
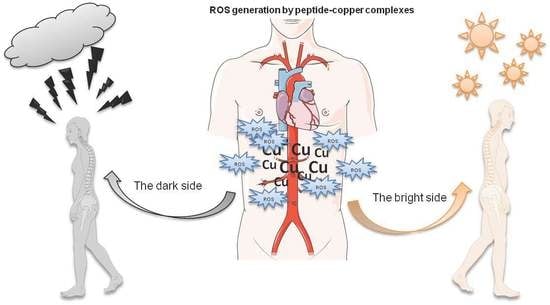
The Bright and Dark Sides of Reactive Oxygen Species Generated by Copper–Peptide Complexes


Abstract
:
{kind=link}
{kind=link}
{kind=link}
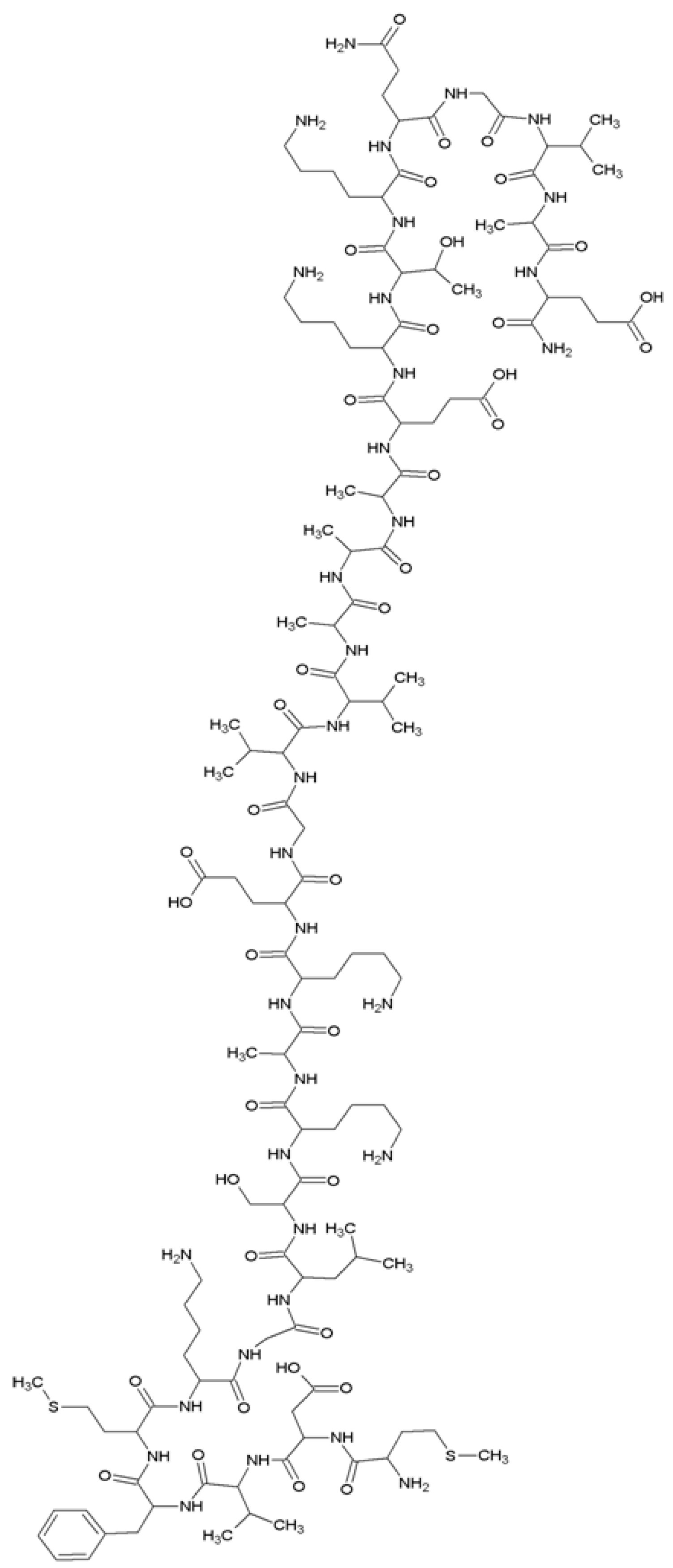{kind=link}
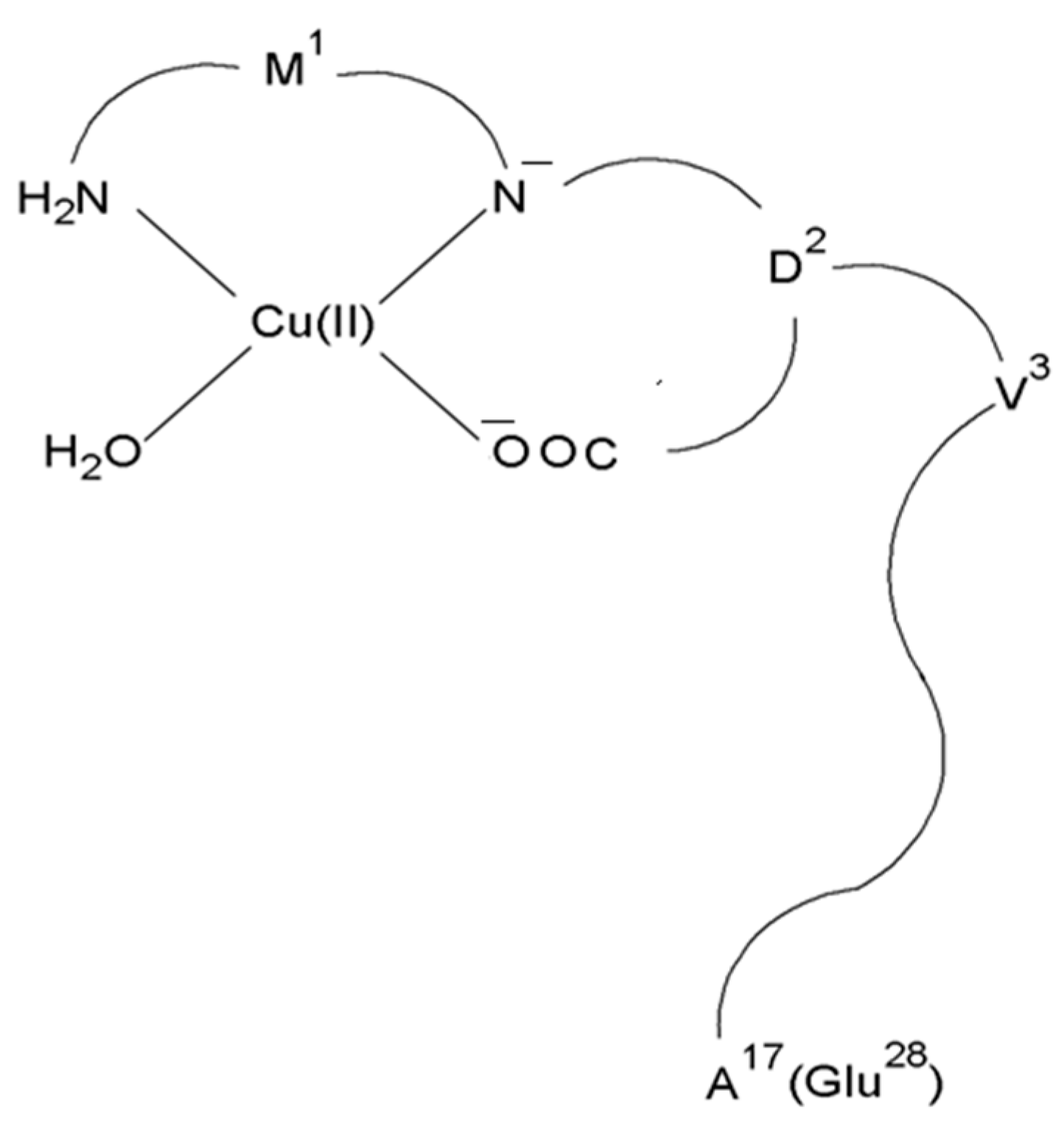{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Dark Side of Copper–Peptide Complexes
2.1. Neurodegenerative Disorders
2.1.1. Tau (T) Protein
2.1.2. α-Synuclein (αSyn) Protein
2.1.3. Presenilin 1 (Prs1) Protein
2.1.4. β-Amyloid (Aβ) Protein
2.1.5. Cellular Prion (PrPC) Protein
2.2. Cancer
2.2.1. Major Outer Membrane (FomA) Protein
2.2.2. Adhesion (FadA) Protein
2.3. Respiratory System Diseases
Spike (S) Protein
3. Bright Side of Copper–Peptide Complexes
3.1. Cancer Treatment
3.1.1. Copper Complexes with Amino Acids and Peptide
3.1.2. Copper Complexes with Peptide and Diimines
3.1.3. Copper Complexes with Peptide and Imidazole
3.1.4. Copper Complexes with Phosphines
3.2. Neurodegenerative Diseases
3.3. Antiviral Properties
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Ac | acetyl group |
| Dmp | neocuproine; 2,9-dimethyl-1,10-phenatroline |
| SarGly | sarcosyl-glycine |
| DSBs | double strand breaks |
| DNA | deoxyribonucleic acid |
| Phen | 1,10-phenanthroline |
| ROS | reactive oxygen species |
| Prs1 | Presenilin 1 |
| αSyn | alpha-synuclein |
| MDA | malondialdehyde |
| 4-HNE | 4-hydroxynonenal |
| AD | Alzheimer’s disease |
| MCO | metal-ion-catalyzed oxidation |
| FAD | Familial Alzheimer’s disease |
| APP protein | amyloid precursor protein |
| Fn | Fusobacterium nucleatum |
| AC | amino acids |
| DSBs | double strand breaks |
| GGH | Gly-Gly-His |
| MPP | FrFKFrFK-CONH2 (Phe-r-Phe-Lys-Phe-r-Phe-Lys-CONH2, where r = D-arginine) |
| HDPs | host defense peptides |
| MDA-MB-231 | human metastatic breast adenocarcinoma |
| MCF-7 | human metastatic breast adenocarcinoma |
| MCF-10A | human normal breast cells |
| A549 | human lung epithelial carcinoma |
| MRC-5 | human lung epithelial cells |
| AA | ascorbic acid |
| TBZ | 2-(4′-thiazolyl)benzimidazole) |
| DOPA | 3,4-dihydroxyphenylalanine |
| HSV | herpes simplex virus |
| FCV | feline calicivirus |
References
- Harris, E.D. Cellular copper transport and metabolism. Annu. Rev. Nutr. 2000, 20, 291–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Guo, Z.J. Copper in medicine: Homeostasis, chelation therapy and antitumor drug design. Curr. Med. Chem. 2006, 13, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Duncan, C.; White, A.R. Copper complexes as therapeutic agents. Metallomics 2012, 4, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Santini, C.; Pellei, M.; Gandin, V.; Porchia, M.; Tisato, F.; Marzano, C. Advances in copper complexes as anticancer agents. Chem. Rev. 2014, 114, 815–862. [Google Scholar] [CrossRef] [PubMed]
- Serment-Guerrero, J.; Bravo-Gomez, M.E.; Lara-Rivera, E.; Ruiz-Azuara, L. Genotoxic assessment of the copper chelated compounds Casiopeinas: Clues about their mechanisms of action. J. Inorg. Biochem. 2017, 166, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Finkel, T. Radical medicine: Treating ageing to cure disease. Nat. Rev. Mol. Cell Biol. 2005, 6, 971–976. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; He, Z. ROS-responsive drug delivery systems for biomedical. Asian J. Pharm. Sci. 2018, 13, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, N.D.; Diana Viña, D.; Leite, C.M.; Mendes; Batista, A.A.; Ellena, J.; Costa-Filho, A.J.; Facchin, G. Synthesis and structural characterization of a series of ternary copper(II)-L-dipeptide-neocuproine complexes. Study of their cytotoxicity against cancer cells including MDA-MB-231, triple negative breast cancer cells. J. Inorg. Biochem. 2020, 203, 110930–110941. [Google Scholar] [CrossRef] [PubMed]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharm. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Karogodina, T.Y.; Sergeeva, S.V.; Stass, D.V. Stability and reactivity of free radicals: A physicochemical perspective with biological implications. Hemoglobin 2011, 35, 262–275. [Google Scholar] [CrossRef]
- Lushchak, V.I. Free radicals, reactive oxygen species, oxidative stress and its classification. Chem. Biol. Interact. 2014, 224, 164–175. [Google Scholar] [CrossRef] [PubMed]
- Studer, A.; Curran, D.P. Catalysis of radical reactions: A radical chemistry perspective. Angew. Chem. Int. Ed. 2016, 55, 58–102. [Google Scholar] [CrossRef] [PubMed]
- Hayyan, M.; Hashim, M.A.; AlNashef, I.M. Superoxide ion: Generation and chemical implications. Chem. Rev. 2016, 116, 3029–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clausen, M.H.; Sassetti, E.; Clausen, M.H.; Laraia, L. Small-Molecule Inhibitors of Reactive Oxygen Species Production. J. Med. Chem. 2021, 64, 5252–5275. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 2015; Volume 5, p. 145. [Google Scholar]
- Thannickal, V.J.; Fanburg, B.L. Reactive oxygen species in cell signaling. Am. J. Physiol. Cell. Mol. Physiol. 2000, 279, 1005–1028. [Google Scholar] [CrossRef] [Green Version]
- Curtin, J.F.; Donovan, M.; Cotter, T.G. Regulation and measurement of oxidative stress in apoptosis. J. Immunol. Methods 2002, 265, 49–72. [Google Scholar] [CrossRef] [Green Version]
