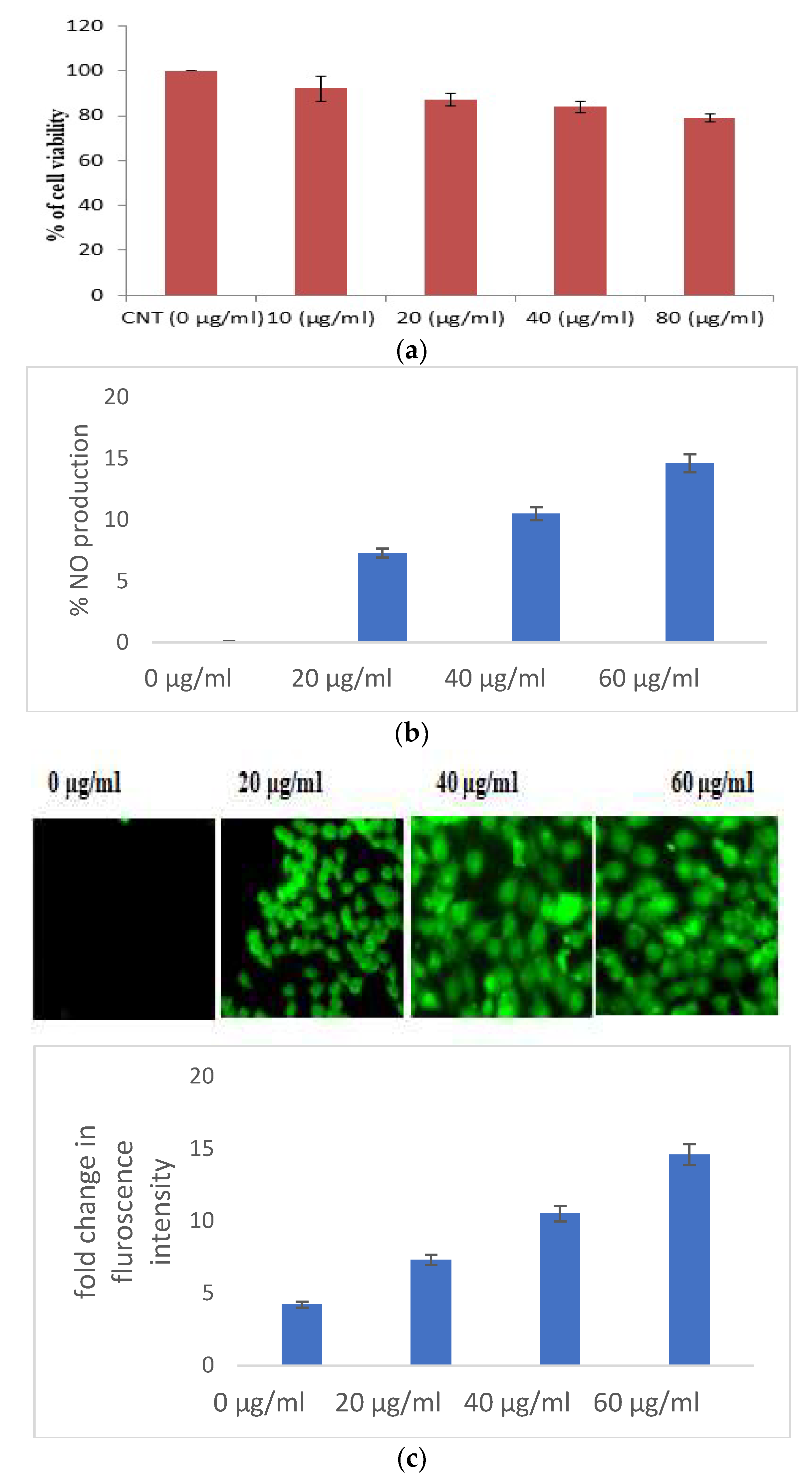
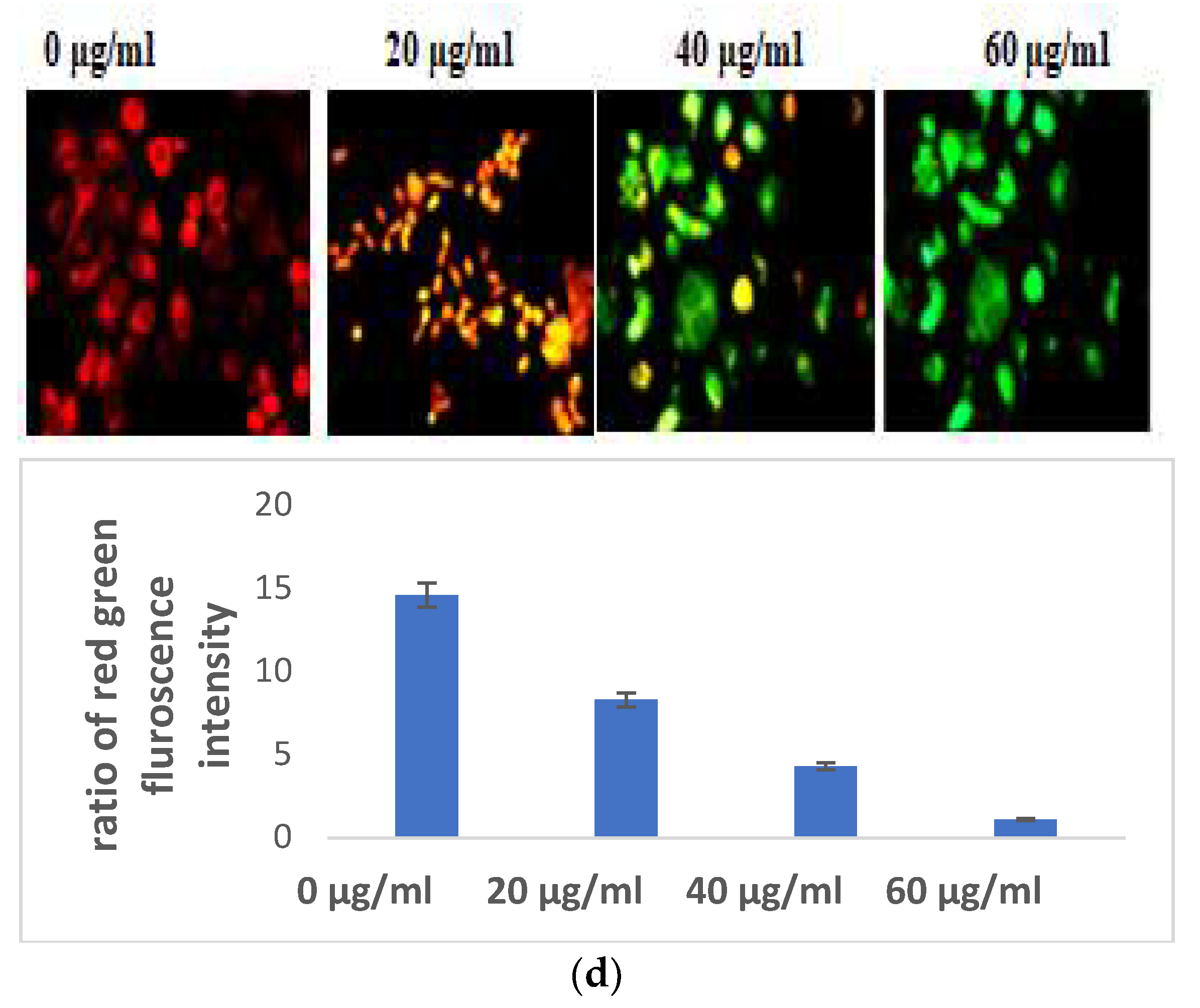
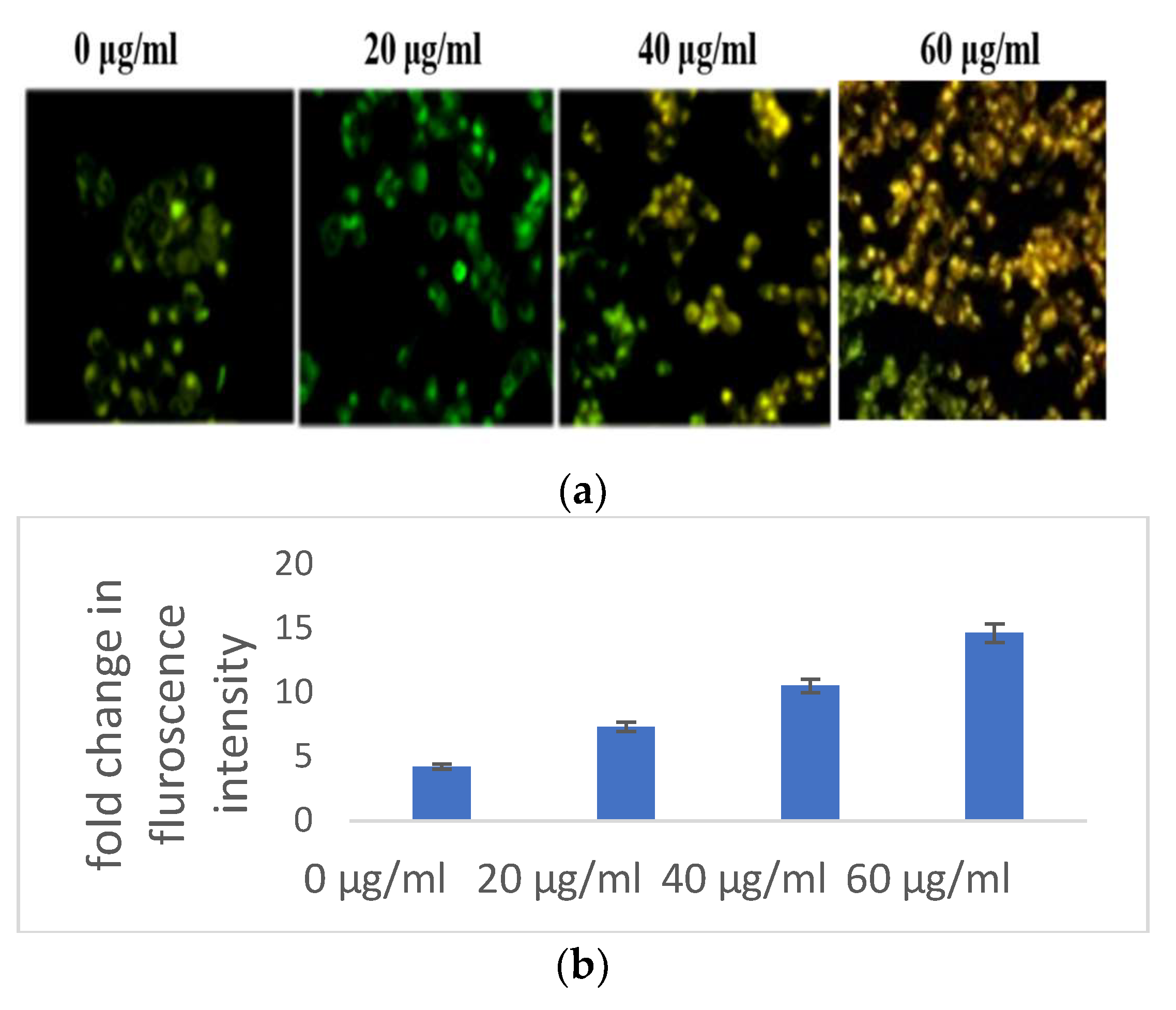
3.2.1. Selaginella bryopteris Induces Oxidative Stress in HaCaT Cells
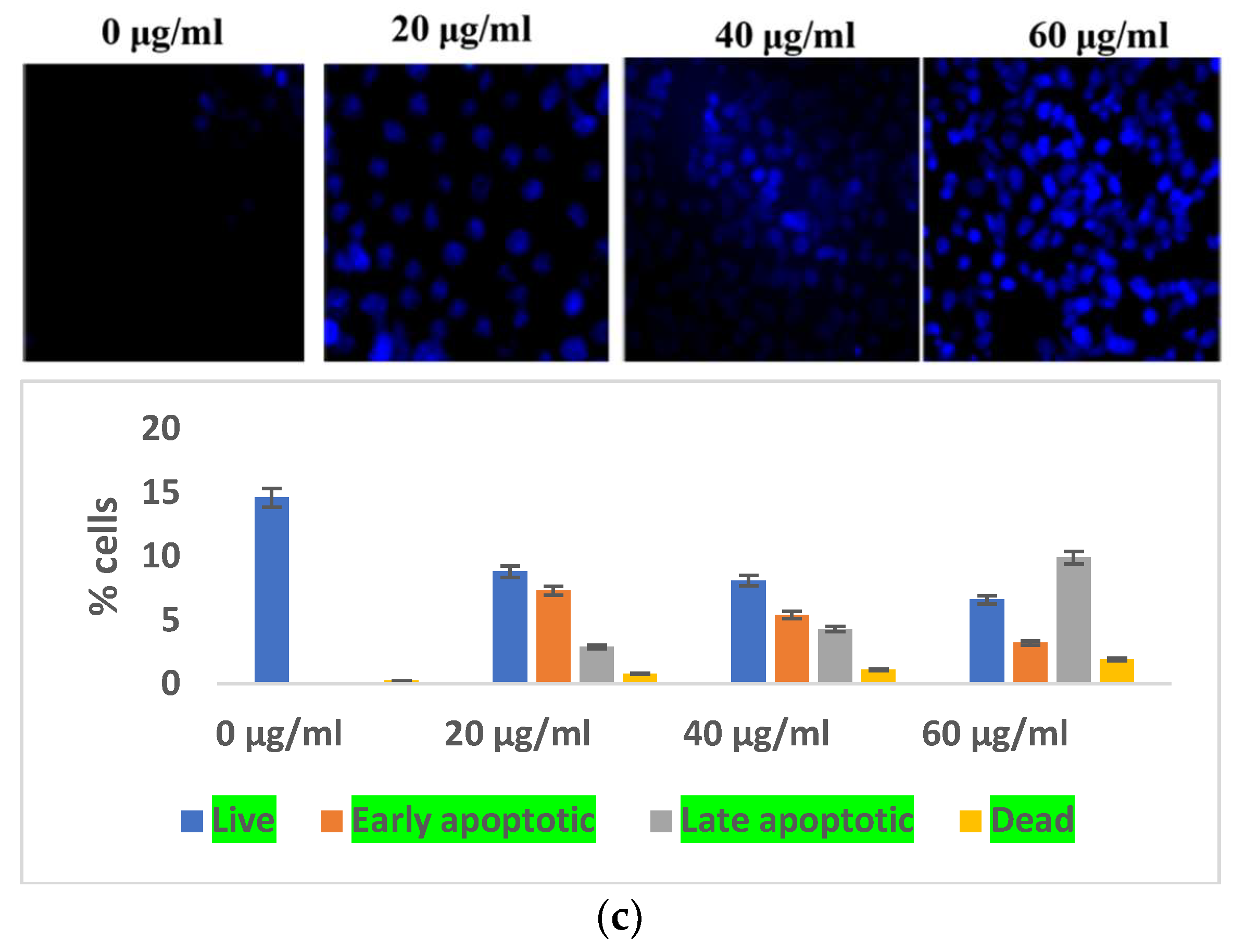
3.2.2. Selaginella bryopteris Causes Apoptotic-Mediated Cell Death
3.2.3. Selaginella bryopteris Affects Cell Organelle Health in HaCaT Cells
3.3. In Vivo Study
3.3.1. Antioxidant Markers
3.3.2. Elisa Quantified Interleukins and TNF-α
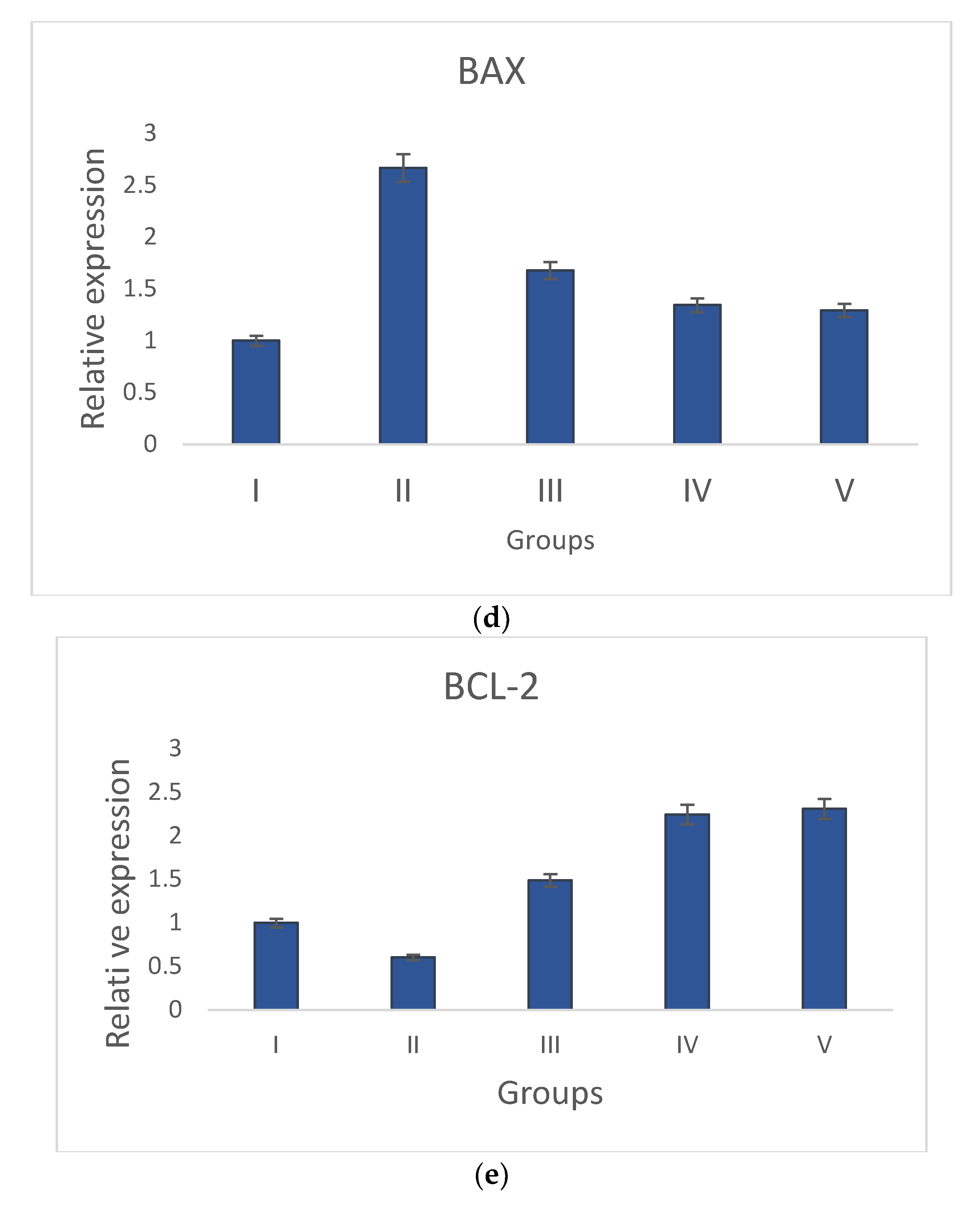
3.3.3. Gene Expression Analysis of Bcl-2, p53, Bax, Caspase-3, and Caspase-9 by qRT-PCR
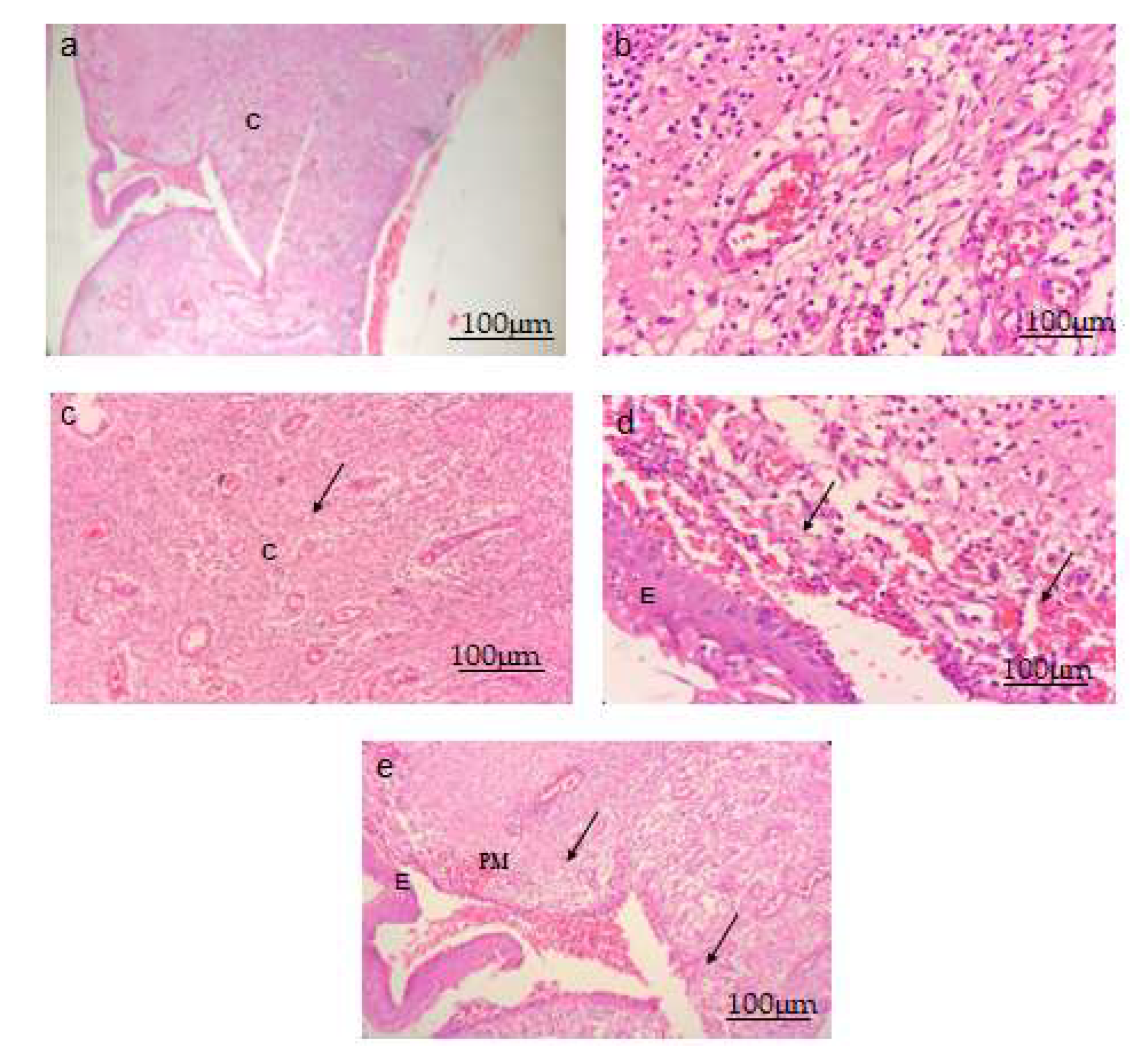
3.4. Histopathological Analysis
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Diabetes Federations. IDF Diabetes Atlas, 7th ed.; International Diabetes Federation: Brussels, Belgium, 2015. [Google Scholar]
