Nickel Metalloregulators and Chaperones

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Nickel-Responsive Metalloregulators
2.1. NikR
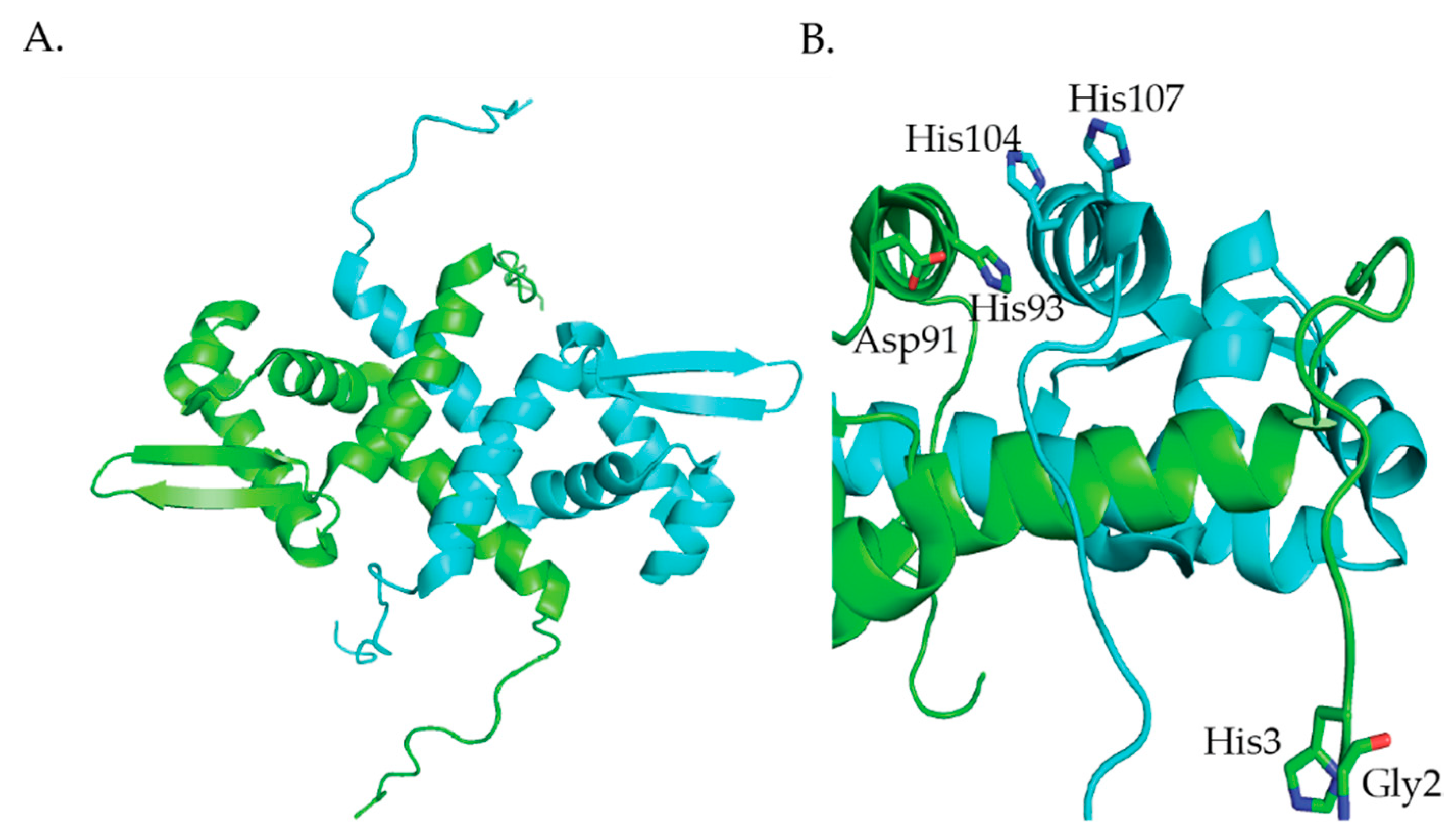
2.2. Nur
2.3. RcnR/Csor Family
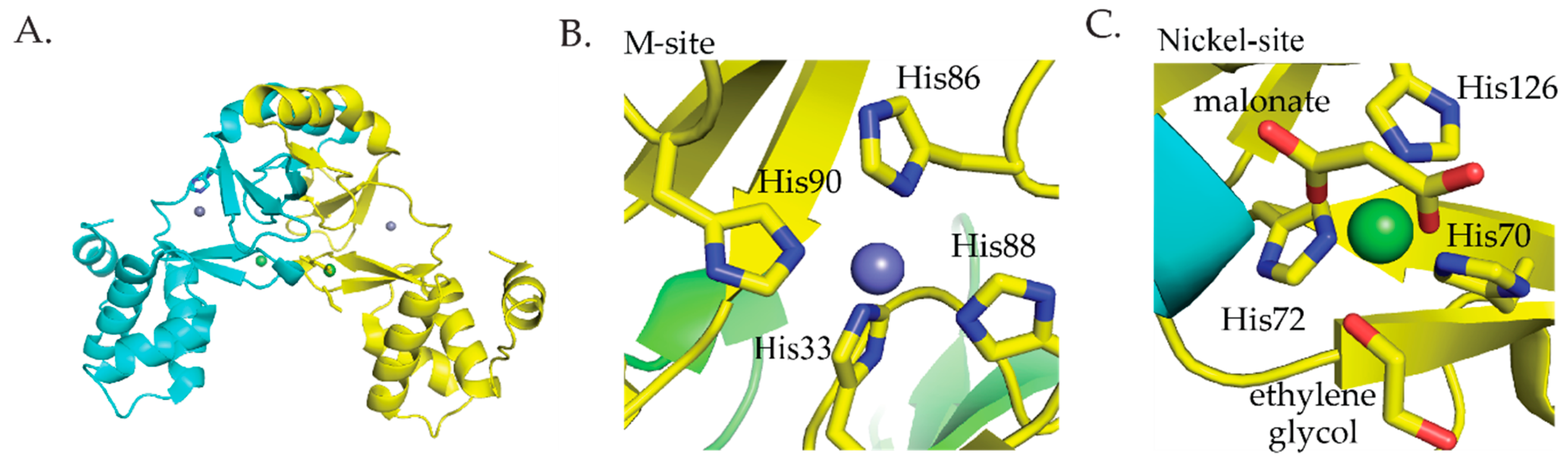
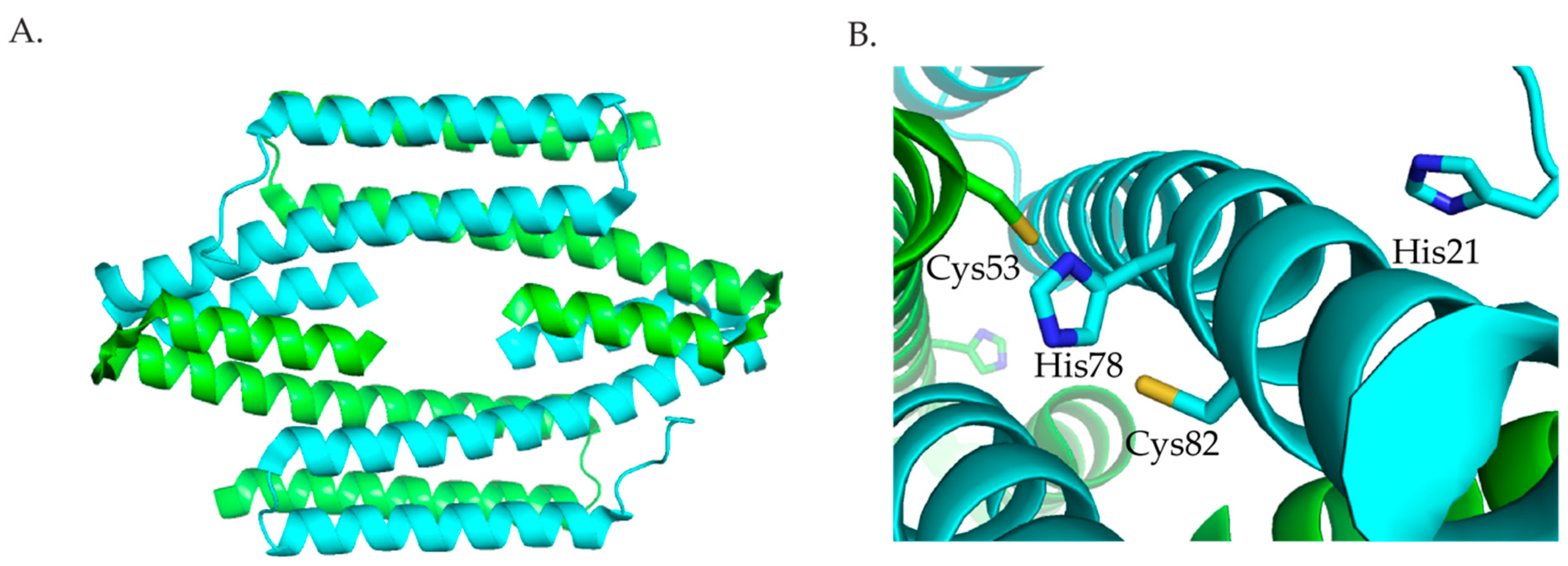
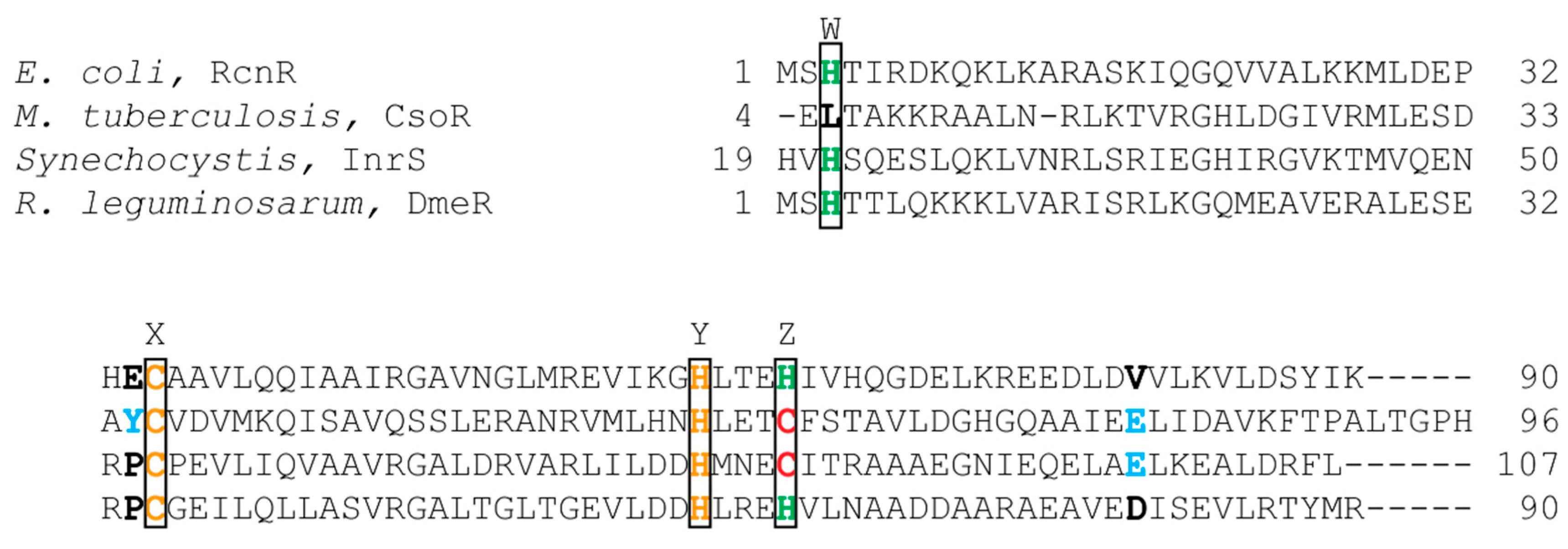
2.3.1. RcnR
2.3.2. InrS
2.3.3. DmeR
2.4. NmtR
2.5. Other Transcriptional Metalloregulators
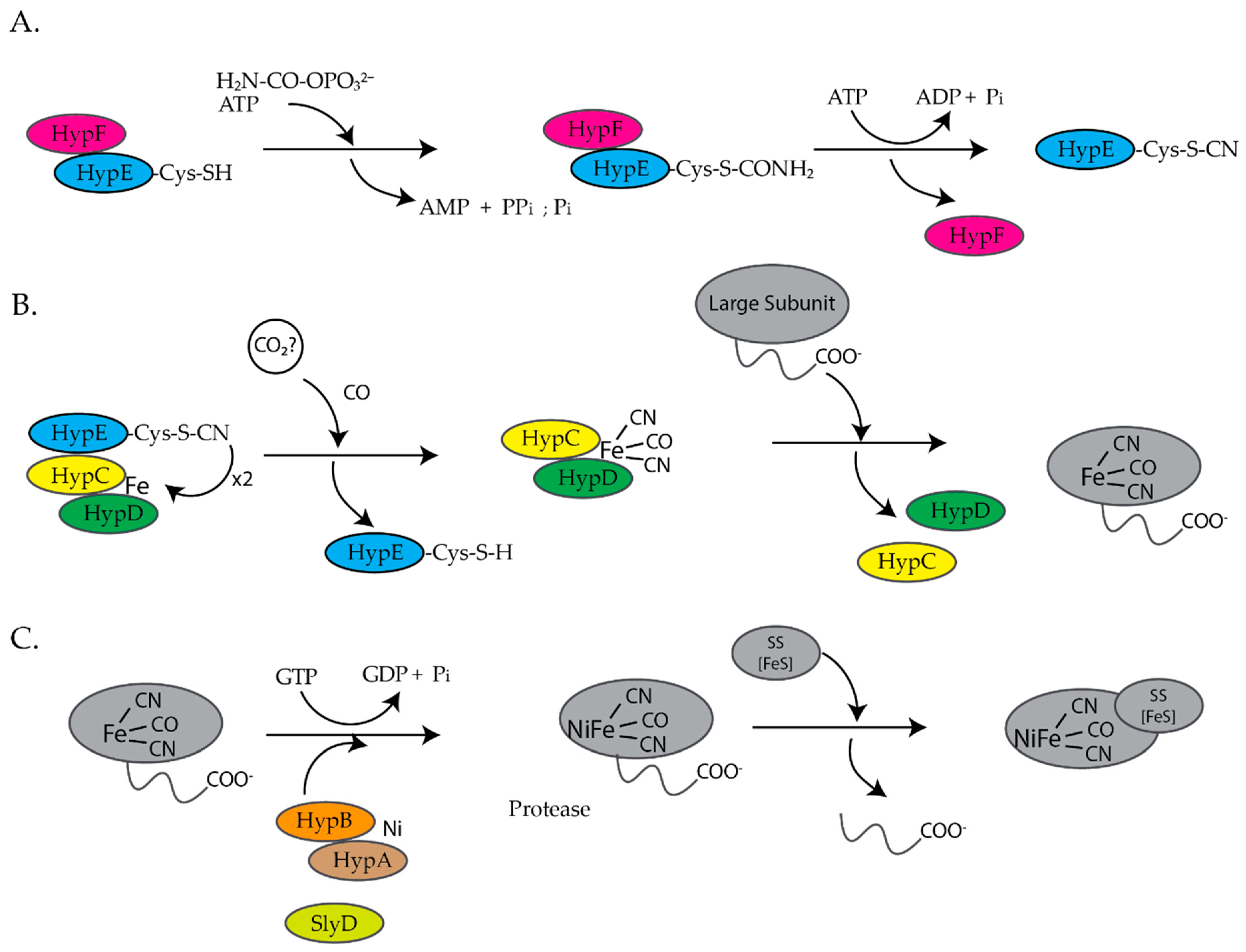
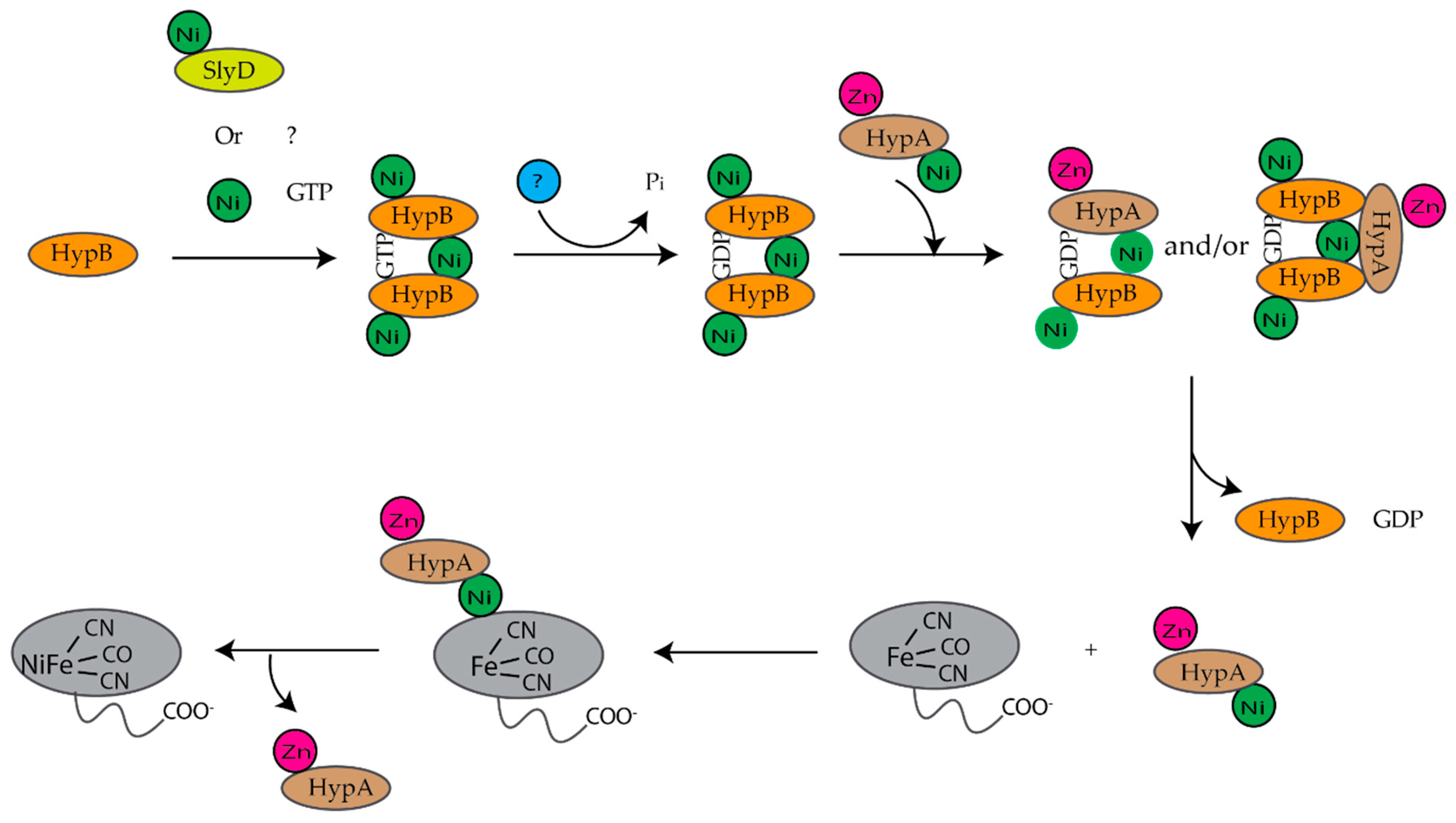
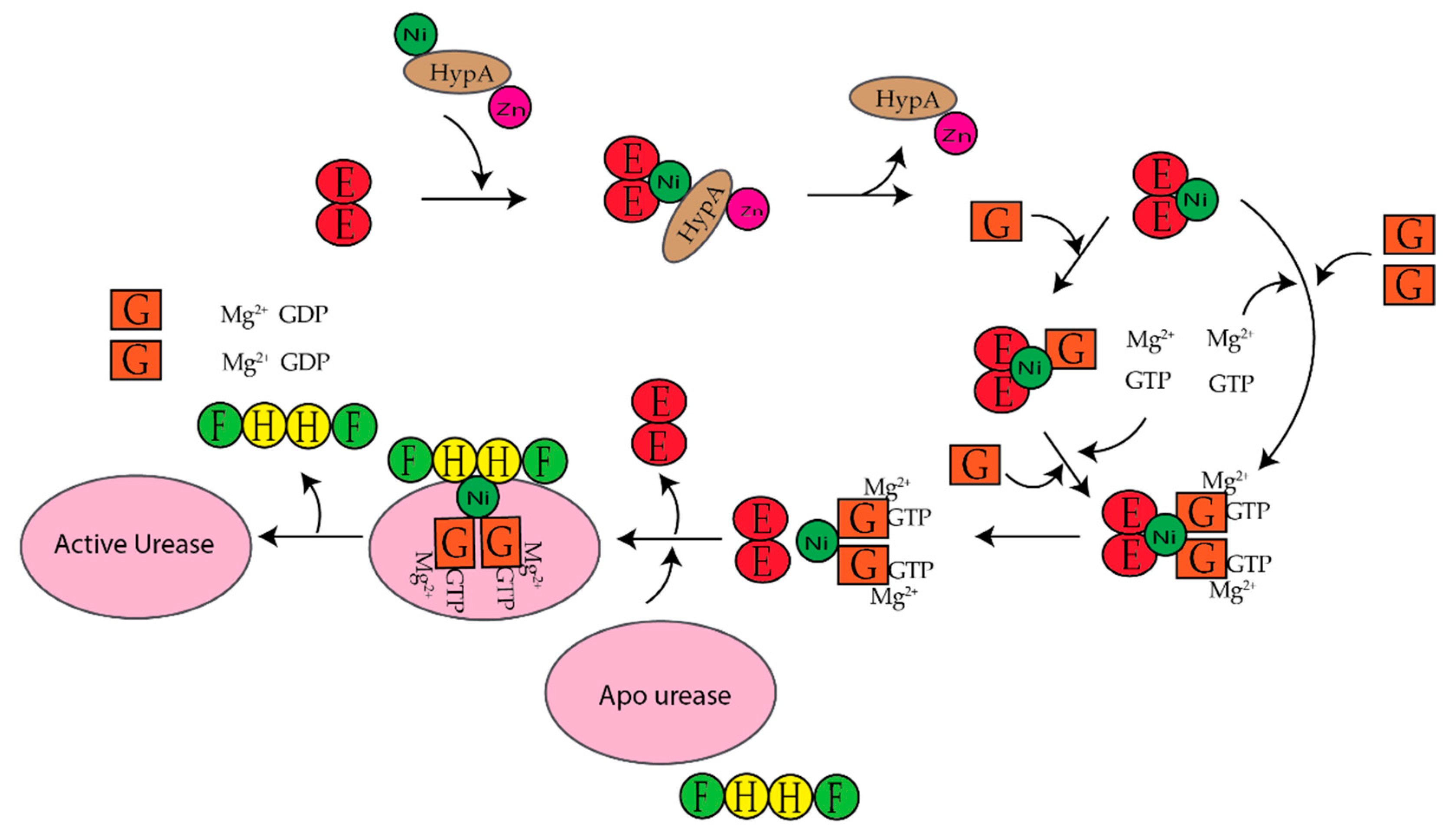
3. Ni(II) Chaperones Associated with [NiFe]-Hydrogenase and Urease
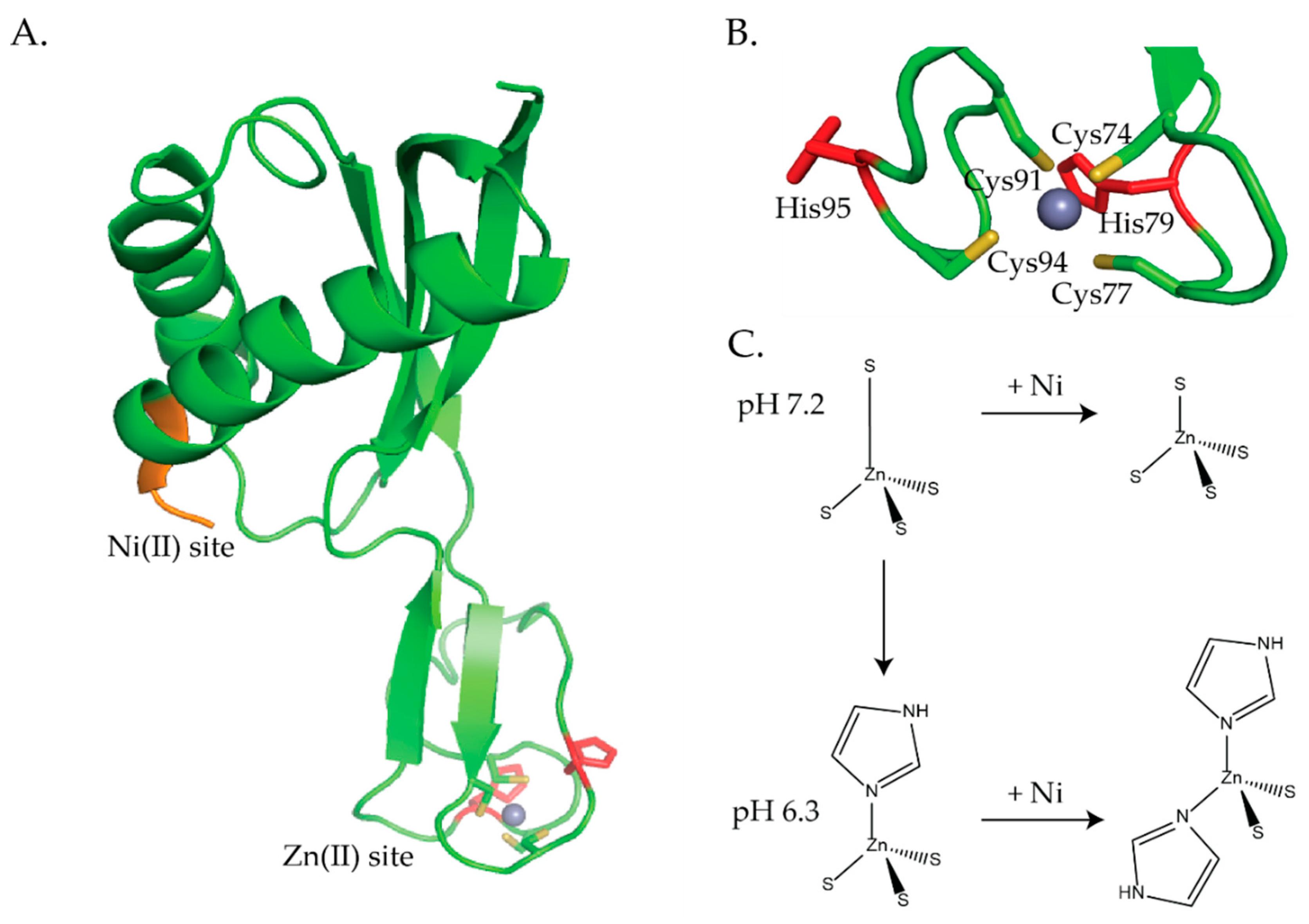
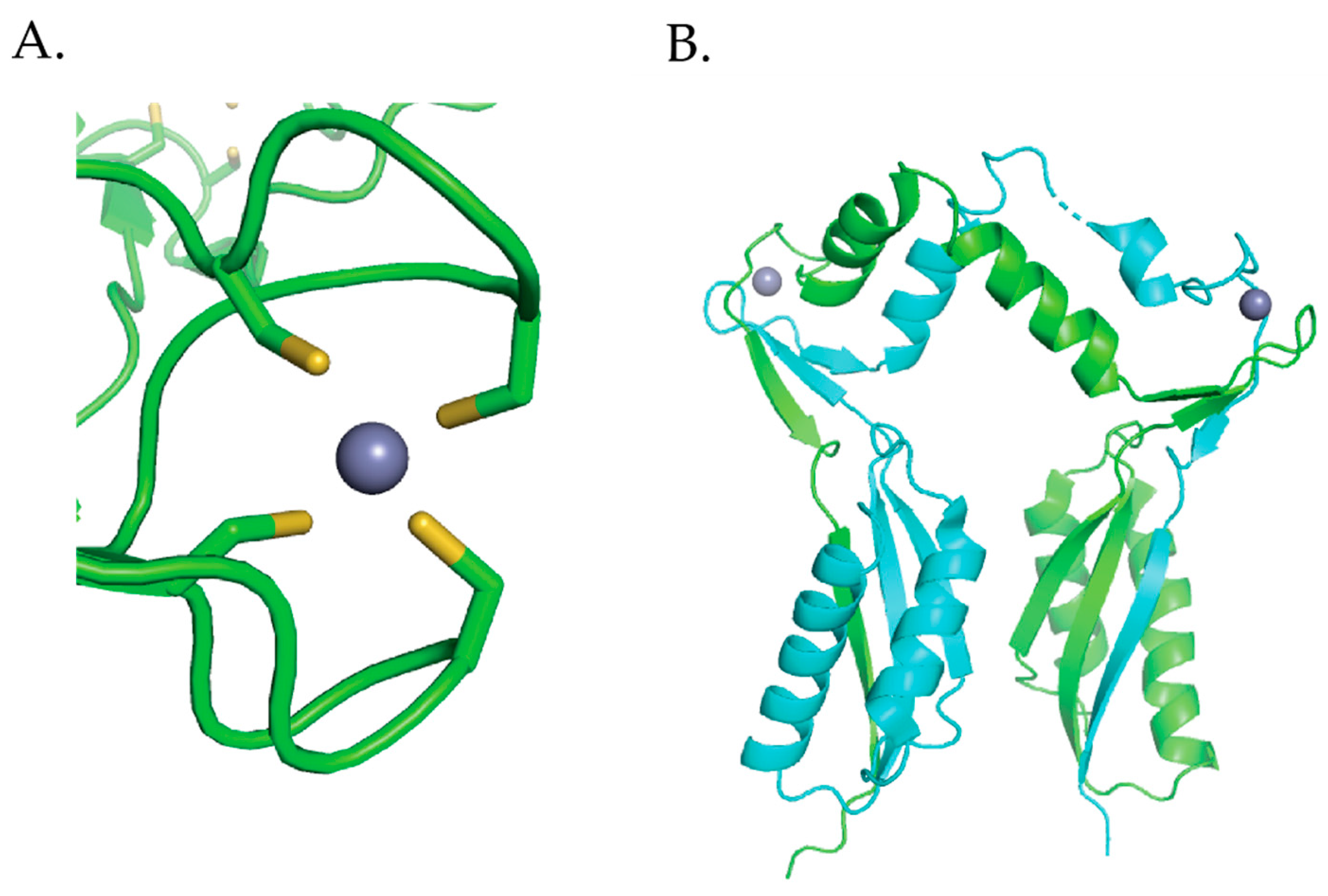
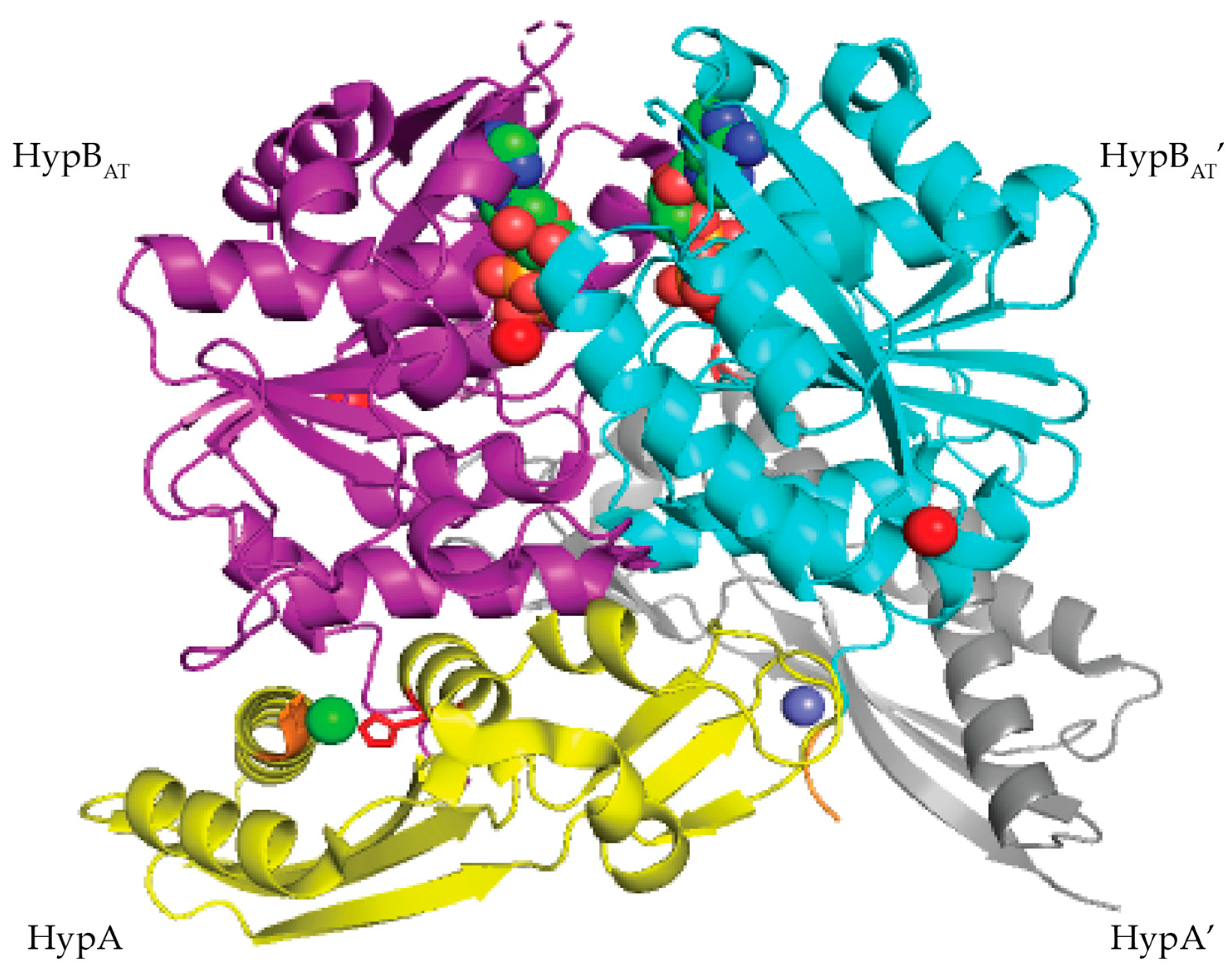
3.1. HypA
3.2. HypB
3.3. SlyD
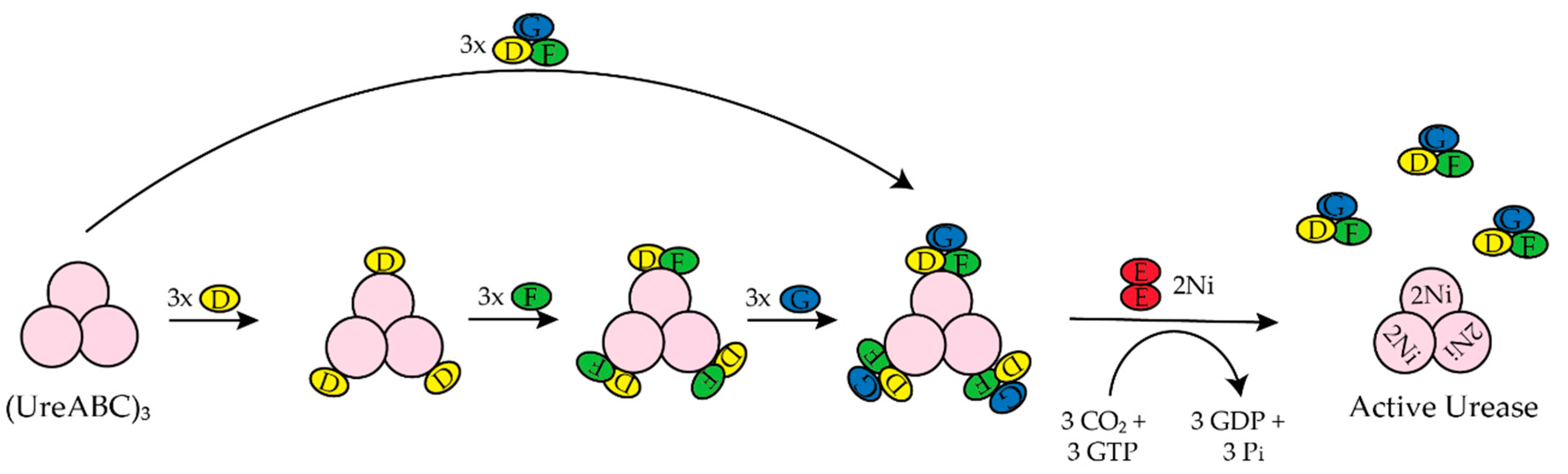
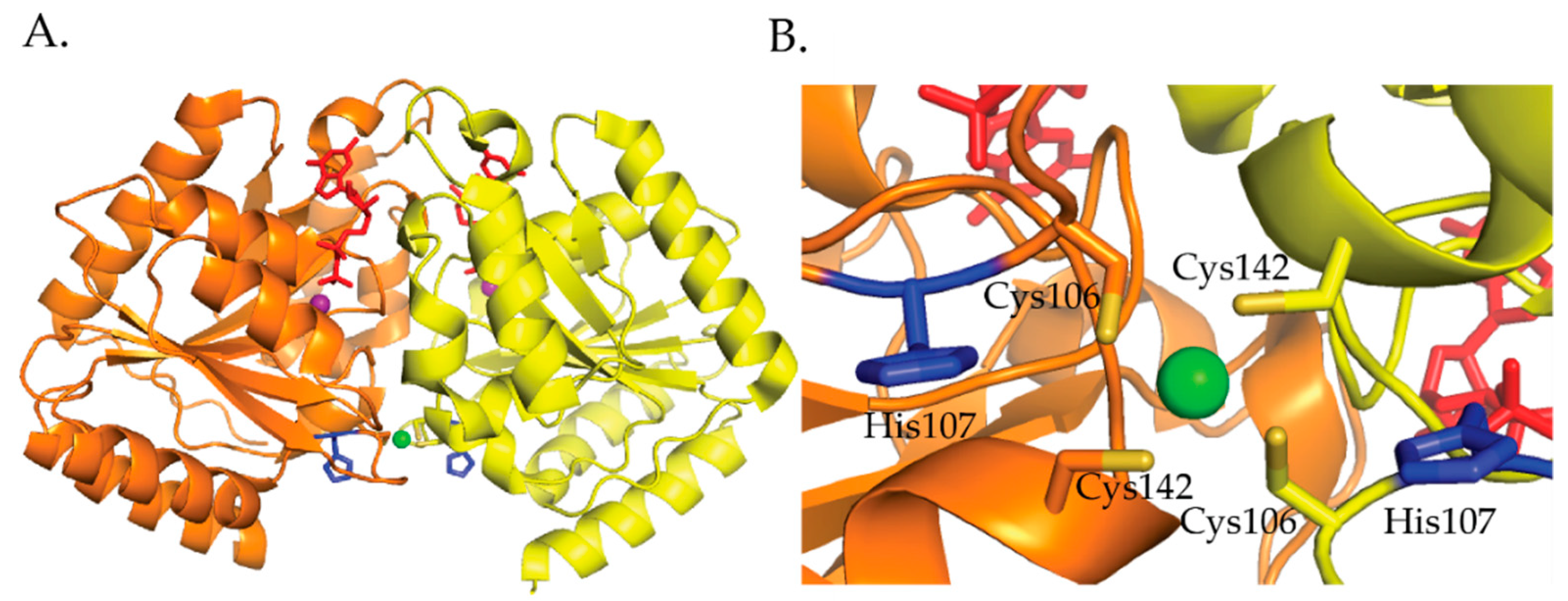
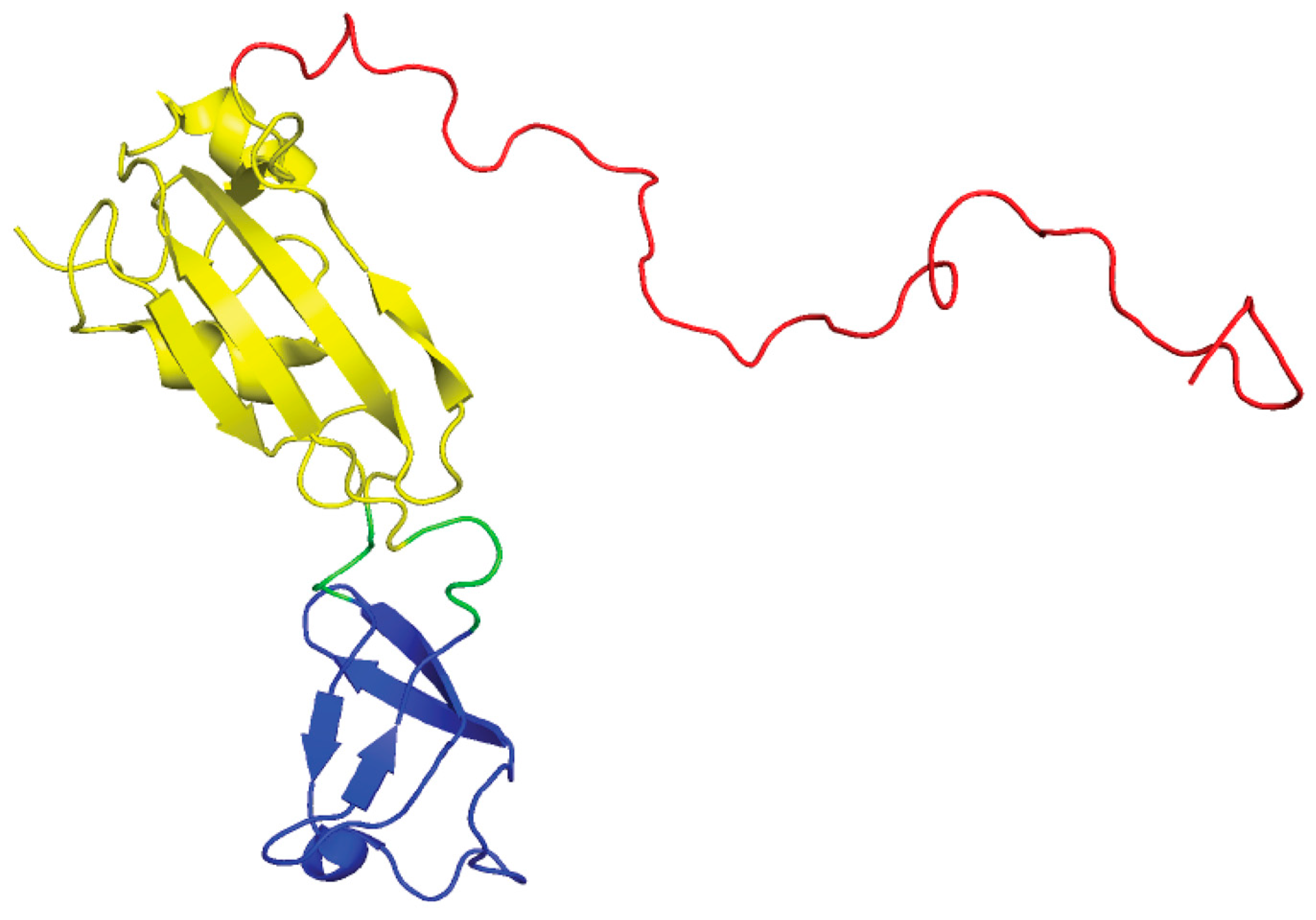
3.4. UreE
4. Protein–Protein Interactions
5. Conclusions
Supplementary Materials
Funding
Conflicts of Interest
