
The Redox Active [2Fe-2S] Clusters: Key-Components of a Plethora of Enzymatic Reactions—Part I: Archaea



Abstract
:
1. Introduction
2. Archaeal Enzymatic Reactions
2.1. Archaeoglobus fulgidus
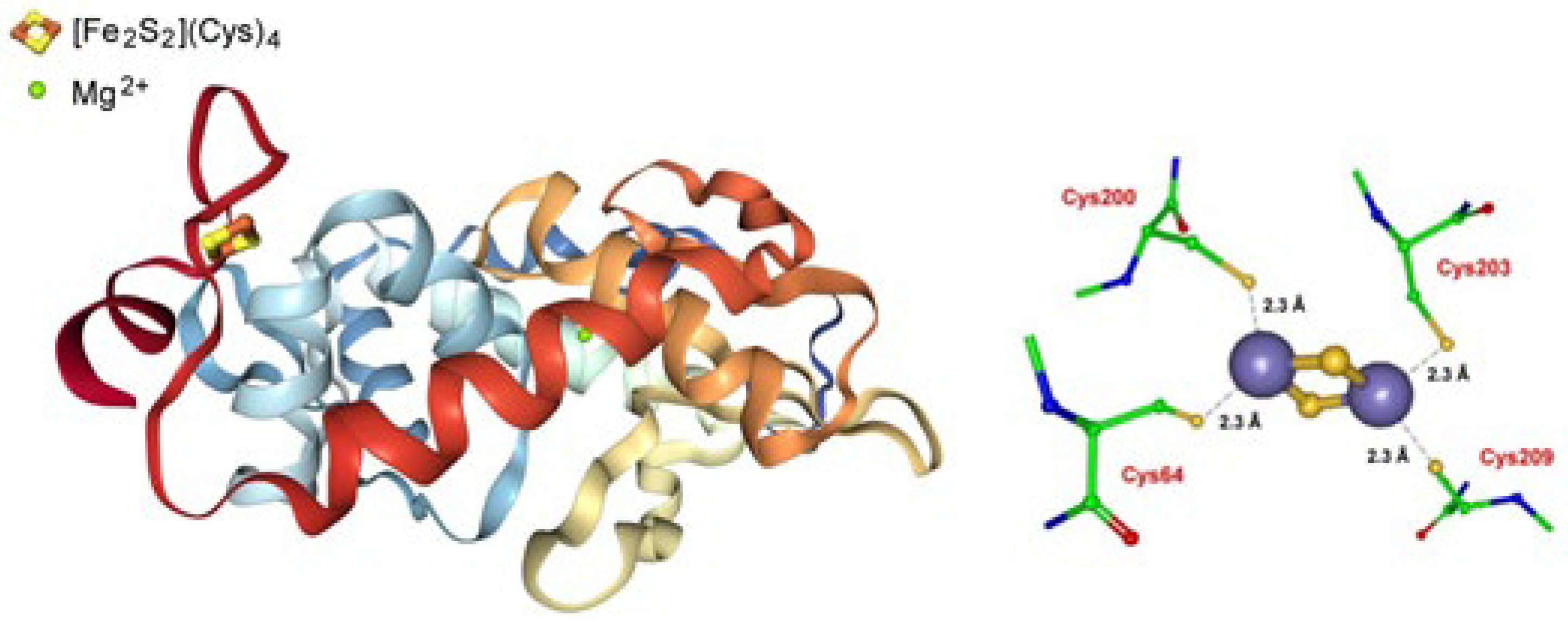
2.1.1. Copper Chaperone CopZ
2.1.2. Cysteine Desulfurase
2.2. Methanothrix thermoacetophila
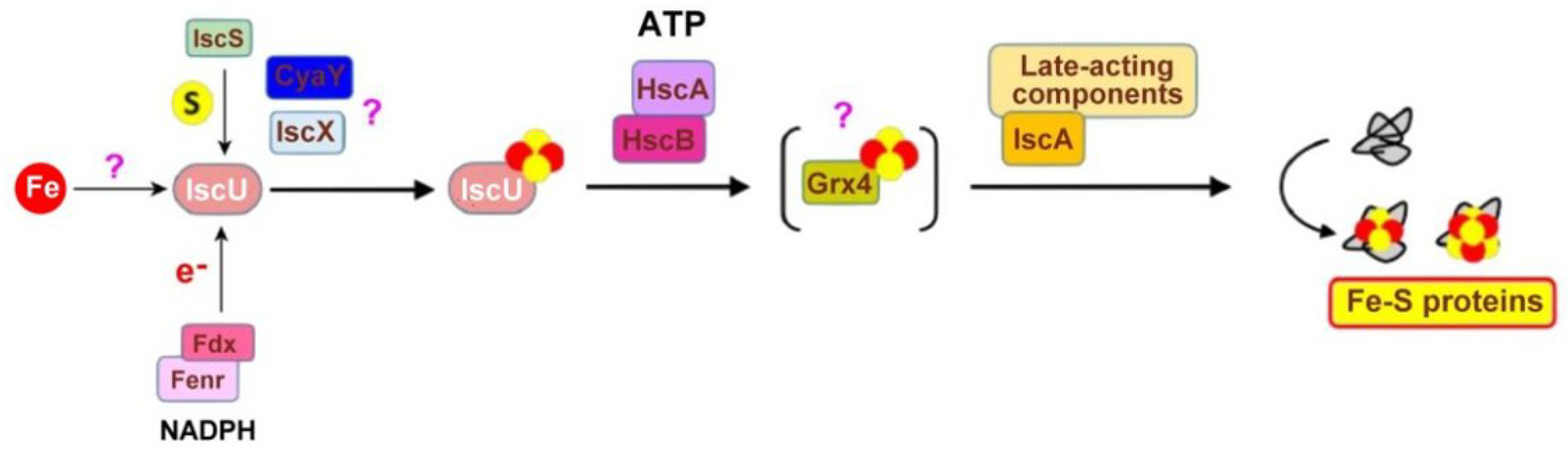
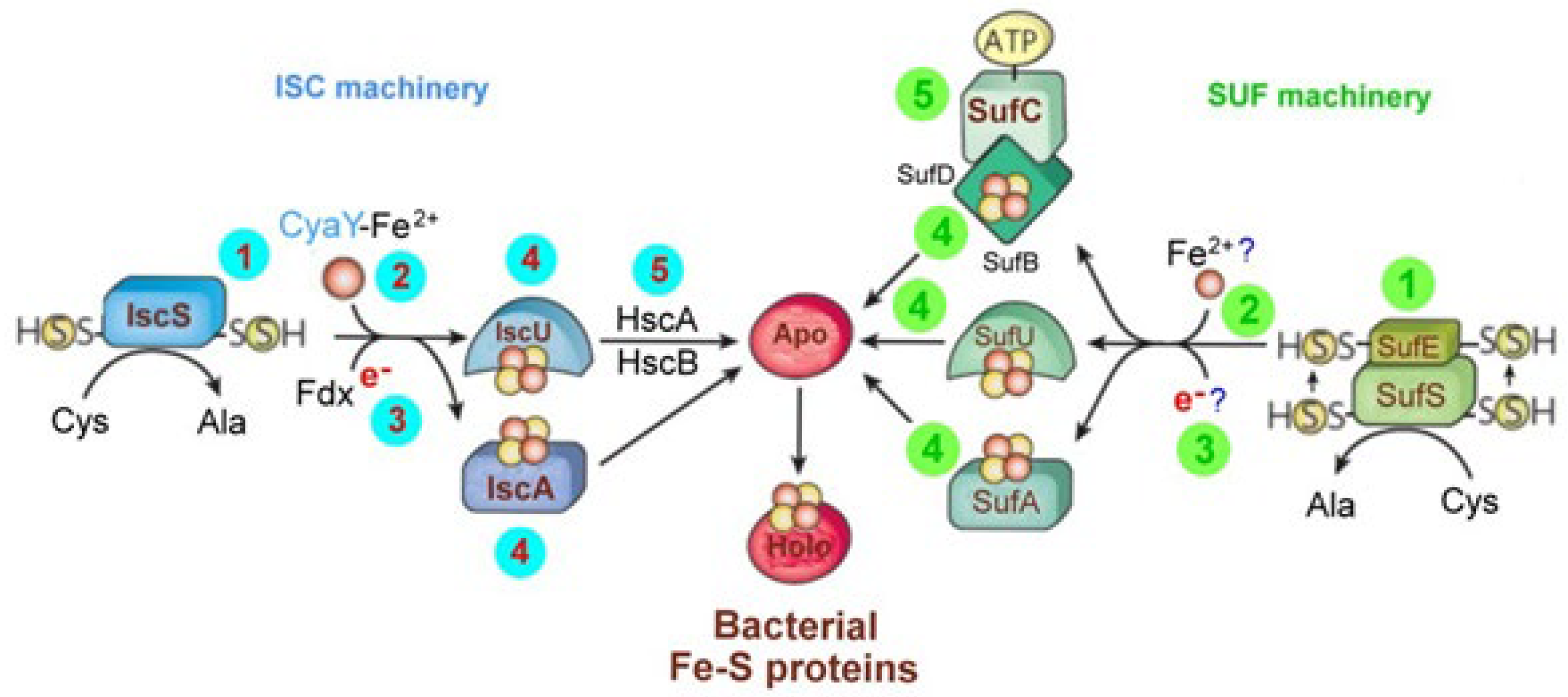
The IscU System
2.3. Pyrobaculum calidifontis
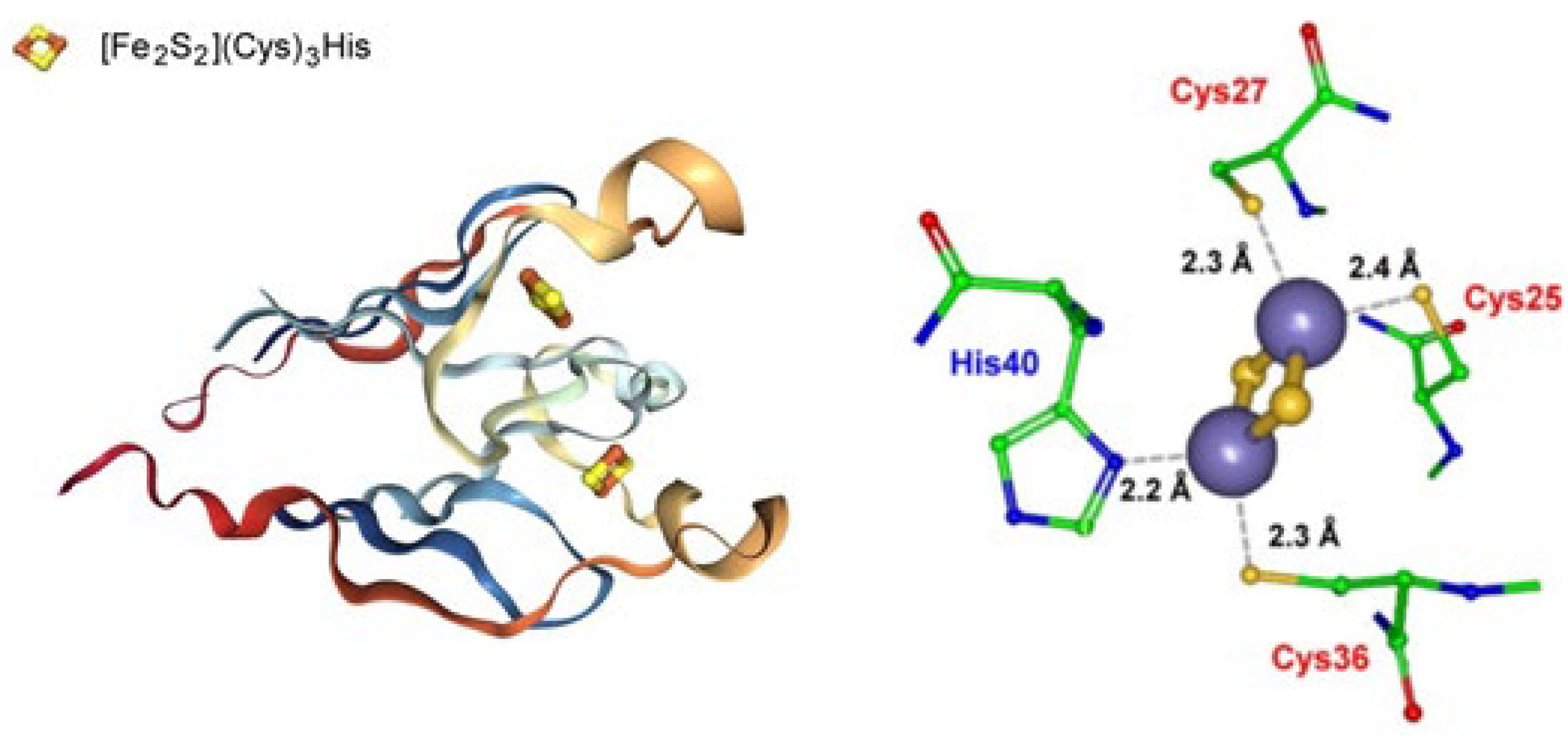
2.3.1. CRISPR-Associated Exonuclease Cas4
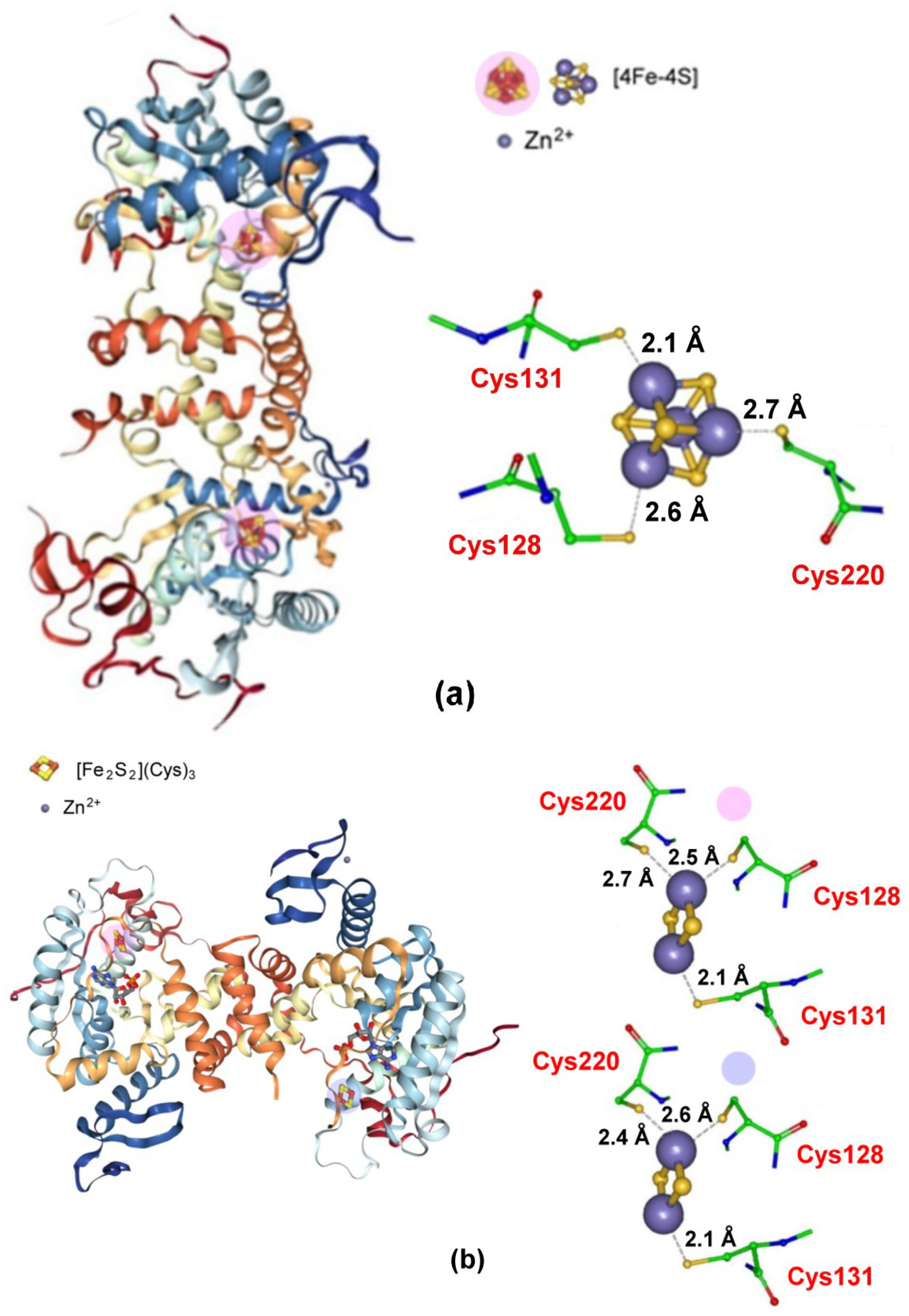
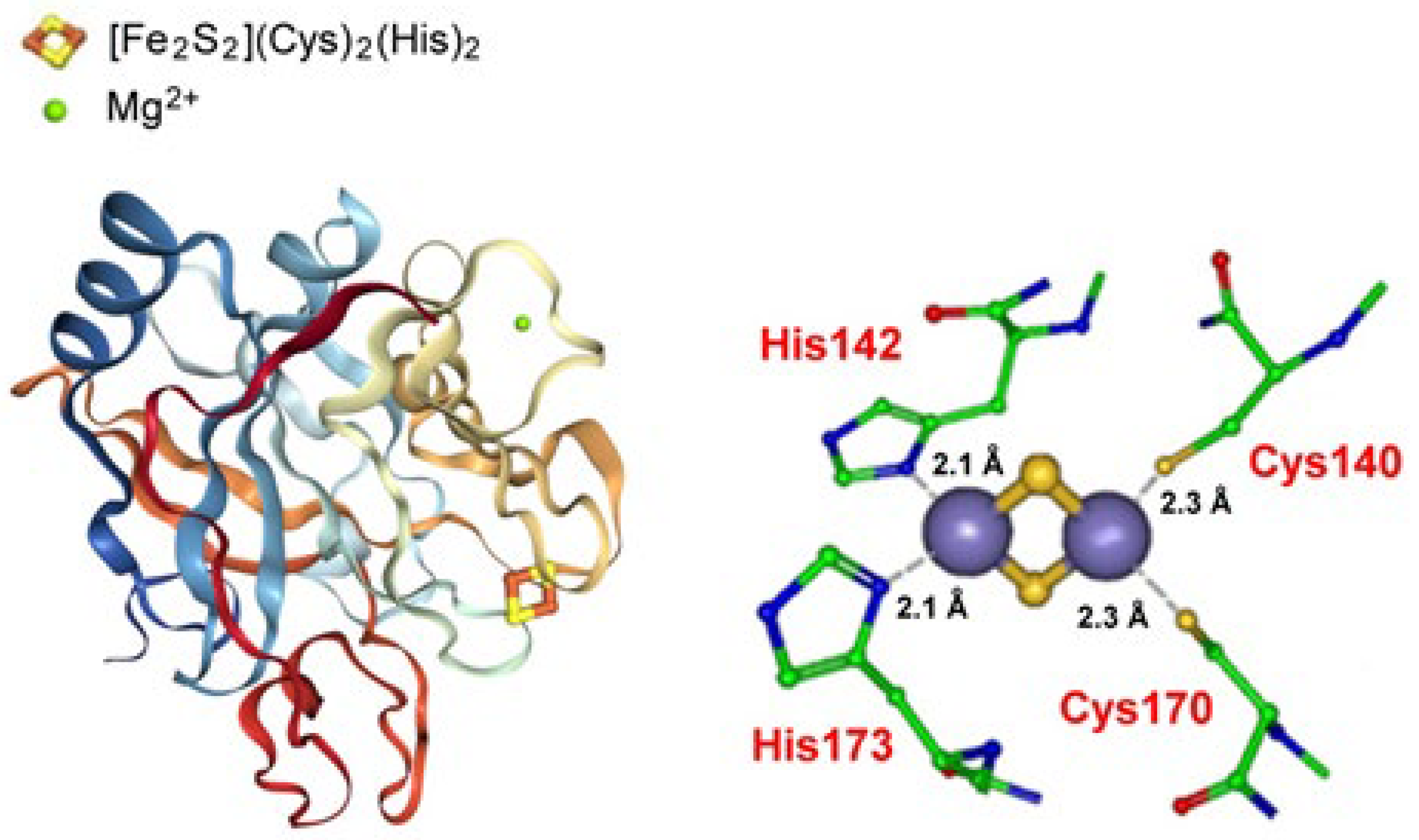
2.3.2. Zinc Finger, CDGSH-Type Domain Proteins