- Bhan, S.; Mitra, R.; Arya, A.K.; Pandey, H.P.; Tripathi, K. A study on evaluation of apoptosis and expression of bcl-2-related marker in wound healing of streptozotocin-induced diabetic rats. Dermatology 2013, 2013, 739054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giannone, G.; Mazzone, O.; Russo, A.; Trovato, G.M. Electrocardiographic findings during colonoscopy. Boll. Soc. Ital. Cardiol. 1978, 23, 441–446. [Google Scholar] [PubMed]
- Siqueira, M.F.; Li, J.; Chehab, L.; Desta, T.; Chino, T.; Krothpali, N.; Behl, Y.; Alikhani, M.; Yang, J.; Braasch, C.; et al. Impaired wound healing in mouse models of diabetes is mediated by TNF-alpha dysregulation and associated with enhanced activation of forkhead box O1 (FOXO1). Diabetologia 2010, 53, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arya, A.K.; Pokharia, D.; Tripathi, K. Relationship between oxidative stress and apoptotic markers in lymphocytes of diabetic patients with chronic non healing wound. Diabetes Res. Clin. Pract. 2011, 94, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Chen, L. The worldwide epidemiology of type 2 diabetes mellitus–present and future 190 perspectives. Nat. Rev. Endocrinol. 2012, 8, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Wang, D.; Yu, B.; Yu, X.; Li, W. Maintenance or collapse: Responses of extraplastidic membrane lipid composition to desiccation in the resurrection plant Paraisometrummileense. PLoS ONE 2014, 9, 103430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sah, N.K.; Singh, S.N.P.; Sahdev, S.; Banerji, S.; Jha, V.; Khan, Z.; Hasnain, S.E. Indian herb ‘Sanjeevani’ (Selaginella bryopteris) can promote growth and protect against heat shock and apoptotic activities of ultra violet and oxidative stress. J. Biosci. 