
Simple Isolation of Cordycepin from Cordyceps militaris by Dual-Normal Phase Column Chromatography and Its Potential for Making Kombucha Functional Products





Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Instrumentations
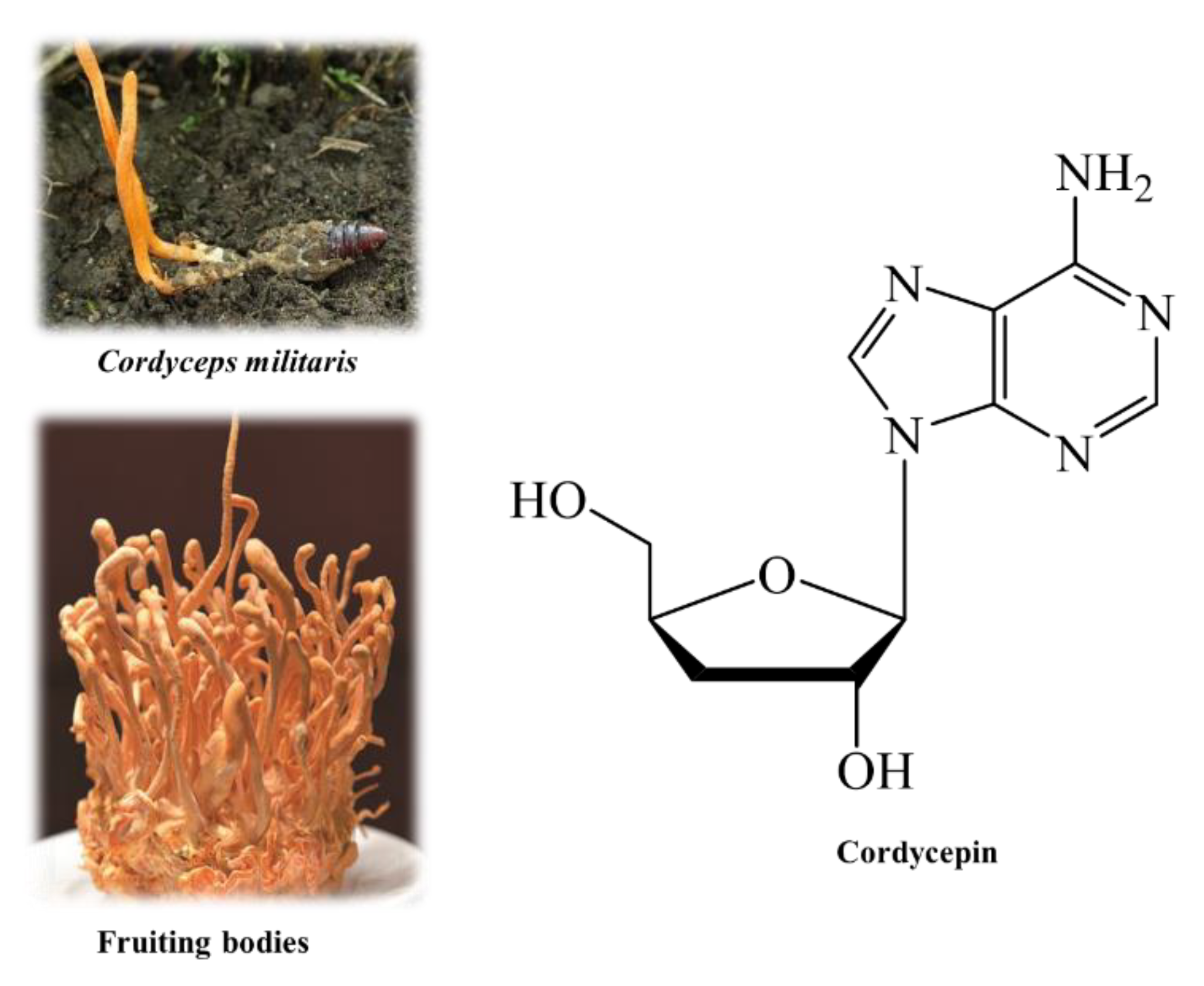
2.2. Cordyceps Millitaris Sample
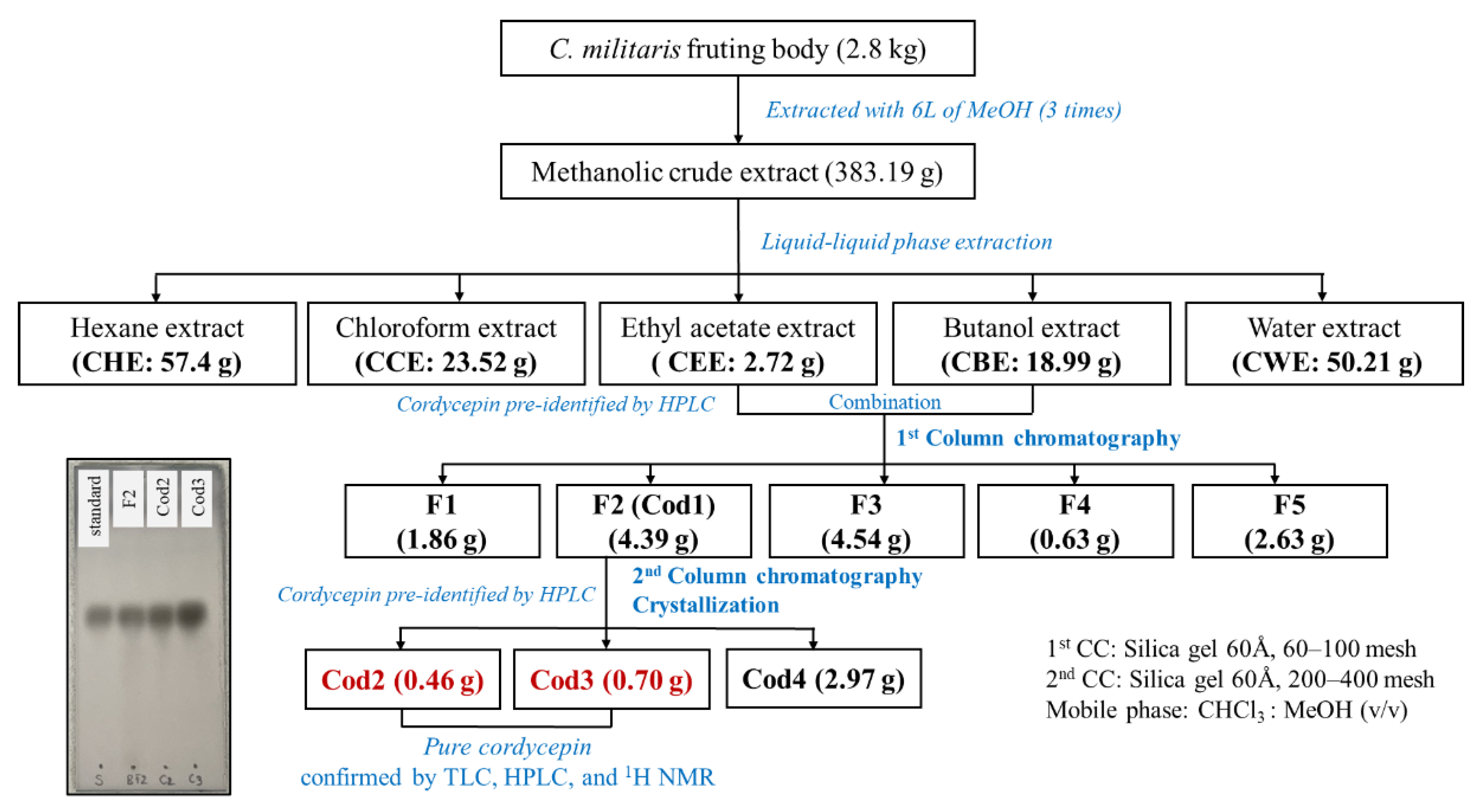
2.3. Sample Preparation and Extraction
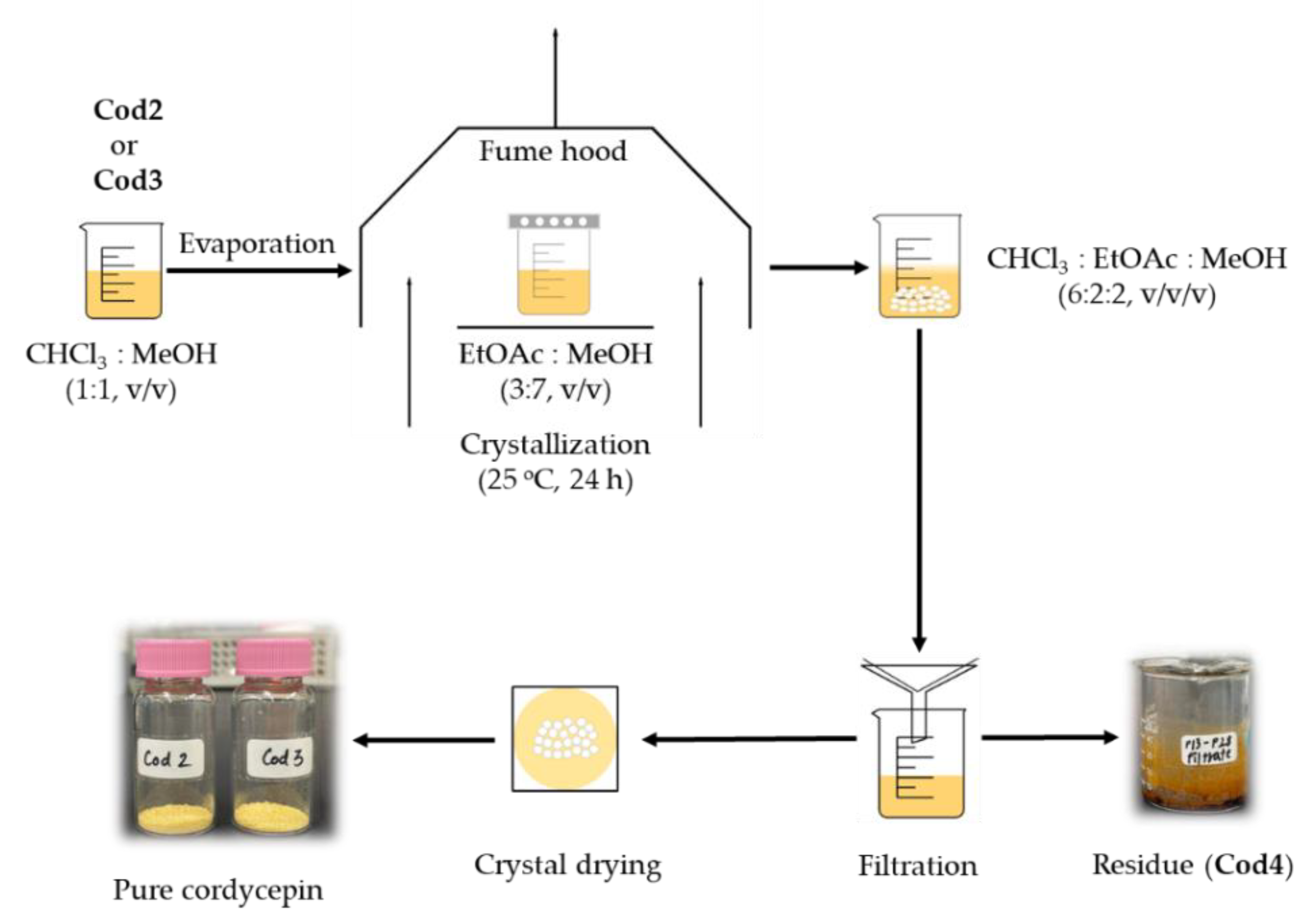
2.4. Fractionation, Isolation, Purification, and Crystalization of Cordycepin
2.5. Thin-Layer Chromatography and Spectroscopic Analyses
2.6. Kombucha Fermentation
2.7. Biological Acitivities
2.7.1. Antioxidant
2.7.2. α-Glucosidase Inhibitory Assay
2.7.3. Cytotoxic Activity against HL-60 and Meg-01 Cell Lines