2.4. Pyrococcus horikoshii
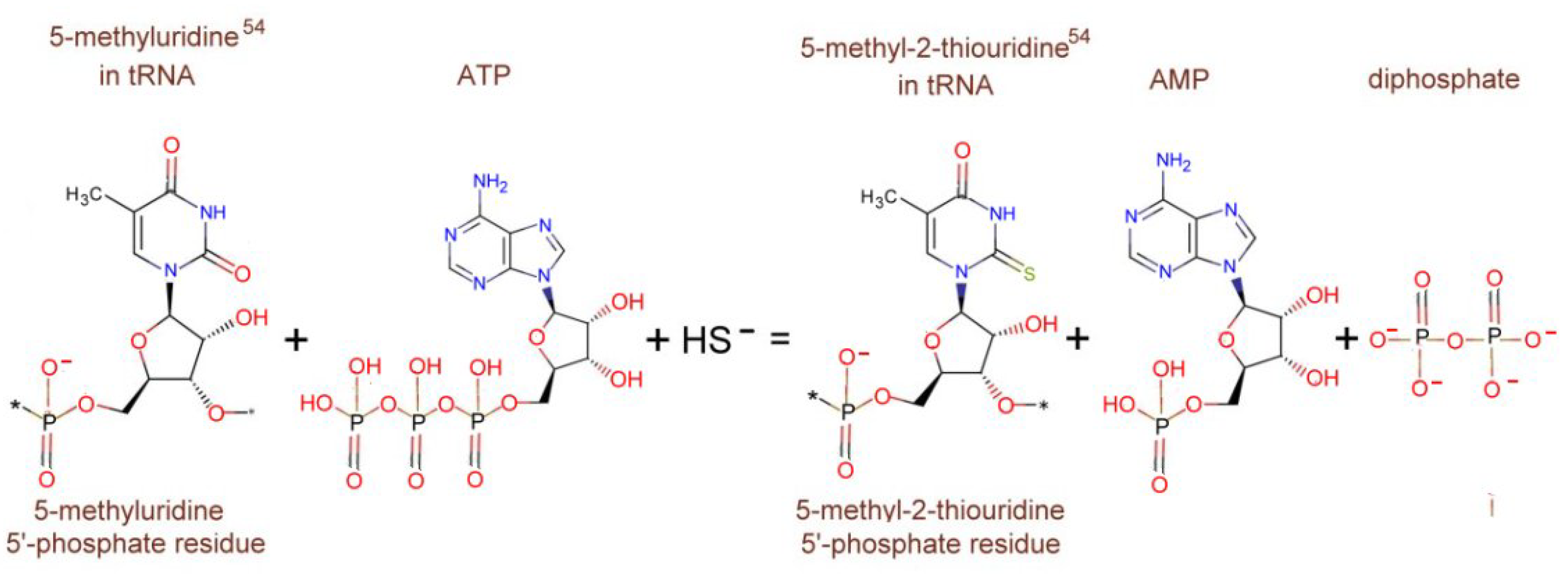
tRNA-5-methyluridine(54) 2-sulfurtransferase
2.5. Sulfolobus acidocaldarius
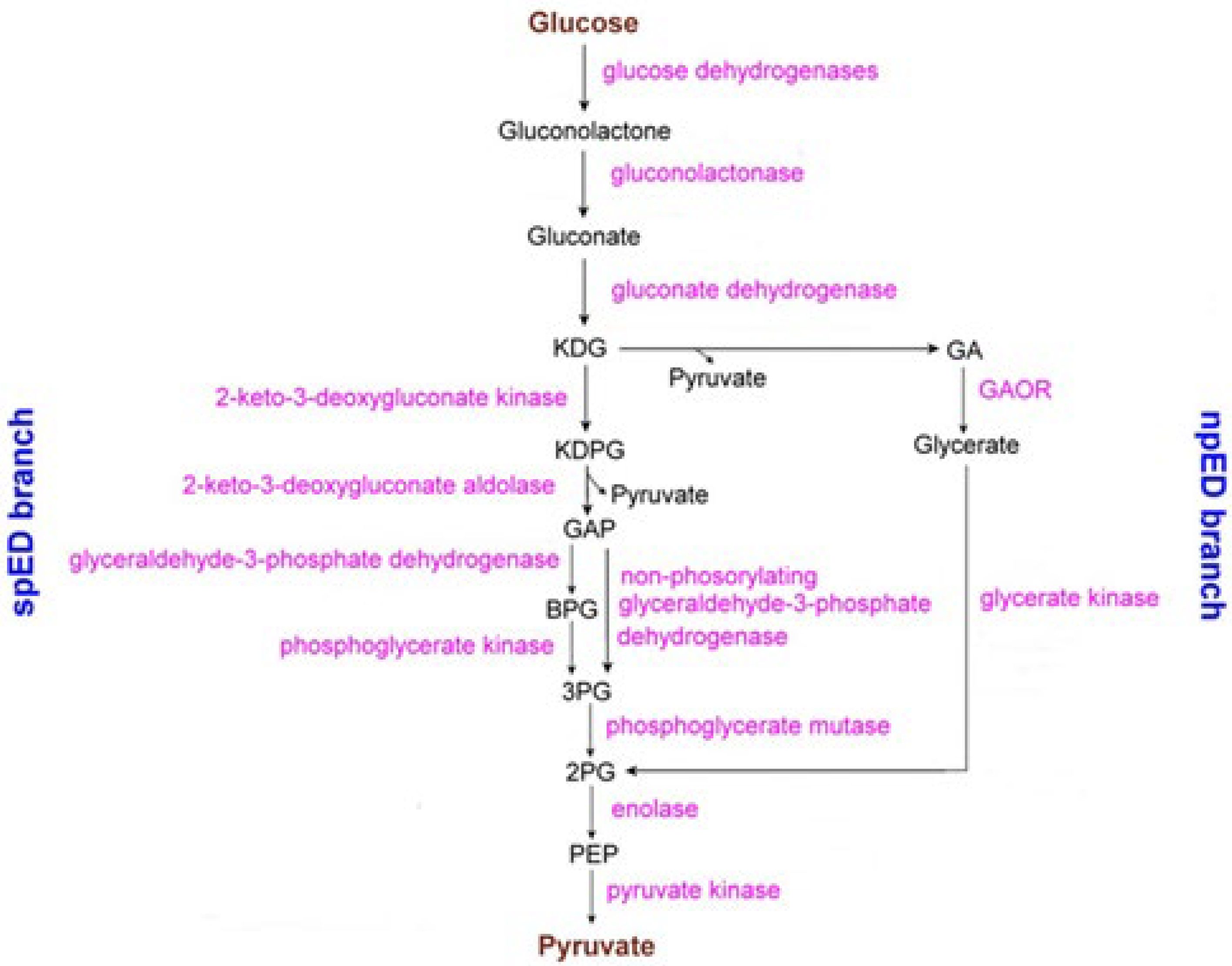
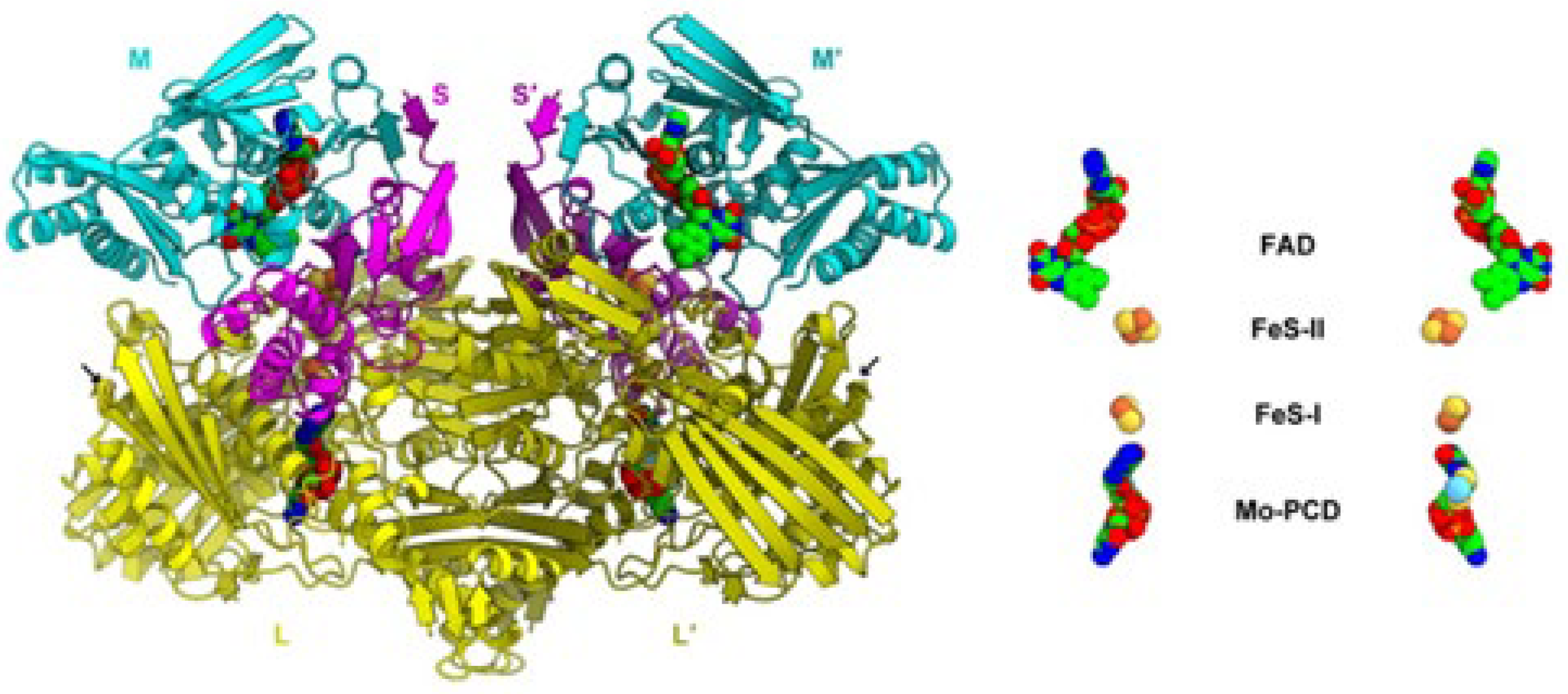
The Respiratory System
- The SoxABCD complex (that acts as a proton pump in reconstituted in vitro system [128] as well as in vivo [129]) with its subunits SoxA (no prostethic group), SoxB, and SoxC (both having the prostethic group cytochrome As (the pedice S means Sulfolobus) [130,131]), and SoxD (we do not deal with SoxABCD as it does not contain any Fe-S cluster; for more details about such a cytochrome oxygenase, see reference [131]).