2005, 30, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Antony, R.; Thomas, R. A mini review on medicinal properties of the resurrecting plant Selaginella bryopteris (Sanjeevani). Int. J. Pharm. Life Sci. 2011, 2, 7. [Google Scholar]
- Kunert, O.; Swamy, R.C.; Kaiser, M.; Presser, A.; Buzzi, S.; Rao, A.A.; Schühly, W. Anti-plasmodial and leishmanicidal activity of bioflavonoid from Indian Selaginella bryopteris. Phytochem. Lett. 2008, 1, 171–174. [Google Scholar] [CrossRef]
- Paswan, S.K.; Gautam, A.; Verma, P.; Rao, C.V.; Sidhu, O.P.; Singh, A.P.; Srivastava, S. The Indian magical herb ‘Sanjeevni’ (Selaginella bryopteris L.)-a promising anti -inflammatory phytomedicine for the treatment of patients with inflammatory skin diseases. J. Pharmacopunct. 2017, 20, 93. [Google Scholar]
- Sah, P. Does the magical himalayan herb “Sanjeevani Booti” really exist in nature. J. Am. Sci. 2008, 4, 65–67. [Google Scholar]
- Ragazzi, E.; Veronese, G. Quantitative analysis of flavonoid compounds after thin layer chromatographic separation. J. Chrom. 1973, 77, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Oyaizu, M. Studies on products of browning reaction: Antioxidative activity of products of browning reaction. J. Nutr. Diet 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, A.; Bharti, S.K.; Tripathi, T.; Mishra, A.; Sidhu, O.P.; Roy, R.; Nautiyal, C.S. Metabolic profiling of Commiphora wightii (guggul) reveals a potential source for pharmaceuticals and nutraceuticals. Phytochemistry 2015, 110, 29–36. [Google Scholar] [CrossRef]
- Yadav, P.P.; Arora, A.; Bid, H.K.; Konwar, R.R.; Kanojiya, S. New cassane butenolide hemiketal diterpenes from the marine creeper Caesalpinia bonduc and their antiproliferative activity. Tetrahedron Lett. 2007, 48, 7194–7198. [Google Scholar] [CrossRef]
