
Modulation of Rab GDP-Dissociation Inhibitor Trafficking and Expression by the Transmembrane Protein 59 (TMEM59)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA Extraction and cDNA Synthesis
2.3. cDNA Cloning and Plasmid Constructing
2.4. Transfection
2.5. Subcellular Localization of TMEM59
2.6. Live Cell Imaging of TMEM59
2.7. Western Blotting
3. Results
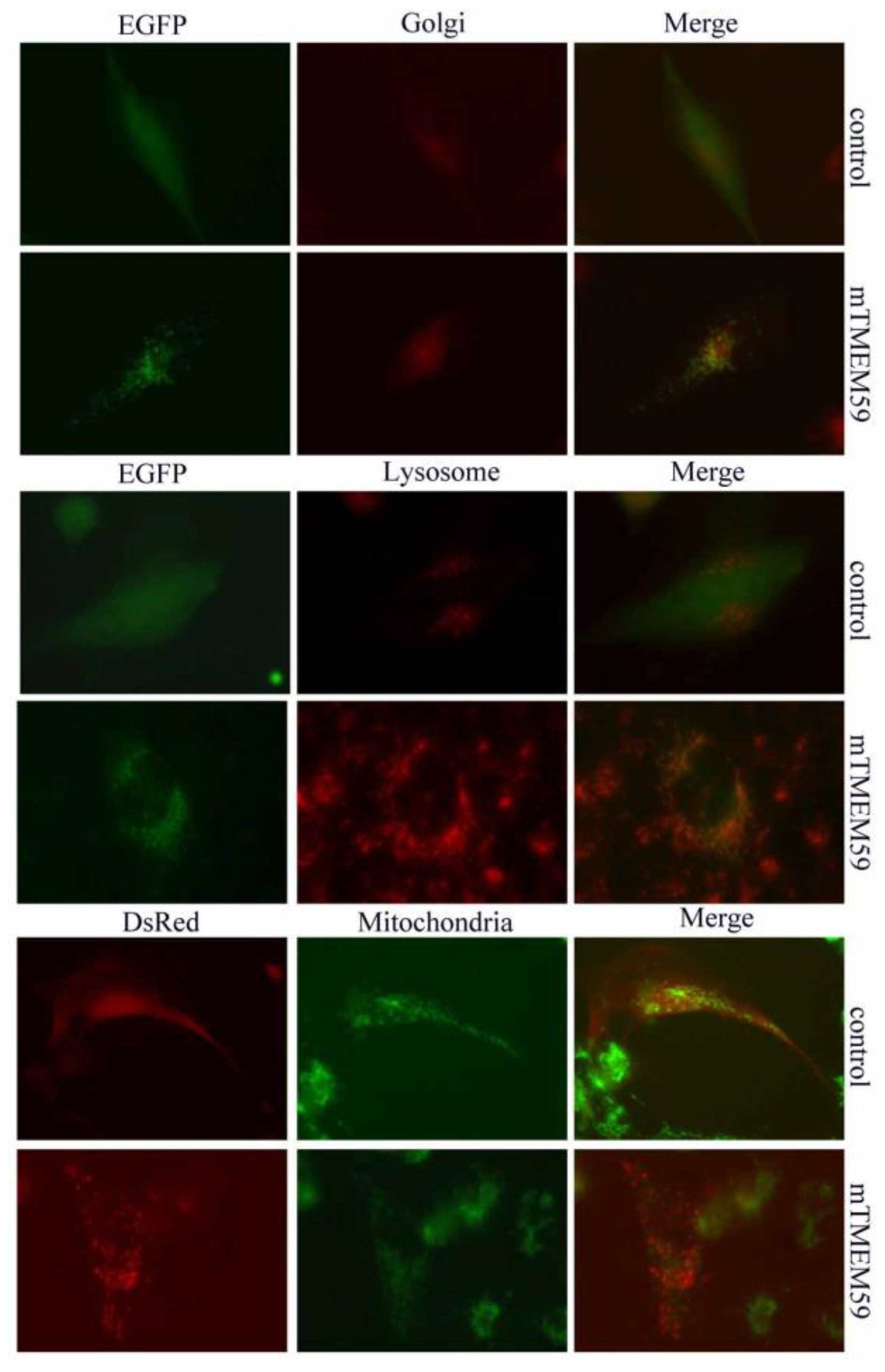
3.1. TMEM59 Localizes to Vesicular Structures
3.2. Movement of TMEM59 in the Cytoplasm
3.3. TMEM59 Localization in Lysosomes and Acidic Vesicles
3.4. TMEM59 Increases APP Level but Has No Effect on BACE2
3.5. Expression and Localization of GDI1 and GDI2 Were Changed by TMEM59
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Chen, Y.; Deng, Y.; Zhang, J.; Yang, L.; Xie, X.; Xu, T. GDI-1 preferably interacts with Rab10 in insulin-stimulated GLUT4 translocation. Biochem. J. 2009, 422, 229–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duce, J.A.; Tsatsanis, A.; Cater, M.A.; James, S.A.; Robb, E.; Wikhe, K.; Leong, S.L.; Perez, K.; Johanssen, T.; Greenough, M.A. Iron-export ferroxidase activity of β-amyloid precursor protein is inhibited by zinc in Alzheimer’s disease. Cell 2010, 142, 857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priller, C.; Bauer, T.; Mitteregger, G.; Krebs, B.; Kretzschmar, H.A.; Herms, J. Synapse formation and function is modulated by the amyloid precursor protein. J. Neurosci. 2006, 26, 7212–7221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, P.R.; O’Connor, K.; Tate, W.P.; Abraham, W.C. Roles of amyloid precursor protein and its fragments in regulating neural activity, plasticity and memory. Prog. Neurobiol. 2003, 70, 1–32. [Google Scholar] [CrossRef]
- Bakulski, K.M.; Dolinoy, D.C.; Sartor, M.A.; Paulson, H.L.; Konen, J.R.; Lieberman, A.P.; Albin, R.L.; Hu, H.; Rozek, L.S. Genome-wide DNA methylation differences between late-onset Alzheimer’s disease and cognitively normal controls in human frontal cortex. J. Alzheimer’s Dis. 2012, 29, 571–588. [Google Scholar] [CrossRef] [Green Version]
- Xi, L.; Zhe, W.; Yili, W.; Jianping, W.; Weihong, S. BACE2 degradation mediated by the macroautophagy–lysosome pathway. Eur. J. Neurosci. 2013, 37, 1970–1977. [Google Scholar]
- Wang, J.; Wang, Q.; Zhou, F.; Li, J.; Li, Q.; Zhou, H.; Li, S.; Ma, S.; Wen, T. The antitumor effect of TAT-DCF1 peptide in glioma cells. Neuropeptides 2018, 71, 21–31. [Google Scholar] [CrossRef]