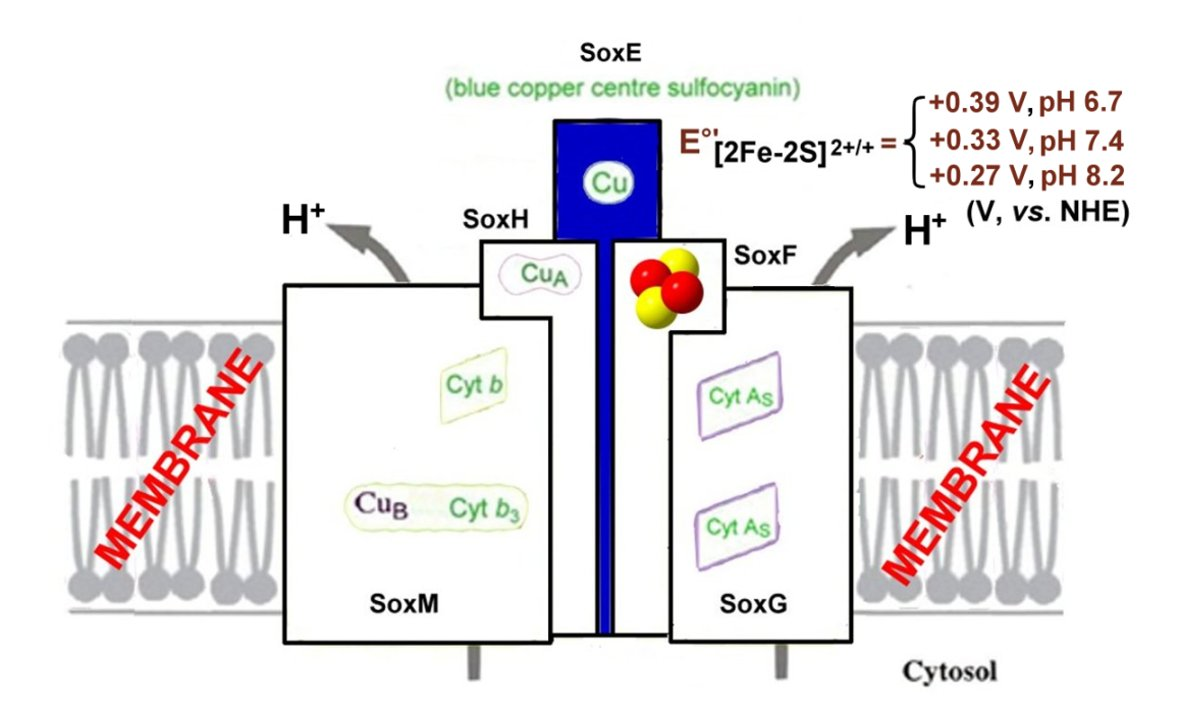
- The SoxM complex constitutes a unique respiratory supercomplex combining features of a cytochrome bc1 complex (subunits SoxF, SoxG, and SoxE) and cytochrome c oxidase (subunits SoxM, SoxH, and SoxL). In fact, SoxM hosts two cytochrome-b types, namely b and b3, that constitute a quinol bb3 oxidase [132]), where cytochrome b encodes a CuA site located in the subunit II (SoxH) typical of cytochrome c oxidase that constitutes the primary electron acceptor (it is probably linked to two histidines, a cysteine and a methionine of cupredoxin [133]) and the cytochrome b3 that is coupled to a CuB center that catalyzes the dioxygen reduction to two water molecules, thus completing the cellular respiration process [134]; in fact, SoxH bears a prostethic group CuA that resides in subunit II of cytochrome c oxidase and hosts two heme As. SoxF hosts a iron-sulfur protein II. SoxE bears the blue copper centre sulfocyanin, and finally SoxI hosts an iron-sulfur protein I.
2.6. Sulfurisphaera tokodaii
3. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kaiser, G. Classification—The Three Domain System. Community College of Baltimore Country (Cantonsville). Available online: https://bio.libretexts.org/@go/page/2699 (accessed on 3 January 2021).
- Bar-On, Y.M.; Phillips, R.; Milo, R. The biomass distribution on Earth. Proc. Natl. Acad. Sci. USA 2018, 115, 6506–6511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woese, C.R.; Fox, G.E. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proc. Natl. Acad. Sci. USA 1977, 74, 5088–5090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woese, C.R. These unusual bacteria are genealogically neither prokaryotes nor eukaryotes. This discovery means there are not two lines of descent but three: The archaebacteria, the true bacteria and the eukaryotes. Sci. Am. 1981, 244, 98–125. [Google Scholar] [CrossRef]
- Woese, C.R. There must be a prokaryote somewhere: Microbiology’s search for itself. Microbiol. Rev. 1994, 58, 1–9. [Google Scholar] [CrossRef]
- Woese, C.R. The universal ancestor. Proc. Natl. Acad. Sci USA 1998, 95, 6854–6859. [Google Scholar] [CrossRef] [Green Version]
- Woese, C.R. Interpreting the universal phylogenetic tree. Proc. Natl. Acad. Sci. USA 2000, 97, 8392–8396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woese, C.R. On the evolution of cells. Proc. Natl. Acad. Sci. USA 2002, 99, 8742–8747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Organismal Biology. Prokaryotes: Bacteria & Archaea (See Biodiversity). Available online: https://organismalbio.biosci.gatech.edu (accessed on 26 June 2021).
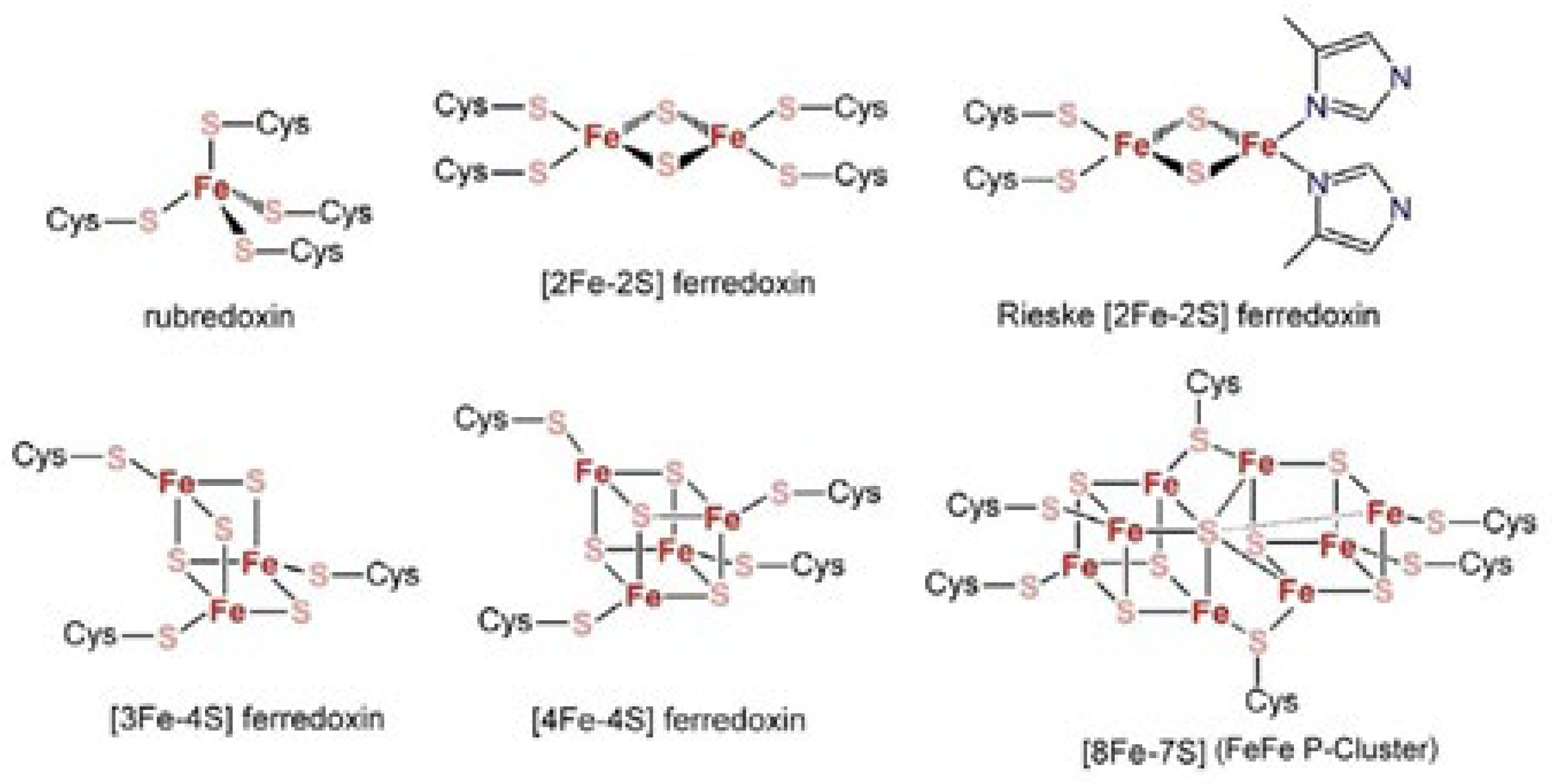
- Fabrizi de Biani, F.; Zanello, P. The competition between chemistry and biology in assembling iron-sulfur derivatives. Molecular structures and electrochemistry. Part IV. [Fe3S4](SCys)3 proteins. Inorg. Chim. Acta 2017, 455, 319–328. [Google Scholar] [CrossRef]
- Beinert, H. Iron-sulfur proteins: Ancient structures, still full of surprises. J. Biol. Inorg. Chem. 2000, 5, 2–15. [Google Scholar] [CrossRef]
- Johnson, D.C.; Dean, D.; Smith, A.D.; Johnson, M.K. Structure, function, and formation of biological iron-sulfur clusters. Annu. Rev. Biochem. 2005, 74, 247–281. [Google Scholar] [CrossRef] [PubMed]
- Lukianova, O.A.; David, S.S. A role for ironsulfur clusters in DNA repair. Curr. Opin. Chem. Biol. 2005, 9, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Fontecave, M. Iron-sulfur clusters: Ever-expanding roles. Nat. Chem. Biol. 2006, 4, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Outten, F.W. Iron-sulfur clusters as oxygen-responsive molecular switches. Nat. Chem. Biol. 2007, 3, 206–207. [Google Scholar] [CrossRef] [PubMed]
- Wollers, S.; Layer, G.; Garcia-Serres, R.; Signor, L.; Clemancey, M.; Latour, J.-M.; Fontecave, M.; Ollagnier de Choudens, S. Iron-Sulfur (Fe-S) Cluster Assembly. J. Biol. Chem. 2010, 285, 23331–23341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crack, J.C.; Green, J.; Thomson, A.J.; Le Brun, N.E. Iron–sulfur cluster sensor-regulators. Curr. Opin. Chem. Biol. 2012, 16, 35–44. [Google Scholar] [CrossRef]
- Peters, J.W.; Broderick, J.B. Emerging Paradigms for Complex Iron-Sulfur Cofactor Assembly and Insertion. Annu. Rev. Biochem. 2012, 81, 429–450. [Google Scholar] [CrossRef]
- Balk, J.; Schaedler, T.A. Iron Cofactor Assembly in Plants. Annu. Rev. Plant Biol. 2014, 65, 125–153. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Sivalingam, K.; Neese, F.; Chan, G.K.-L. Low-energy spectrum of iron–sulfur clusters directly from many-particle quantum mechanics. Nat. Chem. 2014, 6, 927–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiban, J.; So, M.; Kaguni, L.S. Iron-sulfur clusters in mitochondrial metabolism: Multifaceted roles of a simple cofactor. Biochemistry 2016, 81, 1066–1080. [Google Scholar] [CrossRef] [PubMed]
- Zanello, P. The competition between chemistry and biology in assembling iron–sulfur derivatives. Molecular structures and electrochemistry. Part I. {Fe(SγCys)4} proteins. Coord. Chem. Rev. 2013, 257, 1777–1805. [Google Scholar] [CrossRef]
- Zanello, P. The competition between chemistry and biology in assembling iron–sulfur derivatives. Molecular structures and electrochemistry. Part II. {[Fe2S2](SγCys)4} proteins. Coord. Chem. Rev. 2014, 280, 54–83. [Google Scholar] [CrossRef]
- Zanello, P. The competition between chemistry and biology in assembling iron–sulfur derivatives. Molecular structures and electrochemistry. Part III. {[Fe2S2](Cys)3(X)} (X = Asp, Arg, His) and {[Fe2S2](Cys)2(His)2} proteins. Coord. Chem. Rev. 2016, 306, 420–442. [Google Scholar] [CrossRef]
- Zanello, P. The competition between chemistry and biology in assembling iron–sulfur derivatives. Molecular structures and electrochemistry. Part V. {[Fe4S4](SγCys)4} proteins. Coord. Chem. Rev. 2017, 335, 172–227. [Google Scholar] [CrossRef]
- Zanello, P.; Corsini, M. The competition between chemistry and biology in assembling iron sulfur derivatives: Molecular structures and electrochemistry. Part VI. [Fe4-S4](SCys)3 (nonthiolate ligand) proteins. In Reference Module in Chemistry, Molecular Sciences and Chemical Engineering; Elsevier: Amsterdam, The Netherlands, 2014; Available online: https://www.sciencedirect.com/science/article/pii/B9780124095472127088 (accessed on 3 January 2022).