- Edge, R. Radiolytic and photolytic production of free radicals and reactive oxygen species: Interactions with antioxidants and biomolecules. In Applied Photochemistry; Evans, R.C., Douglas, P., Burrows, H.D., Eds.; Springer Science: Dordrecht, The Netherlands, 2013; pp. 305–330. [Google Scholar]
- Swartz, H.M.; Mason, R.P.; Hogg, N.; Kalyanaraman, B.; Sarna, T.; Plonka, P.M.; Zareb, M.; Gutierrez, P.L.; Berliner, L.J. Biomedical EPR, Part A: Free Radicals, Metals, Medicine, and Physiology; Springer: Boston, MA, USA, 2005; Volume 23, pp. 25–74. [Google Scholar] [CrossRef]
- Halliwell, B.; Adhikary, A.; Dingfelder, M.; Dizdaroglu, M. Hydroxyl radical is a significant player in oxidative DNA damage in vivo. Chem. Soc. Rev. 2021, 50, 8355–8360. [Google Scholar] [CrossRef]
- Edge, R.; Truscott, T.G. The Reactive Oxygen Species Singlet Oxygen, Hydroxyadicals, and the Superoxide Radical Anion—Examples of Their Roles in Biology and Medicine. Oxygen 2021, 1, 9. [Google Scholar] [CrossRef]
- Ohtsuki, A.; Lei, L.; Tanishima, M.; Goto, A.; Kaji, H. Photocontrolled organocatalyzed living radical polymerization feasible over a wide range of wavelengths. J. Am. Chem. Soc. 2015, 137, 5610–5617. [Google Scholar] [CrossRef]
- Shanmugam, S.; Xu, J.; Boyer, C. Exploiting metalloporphyrins for selective living radical polymerization tunable over visible wavelengths. J. Am. Chem. Soc. 2015, 137, 9174–9185. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Q.; Gao, F.; Zhang, X.Z. Initiator-loaded gold nanocages as a light-induced free-radical generator for cancer therapy. Angew. Chem. Int. Ed. 2017, 56, 9029–9033. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, H.; Kleiman, M.; Esser-Kahn, A.P. Mechanically controlled radical polymerization initiated by ultrasound. Nat. Chem. 2017, 9, 135. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, K.; Bu, W.; Ni, D.; Liu, Y.; Feng, J.; Shi, J. Marriage of scintillator and semiconductor for synchronous radiotherapy and deep photodynamic therapy with diminished oxygen dependence. Angew. Chem. Int. Ed. 2015, 54, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, N.; Dong, X.; Wang, C.; Du, Z.; Mei, L.; Yong, Y.; Huang, C.; Li, Y.; Gu, Z.; et al. Graphdiyne nanoparticles with high free radical scavenging activity for radiation protection. ACS Appl. Mater. Interfaces 2019, 11, 2579–2590. [Google Scholar] [CrossRef] [PubMed]
- Buck, A.T.; Paletta, J.T.; Khindurangala, S.A.; Beck, C.L.; Winter, A.H. Anoncovalently reversible paramagnetic switch in water. J. Am. Chem. Soc. 2013, 135, 10594–10597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, B.; Li, W.-Y.; Chang, Y.; Yuan, B.; Wu, Y.; Zhang, M.-T.; Xu, J.-F.; Li, J.; Zhang, X. A supramolecular radical dimer: High-efficiency NIR-II photothermal conversion and therapy. Angew. Chem. 2019, 131, 15672–15677. [Google Scholar] [CrossRef]
- Magennis, E.P.; Fernandez-Trillo, F.; Sui, C.; Spain, S.P.; Bradshaw, D.J.; Churchley, D.; Mantovani1, G.; Winzer, K.; Alexander, C. Bacteria-instructed synthesis of polymers for self-selective microbial binding and labelling. Nat. Mater. 2014, 13, 748–755. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.; Lunn, D.J.; Pusuluri, A.; Yoo, J.I.; O′Malley, M.A.; Mitragotri, S.; Soh, H.T.; Hawker, C.J. Engineering live cell surfaces with functional polymers via cytocompatible controlled radical polymerization. Nat. Chem. 2017, 9, 537. [Google Scholar] [CrossRef]
- Geng, J.; Li, W.; Zhang, Y.; Thottappillil, N.; Clavadetscher, J.; Lilienkampf, A.; Bradley, M. Radical polymerization inside living cells. Nat. Chem. 2019, 11, 578–586. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, J.W. Free radical catalysis by galactose oxidase. Chem. Rev. 2003, 103, 2347–2364. [Google Scholar] [CrossRef]
- Orlando, J.J.; Tyndall, G.S.; Wallington, T.J. The atmospheric chemistry of alkoxy radicals. Chem. Rev. 2003, 103, 4657–4690. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Harmful and Beneficial Role of ROS. Oxid Med. Cell Longev. 2016, 2016, 7909186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Auten, R.L.; Davis, J.M. Oxygen toxicity and reactive oxygen species: The devil is in the details. Pediatr Res. 2009, 66, 121–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Ren, Z.; Zhang, J.; Chuang, C.C.; Kandaswamy, E.; Zhou, T.; Zuo, L. Role of ROS and Nutritional Antioxidants in Human Diseases. Front. Physiol. 2018, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Barrera, G.; Pizzimenti, S.; Daga, M.; Dianzani, C.; Arcaro, A.; Cetrangolo, G.P.; Giordano, G.; Cucci, M.A.; Graf, M.; Gentile, F. Lipid Peroxidation-Derived Aldehydes, 4-Hydroxynonenal and Malondialdehyde in Aging-Related Disorders. Antioxidants 2018, 7, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef]
- Saieva, C.; Peluso, M.; Palli, D.; Cellai, F.; Ceroti, M.; Selvi, V.; Bendinelli, B.; Assedi, M.; Munnia, A.; Masala, G. Dietary and lifestyle determinants of malondialdehyde DNA adducts in a representative sample of the Florence City population. Mutagenesis 2016, 31, 475–480. [Google Scholar] [CrossRef]