- Wang, L.; Wang, J.; Wu, Y.; Wu, J.; Pang, S.; Pan, R.; Wen, T. A novel function of dcf1 during the differentiation of neural stem cells in vitro. Cell Mol. Neurobiol. 2008, 28, 887–894. [Google Scholar] [CrossRef]
- Xie, Y.; Li, Q.; Yang, Q.; Yang, M.; Zhang, Z.; Zhu, L.; Yan, H.; Feng, R.; Zhang, S.; Huang, C.; et al. Overexpression of DCF1 inhibits glioma through destruction of mitochondria and activation of apoptosis pathway. Sci. Rep. 2014, 4, 3702. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Chen, Y.; Li, Q.; Wu, L.; Wen, T. Dcf1 regulates neuropeptide expression and maintains energy balance. Neurosci. Lett. 2017, 650, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Qiang, L.; Linhua, G.; Jian, N.; Yu, C.; Yanlu, C.; Zhili, H.; Xu, H.; Tieqiao, W. Dcf1 improves behavior deficit in drosophila and mice caused by optogenetic suppression. J. Cell Biochem. 2017, 118, 4210–4215. [Google Scholar]
- Shi, J.; Li, Q.; Wen, T. Dendritic cell factor 1-knockout results in visual deficit through the GABA system in mouse primary visual cortex. Neurosci. Bull. 2018, 34, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Feng, R.; Li, Y.; Gan, L.; Zhou, F.; Meng, S.; Li, Q.; Wen, T. Degradation of alpha-synuclein by dendritic cell factor 1 delays neurodegeneration and extends lifespan in Drosophila. Neurobiol. Aging 2018, 67, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Romero, E.B.; Letek, M.; Fleischer, A.; Pallauf, K.; Ramón, C.; Pimentel-Muinos, F. TMEM59 defines a novel ATG16L1-binding motif that promotes local activation of LC3. EMBO J. 2013, 32, 566–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boada-Romero, E.; Serramito-Gómez, I.; Sacristán, M.P.; Boone, D.L.; Xavier, R.J.; Pimentel-Muiños, F.X. The T300A Crohn’s disease risk polymorphism impairs function of the WD40 domain of ATG16L1. Nat. Commun. 2016, 7, 11821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Shi, J.; Lin, R.; Wen, T. Dopamine and dopamine receptor D1 associated with decreased social interaction. Behav. Brain Res. 2017, 324, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhou, F.; Wang, D.; Li, J.; Lu, D.; Li, Q.; Zhou, H.; Li, W.; Wang, Q.; Wu, Y.; et al. Interaction of DCF1 with ATP1B1 induces impairment in astrocyte structural plasticity via the P38 signaling pathway. Exp. Neurol. 2018, 302, 214–229. [Google Scholar] [CrossRef]