- Zanello, P. Structure and electrochemistry of proteins harboring iron-sulfur clusters of different nuclearities. Part I. [4Fe-4S] + [2Fe-2S] iron-sulfur proteins. J. Struct. Biol. 2017, 200, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Zanello, P. Structure and electrochemistry of proteins harboring iron-sulfur clusters of different nuclearities. Part II. [4Fe-4S] and [3Fe-4S] iron-sulfur proteins. J. Struct. Biol. 2018, 202, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Zanello, P. Structure and electrochemistry of proteins harboring iron-sulfur clusters of different nuclearities. Part III. [4Fe-4S] and [3Fe-4S] iron-sulfur proteins. J. Struct. Biol. 2018, 202, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Zanello, P. Structure and electrochemistry of proteins harboring iron-sulfur clusters of different nuclearities. Part IV. Canonical, non-canonical and hybrid iron-sulfur proteins. J. Struct. Biol. 2019, 205, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Zanello, P. Structure and electrochemistry of proteins harboring iron-sulfur clusters of different nuclearities. Part V. Nitrogenases. Coord. Chem. Rev. 2019, 398, 113004. [Google Scholar] [CrossRef]
- Bonfio, C.; Godino, E.; Corsini, M.; de Biani, F.F.; Guella, F.G.; Mansy, S.S. Prebiotic ironsulfur peptide catalysts generate a pH gradient across model membranes of late protocells. Nat. Catal. 2018, 1, 616–623. [Google Scholar] [CrossRef]
- Klenk, H.-P.; Clayton, R.A.; Tomb, J.-F.; White, O.; Nelson, K.E.; Ketchum, K.A.; Dodson, R.J.; Gwinn, M.; Hickey, E.K.; Peterson, J.D.; et al. The complete genome sequence of the hyperthermophilic, sulphate-reducing archaeon Archaeoglobus fulgidus. Nature 1997, 390, 591–594. [Google Scholar] [CrossRef]
- Birkeland, N.-K.; Schönheit, P.; Poghosyan, L.; Fiebig, A.; Klenk, H.-P. Complete genome sequence analysis of Archaeoglobus fulgidus strain 7324 (DSM 8774), a hyperthermophilic archaeal sulfate reducer from a North Sea oil field. Stand. Genomic Sci. 2017, 12, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGuirl, M.A.; Dooley, D.M. Copper-containing oxidases. Curr. Opin. Chem. Biol. 1999, 3, 138–144. [Google Scholar] [CrossRef]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82/83, 969–974. [Google Scholar] [CrossRef]
- Prousek, Fenton chemistry in biology and medicine. J. Pure Appl. Chem. 2007, 79, 2325–2338. [CrossRef]
- Harrison, M.D.; Jones, C.E.; Dameron, C.T. Copper chaperones: Function, structure and copper-binding properties. J. Biol. Inorg. Chem. 1999, 4, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Markossian, K.A.; Kurganov, B.I. Copper chaperones, intracellular copper trafficking proteins. Function, structure, and mechanism of action. Biochemistry 2003, 68, 827–837. [Google Scholar]
- Pham, A.N.; Xing, G.; Miller, C.J.; Waite, T.D. Fenton-like copper redox chemistry revisited: Hydrogen peroxide and superoxide mediation of copper-catalyzed oxidant production. J. Catal. 2013, 301, 54–64. [Google Scholar] [CrossRef]
- Kim, B.-E.; Nevitt, T.; Thiele, D.J. Mechanisms for copper acquisition, distribution and regulation. Nat. Chem. Biol. 2008, 4, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Macomber, L.; Imlay, J.A. The iron-sulfur clusters of dehydratases are primary intracellular targets of copper toxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 8344–8349. [Google Scholar] [CrossRef] [Green Version]
- Kühlbrandt, W. Biology, structure and mechanism of P-type ATPases. Nat. Rev. Mol. Cell Biol. 2004, 5, 282–295. [Google Scholar] [CrossRef]
- Palmgren, M.G.; Nissen, P. P-Type ATPases. Annu. Rev. Biophys. 2011, 40, 243–266. [Google Scholar] [CrossRef]
- Bublitz, M.; Morth, J.P.; Nissen, P. P-type ATPases at a glance. J. Cell Sci. 2012, 124, 2515–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inesi, G.; Pilankatta, R.; Tadini-Buoninsegni, F. Biochemical characterization of P-type copper ATPases. Biochem. J. 2014, 463, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumaa, P. Copper chaperones. The concept of conformational control in the metabolism of copper. FEBS Lett. 2013, 587, 1902–1910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Guerrero, M.; Argüello, J.M. Mechanism of Cu+-transporting ATPases: Soluble Cu+ chaperones directly transfer Cu+ to transmembrane transport sites. Proc. Natl. Acad. Sci. USA 2008, 105, 5992–5997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
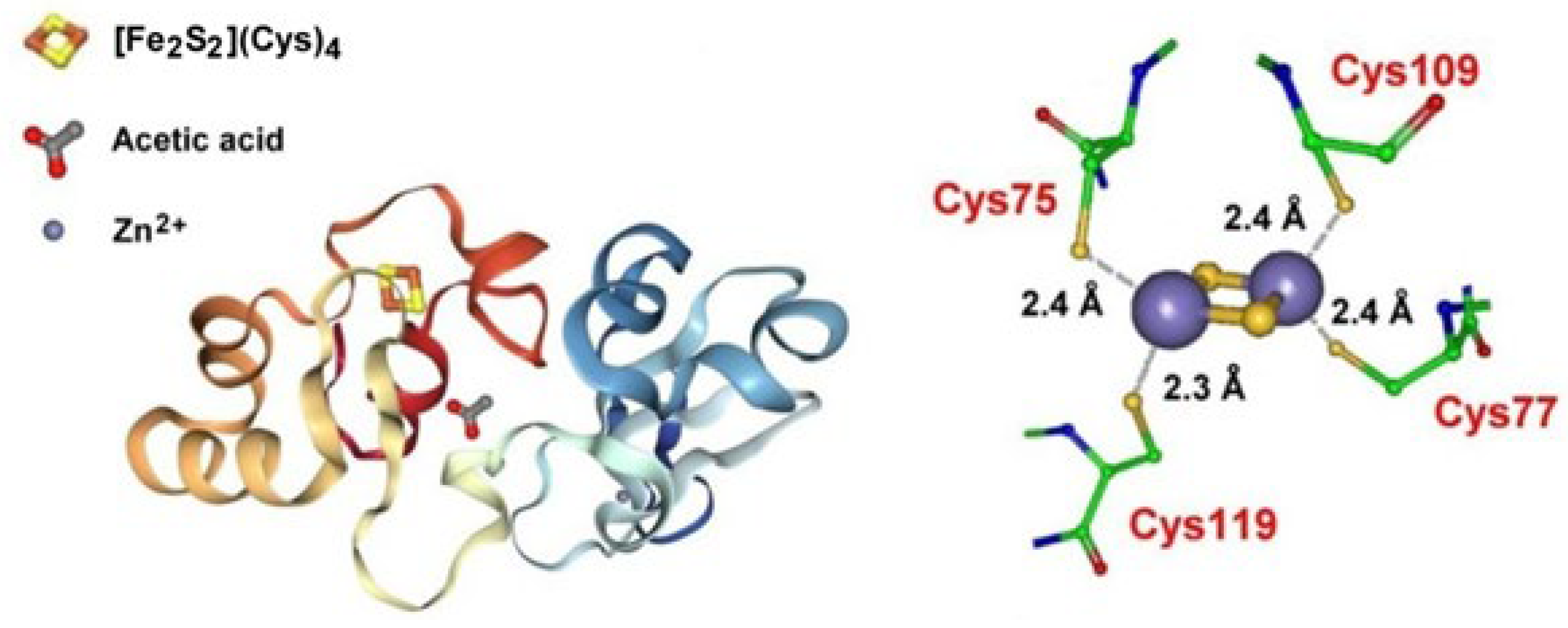
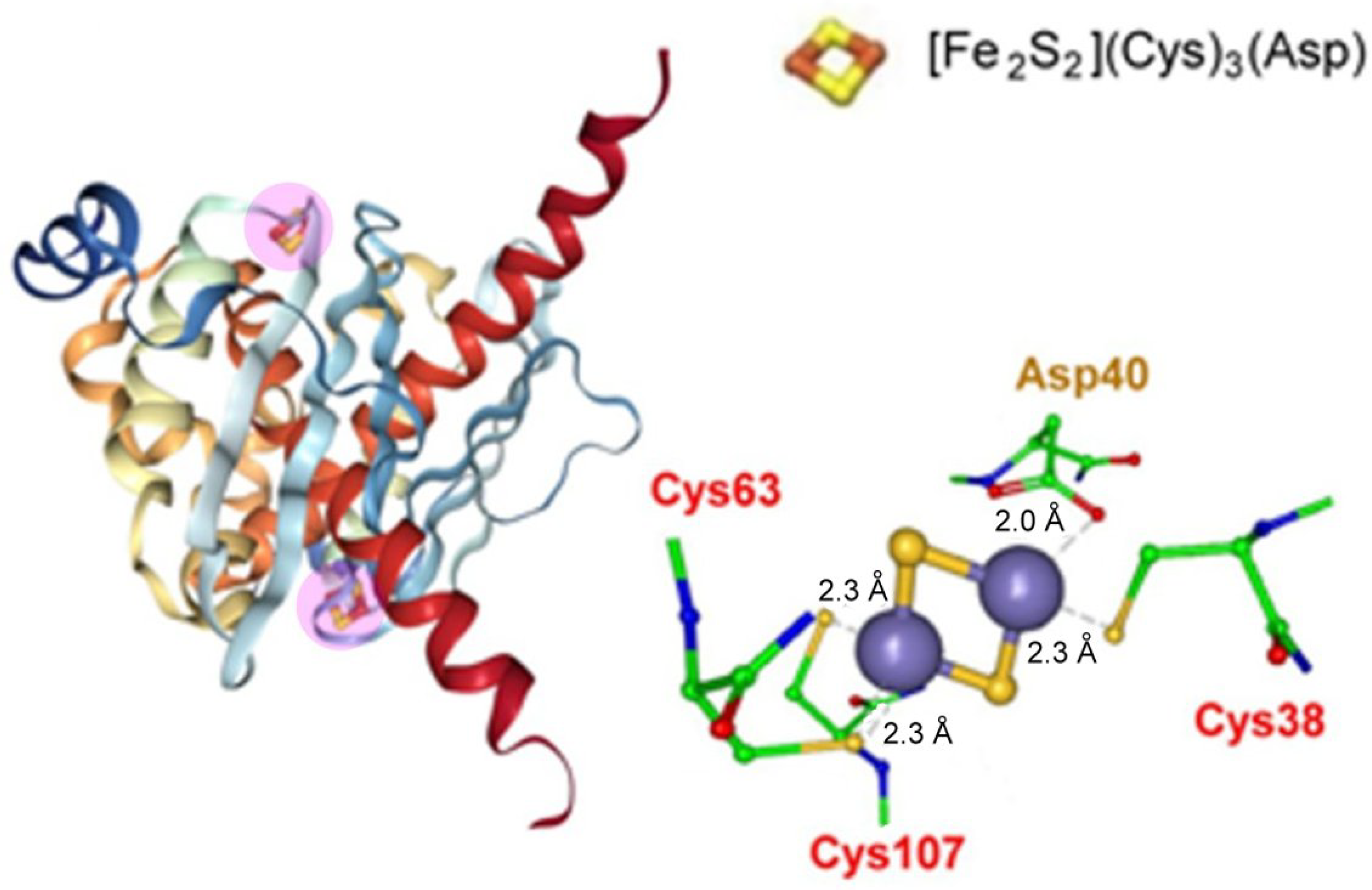
- Sazinsky, M.H.; LeMoine, B.; Orofino, M.; Davydov, R.; Bencze, K.Z.; Stemmler, T.L.; Hoffman, B.M.; Argüello, J.M.; Rosenzweig, A.C. Characterization and Structure of a Zn2 and [2Fe-2S]-containing Copper Chaperone from Archaeoglobus fulgidus. J. Biol. Chem. 2007, 282, 25950–25959. [Google Scholar] [CrossRef] [Green Version]
- Boal, A.K.; Rosenzweig, A.C. Structural Biology of Copper Trafficking. Chem. Rev. 2009, 109, 4760–4779. [Google Scholar] [CrossRef] [Green Version]
- Mayhew, S.G. The Redox Potential of Dithionite and SO-2 from Equilibrium Reactions with Flavodoxins, Methyl Viologen and Hydrogen plus Hydrogenase. Eur. J. Biochem. 1978, 85, 535–547. [Google Scholar] [CrossRef]