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; TsouhFokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front Physiol. 2020, 11, 694. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Detection, identification, and quantification of oxidative protein modifications. J. Biol. Chem. 2019, 294, 19683–19708. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chande, N.S. Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 19, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sai, D.L.; Lee, J.; Nguyen, D.L.; Kim, Y.-P. Tailoring photosensitive ROS for advanced photodynamic therapy. Exp. Mol. Med. 2021, 52, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Escudero, A. Photodynamic therapy: Photosensitizers and nanostructures. Mater. Chem. Front. 2021, 5, 3788–3812. [Google Scholar] [CrossRef]
- Milkovic, L. Overview of ROS as Cell Function Regulators and Their Implications in Therapy Concepts. Cells 2019, 8, 793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forte, M. Targeting Nitric Oxide with Natural Derived Compounds as a Therapeutic Strategy in Vascular Diseases. Oxid. Med. Cell Longev. 2016, 1155, 7364138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço, F.C. Nitric Oxide Pathways in Neurovascular Coupling Under Normal and Stress Conditions in the Brain: Strategies to Rescue Aberrant Coupling and Improve Cerebral Blood Flow. Front. Physiol. 2021, 12, 729201. [Google Scholar] [CrossRef] [PubMed]
- Lukács, M.; Szunyog, G.; Grenács, Á.; Lihi, N.; Kállay, C.; Di Natale, G.; Campagna, T.; Lanza, V.; Tabbi, G.; Pappalardo, G.; et al. Copper(II) Coordination Abilities of the Tau Protein’s N-Terminus Peptide Fragments: A Combined Potentiometric, Spectroscopic and Mass Spectrometric Study. Chempluschem 2019, 84, 1697–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muralidar, S.; Ambi, S.V.; Sekaran, S.; Thirumalai, D.; Palaniappan, B. Role of tau protein in Alzheimer’s disease: The prime pathological player. Int. J. Biol. Macromol. 2020, 163, 1599–1617. [Google Scholar] [CrossRef]
- Leal, S.S.; Botelho, H.M.; Gomes, C.M. Metal ions as modulators of protein conformation and misfolding in neurodegeneration. Coord. Chem. Rev. 2012, 256, 2253–2270. [Google Scholar] [CrossRef]
- Sun, X.-Y.; Wei, Y.-P.; Xiong, Y.; Wang, X.-C.; Xie, A.-J.; Wang, X.-L.; Yang, Y.; Wang, Q.; Lu, Y.-M.; Liu, R.; et al. Synaptic released zinc promotes tau hyperphosphorylation by inhibition of protein phosphatase 2A (PP2A). J. Biol. Chem. 2012, 287, 11174–11182. [Google Scholar] [CrossRef] [Green Version]
- Petersen, J.D.; Kaech, S.; Banker, G. Selective microtubule-based transport of dendritic membrane proteins arises in concert with axon specification. J. Neurosci. 2014, 34, 4135–4147. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; Manczak, M.; Yin, X.; Wang, R.; Reddy, P.H. Hippocampal phosphorylated tau induced cognitive decline, dendritic spine loss and mitochondrial abnormalities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dash, S. Tau pathology in Alzheimer’s disease and associated hypotheses. Life Res. 2019, 17, 115–124. [Google Scholar] [CrossRef]
- Wang, Y.; Mandelkow, E. Tau in physiology and pathology. Nat. Rev. Neurosci. 2016, 17, 5–21. [Google Scholar] [CrossRef]
- Ahmadi, S.; Zhu, S.; Sharma, R.; Wu, B.; Soong, R.; Dutta Majumdar, R.; Wilson, D.J.; Simpson, A.J.; Kraatz, H.B. Aggregation of Microtubule Binding Repeats of Tau Protein is Promoted by Cu2+. ACS Omega 2019, 4, 5356–5366. [Google Scholar] [CrossRef] [Green Version]
- Golec, C.; Mortensen, S.; Anwar, S.; Martic-Milne, S. Dual roles of tau R peptides on Cu(II)/(I)-mediated reactive oxygen species formation. J. Biol. Inorg. Chem. 2021, 26, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Burré, J.; Sharma, M.; Tsetsenis, T.; Buchman, V.; Etherton, M.R.; Südhof, T.C. Alpha-synuclein promotes SNARE-complex assembly in vivo and in vitro. Science 2010, 329, 1663–1667. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Uéda, K.; Chan, P. Alpha-synuclein and dopamine metabolism. Mol. Neurobiol. 2005, 31, 243–254. [Google Scholar] [CrossRef]
- Spillantini, M.G.; Schmidt, M.L.; Lee, V.M.; Trojanowski, J.Q.; Jakes, R.; Goedert, M. α-Synuclein in Lewy bodies. Nature 1997, 388, 839–840. [Google Scholar] [CrossRef]
- Lee, H.-J.; Bae, E.-J.; Lee, S.-J. Extracellular α-synuclein—A novel and crucial factor in Lewy body diseases. Nat. Rev. Neurol. 2014, 10, 92–98. [Google Scholar] [CrossRef]
- Binolfi, A.; Quintanar, L.; Bertoncini, C.W.; Griesinger, C.; Fernández, C.O. Bioinorganic chemistry of copper coordination to alpha-synuclein: Relevance to Parkinson’s disease. Coord. Chem. Rev. 2012, 256, 2188–2201. [Google Scholar] [CrossRef] [Green Version]
- Dell’Acqua, S.; Pirota, V.; Anzani, C.; Rocco, M.M.; Nicolis, S.; Valensin, D.; Monzani, E.; Casella, L. Reactivity of copper-α-synuclein peptide complexes relevant to Parkinson’s disease. Metallomics 2015, 7, 1091–1102. [Google Scholar] [CrossRef] [PubMed]
- Kowalik-Jankowska, T.; Rajewska, A.; Wiśniewska, K.; Grzonka, Z.; Jezierska, J. Coordination abilities of N-terminal fragments of alpha-synuclein towards copper(II) ions: A combined potentiometric and spectroscopic study. J. Inorg. Biochem. 2005, 99, 2282–2291. [Google Scholar] [CrossRef] [PubMed]