References
- Maroney, M.J.; Ciurli, S. Nonredox nickel enzymes. Chem. Rev. 2014, 114, 4206–4228. [Google Scholar] [CrossRef] [PubMed]
- Maier, R.J. Use of molecular hydrogen as an energy substrate by human pathogenic bacteria. Biochem. Soc. Trans. 2005, 33, 83–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulrooney, S.B.; Hausinger, R.P. Nickel uptake and utilization by microorganisms. FEMS Microbiol. Rev. 2003, 27, 239–261. [Google Scholar] [CrossRef]
- Macomber, L.; Hausinger, R.P. Mechanisms of nickel toxicity in microorganisms. Metallomics 2011, 3, 1153–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, K.A.; Carr, C.E.; Maroney, M.J. Specific metal recognition in nickel trafficking. Biochemistry 2012, 51, 7816–7832. [Google Scholar] [CrossRef]
- De Pina, K.; Desjardin, V.; Mandrand-Berthelot, M.A.; Giordano, G.; Wu, L.F. Isolation and characterization of the nikR gene encoding a nickel-responsive regulator in Escherichia coli. J. Bacteriol. 1999, 181, 670–674. [Google Scholar] [PubMed]
- Blériot, C.; Effantin, G.; Lagarde, F.; Mandrand-Berthelot, M.A.; Rodrigue, A. RcnB is a periplasmic protein essential for maintaining intracellular Ni and Co concentrations in Escherichia coli. J. Bacteriol. 2011, 193, 3785–3793. [Google Scholar] [CrossRef]
- Iwig, J.S.; Rowe, J.L.; Chivers, P.T. Nickel homeostasis in Escherichia coli - the rcnR-rcnA efflux pathway and its linkage to NikR function. Mol. Microbiol. 2006, 62, 252–262. [Google Scholar] [CrossRef]
- Böck, A.; King, P.W.; Blokesch, M.; Posewitz, M.C. Maturation of hydrogenases. Adv. Microb. Physiol. 2006, 51, 1–71. [Google Scholar]
- Forzi, L.; Sawers, R.G. Maturation of [NiFe]-hydrogenases in Escherichia coli. Biometals 2007, 20, 565–578. [Google Scholar] [CrossRef]
- Fontecilla-Camps, J.C.; Amara, P.; Cavazza, C.; Nicolet, Y.; Volbeda, A. Structure–function relationships of anaerobic gas-processing metalloenzymes. Nature 2009, 460, 814–822. [Google Scholar] [CrossRef]