2.8. Statistical Analysis
3. Results
3.1. Isolation and Purification of Cordycepin
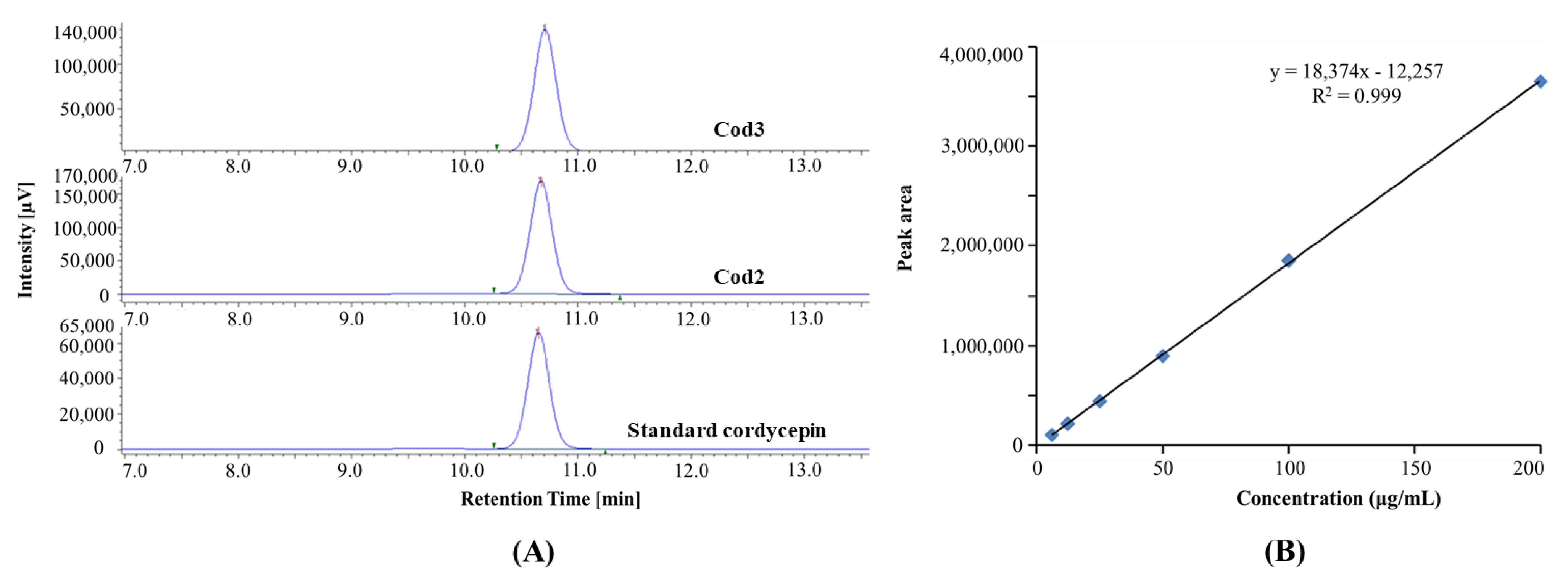
3.1.1. Identification of Cordycepin by HPLC
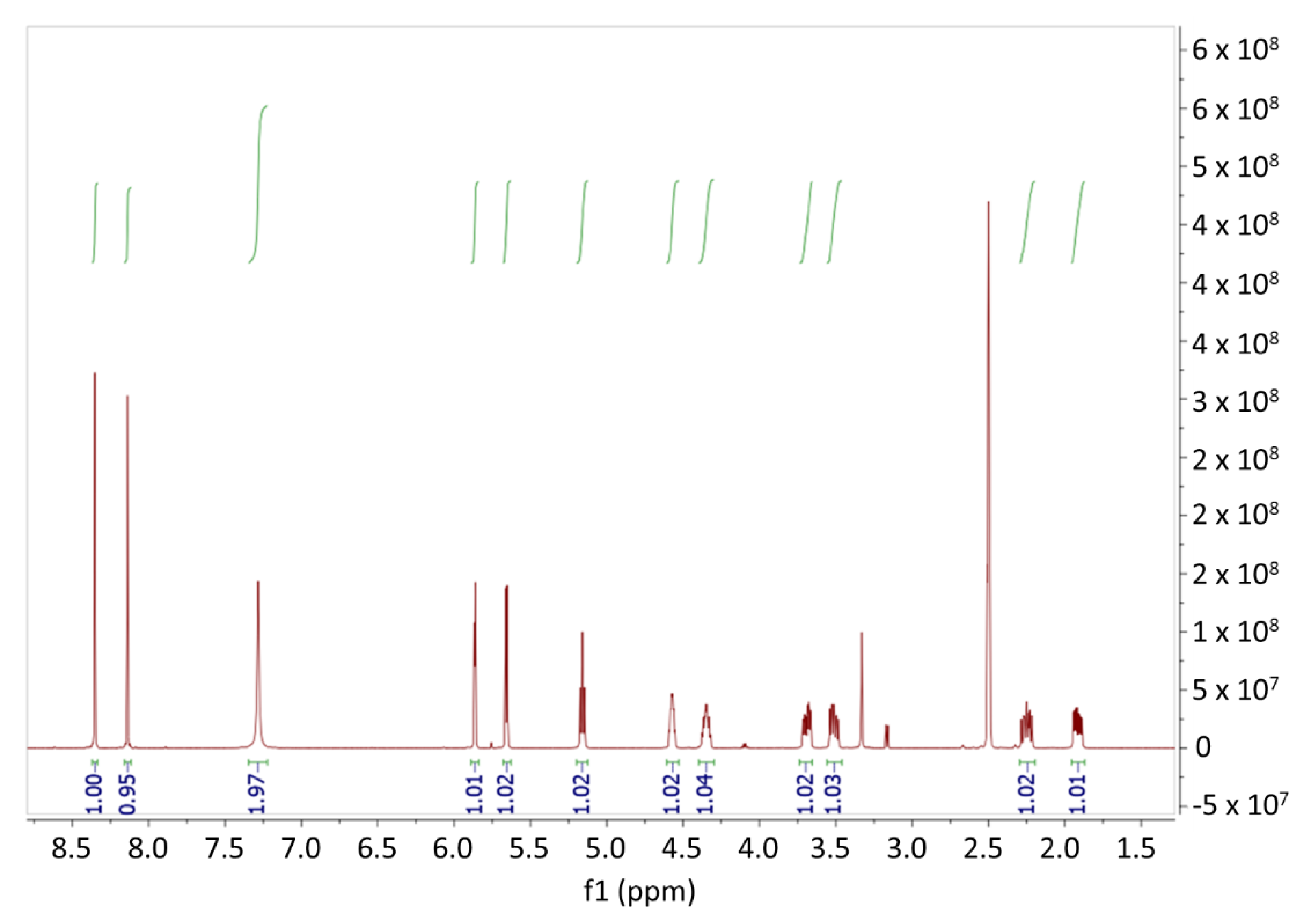
3.1.2. Confirmation of Cordycepin by 1H NMR
3.1.3. Isolation and Purification Efficiency of Cordycepin
3.2. Biological Activities
3.2.1. Antioxidant Activity
3.2.2. α-Glucosidase Inhibitory Activity
3.2.3. Cytotoxic Activity
3.3. Effect of Kombucha Fermentation on Cordycepin Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Paterson, R.R.M. Cordyceps—A traditional Chinese medicine and another fungal therapeutic biofactory? Phytochemistry 2008, 69, 1469–1495. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Gong, Z.; Su, Y.; Lin, J.; Tang, K. Cordyceps fungi: Natural products, pharmacological functions and developmental products. J. Pharm. Pharmacol. 2009, 61, 279–291. [Google Scholar] [CrossRef]
- Zhu, J.-S.; Halpern, G.M.; Jones, K. The scientific rediscovery of an ancient Chinese herbal medicine: Cordyceps sinensis part I. J. Altern. Complement. Med. 1998, 4, 289–303. [Google Scholar] [CrossRef]
- Jędrejko, K.J.; Lazur, J.; Muszyńska, B. Cordyceps militaris: An overview of its chemical constituents in relation to biological activity. Foods 2021, 10, 2634. [Google Scholar] [CrossRef] [PubMed]
- Cohen, N.; Cohen, J.; Asatiani, M.D.; Varshney, V.K.; Yu, H.T.; Yang, Y.C.; Li, Y.H.; Mau, J.L.; Wasser, S.P. Chemical composition and nutritional and medicinal value of fruit bodies and submerged cultured mycelia of culinary-medicinal higher Basidiomycetes mushrooms. Int. J. Med. Mushrooms 2014, 16, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Zhou, X.-H.; Li, H.-H.; Huang, W.-F. Column chromatographic extraction and preparation of cordycepin from Cordyceps militaris waster medium. J. Chromatogr. B 2009, 877, 2135–2141. [Google Scholar] [CrossRef]
- Tuli, H.S.; Kashyap, D.; Sharma, A.K. Cordycepin: A cordyceps metabolite with promising therapeutic potential. In Fungal Metabolites; Merillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2015; pp. 1–22. [Google Scholar]
- Ahn, Y.-J.; Park, S.-J.; Lee, S.-G.; Shin, S.-C.; Choi, D.-H. Cordycepin: Selective growth inhibitor derived from liquid culture of Cordyceps militaris against Clostridium spp. J. Agric. Food Chem. 2000, 48, 2744–2748. [Google Scholar] [CrossRef]
- Luo, Q.Y.; Cao, H.F.; Liu, S.K.; Wu, M.; Li, S.S.; Zhang, Z.Q.; Chen, A.J.; Shen, G.H.; Wu, H.J.; Li, M.L.; et al. Novel liquid fermentation medium of Cordyceps militaris and optimization of hydrothermal reflux extraction of cordycepin. J. Asian Nat. Prod. Res. 2020, 22, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Fortune Business Insights. Available online: https://www.fortunebusinessinsights.com/industry-reports/kombucha-market-100230 (accessed on 14 October 2021).