- Kowalik-Jankowska, T.; Rajewska, A.; Jankowska, E.; Grzonka, Z. Products of Cu(II)-catalyzed oxidation of alpha-synuclein fragments containing M1-D2 and H50 residues in the presence of hydrogen peroxide. Dalton Trans. 2008, 6, 832–838. [Google Scholar] [CrossRef]
- Kowalik-Jankowska, T.; Rajewska, A.; Jankowska, E.; Wiśniewska, K.; Grzonka, Z. Products of Cu(II)-catalyzed oxidation of the N-terminal fragments of α-synuclein in the presence of hydrogen peroxide. J. Inorg. Biochem. 2006, 100, 1623–1631. [Google Scholar] [CrossRef]
- Somavarapu, A.K.; Kepp, K.P. Loss of stability and hydrophobicity of presenilin 1 mutations causing Alzheimer’s disease. J. Neurochem. 2016, 137, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.; Cheung, K.H.; Foskett, J.K. Enhanced ROS generation mediated by Alzheimer’s disease presenilin regulation of InsP3R Ca2+ signaling. Antioxid. Redox. Signal. 2011, 14, 1225–1235. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Park, H.; Maharana, C.; Gwon, A.R.; Park, J.; Baek, S.H.; Bae, H.G.; Cho, Y.; Kim, H.K.; Sul, J.H.; et al. Alzheimer’s disease-causing presenilin-1 mutations have deleterious effects on mitochondrial function. Theranostics 2021, 11, 8855–8873. [Google Scholar] [CrossRef]
- Montes, S.; Rivera-Mancia, S.; Diaz-Ruiz, A.; Tristan-Lopez, L.; Rios, C. Copper and Copper Proteins in Parkinson’s Disease. Oxid. Med. Cell. Longev. 2014, 2014, 147251. [Google Scholar] [CrossRef] [Green Version]
- Lesiów, M.K.; Krupa, K. The impact of the histidyl residue position on the formation and stability of Cu(II) complexes and their ability of ROS generation. New J. Chem. 2021, 45, 8543–8556. [Google Scholar] [CrossRef]
- Kowalik-Jankowska, T.; Ruta, M.; Wiśniewska, K.; Łankiewicz, L.; Dyba, M. Products of Cu(II)-catalyzed oxidation in the presence of hydrogen peroxide of the 1–10, 1–16 fragments of human and mouse β-amyloid peptide. J. Inorg. Biochem. 2004, 98, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Swomley, A.M.; Sultana, R. Amyloid β-peptide (1-42)-induced oxidative stress in Alzheimer disease: Importance in disease pathogenesis and progression. Antiox. Redox Signal. 2013, 19, 823–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalik-Jankowska, T.; Ruta, M.; Wiśniewska, K.; Łankiewicz, L. Coordination abilities of the 1-16 and 1-28 fragments of β-amyloid peptide towards copper(II) ions: A combined potentiometric and spectroscopic study. J. Inorg. Biochem. 2003, 95, 270–282. [Google Scholar] [CrossRef]
- WilochM, Z.; Wawrzyniak, U.E.; Ufnalska, I.; Bonna, A.; Bal, W.; Drew, S.C.; Wróblewski, W. Tuning the Redox Properties of Copper(II) Complexes with Amyloid-β Peptides. J. Electrochem. Soc. 2016, 163, 196–199. [Google Scholar] [CrossRef]
- Jiang, D.; Men, L.; Wang, J.; Zhang, Y.; Chickenyen, S.; Wang, Y.; Zhou, F. Redox reactions of copper complexes formed with different beta-amyloid peptides and their neuropathalogical relevance. Biochemistry 2007, 46, 9270–9282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo-Morantes, C.Y.; Wille, H. The structure of human prions: From biology to structural models-considerations and pitfalls. Viruses 2014, 6, 3875–3892. [Google Scholar] [CrossRef] [PubMed]
- Wulf, M.A.; Senatore, A.; Aguzzi, A. The biological function of the cellular prion protein: An update. BMC Biol. 2017, 15, 34. [Google Scholar] [CrossRef] [Green Version]
- Linden, R. The Biological Function of the Prion Protein: A Cell Surface Scaffold of Signaling Modules. Front. Mol. Neurosci. 2017, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Zidar, J.; Pirc, E.T.; Hodoscek, M.; Bukovec, P. Copper(II) ion binding to cellular prion protein. J. Chem. Inf. Model 2008, 48, 283–287. [Google Scholar] [CrossRef]
- Bonomo, R.P.; Cucinotta, V.; Giuffrida, A.; Impellizzeri, G.; Magri, A.; Pappalardo, G.; Rizzarelli, E.; Santoro, A.M.; Tabbi, G.; Vagliasindi, L.I. A re-investigation of copper coordination in the octa-repeats region of the prion protein. Dalton Trans. 2005, 1, 150–158. [Google Scholar] [CrossRef]
- Srikanth, R.; Wilson, J.; Burns, C.S.; Vachet, R.W. Identification of the copper(II) coordinating residues in the prion protein by metal-catalyzed oxidation mass spectrometry: Evidence for multiple isomers at low copper(II) loadings. Biochemistry 2008, 47, 9258–9268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pocanschi, C.L.; Apell, H.-J.; Puntervoll, P.; Høgh, B.; Jensen, H.B.; Welte, W.; Kleinschmidt, J.-H. The major outer membrane protein of Fusobacterium nucleatum (FomA) folds and inserts into lipid bilayers via parallel folding pathways. J. Mol. Biol. 2006, 20, 548–561. [Google Scholar] [CrossRef] [Green Version]
- Nobbs, A.H.; Jenkinson, H.F.; Jakubovics, N.S. Stick to your gums: Mechanisms of oral microbial adherence. J. Dent. Res. 2011, 90, 1271–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.F.; Shi, W.; Zhu, W.; Smith, J.W.; Hsieh, S.L.; Gallo, R.L.; Huang, C.M. Vaccination targeting surface FomA of Fusobacterium nucleatum against bacterial co-aggregation: Implication for treatment of periodontal infection and halitosis. Vaccine 2010, 28, 3496–3505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Cai, S.; Ma, Y. Association between Fusobacterium nucleatum and colorectal cancer: Progress and future directions. J. Cancer 2018, 9, 1652–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hordyjewska, A.; Popiołek, Ł.; Kocot, J. The many “faces” of copper in medicine and treatment. Biometals 2014, 27, 611–621. [Google Scholar] [CrossRef] [Green Version]