- Peters, J.W.; Schut, G.J.; Boyd, E.S.; Mulder, D.W.; Shepard, E.M.; Broderick, J.B.; King, P.W.; Adams, M.W.W. [FeFe]- and [NiFe]-hydrogenase diversity, mechanism, and maturation. Biochim. Biophys. Acta-Mol. Cell Res. 2015, 1853, 1350–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, S.; Sasaki, D.; Tominaga, T.; Miki, K. Structural basis of [NiFe] hydrogenase maturation by Hyp proteins. Biol. Chem. 2012, 393, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Leach, M.R.; Zamble, D.B. Metallocenter assembly of the hydrogenase enzymes. Curr. Opin. Chem. Biol. 2007, 11, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.W.; Butland, G.; Greenblatt, J.F.; Emili, A.; Zamble, D.B. A role for SlyD in the Escherichia coli hydrogenase biosynthetic pathway. J. Biol. Chem. 2005, 280, 4360–4366. [Google Scholar] [CrossRef] [PubMed]
- Pinske, C.; Sargent, F.; Sawers, R.G. SlyD-dependent nickel delivery limits maturation of [NiFe]-hydrogenases in late-stationary phase Escherichia coli cells. Metallomics 2015, 7, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Lacasse, M.J.; Zamble, D.B. [NiFe]-Hydrogenase maturation. Biochemistry 2016, 55, 1689–1701. [Google Scholar] [CrossRef] [PubMed]
- Hube, M.; Blokesch, M.; Böck, A. Network of hydrogenase maturation in Escherichia coli: Role of accessory proteins HypA and HybF. J. Bacteriol. 2002, 184, 3879–3885. [Google Scholar] [CrossRef] [PubMed]
- Reissmann, S.; Hochleitner, E.; Wang, H.; Paschos, A.; Lottspeich, F.; Glass, R.S.; Böck, A. Taming of a poison: Biosynthesis of the NiFe-hydrogenase cyanide ligands. Science 2003, 299, 1067–1070. [Google Scholar] [CrossRef]
- Li, Y.; Zamble, D.B. Nickel homeostasis and nickel regulation: An overview. Chem. Rev. 2009, 109, 4617–4643. [Google Scholar] [CrossRef]
- Lubitz, W.; Ogata, H.; Rüdiger, O.; Reijerse, E. Hydrogenases. Chem. Rev. 2014, 114, 4081–4148. [Google Scholar] [CrossRef] [PubMed]
- Dunn, B.E.; Cohen, H.; Blaser, M.J. Helicobacter pylori. Clin. Microbiol. Rev. 1997, 10, 720–741. [Google Scholar] [CrossRef] [PubMed]