- Melber, A.; Winge, D.R. Steps Toward Understanding Mitochondrial Fe/S Cluster Biogenesis. Methods Enzymol. 2018, 599, 265–292. [Google Scholar]
- Frazzon, J.; Dean, D.R. Feedback regulation of iron-sulfur cluster biosynthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 14751–14753. [Google Scholar] [CrossRef] [Green Version]
- Mihara, H.; Esaki, N. Bacterial cysteine desulfurases: Their function and mechanisms. Appl. Microbiol. Biotechnol. 2002, 60, 12–23. [Google Scholar] [PubMed]
- Kato, S.-I.; Mihara, H.; Kurihara, T.; Takahashi, Y.; Tokumoto, U.; Yoshimura, T.; Esaki, N. The iscS gene is essential for the biosynthesis of 2-selenouridine in tRNA and the selenocysteine-containing formate dehydrogenase H. Proc. Natl. Acad. Sci. USA 2002, 99, 5948–5952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazzon, J.; Fickt, J.R.; Dean, D.R. Biosynthesis of iron-sulphur clusters is a complex and highly conserved process. Biochem. Soc. Trans. 2002, 30, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Frazzon, J.; Dean, D.R. Formation of iron-sulfur clusters in bacteria: An emerging field in bioinorganic chemistry. Curr. Opin. Chem. Biol. 2003, 7, 166–173. [Google Scholar] [CrossRef]
- Fontecave, M.; Ollagnier-de-Choudens, S. Iron-sulfur cluster biosynthesis in bacteria: Mechanisms of cluster assembly and transfer. Arch. Biochem. Biophys. 2008, 474, 226–237. [Google Scholar] [CrossRef]
- Ali, V.; Nozak, T. Iron–Sulphur Clusters, Their Biosynthesis, and Biological Functions in Protozoan Parasites. Adv. Parasitol. 2013, 83, 1–92. [Google Scholar]
- Roche, B.; Aussel, L.; Ezraty, B.; Mandin, P.; Py, B.; Barras, F. Iron/sulfur proteins biogenesis in prokaryotes: Formation, regulation and diversity. Biochim. Biophys. Acta 2013, 1827, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Kesawat, M.S.; Das, B.K.; Kumar, M.; Bhaganagare, G.R.; Manorama. An overview on Fe–S protein biogenesis from prokaryotes to eukaryotes. In Biological Nitrogen Fixation; de Bruijn, F.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; Volume 1, Chapter 6; pp. 57–74. [Google Scholar]
- Rocha, A.G.; Dancis, A. Life without Fe-S cluster. Mol. Microbiol. 2016, 99, 821–826. [Google Scholar] [CrossRef]
- Dos Santos, P.C. B. subtilis as a Model for Studying the Assembly of FeS Clusters in Gram-Positive Bacteria. Methods Enzymol. 2017, 595, 185–212. [Google Scholar] [PubMed]
- Braymer, J.J.; Lill, R. Iron sulfur cluster biogenesis and trafficking in mitochondria. J. Biol. Chem. 2017, 292, 12754–12763. [Google Scholar] [CrossRef] [Green Version]
- Boniecki, M.T.; Freibert, S.A.; Mühlenhoff, U.; Lill, R.; Cygler, M. Structure and functional dynamics of the mitochondrial Fe/S cluster synthesis complex. Nat. Commun. 2017, 8, 1287. [Google Scholar] [CrossRef] [Green Version]
- Di Maio, D.; Chandramouli, B.; Yan, R.; Brancato, G.; Pastore, A. Understanding the role of dynamics in the iron sulfur cluster molecular machine. Biochim. Biophys. Acta 2017, 1861, 3154–3163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardenas-Rodriguez, M.; Chatzi, A.; Tokatlidis, K. Iron sulfur clusters: From metals through mitochondria biogenesis to disease. J. Biol. Inorg. Chem. 2018, 23, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Dos Santos, P.C. Metallocluster transactions: Dynamic protein interactions guide the biosynthesis of FeS clusters in bacteria. Biochem. Soc. Trans. 2018, 46, 1593–1603. [Google Scholar] [CrossRef] [PubMed]
- Wachnowsky, C.; Fidai, I.; Cowan, J.A. Iron-sulfur cluster biosynthesis and trafficking-impact on human disease conditions. Metallomics 2018, 10, 9–29. [Google Scholar] [CrossRef] [PubMed]
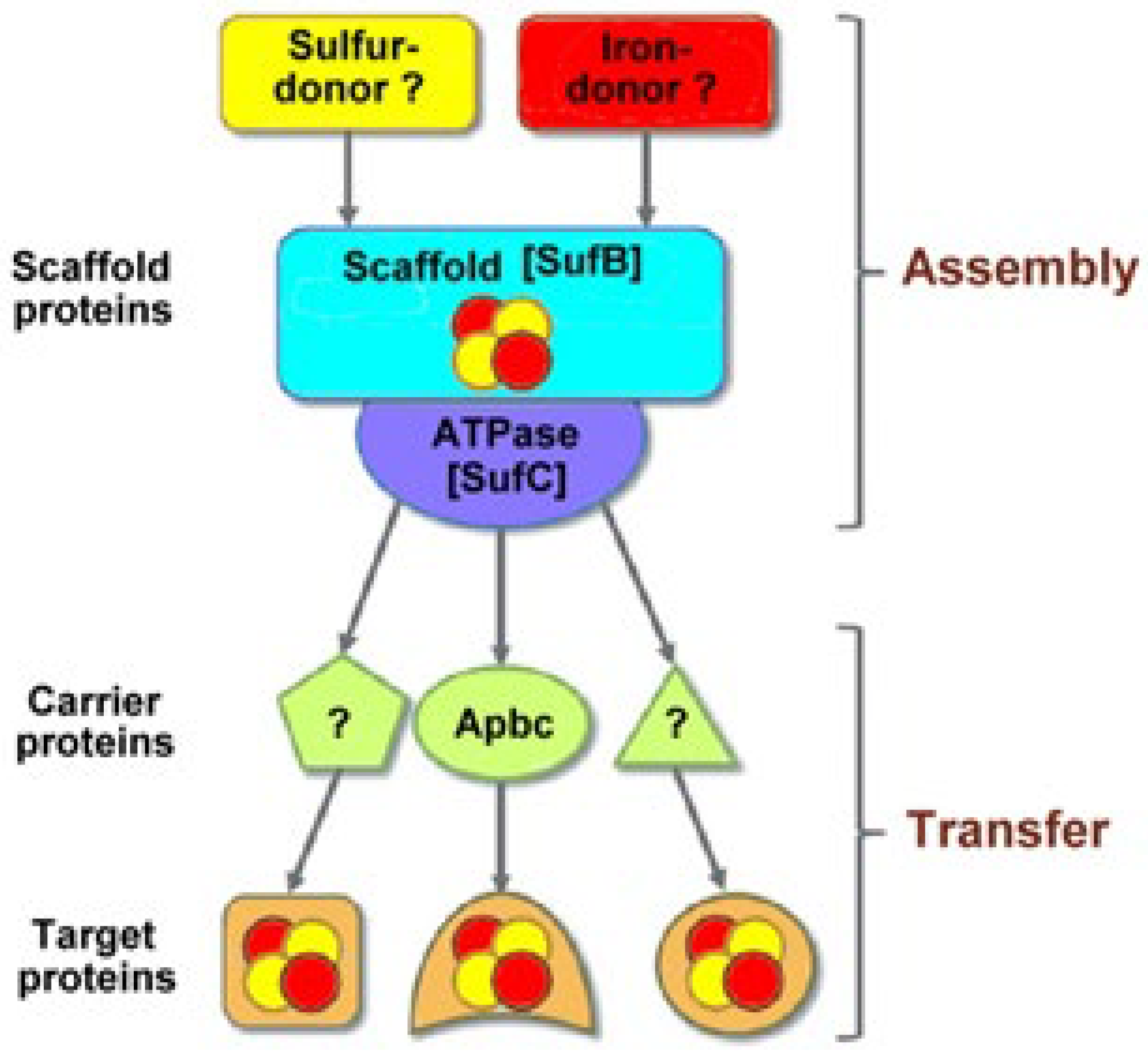
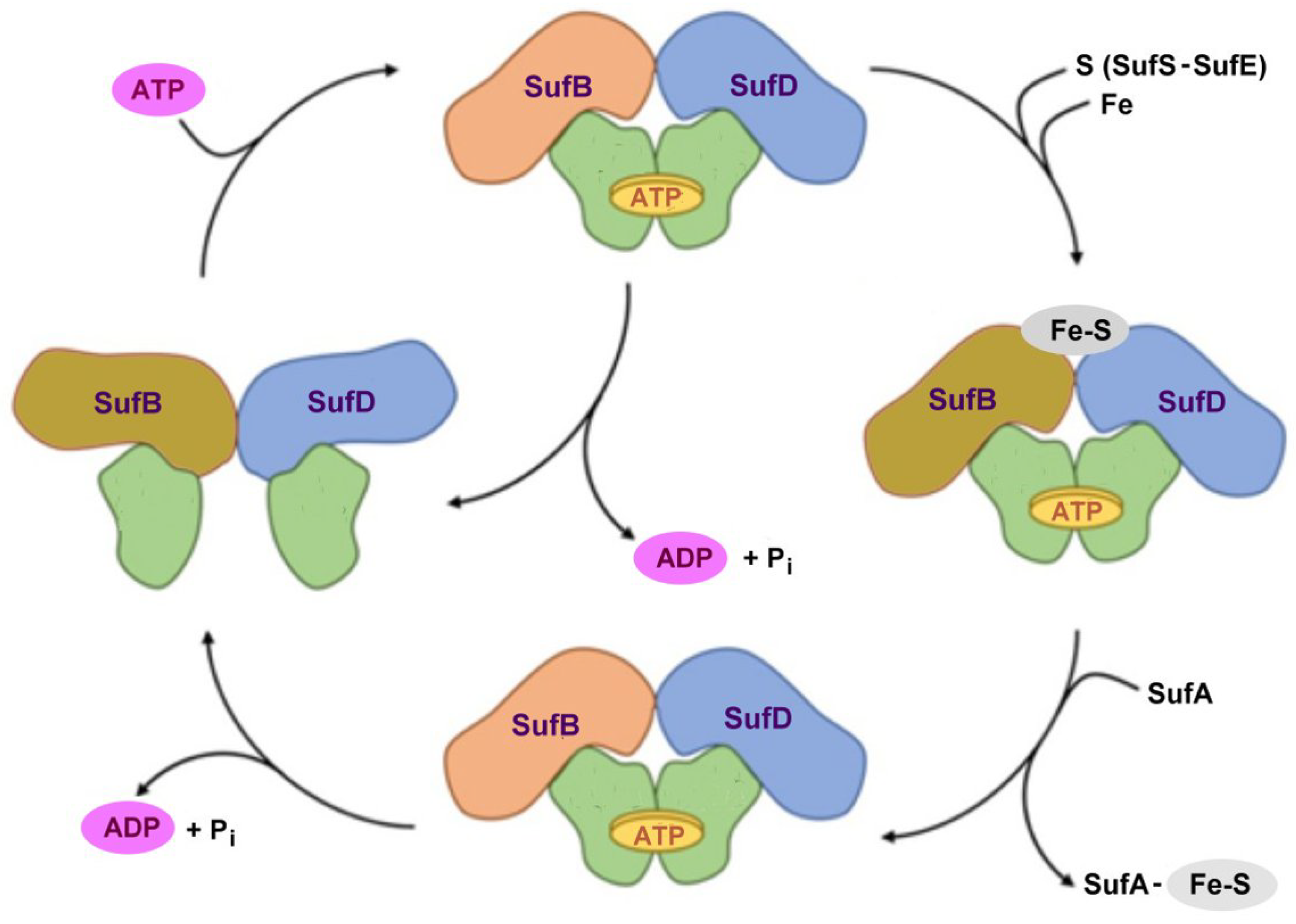
- Pérard, J.; Ollagnier de Choudens, S. Iron-sulfur clusters biogenesis by the SUF machinery: Close to the molecular mechanism understanding. J. Biol. Inorg. Chem. 2018, 23, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Chen, T.; Happe, T.; Lu, Y.; Sawyer, A. Iron–sulphur cluster biogenesis via the SUF pathway. Metallomics 2018, 10, 1038–1052. [Google Scholar] [CrossRef] [PubMed]
- Yuda, E.; Tanaka, N.; Fujishiro, T.; Yokoyama, N.; Hirabayashi, K.; Fukuyama, K.; Wada, K.; Takahashi, Y. Mapping the key residues of SufB and SufD essential for biosynthesis of iron-sulfur clusters. Sci. Rep. 2017, 7, 9387. [Google Scholar] [CrossRef] [Green Version]
- Nicolet, Y.; Fontecilla-Camps, J.C. Fe-S clusters: Biogenesis and redox, catalytic, and regulatory properties. In Bioinspired Catalysis: Metal-Sulfur Complexes; Weigand, W., Schollhammer, P., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2015. [Google Scholar]
- Srour, B.; Gervason, S.; Monfort, B.; D’Autréaux, B. Mechanism of Iron–Sulfur Cluster Assembly: In the Intimacy of Iron and Sulfur Encounter. Inorganics 2020, 8, 55. [Google Scholar] [CrossRef]
- Takahashi, Y.; Nakamura, M. Functional Assignment of the ORF2-iscS-iscU-iscA-hscB-hscA-fdx-0RF3 Gene Cluster Involved in the Assembly of Fe-S Clusters. J. Biochem. 1999, 126, 917–926. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.-H.; Huynh, K.-H.; Nguyen, D.Q.; Park, H.; Jung, K.; Sur, B.; Ahn, Y.-J.; Cha, S.-S.; Kang, L.-W. Catalytic Intermediate Crystal Structures of Cysteine Desulfurase from the Archaeon Thermococcus onnurineus NA1. Archaea 2017, 2017, 5395293. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sieprawska-Lupa, M.; Whitman, W.B.; White, R.H. Cysteine Is Not the Sulfur Source for Iron-Sulfur Cluster and Methionine Biosynthesis in the Methanogenic Archaeon Methanococcus maripaludis. J. Biol. Chem. 2010, 285, 31923–31949. [Google Scholar] [CrossRef] [Green Version]