- Lesiów, M.K.; Pietrzyk, P.; Kyzioł, A.; Komarnicka, U.K. Cu(II) complexes with fomA protein fragments of Fusobacterium nucleatum increase oxidative stress and malondialdehyde level. Chem. Res. Toxicol. 2019, 32, 2227–2237. [Google Scholar] [CrossRef]
- Lesiów, M.K.; Pietrzyk, P.; Bieńko, A.; Kowalik-Jankowska, T. Stability of Cu(II) complexes with FomA protein fragments containing two His residues in the peptide chain. Metallomics 2019, 11, 1518–1531. [Google Scholar] [CrossRef]
- Lesiów, M.K.; Komarnicka, U.K.; Kyzioł, A.; Bieńko, A.; Pietrzyk, P. ROS-mediated lipid peroxidation as a result of Cu(II) interaction with FomA protein fragments of F. nucleatum: Relevance to colorectal carcinogenesis. Metallomics 2019, 11, 2066–2077. [Google Scholar] [CrossRef]
- Témoin, S.; Wu, K.L.; Wu, V.; Shoham, M.; Han, Y.W. Signal peptide of FadAadhesin from Fusobacterium nucleatum plays a novel structural role by modulating the filament’s length and width. FEBS Lett. 2012, 586, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Guo, P.; Tian, Z.; Kong, X.; Yang, L.; Shan, X.; Dong, B.; Ding, X.; Jing, X.; Jiang, C.; Jiang, N.; et al. FadA promotes DNA damage and progression of Fusobacterium nucleatum-induced colorectal cancer through up-regulation of chk2. J. Exp. Clin. Cancer Res. 2020, 39, 202. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, P.; Eslami, H.; Kafil, H.S. Carcinogenesis mechanisms of Fusobacterium nucleatum. Biomed. Pharmacother. 2017, 89, 918–925. [Google Scholar] [CrossRef] [PubMed]
- Krupa, K.; Lesiów, M.; Stokowa-Sołtys, K.; Starosta, R.; Ptaszyńska, N.; Łęgowska, A.; Rolka, K.; Wernecki, M.; Cal, M.; Jeżowska-Bojczuk, M. Copper(II) complexes with Fusobacterium nucleatum adhesin FadA: Coordination pattern, physicochemical properties and reactivity. J. Inorg. Biochem. 2018, 189, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.M.; Ashwaq, O.; Sarief, A.; Azad John Mohamed, A.K. A comprehensive review about SARS-CoV-2. Future Virol. 2020, 15, 625–648. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.J. The viral protein fragment theory of COVID-19 pathogenesis. Med. Hypotheses 2020, 144, 110267. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, U.C.; Shrivastava, R. Interaction of viral proteins with metal ions: Role in maintaining the structure and functions of viruses. FEMS Microbiol. Immunol. 2005, 43, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Li, J.; Wang, P.H.; Yang, N.; Huang, J.; Ou, J.; Xu, T.; Zhao, X.; Liu, T.; Huang, X.; et al. SARS-CoV-2 spike promotes inflammation and apoptosis through autophagy by ROS-suppressed PI3K/AKT/mTOR signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166260. [Google Scholar] [CrossRef]
- Raha, S.; Mallick, R.; Basak, S.; Duttaroy, A.K. Is copper beneficial for COVID-19 patients? Med. Hypotheses 2020, 142, 109814. [Google Scholar] [CrossRef]
- Leng, L.; Cao, R.; Ma, J.; Mou, D.; Zhu, Y.; Li, W.; Lv, L.; Gao, D.; Zhang, S.; Gong, F.; et al. Pathological features of COVID-19-associated lung injury: A preliminary proteomics report based on clinical samples, Signal Transduct. Target Ther. 2020, 5, 240. [Google Scholar] [CrossRef]
- Colunga Biancatelli, R.M.L.; Solopov, P.A.; Sharlow, E.R.; Lazo, J.S.; Marik, P.E.; Catravas, J.D. The SARS-CoV-2 spike protein subunit S1 induces COVID-19-like acute lung injury in Κ18-hACE2 transgenic mice and barrier dysfunction in human endothelial cells. Am. J. Physiol. Lung Cell Mol. Physiol. 2021, 321, L477–L484. [Google Scholar] [CrossRef] [PubMed]
- Besold, A.N.; Culbertson, E.M.; Culotta, V.C. The Yin and Yang of copper during infection. J. Biol. Inorg. Chem. 2016, 21, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Barhoumi, T.; Alghanem, B.; Shaibah, H.; Mansour, F.A.; Alamri, H.S.; Akiel, M.A.; Alroqi, F.; Boudjelal, M. SARS-CoV-2 Coronavirus Spike Protein-Induced Apoptosis, Inflammatory, and Oxidative Stress Responses in THP-1-Like-Macrophages: Potential Role of Angiotensin-Converting Enzyme Inhibitor (Perindopril). Front Immunol. 2021, 12, 728896. [Google Scholar] [CrossRef] [PubMed]
- Miripour, Z.S.; Sarrami-Forooshani, R.; Sanati, H.; Makarem, J.; Taheri, M.S.; Shojaeian, F.; Eskafi, A.H.; Abbasvandi, F.; Namdar, N.; Ghafari, H.; et al. Real-time diagnosis of reactive oxygen species (ROS) in fresh sputum by electrochemical tracing; correlation between COVID-19 and viral-induced ROS in lung/respiratory epithelium during this pandemic. Biosens. Bioelectron. 2020, 165, 112435. [Google Scholar] [CrossRef] [PubMed]