- Mousavi, S.M.; Hashemi, S.A.; Zarei, M.; Gholami, A.; Lai, C.W.; Chiang, W.H.; Omidifar, N.; Bahrani, S.; Mazraedoost, S. Recent progress in chemical composition, production, and pharmaceutical effects of kombucha beverage: A complementary and alternative medicine. Evid. Based Complement. Alternat. Med. 2020, 2020, 4397543. [Google Scholar] [CrossRef]
- Aloulou, A.; Hamden, K.; Elloumi, D.; Ali, M.B.; Hargafi, K.; Jaouadi, B.; Ayadi, F.; Elfeki, A.; Ammar, E. Hypoglycemic and antilipidemic properties of kombucha tea in alloxan-induced diabetic rats. BMC Complement. Altern. Med. 2012, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Dufresne, C.; Farnworth, E. Tea, kombucha, and health: A review. Food Res. Int. 2000, 33, 409–421. [Google Scholar] [CrossRef]
- Coelho, R.M.D.; de Almeida, A.L.; do Amaral, R.Q.G.; da Mota, R.N.; de Sousa, P.H.M. Kombucha: Review. Int. J. Gastron. Food Sci. 2020, 22, 100272. [Google Scholar] [CrossRef]
- Shahbazi, H.; Gahruie, H.H.; Golmakani, M.-T.; Eskandari, M.H.; Movahedi, M. Effect of medicinal plant type and concentration on physicochemical, antioxidant, antimicrobial, and sensorial properties of kombucha. Food Sci. Nutr. 2018, 6, 2568–2577. [Google Scholar] [CrossRef]
- Quy, T.N.; Xuan, T.D.; Andriana, Y.; Tran, H.-D.; Khanh, T.D.; Teschke, R. Cordycepin isolated from Cordyceps militaris: Its newly discovered herbicidal property and potential plant-based novel alternative to glyphosate. Molecules 2019, 24, 2901. [Google Scholar] [CrossRef] [Green Version]
- Quan, N.V.; Xuan, T.D.; Tran, H.-D.; Thuy, N.T.D.; Trang, L.T.; Huong, C.T.; Andriana, Y.; Tuyen, P.T. Antioxidant, α-amylase and α-glucosidase inhibitory activities and potential constituents of Canarium tramdenum Bark. Molecules 2019, 24, 605. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.G.; Shrestha, B.; Lim, S.Y.; Yoon, D.H.; Chang, W.C.; Shin, D.J.; Han, S.K.; Park, S.M.; Park, J.H.; Park, H.I.; et al. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-κB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur. J. Pharmacol. 2006, 545, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Hyun, S.H.; Sung, G.H.; Choi, H.K. Simple and rapid determination of cordycepin in Cordyceps militaris fruiting bodies by quantitative nuclear magnetic resonance spectroscopy. Anal. Lett. 2014, 47, 1031–1042. [Google Scholar] [CrossRef]
- Masuda, M.; Hatashita, M.; Fujihara, S.; Suzuki, Y.; Sakurai, A. Simple and efficient isolation of cordycepin from culture broth of a Cordyceps militaris mutant. J. Biosci. Bioeng. 2015, 120, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.; Tang, S.; Azi, F.; Hu, W.; Dong, M. Use of kombucha consortium to transform soy whey into a novel functional beverage. J. Funct. Foods 2019, 52, 81–89. [Google Scholar] [CrossRef]
- Quan, N.V.; Tran, H.-D.; Xuan, T.D.; Ahmad, A.; Dat, T.D.; Khanh, T.D.; Teschke, R. Momilactones A and B are α-amylase and α-glucosidase inhibitors. Molecules 2019, 24, 482. [Google Scholar] [CrossRef]
- Anh, L.H.; Quan, N.V.; Lam, V.Q.; Iuchi, Y.; Takami, A.; Teschke, R.; Xuan, T.D. Antioxidant, anti-tyrosinase, anti-α-amylase, and cytotoxic potentials of the invasive weed Andropogon virginicus. Plants 2020, 10, 69. [Google Scholar] [CrossRef]
- Radhi, M.; Ashraf, S.; Lawrence, S.; Tranholm, A.A.; Wellham, P.A.D.; Hafeez, A.; Khamis, A.S.; Thomas, R.; McWilliams, D.; de Moor, C.H. A systematic review of the biological effects of cordycepin. Molecules 2021, 26, 5886. [Google Scholar] [CrossRef]
- Kredich, N.M.; Guarino, A.J. An improved method of isolation and determination of cordycepin. Biochim. Biophys. Acta 1960, 41, 363–365. [Google Scholar] [CrossRef] [Green Version]