- Boyd, E.S.; Thomas, K.M.; Dai, Y.; Boyd, J.M.; Outten, F.W. Interplay between Oxygen and Fe-S Cluster Biogenesis: Insights from the Suf Pathway. Biochemistry 2014, 53, 5834–5847. [Google Scholar] [CrossRef] [Green Version]
- Boyd, J.M.; Drevland, R.M.; Downs, D.M.; Graham, D.E. Archaeal ApbC/Nbp35 Homologs Function as Iron-Sulfur Cluster Carrier Proteins. J. Bacteriol. 2009, 191, 1490–1497. [Google Scholar] [CrossRef] [Green Version]
- Garcia, P.S.; Gribaldo, S.; Py, B.; Barras, F. The SUF system: An ABC ATPase-dependent protein complex with a role in Fe-S cluster biogenesis. Res. Microbiol. 2019, 170, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Beer, L.L.; Whitman, W.B. Sulfur metabolism in archaea reveals novel processes. Environ. Microbiol. 2012, 14, 2632–2644. [Google Scholar] [CrossRef] [PubMed]
- Boyd, J.M.; Pierik, A.J.; Netz, D.J.A.; Lill, R.; Downs, D.M. Bacterial {ApbC} Can Bind and Effectively Transfer Iron-Sulfur Clusters. Biochemistry 2008, 47, 8195–8202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, J.M.; Sondelski, J.L.; Downs, D.M. Bacterial ApbC Protein Has Two Biochemical Activities That Are Required for in Vivo Function. J. Biol. Chem. 2009, 284, 110–118. [Google Scholar] [CrossRef] [Green Version]
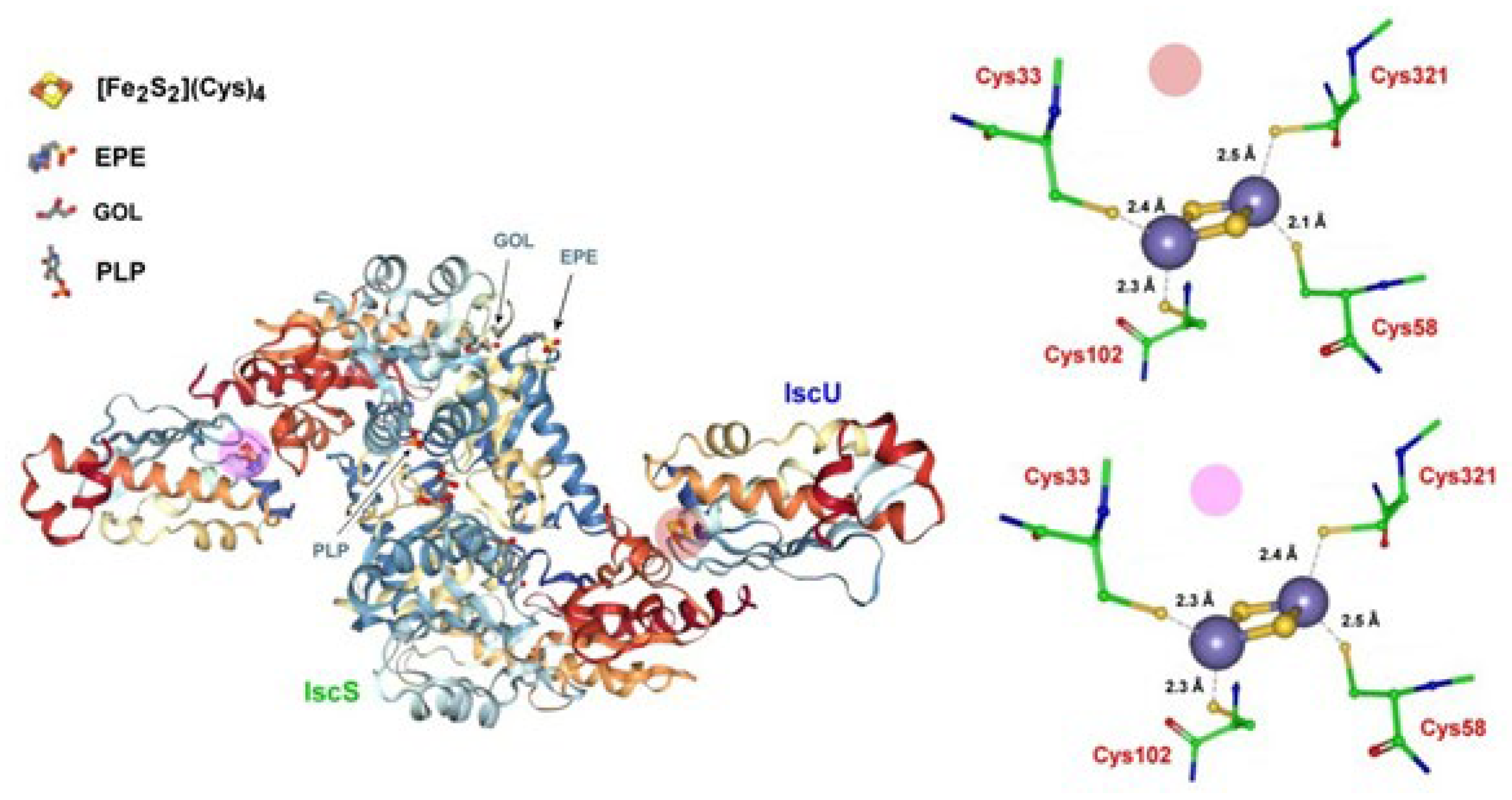
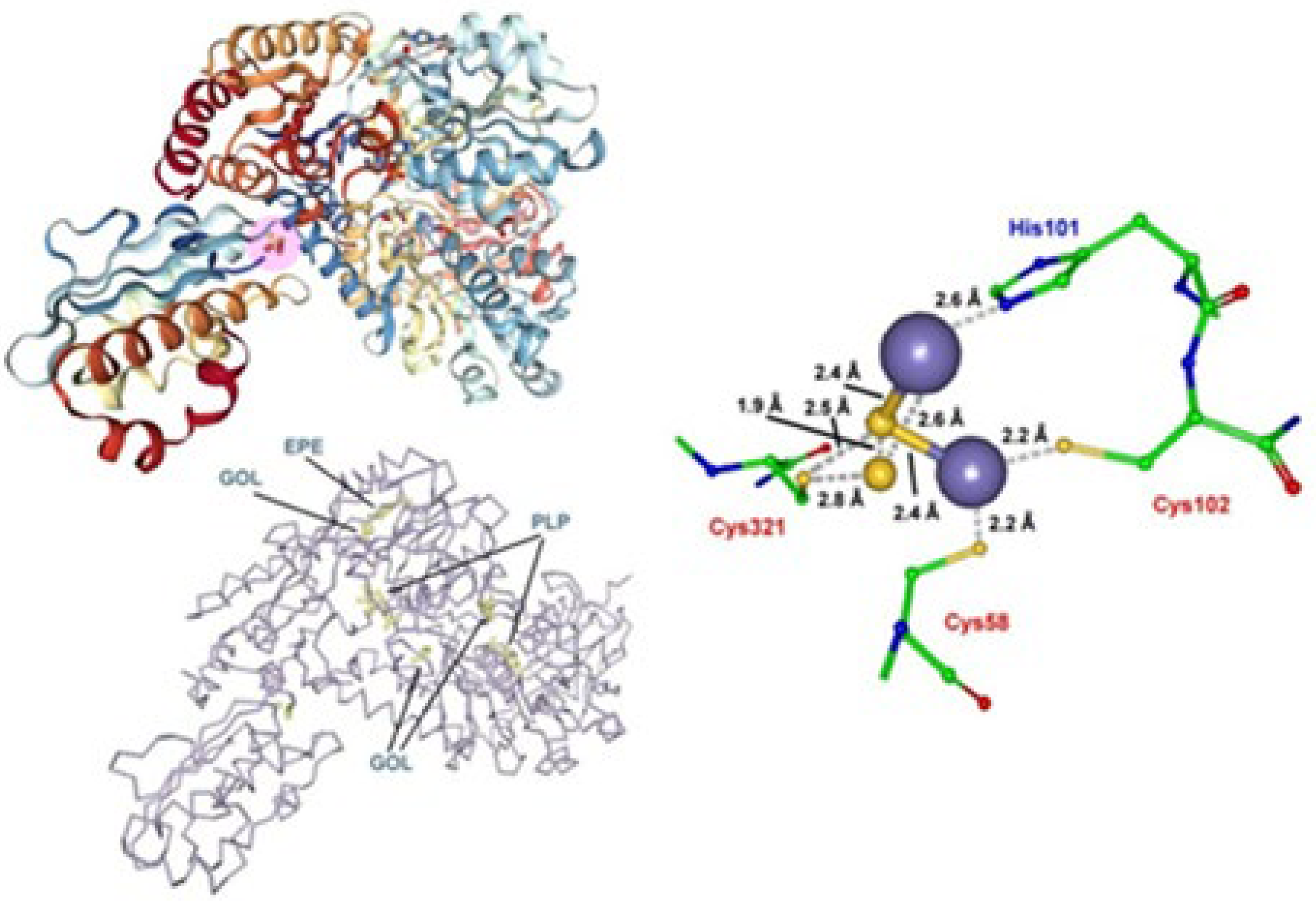
- Marinoni, E.N.; de Oliveira, J.S.; Nicolet, Y.; Raulfs, E.C.; Amara, P.; Dean, D.R.; Fontecilla-Camps, J.C. (IscS-IscU)2 Complex Structures Provide Insights into Fe2S2 Biogenesis and Transfer. Angew. Chem. Int. Ed. 2012, 51, 5439–5442. [Google Scholar] [CrossRef] [PubMed]
- Raulfs, E.C.; O’Carroll, I.P.; Dos Santos, P.C.; Unciuleac, M.-C.; Dean, D.R. In vivo iron-sulfur cluster formation. Proc. Natl. Acad. Sci. USA 2008, 105, 8591–8596. [Google Scholar] [CrossRef] [Green Version]
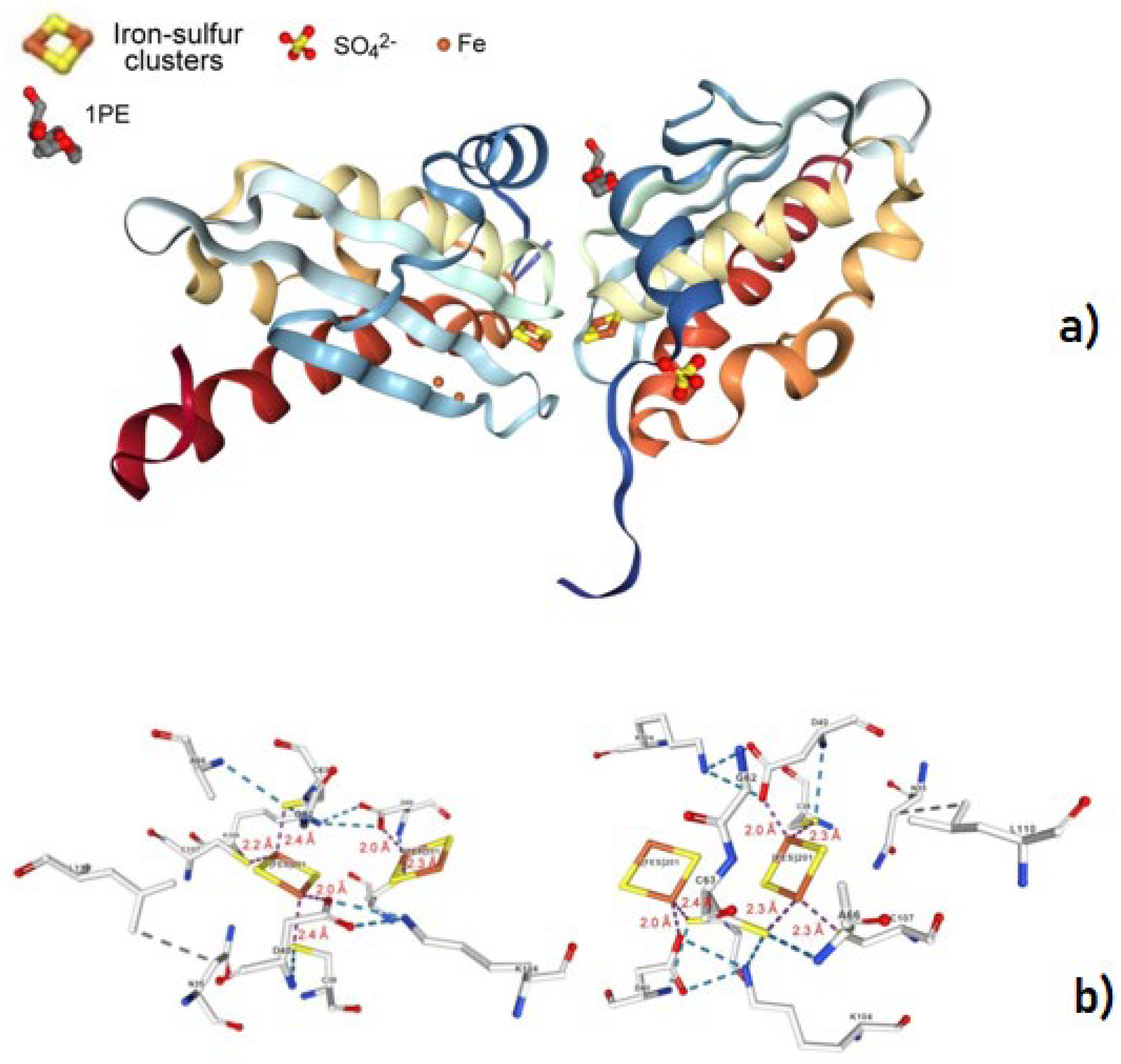
- Yamanaka, Y.; Zeppieri, L.; Nicolet, Y.; Marinoni, E.N.; de Oliveira, J.S.; Odaka, M.; Dean, D.R.; Fontecilla-Camps, J.C. Crystal structure and functional studies of an unusuall-cysteine desulfurase from Archaeoglobus fulgidus. Dalton Trans. 2013, 42, 3092–3099. [Google Scholar] [CrossRef] [PubMed]
- Pagnier, A.; Nicolet, Y.; Fontecilla-Camps, J.C. IscS from Archaeoglobus fulgidus has no desulfurase activity but may provide a cysteine ligand for [Fe2S2] cluster assembly. Biochim. Biophys. Acta 2015, 1853, 1457–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nývltová, E.; Suták, R.; Harant, K.; Sedinová, M.; Hrdý Jan Pačes, I.; Vlčk, Č.; Tachezy, J. NIF-type iron-sulfur cluster assembly system is duplicated and distributed in the mitochondria and cytosol of Mastigamoeba balamuthi. Proc. Natl. Acad. Sci. USA 2013, 110, 7371–7376. [Google Scholar] [CrossRef] [Green Version]
- Bell, S.D.; Jackson, S.P. Mechanism and regulation of transcription in archaea. Curr. Opin. Microbiol. 2001, 4, 208–213. [Google Scholar] [CrossRef]
- Brochier, C.; Forterre, P.; Gribaldo, S. An emerging phylogenetic core of Archaea: Phylogenies of transcription and translation machineries converge following addition of new genome sequences. BMC Evol. Biol. 2005, 5, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barry, E.R.; Bell, S.D. DNA Replication in the Archaea. Microbiol. Mol. Biol. Rev. 2006, 70, 876–887. [Google Scholar] [CrossRef] [Green Version]
- Greci, M.D.; Bell, S.D. Archaeal DNA Replication. Annu. Rev. Microbiol. 2020, 74, 65–80. [Google Scholar] [CrossRef]
- Lindås, A.C.; Karlsson, E.A.; Lindgren, M.T.; Ettema, T.J.G.; Bernander, R. A unique cell division machinery in the Archaea. Proc. Natl. Acad. Sci. USA 2008, 1051, 18942–18946. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, T. Iron-Sulfur World in Aerobic and Hyperthermoacidophilic Archaea Sulfolobus. Archaea 2010, 2010, 842639. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, M.; Langston, L.; Stillman, B. Principles and concepts of DNA replication in bacteria, archaea, and bacteria. Cold Spring Harb. Perspect. Biol. 2013, 5, a010108. [Google Scholar]
- Han, W.; Shen, Y.; She, Q. Nanobiomotors of archaeal DNA repair machineries: Current research status and application potential. Cell Biosci. 2014, 4, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phung, D.K.; Etienne, C.; Batista, M.; Langendiik-Genevaux, P.; Moalic, Y.; Laurent, S.; Liu, S.; Morales, V.; Jebbar, M.; Fichant, G.; et al. RNA processing machineries in Archaea: The 5′-3′ exoribonuclease aRNase J of the β-CASP family is engaged specifically with the helicase ASH-Ski2 and the 3′-5′ exoribonucleolytic RNA exosome machinery. Nucleic Acid Res. 2020, 48, 3832–3847. [Google Scholar] [CrossRef] [PubMed]
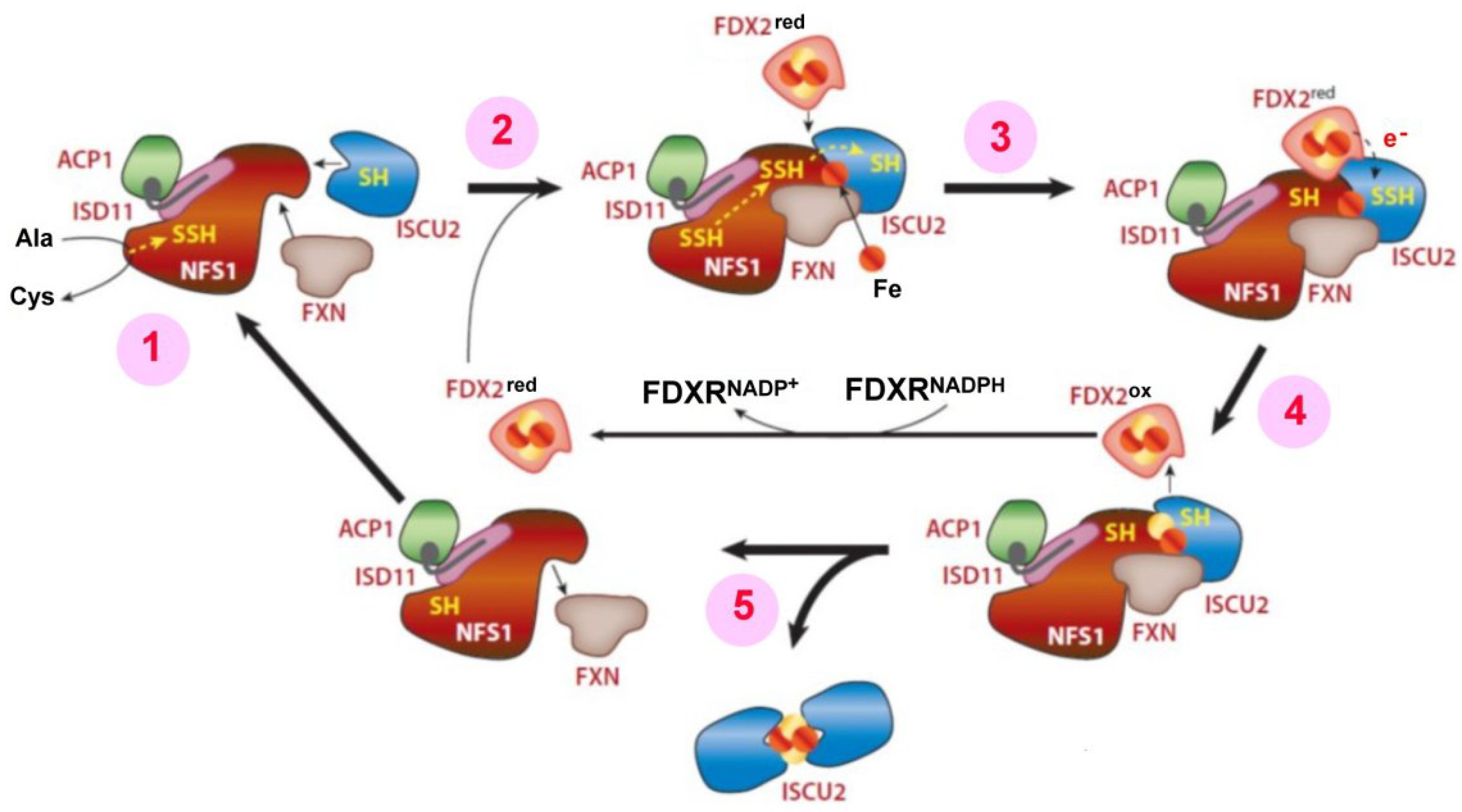
- Lill, R.; Freibert, S.-A. Mechanisms of Mitochondrial Iron-Sulfur Protein Biogenesis. Annu. Rev. Biochem. 2020, 89, 471–499. [Google Scholar] [CrossRef] [PubMed]