- Reshi, M.L.; Su, Y.C.; Hong, J.R. RNA Viruses: ROS-Mediated Cell Death. Int. J. Cell Biol. 2014, 2014, 467452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razzaq, Z.; Malik, A. Viral load is associated with abnormal serum levels of micronutrients and glutathione and glutathione-dependent enzymes in genotype 3 HCV patients. BBA Clin. 2014, 2, 72–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, M.; Rathje, O.; Levina, A.; Lay, P.A. High cytotoxicity of vanadium(IV) complexes with 1,10-phenanthroline and related ligands is due to decomposition in cell culture medium. Biol. Inorg. Chem. 2017, 22, 663–672. [Google Scholar] [CrossRef]
- Martínez-Valencia, B.; Corona-Motolinia, N.D.; Sánchez-Laraa, E.; Norieg, L.; Sánchez-Gaytán, B.L.; Castro, M.E.; Meléndez-Bustamante, F.; González-Vergara, E. Cyclo-tetravanadate bridged copper complexes as potential double bullet pro-metallodrugs for cancer treatment. J. Inorg. Biochem. 2020, 208, 111081–111092. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowska, A.; Szuster-Ciesielska, A.; Sztandera, M.; Bregier-Jarzebowska, R.; Jarzab, A.; Rojek, T.; Komarnicka, U.K.; Bojarska-Junak, T.; Jezierska, J. L-argininatocopper(II) complexes in solution exert significant selective anticancer and antimicrobial activities. Appl. Organomet. Chem. 2020, 34, 5698–5706. [Google Scholar] [CrossRef]
- Wojciechowska, A.; Gągor, A.; Zierkiewicz, W.; Jarząb, A.; Dylonga, A.; Duczmala, M. Metal–organic framework in an l-arginine copper(ii) ion polymer: Structure, properties, theoretical studies and microbiological activity. RSC Adv. 2015, 5, 36295–36307. [Google Scholar] [CrossRef]
- Badetti, E.; Calgaro, L.; Falchi, L.; Bonetto, A.; Bettiol, C.; Leonetti, B.; Ambrosi, E.; Zendri, E.; Marcomini, P. Interaction between Copper Oxide Nanoparticles and Amino Acids: Influence on the Antibacterial Activity. Nanomaterials 2019, 9, 792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hecke, K.; Nam, P.C.; Nguyen, M.T.; Van Meervelt, L. Netropsin interactions in the minor groove of d(GGCCAATTGG) studied by a combination of resolution enhancement and ab initio calculations. FEBS J. 2005, 272, 3531–3539. [Google Scholar] [CrossRef] [PubMed]
- Mashima, T.; Nishikawa, F.; Kamatari, Y.O.; Fujiwara, H.; Saimura, M.; Nagata, T.; Kodaki, T.; Nishikawa, S.; Kuwata, K.; Katahira, M. Anti-prion activity of an RNA aptamer and its structural basis. Nucleic Acids Res. 2013, 41, 1355–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macmaster, R.; Sedelnikova, S.; Baker, P.J.; Bolt, E.L.; Lloyd, R.G.; Rafferty, J.B. RusA Holliday junction resolvase: DNA complex structure—Insights into selectivity and specificity. Nucleic Acids Res. 2006, 34, 5577–5582. [Google Scholar] [CrossRef] [PubMed]
- Liao, S.R.; Le, X.Y.; Feng, X.L. Syntheses, characterizations and SOD-like activities of ternary copper(II) complexes with 1,10-phenanthroline and L-α-amino acids. J. Coord. Chem. 2008, 61, 847–854. [Google Scholar] [CrossRef]
- Harada, K.; Franke, A.D. Identification of two novel arginine binding DNAs. EMBO 1995, 14, 5798–5809. [Google Scholar] [CrossRef]
- Conklin, K.A. Chemotherapy-associated oxidative stress: Impact on chemotherapeutic effectiveness. Integr. Cancer Ther. 2004, 3, 294–300. [Google Scholar] [CrossRef]
- Srinivasa, S.U.; Bryce, W.Q.T.; Vellayappan, B.V.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef]
- Wojciechowska, A.; Rojek, T.; Gorzsas, A.; Malik-Gajewska, M.; Duczmal, M. Isothiocyanate controlled architecture, spectroscopic, and magnetic behavior of copper(II) l–arginine complexes. J. Coord. Chem. 2019, 72, 1358–1362. [Google Scholar] [CrossRef]
- Hosseininezhad, S.M.; Hosseinali, R. An analysis of the reactions of L-arginine with Cu(II), Co(II), Fe(III), Zn(II), and Cr(III). Adv. Environ. Biol. 2014, 7, 315. [Google Scholar]
- Wojciechowska, A.; Bregier, R.; Komarnicka, U.K.; Kozieł, S.; Szuster, A.; Sztandera, M.; Jarząb, A.; Staszak, Z.; Witkowska, D.; Bojarska, A.; et al. Isothiocyanate l−argininato copper(II) complexes—Solution structure, DNA interaction, anticancer and antimicrobial activity. Chem. Biol. Interact. 2021, 348, 109636–109641. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hao, S.; Han, A.; Yang, Y.; Fang, G.; Wang, S. Intracellular Fenton reaction based on mitochondria-targeted copper(II)–peptide complex for induced apoptosis. J. Mater. Chem. 2019, 7, 4008–4016. [Google Scholar] [CrossRef]
- Chekmenev, E.Y.; Vollmar, B.S.; Forseth, K.T.; Manion, M.C.; Jones, S.M.; Wagner, T.J.; Endicott, R.M.; Kyriss, B.P.; Homem, L.M.; Pate, M.; et al. Investigating molecular recognition and biological function at interfaces using piscidins, antimicrobial peptides from fish. Biochim. Biophys. Acta 2006, 1758, 1359–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chekmenev, E.Y.; Jones, S.M.; Nikolayeva, Y.M.; Vollmar, B.S.; Wagner, T.J.; Gor’kov, P.L.; Brey, W.W.; Manion, M.N.; Cotton, D.M. High-field NMR studies of molecular recognition and structure-function relationships in antimicrobial piscidins at the water-lipid bilayer interface. J. Am. Chem. Soc. 2006, 128, 5308–5309. [Google Scholar] [CrossRef] [PubMed]
- Comert, F.; Heinrich, F.; Chowdhury, A.; Schoeneck, M.; Darling, D.; Anderson, K.W.; Daben, M.; Angeles-Boza, A.M.; Silin, V.; Cotton, M.L.; et al. Copper-binding anticancer peptides from the piscidin family: An expanded mechanism that encompasses physical and chemical bilayer disruption. Sci. Rep. 2021, 11, 12620–12631. [Google Scholar] [CrossRef] [PubMed]