- Li, X.; Feng, R.; Huang, C.; Wang, H.; Wang, J.; Zhang, Z.; Yan, H.; Wen, T. MicroRNA-351 regulates TMEM 59 (DCF1) expression and mediates neural stem cell morphogenesis. RNA Biol. 2012, 9, 292–301. [Google Scholar] [CrossRef] [Green Version]
- Gerlach, J.P.; Jordens, I.; Tauriello, D.V.F.; van ‘t Land-Kuper, I.; Bugter, J.M.; Noordstra, I.; van der Kooij, J.; Low, T.Y.; Pimentel-Muiños, F.X.; Xanthakis, D.; et al. TMEM59 potentiates Wnt signaling by promoting signalosome formation. Proc. Natl. Acad. Sci. USA 2018, 115, E3996–E4005. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ju, X.; Cheng, Y.; Guo, X.; Wen, T. Identifying Tmem59 related gene regulatory network of mouse neural stem cell from a compendium of expression profiles. BMC Syst. Biol. 2011, 5, 152. [Google Scholar] [CrossRef] [Green Version]
- Abdul-Hay, S.O.; Sahara, T.; McBride, M.; Kang, D.; Leissring, M.A. Identification of BACE2 as an avid ß-amyloid-degrading protease. Mol. Neurodegener. 2012, 7, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bächner, D.; Sedlacek, Z.; Korn, B.; Hameister, H.; Poustka, A. Expression patterns of two human genes coding for different rab GDP-dissociation inhibitors (GDIs), extremely conserved proteins involved in cellular transport. Hum. Mol. Genet. 1995, 4, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Shisheva, A.; Buxton, J.; Czech, M. Differential intracellular localizations of GDP dissociation inhibitor isoforms. Insulin-dependent redistribution of GDP dissociation inhibitor-2 in 3T3-L1 adipocytes. J. Biol. Chem. 1994, 269, 23865–23868. [Google Scholar] [CrossRef]
- Shisheva, A.; Doxsey, S.J.; Buxton, J.M.; Czech, M.P. Pericentriolar targeting of GDP-dissociation inhibitor isoform 2. Eur. J. Cell Biol. 1995, 68, 143–158. [Google Scholar] [PubMed]
- Sedlacek, Z.; Konecki, D.S.; Korn, B.; Klauck, S.M.; Poustka, A. Evolutionary conservation and genomic organization of XAP-4, an Xq28 located gene coding for a human RAB GDP-dissociation inhibitor (GDI). Mamm. Genome 1994, 5, 633. [Google Scholar] [CrossRef] [PubMed]