- Weiler, B.D.; Brück, M.-C.; Kothe, I.; Bill, E.; Lill, R.; Mühlenhoff, U. Mitochondrial [4Fe-4S] protein assembly involves reductive [2Fe-2S] cluster fusion on ISCA1-ISCA2 by electron flow from ferredoxin FDX2. Proc. Natl. Acad. Sci. USA 2020, 117, 20555–20565. [Google Scholar] [CrossRef] [PubMed]
- Lill, R. Function and biogenesis of iron-sulphur proteins. Nature 2009, 460, 831–838. [Google Scholar] [CrossRef]
- Nozhevnikova, A.N.; Chudina, V.I. Morphology of the thermophilic acetate bacterium Methanothrix thermoacetophila. Mikrobiologiia 1984, 53, 756–760. [Google Scholar]
- Boone, D.R.; Kamagata, Y.J. Rejection of the species Methanothrix soehngenii VP and the genus Methanothrix VP as nomina confusa, and transfer of Methanothrix thermophila VP to the genus Methanosaeta VP as Methanosaeta thermophila comb. nov. Request for an Opinion. Syst. Bacteriol. 1998, 48, 1079–1080. [Google Scholar] [CrossRef] [Green Version]
- Methanothrix thermoacetophila PT strain: PT. Available online: https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA15765 (accessed on 3 January 2022).
- Methanothrix thermoacetophila (strain DSM 6194/JCM 14653/NBRC 101360/PT) (Methanothrix thermoacetophila). Available online: https://www.uniprot.org/proteomes/UP000000674 (accessed on 3 January 2022).
- Kunichika, K.; Nakamura, R.; Fujishiro, T.; Takahashi, Y. The Structure of the Dimeric State of IscU Harboring Two Adjacent [2Fe-2S] Clusters Provides Mechanistic Insights into Cluster Conversion to [4Fe-4S]. Biochemistry 2021, 60, 1569–1572. [Google Scholar] [CrossRef] [PubMed]
- Kunichika, K.; Fujishiro, T.; Takahashi, Y. Crystal Structure of IscU H106C Variant. Available online: https://www.rcsb.org/structure/7CNV (accessed on 3 January 2022).
- Amo, T.; Luz, M.; Paje, F.; Inagaki, A.; Ezaki, S.; Atomi, H.; Imanaka, T. Pyrobaculum calidifontissp. nov., a novel hyperthermophilic archaeon that grows in atmospheric air. Archaea 2002, 1, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Lemak, S. Structural and Biochemical Characterization of CRISPR-Associated Cas4 Nucleases from a Prokaryotic Defense System. Master’s Thesis, Department of Chemical Engineering and Applied Chemistry, University of Toronto, Toronto, ON, Canada, 2013; p. 866. [Google Scholar]
- Zhang, J.; Kasciukovic, T.; White, M.F. The CRISPR Associated Protein Cas4 Is a 5’ to 3’ DNA Exonuclease with an Iron-Sulfur Cluster. PLoS ONE 2012, 7, e47232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shmakova, S.A.; Makarova, K.S.; Wolf, Y.I.; Severinova, K.V.; Koonin, E.V. Systematic prediction of genes functionally linked to CRISPR-Cas systems by gene neighborhood analysis. Proc. Natl. Acad. Sci. USA 2018, 115, E5307–E5316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarov, K.S.; Wolf, Y.I.; Koonin, E.V. Classification and Nomenclature of CRISPR-Cas Systems: Where from Here? CRISPR J. 2018, 1, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Makarova, K.S. Origins and evolution of CRISPR-Cas systems. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20180087. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR-Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Burstein, D.; Harrington, L.B.; Strutt, S.C.; Probst, A.J.; Anantharaman, K.; Thomas, B.C.; Doudna, J.A.; Banfield, J.F. New CRISPR-Cas systems from uncultivated microbes. Nature 2017, 542, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Lemak, S.; Nocek, B.; Beloglazova, N.; Skarina, T.; Flick, R.; Brown, G.; Joachimiak, A.; Savchenko, A.; Yakunin, A.F. The CRISPR-associated Cas4 protein Pcal_0546 from Pyrobaculum calidifontis contains a [2Fe-2S] cluster: Crystal structure and nuclease activity. Nucleic Acids Res. 2014, 42, 11144–11155. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Zhang, L.; Lai, S.; Ye, K. Structure and Molecular Evolution of CDGSH Iron-Sulfur Domains. PLoS ONE 2011, 6, e24790. [Google Scholar] [CrossRef]
- Sengupta, S.; Nechushtai, R.; Jennings, P.A.; Onuchic, J.N.; Padilla, P.A.; Azad, R.K.; Mittler, R. Phylogenetic analysis of the CDGSH iron-sulfur binding domain reveals its ancient origin. Sci. Rep. 2018, 8, 4840. [Google Scholar] [CrossRef]
- González, J.M.; Masuchi, Y.; Robb, F.T.; Ammerman, J.W.; Maeder, D.L.; Yanagibayashi, M.; Tamaoka, J.; Kato, C. Pyrococcus horikoshii sp. nov., a hyperthermophilic archaeon isolated from a hydrothermal vent at the Okinawa Trough. Extremophiles 1998, 2, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Kawarbayasi, Y.; Sawada, M.; Horikawa, H.; Haikawa, Y.; Hino, Y.; Yamamoto, S.; Sekine, M.; Baba, S.-I.; Kosugi, H.; Hosoyama, A.; et al. Complete Sequence and Gene Organization of the Genome of a Hyper-thermophilic Archaebacterium, Pyrococcus horikoshii OT3 (Supplement). DNA Res. 1998, 5, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Shigi, N. Recent Advances in Our Understanding of the Biosynthesis of Sulfur Modifications in tRNAs. Front. Microbiol. 2018, 9, 2679. [Google Scholar] [CrossRef] [PubMed]
- ttuA-tRNA-5-methyluridine(54) 2-sulfurtransferase—Pyrococcus horikoshii (Strain ATCC 700860/DSM 12428/JCM 9974/NBRC 100139/OT-3)—ttuA Gene & Protein. Available online: https://www.uniprot.org/uniprot/O58038 (accessed on 26 June 2021).