- Mihailescu, M.; Sorci, M.; Seckute, J.; Silin, V.I.; Hammer, J.; Perrin, P.S., Jr.; Hernandez, J.I.; Smajic, N.; Shrestha, A.; Bogardus, K.A.; et al. Structure and function in antimicrobial piscidins: Histidine position, directionality of membrane insertion, and pH-dependent permeabilization. J. Am. Chem. Soc. 2019, 141, 9837–9853. [Google Scholar] [CrossRef] [PubMed]
- Comert, F.; Greenwood, A.; Maramba, J.; Acevedo, R.; Lucas, L.; Kulasinghe, T.; Cairns, L.S.; Wen, Y.; Fu, R.; Hammer, J.; et al. The host-defense peptide piscidin P1 reorganizes lipid domains in membranes and decreases activation energiesin mechanosensitive ion channels. J. Biol. Chem. 2019, 294, 18557–18570. [Google Scholar] [CrossRef] [PubMed]
- Libardo, M.D.J.; Bahar, A.A.; Ma, B.; Fu, R.; McCormick, L.E.; Zhao, J.; McCallum, S.A.; Nussinov, R.; Ren, D.; Angeles-Boza, A.M.; et al. Nuclease activity gives an edge to host-defense peptide piscidin 3 over piscidin 1, rendering it more effective against persisters and biofilms. FEBS J. 2017, 284, 3662–3683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Z.; Liu, Q.; Zhu, Q.; Yang, B.; Khaliq, H.; Sun, A.; Qi, Y.; Moku, G.K.; Su, Y.; Wang, J.; et al. Comparative pharmacokinetics and preliminary pharmacodynamics evaluation of piscidin 1 against PRV and PEDVin rats. Front Chem. 2018, 6, 244. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.; Guo, N.; Chen, S.; Wang, Y.; Liu, B.; He, O. Antiviral activity of piscidin 1 against pseudorabies virus both in vitro and in vivo. Virol. J. 2019, 16, 95. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.-J.; Huang, T.-C.; Muthusamy, S.; Lee, J.-F.; Duann, Y.-F.; Lin, C.-H. Piscidin-1, an antimicrobial peptide from fish (Hybrid Striped Bass Moronesaxatilis x M. chrysops), induces apoptotic and necrotic activity in HT1080 cells. Zool. Sci. 2012, 29, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Facchin, G.; Torre, M.H.; Kremer, E.; Piro, O.E.; Castellano, E.E.; Baran, E.J. Synthesis and characterization of three new Cu(II)-dipeptide complexes. J. Inorg. Biochem. 2002, 89, 174–180. [Google Scholar] [CrossRef]
- Facchin, G.; Torre, M.E. Cu (II) complexation with His–Gly and His–Ala. X-ray structure of [Cu (his–gly)2(H2O)2]·6H2O. Inorg. Chim. Acta 2003, 355, 408–413. [Google Scholar] [CrossRef]
- Sanchiz, J.; Kremer, C. Magnetic properties of copper (II) complexes containing peptides. Crystal structure of [Cu (phe-leu)]. J. Mol. Struct. 2006, 797, 179–183. [Google Scholar] [CrossRef]
- Facchin, G.; Kremer, E. Interaction of Cu-dipeptide complexes with calf thymus DNA and antiproliferative activity of [Cu (ala-phe)] in osteosarcoma-derived cells. Polyhedron 2009, 28, 2329–2334. [Google Scholar] [CrossRef]
- Iglesias, S.; Alvarez, N.; Ribeiro, R.R.; Barroso, R.P.; Costa-Filho, A.J.; Kramer, M.G.; Facchin, G. Synthesis, structural characterization and cytotoxic activity of ternary copper (II)–dipeptide–phenanthroline complexes. A step towards the development of new copper compounds for the treatment of cancer. J. Inorg. Biochem. 2014, 139, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.; Sinn, E.; Martin, R.B. Crystal structure of a mixed-ligand complex of copper (II), 1, 10-phenanthroline, and glycylglycinedianion: Glycylglycinato (1, 10-phenanthroline) copper (II) trihydrate. Inorg. Chem. 1976, 15, 807–811. [Google Scholar] [CrossRef]
- Bhirud, R.G.; Srivastava, T.S. Synthesis, characterization and superoxide dismutase activity of some ternary copper (II) dipeptide-2,2′-bipyridine, 1,10-phenanthroline and 2,9-dimethyl-1,10-phenanthroline complexes. Inorg. Chim. Acta 1991, 179, 125–131. [Google Scholar] [CrossRef]
- Alvarez, S.N.; Kramer, M.; Torre, M.H.; Kremer, E.; Ellena, J.; Filho, A.; Facchin, G. Structural characterization and cytotoxic activity of heteroleptic copper (II) complexes with L-dipeptides and 5-NO2-phenanthroline. Crystal structure of [Cu(Phe-Ala)(5-NO2-Phen)]·4H2O. Struct. Chem. Crystallogr. Commun. 2015, 1, 1–7. [Google Scholar]
- Vieira, E.D.; Casado, N.M. Weak exchange interaction supported by a biologically relevant long chemical bridge in a Cu-peptide model compound. Inorg. Chem. 2006, 45, 2942–2947. [Google Scholar] [CrossRef]
- Ng, C.; Kong, S.M.; Tiong, Y.L.; Maah, M.J.; Sukram, N.; Ahmad, M.; Khoo, B.A. Selective anticancer copper(II)-mixed ligand complexes: Targeting of ROS and proteasomes. Metallomics 2014, 6, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Facchin, G.; Kremer, E. Structural characterization of a series of new Cu-dipeptide complexes in solid state and in solution. Polyhedron 2006, 25, 2597–2604. [Google Scholar] [CrossRef]
- Facchin, G.; Veiga, N.; Kramer, M.G.; Batista, A.A.; Várnagy, K.; Farkas, E.; Moreno, V.; Torre, M.H. Experimental and theoretical studies of copper complexes with isomeric dipeptides as novel candidates against breast cancer. J. Inorg. Biochem. 2016, 162, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, N.; Noble, C.; Torre, M.H.; Kremer, E.; Costa-Filhod, A.J.; Mendesd, L.F.; Kramere, M.G.; Facchina, G. Synthesis, structural characterization and cytotoxic activity against tumor cells of heteroleptic copper (I) complexes with aromatic diimines and phosphines. Inorg. Chim. Acta 2017, 466, 559–564. [Google Scholar] [CrossRef]