- Frederiksen, S.; Malling, H.; Klenow, H. Isolation of 3′-deoxyadenosine (cordycepin) from the liquid medium of Cordyceps militaris (L. ex Fr.) Link. Biochim. Biophys. Acta 1965, 95, 189–193. [Google Scholar] [CrossRef]
- Zhou, X.; Cai, G.; He, Y.; Tong, G. Separation of cordycepin from Cordyceps militaris fermentation supernatant using preparative HPLC and evaluation of its antibacterial activity as an NAD+-dependent DNA ligase inhibitor. Exp. Ther. Med. 2016, 12, 1812–1816. [Google Scholar] [CrossRef]
- Zhang, Z.; Tudi, T.; Liu, Y.; Zhou, S.; Feng, N.; Yang, Y.; Tang, C.; Tang, Q.; Zhang, J. Preparative isolation of cordycepin, N6 -(2-hydroxyethyl)-adenosine and adenosine from Cordyceps militaris by macroporous resin and purification by recycling high-speed counter-current chromatography. J. Chromatogr. B 2016, 1033–1034, 218–225. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, L.; Zhang, Z.; Guan, Y.; Li, B.; Yang, J.; Fan, H.; Yang, G.; Chen, X.; Zhang, J.; et al. Enhanced cordycepin production in caterpillar medicinal mushroom, Cordyceps militaris (Ascomycetes), mutated by a multifunctional plasma mutagenesis system. Int. J. Med. Mushrooms 2020, 22, 1147–1159. [Google Scholar] [CrossRef]
- Zhu, L.; Liang, Y.; Lao, D.; Zhang, T.; Ito, Y. Preparative separation of high-purity cordycepin from Cordyceps militaris (L.) link by high-speed countercurrent chromatography. J. Liq. Chromatogr. Relat. Technol. 2011, 34, 491–499. [Google Scholar]
- Zhang, Y.; Wan, J.; Cao, X. Synthesis of surface molecularly imprinting polymers for cordycepin and its application in separating cordycepin. Process Biochem. 2016, 51, 517–527. [Google Scholar] [CrossRef]
- Qiu, W.; Wu, J.; Choi, J.H.; Hirai, H.; Nishida, H.; Kawagishi, H. Cytotoxic compounds against cancer cells from Bombyx mori inoculated with Cordyceps militaris. Biosci. Biotechnol. Biochem. 2017, 81, 1224–1226. [Google Scholar] [CrossRef] [Green Version]
- Essawet, N.A.; Cvetkovic, D.; Velićanski, A.; Canadanovic-Brunet, J.; Vulić, J.; Maksimović, V.; Sinisa, M.; Sinisa, M. Polyphenols and antioxidant activities of Kombucha beverage enriched with Coffeeberry® extract. Chem. Ind. Chem. Eng. Q. 2015, 21, 399–409. [Google Scholar] [CrossRef]
- Gamboa-Gómez, C.I.; González-Laredo, R.F.; Gallegos-Infante, J.A.; Pérez, M.D.; Moreno-Jiménez, M.R.; Flores-Rueda, A.G.; Rocha-Guzmán, N.E. Antioxidant and angiotensin-converting enzyme inhibitory activity of Eucalyptus camaldulensis and Litsea glaucescens infusions fermented with Kombucha consortium. Food Technol. Biotechnol. 2016, 54, 367–374. [Google Scholar] [CrossRef]
- Xia, X.; Dai, Y.; Wu, H.; Liu, X.; Wang, Y.; Yin, L.; Wang, Z.; Li, X.; Zhou, J. Kombucha fermentation enhances the health-promoting properties of soymilk beverage. J. Funct. Foods 2019, 62, 103549. [Google Scholar] [CrossRef]
- Nagappan, H.; Pee, P.P.; Kee, S.H.Y.; Ow, J.T.; Yan, S.W.; Chew, L.Y.; Kong, K.W. Malaysian brown seaweeds Sargassum siliquosum and Sargassum polycystum: Low density lipoprotein (LDL) oxidation, angiotensin converting enzyme (ACE), α-amylase, and α-glucosidase inhibition activities. Food Res. Int. 2017, 99, 950–958. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Yao, Q.M.; Zhou, J.; Chang, Y.; Li, J.L.; Wang, Y.Z.; Wu, H.P.; Chen, Y.H.; Liu, Y.R.; Huang, X.J.; et al. Synergistic antitumoral efficacy of a novel replicative adenovirus SG611-PDCD5 and daunorubicin in human leukemic cells. Onco. Targets Ther. 2018, 11, 5121–5132. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.P.; Ye, X.L.; Chen, Z.; Zhong, Y.J.; Li, P.M.; Pu, S.C.; Li, X.G. Synthesis and pharmacokinetic evaluation of novel N-acyl-cordycepin derivatives with a normal alkyl chain. Eur. J. Med. Chem. 2009, 44, 665–669. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | DPPH IC50 (mg/mL) | |