- Arragain, S.; Bimai, O.; Legrand, P.; Caillat, S.; Ravanat, J.-L.; Touati, N.; Binet, L.; Atta, M.; Fontecave, M. Nonredox thiolation in tRNA occurring via sulfur activation by a [4Fe-4S] cluster. Proc. Natl. Acad. Sci. USA 2017, 114, 7355–7360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darland, G.; Brock, T.D. Bacillus acidocaldarius sp. nov., an Acidophilic Thermophilic Spore-forming Bacterium. Gen. J. Microbiol. 1971, 67, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Brock, T.D.; Brock, K.M.; Belli, R.T.; Weiss, R.L. Sulfolobus: A new genus of sulfur-oxidizing bacteria living at low pH and high temperature. Arch. Microbiol. 1972, 84, 54–68. [Google Scholar] [CrossRef]
- Moll, R.; Schäfer, G. Chemiosmotic H+ cycling across the plasma membrane of the thermoacidophilic archaebacterium Sulfolobus acidocaldarius. FEBS Lett. 1988, 232, 359–363. [Google Scholar] [CrossRef] [Green Version]
- Lübben, M.; Schäfer, G. Chemiosmotic energy conversion of the archaebacterial thermoacidophile Sulfolobus acidocaldarius: Oxidative phosphorylation and the presence of an F0-related N,N’-dicyclohexylcarbodiimide-binding proteolipid. J. Bacteriol. 1989, 171, 6106–6116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anemüller, S.; Lübben, M.; Schäfer, G. The respiratory system of Sulfolobus acidocaldarius, a thermoacidophilic archaebacterium. FEBS Lett. 1985, 193, 83–87. [Google Scholar] [CrossRef]
- Castresana, J.; Lübben, M.; Saraste, M.J. New Archaebacterial Genes Coding for Redox Proteins: Implications for the Evolution of Aerobic Metabolism. Mol. Biol. 1995, 250, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Gleißner, M.; Kaiser, U.; Antonopoulos, E.; Schäfer, G. The Archaeal SoxABCD Complex Is a Proton Pump in Sulfolobus acidocaldarius. J. Biol. Chem. 1997, 272, 8417–8426. [Google Scholar] [CrossRef] [Green Version]
- Lübben, M.; Warne, A.; Albracht, S.P.J.; Saraste, M. The purified SoxABCD quinol oxidase complex of Sulfolobus acidocaldarius contains a novel haem. Mol. Microbiol. 1994, 13, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Lübben, M.; Morand, K. Novel prenylated hemes as cofactors of cytochrome oxidases. Archaea have modified hemes A and O. J. Biol. Chem. 1994, 269, 21473–21479. [Google Scholar] [CrossRef]
- Schäfer, G.; Pursckhe, W.; Schmidt, C.L. On the origin of respiration: Electron transport proteins from archaea to man. FEMS Microbiol. Rev. 1996, 18, 173–188. [Google Scholar] [CrossRef]
- Lappalainnen, P.; Sarraste, M. The binuclear CuA centre of cytochrome oxidase. Biochim. Biophys. Acta 1994, 1187, 222–225. [Google Scholar] [CrossRef]
- Komorowski, L.; Verheyen, W.; Schäfer, G. The Archaeal Respiratory Supercomplex SoxM from S. acidocaldarius Combines Features of Quinole and Cytochrome c Oxidases. Biol. Chem. 2002, 383, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Bönisch, H.; Schmidt, C.L.; Schäfer, G.; Ladenstein, R. The Structure of the Soluble Domain of an Archaeal Rieske Iron-Sulfur Protein at 1.1 Å Resolution. J. Mol. Biol. 2002, 319, 791–805. [Google Scholar] [CrossRef]
- Schäfer, G.; Engelhard, M.; Müller, V. Bioenergetics of the Archaea. Microbiol. Mol. Biol. Rev. 1999, 63, 570–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komorowski, L.; Schäfer, G. Sulfocyanin and subunit II, two copper proteins with novel features, provide new insight into the archaeal SoxM oxidase supercomplex. FEBS Lett. 2001, 487, 351–355. [Google Scholar] [CrossRef] [Green Version]
- Anemüller, S.; Schmidt, C.L.; Schäfer, G.; Teixeira, M. Evidence for a Rieske-type FeS center in the thermoacidophilic archaebacterium Sulfolobus acidocaldarius. FEBS Lett. 1993, 318, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Anemüller, S.; Schmidt, C.L.; Schäfer, G.; Bill, E.; Trautwein, A.X.; Teixeira, M. Evidence for a Two-Proton-Dependent Redox Equilibrium in an Archaeal Rieske Iron-Sulfur Cluster. Biochem. Biophys Res. Commun. 1994, 202, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.L.; Hatzfeld, O.M.; Petersen, A.; Link, T.; Schäfer, G. Expression of the Solfolobus acidocaldarius Rieske Iron Sulfur Protein II (SOXF) with the Correctly Inserted [2Fe-2S] Cluster in Escherichia coli. Biochem. Biophys Res. Commun. 1997, 234, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Komorowski, L.; Anemüller, S.; Schäfer, G. Expression and Characterization of a Recombinant CuA-Containing Subunit II from an Archaeal Terminal Oxidase Complex. J. Bioenerg. Biomembr. 2001, 33, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Anemüller, S.; Bill, E.; Schäfer, G.; Trautwein, A.X.; Teixeira, M. EPR studies of cytochrome aa3 from Sulfolobus acidocaldarius. Evidence for a binuclear center in archaebacterial terminal oxidase. Eur. J. Biochem. 1992, 210, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, G.; Anemüller, S.; Moll, R.; Gleissner, M.; Schmidt, C.L. Has Sulfolobus an Archaic Respiratory System? Structure, Function and Genes of its Components. System. Appl. Microbiol. 1993, 16, 544–555. [Google Scholar] [CrossRef]
- Schäfer, G.; Moll, R.; Schmidt, C.L. Respiratory enzymes from Sulfolabus acidocaldarius. Methods Enzymol. 2001, 331, 369–410. [Google Scholar]
- Suzuki, T.; Iwasaki, T.; Uzawa, T.; Hara, K.; Nemoto, N.; Kon, T.; Ueki, T.; Yamagishi, A.; Oshima, T. Sulfolobus tokodaii sp. nov. (f. Sulfolobus sp. strain 7), a new member of the genus Sulfolobus isolated from Beppu Hot Springs, Japan. Extremophiles 2002, 6, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Wakagi, T.; Nishimasu, H.; Miyake, M.; Fushinobu, S. Archaeal Mo-Containing Glyceraldehyde Oxidoreductase Isozymes Exhibit Diverse Substrate Specificities through Unique Subunit Assemblies. S. PLoS ONE 2016, 11, e0147333. [Google Scholar] [CrossRef] [PubMed]
- Bräsen, C.; Esser, D.; Rauch, B.; Siebers, B. Carbohydrate Metabolism in Archaea: Current Insights into Unusual Enzymes and Pathways and Their Regulation. Microbiol. Mol. Biol. Rev. 2014, 78, 89–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Atomi, H. Novel metabolic pathways in Archaea. Curr. Opin. Microbiol. 2011, 14, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Kardinahl, S.; Schmidt, C.L.; Hansen, T.; Anemüller, S.; Petersen, A.; Schäfer, G. The strict molybdate-dependence of glucose-degradation by the thermoacidophile Sulfolobus acidocaldariusreveals the first crenarchaeotic molybdenum containing enzyme—An aldehyde oxidoreductase. Eur. J. Biochem. 1999, 260, 540–548. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, J. A new species of desulfovibrio. J. Bacteriol. 1963, 86, 1120. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, J.M.; Dias, J.M.; Huber, R.; Moura, J.J.G.; Romão, M.J. Structure refinement of the aldehyde oxidoreductase from Desulfovibrio gigas (MOP) at 1.28 Å. J. Biol. Inorg. Chem. 2001, 6, 791–800. [Google Scholar] [CrossRef] [PubMed]
- Correia dos Santos, M.M.; Sousa, P.M.P.; Gonçalves, M.L.S.; Romão, M.J.; Moura, I.; Moura, J.J.G. Direct electrochemistry of the Desulfovibrio gigas aldehyde oxidoreductase. Eur. J. Biochem. 2004, 271, 1329–1338. [Google Scholar] [CrossRef] [PubMed]
- Feio, M.J.; Zinkevich, V.; Beech, I.B.; Llobet-Brossa, E.; Eaton, P.; Schmitt, J.; Guezennec, J. Desulfovibrio alaskensis sp. nov., a sulphate-reducing bacterium from a soured oil reservoir. Int. J. Syst. Evol. Microbiol. 2004, 54, 1747–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, S.L.A.; Brondino, C.D.; Feio, M.J.; Moura, I.; Moura, J.J.G. Aldehyde oxidoreductase activity in Desulfovibrio alaskensis NCIMB 13491. Eur. J. Biochem. 2000, 267, 2054–2061. [Google Scholar] [CrossRef]
- Romão, M.J.; Coelho, C.; Santos-Silva, T.; Foti, A.; Terao, M.; Garattini, E.; Leimkühler, S. Structural basis for the role of mammalian aldehyde oxidases in the metabolism of drugs and xenobiotics. Curr. Opin. Chem. Biol. 2017, 37, 39–47. [Google Scholar] [CrossRef]
- Coelho, C.; Mahro, M.; Trincão, J.; Carvalho, A.T.P.; Ramos, M.J.; Terao, M.; Garattini, E.; Leimkühler, S.; Romão, M.J. The First Mammalian Aldehyde Oxidase Crystal Structure. J. Biol. Chem. 2012, 287, 40690–40702. [Google Scholar] [CrossRef] [Green Version]
- Mahro, M.; Brás, N.F.; Cerqueira, N.M.F.S.A.; Teutloff, C.; Coelho, C.; Romão, M.J.; Leimkühler, S. Identification of Crucial Amino Acids in Mouse Aldehyde Oxidase 3 That Determine Substrate Specificity. PLoS ONE 2013, 8, e82285. [Google Scholar] [CrossRef] [Green Version]
- Iwasaki, T.; Imaii, T.; Urushiyama, A.; Oshima, T. Redox-linked Ionization of Sulredoxin, an Archaeal Rieske-type [2Fe-2S] Protein from Sulfolobus sp. Strain 7. J. Biol. Chem. 1996, 271, 27659–27663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, T.; Kounosu, A.; Dikanov, S.A. The [2Fe-2S] cluster in sulredoxin from the thermoacidophilic Sulfolobus tokodaii strain 7, a novel water-soluble Rieske-type protein. In EPR in the 21st Century; Kawamori, A., Yamauchi, J., Ohta, H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2002; pp. 488–493. [Google Scholar]
- Kounosu, A.; Li, Z.; Cosper, N.J.; Shokes, J.E.; Scott, R.A.; Imai, T.; Urushiyama, A.; Iwasaki, T. Engineering a Three-cysteine, One-histidine Ligand Environment into a New Hyperthermophilic Archaeal Rieske-type [2Fe-2S] Ferredoxin from Sulfolobus solfataricus. J. Biol. Chem. 2004, 279, 12519–12528. [Google Scholar] [CrossRef] [Green Version]
- Bandeiras, T.M.; Freitas, M.C.; Petrasch, D.; Kletzin, A.; Frazão, C. SAD phasing towards structure determination of a thermostable Rieske ferredoxin with a novel stabilizing disulfide bridge. Acta Cryst. 2013, F69, 555–558. [Google Scholar] [CrossRef] [Green Version]
- Botelho, H.M.; Leal, S.S.; Veith, A.; Prosinecki, V.; Bauer, C.; Froöhlich, R.; Kletzin, A.; Gomes, C.M. Role of a novel disulfide bridge within the all-beta fold of soluble Rieske proteins. J. Biol. Inorg. Chem. 2010, 15, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Kletzin, A.; Ferreira, A.S.; Hechler, T.; Bandeiras, T.M.; Teixeira, M.; Gomes, C.M. A Rieske ferredoxin typifying a subtype within Rieske proteins: Spectroscopic, biochemical and stability studies. FEBS Lett. 2005, 579, 1020–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E°’ | pH | E°’ | pH | E°’ | pH | Reference | |
|---|---|---|---|---|---|---|---|
| [2Fe-2S]2+/+ (SoxF) | +0.39 | 6.7 | +0.33 | 7.4 | +0.27 | 8.2 | [138,139] a |
| - | - | +0.38 | 7.5 | - | - | [140] e,d | |
| Cu2+/+ (SoxE) | +0.30 | 8.0 | - | - | - | - | [137] c,f |
| CuA2+/+ (SoxH) | +0.24 | 6.4 | - | - | - | - | [141] f,g |
| CuB2+/+ | +0.37 | 7.4 | - | - | - | - | [142] a |
| Fe3+/2+ (heme AS)1 | +0.03 | 4.5 | - | - | - | - | [134,135] a |
| Fe3+/2+ (heme AS)2 | +0.10 | 4.5 | - | - | - | - | [134,135] a |
| Fe3+/2+ (Cyt b) | +0.20 | 6.5 | - | - | - | - | [134] b |
| Fe3+/2+ (Cyt b3) | +0.35 | 6.5 | - | - | - | - | [134] b |
| Qcal ↔ H2Qcal (±2e−) | +0.11 | 6.5 | - | - | - | - | [143,144] f |
| E°’ [2Fe-2S]I | E°’ [2Fe-2S]II | pH | References | |
|---|---|---|---|---|
| Archaea | ||||
| Sulfolobus acidocaldariusa | −0.30 b | −0.20 b | 7.5 | [149] |
| Bacteria | ||||
| Desulfovibrio gigasc,d [150,151] | −0.28 b,e | −0.29 b,f | 7.6 | [152] |
| Desulfovibrio alaskensis g [153] | −0.28 b | −0.32 b | 9.0 | [154] |
| Eukarya | ||||
| Mus Musculus (house mouse) h [155,156] | ~0.0 b,d | ~−0.1 V b,d | 7.4 | [157] |
| E°’ | pH | E°’ | pH | Reference | |
|---|---|---|---|---|---|
| Sulredoxin from Sulfololobus tokodaii sp. (Strain 7) a | +0.39 | 5.5 | +0.20 | 9.0 | [158,159] b |
| E°’ | pH | ||||
| Sulfolobus solfataricus (strain P-1) c | ~−0.1 | 7.0 | [160,161] b | ||
| E°’ | pH | ||||
| Acidianus ambivalens ferredoxin 2 (RFd2) d | ≥0.0 | 7.0 | [162] d | ||
| Acidianus ambivalens subtype ferredoxin (RFd) e | +0.17 | 7.1 | +0.17 | 7.8 | [163] f |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corsini, M.; Zanello, P. The Redox Active [2Fe-2S] Clusters: Key-Components of a Plethora of Enzymatic Reactions—Part I: Archaea. Inorganics 2022, 10, 14. https://doi.org/10.3390/inorganics10010014
Corsini M, Zanello P. The Redox Active [2Fe-2S] Clusters: Key-Components of a Plethora of Enzymatic Reactions—Part I: Archaea. Inorganics. 2022; 10(1):14. https://doi.org/10.3390/inorganics10010014
Chicago/Turabian StyleCorsini, Maddalena, and Piero Zanello. 2022. "The Redox Active [2Fe-2S] Clusters: Key-Components of a Plethora of Enzymatic Reactions—Part I: Archaea" Inorganics 10, no. 1: 14. https://doi.org/10.3390/inorganics10010014
APA StyleCorsini, M., & Zanello, P. (2022). The Redox Active [2Fe-2S] Clusters: Key-Components of a Plethora of Enzymatic Reactions—Part I: Archaea. Inorganics, 10(1), 14. https://doi.org/10.3390/inorganics10010014