- Beranger, F.; Cadwallader, K.; Porfiri, E.; Powers, S.; Evans, T.; de Gunzburg, J.; Hancock, J. Determination of structural requirements for the interaction of Rab6 with RabGDI and Rab geranylgeranyltransferase. J. Biol. Chem. 1994, 269, 13637–13643. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, R.; Luo, G.; Guo, J.; Wang, Y.; Sun, Y.; Zheng, L.; Wen, T. DCF1 subcellular localization and its function in mitochondria. Biochimie 2018, 144, 50–55. [Google Scholar] [CrossRef]
- Raffaniello, R.; Fedorova, D.; Ip, D.; Rafiq, S. Hsp90 Co-localizes with Rab-GDI-1 and regulates agonist-induced amylase release in AR42J cells. Cell Physiol. Biochem. 2009, 24, 369–378. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Sequence (5′–3′) | Plamsid |
|---|---|---|
| hTMEM59 | 5′-CGGAATTCATGGCGGCGCCGAAGGGGAG-3′ | N2 |
| 5′-CGGGATCCGTAAAATTTCAGAATGAGCA-3 | ||
| 5′-GATCTCGAGACCATGGCGGCGCCGAAGGGGAG-3′ | 3.1(−)A | |
| 5′-CAGGAATTCTAATTTCAGAATGAGCAAG-3′ | ||
| 5′-CGGAATTCATGGCGGCGCCGAAGGGGAG-3′ | N1 | |
| 5′-CGGGATCCAGTAAAATTTCAGAATGAGCA-3′ | ||
| mTMEM59 | 5′-GATCTCGAGACCATGGCGGCGCCAAAGGGGAAG-3′ | N2 |
| 5′-CAGGAATTCGATTTCTGAGTGAGCAAGGTTC-3′ | ||
| 5′-GATCTCGAGACCATGGCGGCGCCAAAGGGGAAG-3′ | 3.1(−)A | |
| 5′-CAGGAATTCTGATTTCTGAGTGAGCAAGGTTC-3′ | ||
| APP | 5′-GACAAGCTTATGCTGCCCGGTTTGG-3′ | N2 |
| 5′-GATCCCGGGGTTCTGCATCTGCTCAAAGAACT-3′ | ||
| BACE2 | 5′-GATCTCGAGATGGGCGCACTGGC-3′ | N2 |
| 5′-GTAGGATCCGTTTCCAGCGATGTCTGACC-3′ | ||
| GDI1 | 5′-GATAAGCTTATGGACGAGGAATACGAT-3′ | N2 |
| 5′-GTAGGATCCGCTGCTCAGCTTCTCCAAAG-3′ | ||
| GDI2 | 5′-GATCTCGAGATGAATGAGGAGTACGAC-3′ | N2 |
| 5′-GTAGGATCCGGTCTTCCCCATAGATGTCA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Wen, T. Modulation of Rab GDP-Dissociation Inhibitor Trafficking and Expression by the Transmembrane Protein 59 (TMEM59). Separations 2022, 9, 341. https://doi.org/10.3390/separations9110341
Wang H, Wen T. Modulation of Rab GDP-Dissociation Inhibitor Trafficking and Expression by the Transmembrane Protein 59 (TMEM59). Separations. 2022; 9(11):341. https://doi.org/10.3390/separations9110341
Chicago/Turabian StyleWang, Haifeng, and Tieqiao Wen. 2022. "Modulation of Rab GDP-Dissociation Inhibitor Trafficking and Expression by the Transmembrane Protein 59 (TMEM59)" Separations 9, no. 11: 341. https://doi.org/10.3390/separations9110341
APA StyleWang, H., & Wen, T. (2022). Modulation of Rab GDP-Dissociation Inhibitor Trafficking and Expression by the Transmembrane Protein 59 (TMEM59). Separations, 9(11), 341. https://doi.org/10.3390/separations9110341