- Sugimori, T.; Shibakawa, K.; Sugimori, T.; Shibakawa, K.; Masuda, H.; Odani, A.; Yamauchi, O. Ternary metal (II) complexes with tyrosine-containing dipeptides. Structures of copper (II) and palladium (II) complexes involving L-tyrosylglycine and stabilization of copper (II) complexes due to intramolecular aromatic ring stacking. Inorg. Chem. 1993, 32, 4951–4959. [Google Scholar] [CrossRef]
- Gaála, A.; Mihucz, V.G.; Bősze, S.; Szabó, I.; Baranyi, M.; Horváth, P.; Streli, C.; Szoboszla, N. Comparative in vitro investigation of anticancer copper chelating agents. Microchem. J. 2018, 136, 227–235. [Google Scholar] [CrossRef]
- Xia-Bing, F.; Zhang, J.-J.; Liu, D.-D.; Gan, Q.; Gao, H.-W.; Mao, Z.-W.; Le, X.-Y. Cu(II)–dipeptide complexes of 2-(4′-thiazolyl)benzimidazole: Synthesis, DNA oxidative damage, antioxidant and in vitro antitumor activity. J. Inorg. Biochem. 2015, 143, 77–87. [Google Scholar]
- Gana, Q.; Zhang, C.-L.; Wang, B.-F.; Xiong, Y.-H.; Fu, Y.-L.; Maoac, Z.-W.; Le, X.-Y. Two new mixed copper(II)–dipeptide complexes of N,N-donor heterocycle ligands: Studies on their non-covalent DNA binding, chemical nuclease, antioxidant and anticancer activities. RSC Adv. 2016, 6, 35952–35965. [Google Scholar] [CrossRef]
- Qi, Y.Y.; Gan, Q.; Liu, Y.X.; Xiong, Y.H.; Mao, Z.W.; Le, X.Y. Two new Cu(II) dipeptide complexes based on 5-methyl-2-(2′-pyridyl)benzimidazole as potential antimicrobial and anticancer drugs: Special exploration of their possible anticancer mechanism. Eur. J. Med. Chem. 2018, 154, 220–232. [Google Scholar] [CrossRef] [PubMed]
- Komarnicka, U.K.; Kozieł, S.; Starosta, R.; Kyzioł, A. Selective Cu(I) complex with phosphine-peptide (SarGly) conjugate contra breast cancer: Synthesis, spectroscopic characterization and insight into cytotoxic action. J. Inorg. Biochem. 2018, 186, 162–175. [Google Scholar] [CrossRef]
- Komarnicka, U.K.; Kozieł, S.; Zabierowski, P.; Kruszyński, R.; Lesiówa, M.K.; Tisato, F.; Porchia, M.; Kyzioł, A. Copper(I) complexes with phosphines P(p-OCH3-Ph)2CH2OH and P(p-OCH3-Ph)2CH2SarGly. Synthesis, multimodal DNA interactions, and prooxidative and in vitro antiproliferative activity. J. Inorg. Biochem. 2020, 203, 110926. [Google Scholar] [CrossRef] [PubMed]
- Hegde, M.L.; Bharathi, P.; Suram, A.; Venugopal, C.; Jagannathan, R.; Poddar, P.; Srinivas, P.; Sambamurti, K.; Rao, K.J.; Scancar, J.; et al. Challenges Associated with Metal Chelation Therapy in Alzheimer’s Disease. J. Alzheimers Dis. 2009, 17, 457–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szeto, J.Y.Y.; Lewis, S.J.G. Current Treatment Options for Alzheimer’s Disease and Parkinson’s Disease Dementia. Curr. Neuropharmacol. 2016, 14, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Pickart, L.; Michelle, J.; Soltero, V.; Margolina, A. The Human Tripeptide GHK-Cu in Prevention of Oxidative Stress and Degenerative Conditions of Aging:Implications for Cognitive Health. Oxidative Med. Cell. Longev. 2012, 5, 8. [Google Scholar]
- Boas, D. A Novel Copper-Binding Peptide That Self-Assembles Into a Transparent Antibacterial and Antiviral Coating Front. Bioeng. Biotechnol. 2021, 20, 11–19. [Google Scholar]
- Andersen, A.; Chen, Y.; Birkedal, H. Bioinspired Metal–Polyphenol Materials: Self-Healing and Beyond. Biomimetics 2019, 4, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagripanti, J.L.; Routson, L.B.; Bonifacino, A.C.; Lytle, C.D. Mechanism of coppermediatedinactivation of herpes simplex virus. Antimicrob. Agents Chemother. 1997, 41, 812–817. [Google Scholar] [CrossRef] [Green Version]
- Betanzos-Cabrera, G.; Rez, F.J.R.; Oz, J.L.M.; Barrn, B.L.; Maldonado, R. Inactivation ofHSV-2 by ascorbate-Cu (II) and its protecting evaluation in CF-1 miceagainst encephalitis. J. Virol. Methods 2004, 120, 8. [Google Scholar] [CrossRef]
- Guo, W.J.; Ye, S.S.; Cao, N.; Huang, J.; Gao, J.; Chen, Q.Y. ROS-mediatedautophagy was involved in cancer cell death induced by novel copper(II)complex. Exp. Toxicol. Pathol. 2010, 62, 577–582. [Google Scholar] [CrossRef]




















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komarnicka, U.K.; Lesiów, M.K.; Witwicki, M.; Bieńko, A. The Bright and Dark Sides of Reactive Oxygen Species Generated by Copper–Peptide Complexes. Separations 2022, 9, 73. https://doi.org/10.3390/separations9030073
Komarnicka UK, Lesiów MK, Witwicki M, Bieńko A. The Bright and Dark Sides of Reactive Oxygen Species Generated by Copper–Peptide Complexes. Separations. 2022; 9(3):73. https://doi.org/10.3390/separations9030073
Chicago/Turabian StyleKomarnicka, Urszula K., Monika K. Lesiów, Maciej Witwicki, and Alina Bieńko. 2022. "The Bright and Dark Sides of Reactive Oxygen Species Generated by Copper–Peptide Complexes" Separations 9, no. 3: 73. https://doi.org/10.3390/separations9030073
APA StyleKomarnicka, U. K., Lesiów, M. K., Witwicki, M., & Bieńko, A. (2022). The Bright and Dark Sides of Reactive Oxygen Species Generated by Copper–Peptide Complexes. Separations, 9(3), 73. https://doi.org/10.3390/separations9030073