|---|---|---|
| BF | AF | |
| Cod1 | 0.88 ± 0.02 a | 9.96 ± 0.06 b |
| Cod23 | na | 4.76 ± 0.03 c |
| BT | 0.06 ± 0.00 b | 11.60 ± 0.05 a |
| Sample | α-Glucosidase Inhibition IC50 (mg/mL) | |
|---|---|---|
| BF | AF | |
| Cod1 | na | 5.66 ± 0.09 |
| Cod23 | na | ne (46.17%) |
| BT | na | ne (22.15%) |
| Sample | Meg-01 Inhibition (%) | HL-60 IC50 (μg/mL) | ||
|---|---|---|---|---|
| BF | AF | BF | AF | |
| Cod1 | 37.89 ± 6.27 a | 14.94 ± 2.14 a | 12.50 ± 1.46 ab | ne |
| Cod23 | 30.60 ± 4.17 a | 8.75 ± 1.90 b | 14.99 ± 0.68 a | 320.49 ± 2.67 |
| BT | ne (3.79%) | 8.64 ± 2.19 b | ne (51.13%) | ne (11.32%) |
| Sample | Cordycepin Content (mg/g) | |
|---|---|---|
| BF | AF | |
| Cod1 | 302.11 ± 2.78 a | 5.27 ± 0.03 b |
| Cod23 | 1203.65 ± 13.43 a | 35.88 ± 0.62 b |
| Study | Material | Stationary Phase | Mobile Phase | Yield (%) |
|---|---|---|---|---|
| Zhu et al. [30] | Whole fungus | Cation-exchange resin and high-speed counter-current chromatography | Two-phase solvents: n-Hexane, n-butanol, methanol, and water (23:80:30:155, v/v/v/v) | (29.97) |
| Zhang et al. [28] | Fruiting bodies | Macroporous resin and high-speed counter-current chromatography | Two-phase solvents: Ethyl acetate, n-butanol, and 1.5% aqueous ammonium hydroxide (1:4:5, v/v/v) | 0.09 |
| Zhang et al. [31] | Mycelia | Molecularly imprinted polymer (MIP) chromatography | Mobile phase: Methanol and acetic acid (9:1, v/v) | 0.23 |
| Ahn et al. [8] | Cultural medium | Amberlite XAD-2 resins (20–50 mesh); silica gel (Merck 230–400 mesh) column; preparatory HPLC (Waters Delta Prep 4000); reverse-phase column (Lichrosorb RP-18); ODS column | Four-phase solvents: water and methanol (for resin extraction); chloroform, methanol, and water (for two-step column chromatography) | 0.04 |
| Qiu et al. [32] | Insect bodies | Silica gel 60N flash column chromatography; C18 cartridges; reverse-phase HPLC (Cosmosil PB) column | Three-phase solvents: Dichloromethane/acetone/methanol; methanol (50% and 100%); 30% methanol | 2.8 × 10−4 (0.09) |
| This study | Fruiting bodies | Normal-phase repeated column chromatography 1st CC: Silica gel 60Å, 60–100 mesh 2nd CC: Silica gel 60Å, 200–400 mesh | Chloroform and methanol (v/v) | 0.04 (5.31) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quan, N.V.; Iuchi, Y.; Anh, L.H.; Hasan, M.; Xuan, T.D. Simple Isolation of Cordycepin from Cordyceps militaris by Dual-Normal Phase Column Chromatography and Its Potential for Making Kombucha Functional Products. Separations 2022, 9, 290. https://doi.org/10.3390/separations9100290
Quan NV, Iuchi Y, Anh LH, Hasan M, Xuan TD. Simple Isolation of Cordycepin from Cordyceps militaris by Dual-Normal Phase Column Chromatography and Its Potential for Making Kombucha Functional Products. Separations. 2022; 9(10):290. https://doi.org/10.3390/separations9100290
Chicago/Turabian StyleQuan, Nguyen Van, Yu Iuchi, La Hoang Anh, Mehedi Hasan, and Tran Dang Xuan. 2022. "Simple Isolation of Cordycepin from Cordyceps militaris by Dual-Normal Phase Column Chromatography and Its Potential for Making Kombucha Functional Products" Separations 9, no. 10: 290. https://doi.org/10.3390/separations9100290
APA StyleQuan, N. V., Iuchi, Y., Anh, L. H., Hasan, M., & Xuan, T. D. (2022). Simple Isolation of Cordycepin from Cordyceps militaris by Dual-Normal Phase Column Chromatography and Its Potential for Making Kombucha Functional Products. Separations, 9(10), 290. https://doi.org/10.3390/separations9100290