- Besra, S.E.; Ray, M.; Dey, S.; Roy, S.; Deb, N. Apoptogenic activity of secretion extract of Bellamya Bengalensis f. annandalei via mitochondrial mediated caspase cascade on human leukemic cell lines. Int. J. Pharm. Sci. Rev. Res. 2013, 20, 146–152. [Google Scholar]
- Nikhil, K.; Sharan, S.; Chakraborty, A.; Bodipati, N.; Peddinti, R.K.; Roy, P. Role of isothiocyanate conjugate of pterostilbene on the inhibition of MCF-7 cell proliferation and tumor growth in Ehrlich ascitic cell induced tumor bearing mice. Exp. Cell Res. 2014, 320, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Kakkar, P.; Das, B.; Viswanathan, P.N. A modified spectrophotometric assay of superoxide dismutase.of superoxide dismutase. Indian J. Biochem. Biophys. 1984, 21, 130–132. [Google Scholar] [PubMed]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Academic Press: Weinheim, Duitsland; NewYork, NY, USA, 1974; pp. 673–684. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys 1959, 82, 70–77. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gautam, A.; Kumar, V.; Azmi, L.; Rao, C.V.; Khan, M.M.; Mukhtar, B.; Kamal, M.; Arif, M.; Mehdi, S.; Alsanad, S.M.; et al. Wound Healing Activity of the Flavonoid-Enriched Fraction of Selaginella bryopteris Linn. against Streptozocin-Induced Diabetes in Rats. Separations 2023, 10, 166. https://doi.org/10.3390/separations10030166
Gautam A, Kumar V, Azmi L, Rao CV, Khan MM, Mukhtar B, Kamal M, Arif M, Mehdi S, Alsanad SM, et al. Wound Healing Activity of the Flavonoid-Enriched Fraction of Selaginella bryopteris Linn. against Streptozocin-Induced Diabetes in Rats. Separations. 2023; 10(3):166. https://doi.org/10.3390/separations10030166
Chicago/Turabian StyleGautam, Arti, Vikas Kumar, Lubna Azmi, Ch. V. Rao, Mohammed Moizuddin Khan, Beenish Mukhtar, Mehnaz Kamal, Muhammad Arif, Seema Mehdi, Saud M. Alsanad, and et al. 2023. "Wound Healing Activity of the Flavonoid-Enriched Fraction of Selaginella bryopteris Linn. against Streptozocin-Induced Diabetes in Rats" Separations 10, no. 3: 166. https://doi.org/10.3390/separations10030166
APA StyleGautam, A., Kumar, V., Azmi, L., Rao, C. V., Khan, M. M., Mukhtar, B., Kamal, M., Arif, M., Mehdi, S., Alsanad, S. M., Al-Khamees, O. A., Jawaid, T., & Alam, A. (2023). Wound Healing Activity of the Flavonoid-Enriched Fraction of Selaginella bryopteris Linn. against Streptozocin-Induced Diabetes in Rats. Separations, 10(3), 166. https://doi.org/10.3390/separations10030166