- Kusters, J.G.; Van Vliet, A.H.M.; Ernst, J.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.-Y.; Sheu, B.S.; Wu, J.J. Helicobacter pylori infection: An overview of bacterial virulence factors and pathogenesis. Biomed. J. 2016, 39, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y. Mechanisms of disease: Helicobacter pylori virulence factors. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 629–641. [Google Scholar] [CrossRef]
- Olson, J.W. Molecular hydrogen as an energy source for Helicobacter pylori. Science 2002, 298, 1788–1790. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, M.A.; Macomber, L.; Hausinger, R.P. Biosynthesis of the urease metallocenter. J. Biol. Chem. 2013, 288, 13178–13185. [Google Scholar] [CrossRef]
- Carter, E.L.; Flugga, N.; Boer, J.L.; Mulrooney, S.B.; Hausinger, R.P. Interplay of metal ions and urease. Metallomics 2009, 1, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Zambelli, B.; Musiani, F.; Benini, S.; Ciurli, S. Chemistry of Ni2+ in urease: Sensing, trafficking, and catalysis. Acc. Chem. Res. 2011, 44, 520–530. [Google Scholar] [CrossRef]
- Zeer-Wanklyn, C.J.; Zamble, D.B. Microbial nickel: Cellular uptake and delivery to enzyme centers. Curr. Opin. Chem. Biol. 2017, 37, 80–88. [Google Scholar] [CrossRef]
- Benoit, S.L.; Mehta, N.; Weinberg, M.V.; Maier, C.; Maier, R.J. Interaction between the Helicobacter pylori accessory proteins HypA and UreE is needed for urease maturation. Microbiology 2007, 153, 1474–1482. [Google Scholar] [CrossRef]
- Olson, J.W.; Mehta, N.S.; Maier, R.J. Requirement of nickel metabolism proteins HypA and HypB for full activity of both hydrogenase and urease in Helicobacter pylori. Mol. Microbiol. 2001, 39, 176–182. [Google Scholar] [CrossRef]
- Johnson, R.C.; Hu, H.Q.; Merrell, D.S.; Maroney, M.J. Dynamic HypA zinc site is essential for acid viability and proper urease maturation in Helicobacter pylori. Metallomics 2015, 7, 674–682. [Google Scholar] [CrossRef]
- Jones, M.D.; Li, Y.; Zamble, D.B. Acid-responsive activity of the Helicobacter pylori metalloregulator NikR. Proc. Natl. Acad. Sci. USA 2018, 115, 8966–8971. [Google Scholar] [CrossRef]
- Wolfram, L.; Bauerfeind, P. Activities of urease and nickel uptake of Helicobacter pylori proteins are media- and host-dependent. Helicobacter 2009, 14, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Higgins, K.A.; Giedroc, D.P. Metal specificity of metallosensors. In Encyclopedia of Inorganic and Bioinorganic Chemistry; John Wiley & Sons, Ltd.: Chichester, UK, 2013; pp. 209–224. [Google Scholar]
- Musiani, F.; Zambelli, B.; Bazzani, M.; Mazzei, L.; Ciurli, S. Nickel-responsive transcriptional regulators. Metallomics 2015, 7, 1305–1318. [Google Scholar] [CrossRef]
- Chivers, P.T.; Sauer, R.T. NikR repressor: High-affinity nickel binding to the C-terminal domain regulates binding to operator DNA. Chem. Biol. 2002, 9, 1141–1148. [Google Scholar] [CrossRef]
- Bloom, S.L.; Zamble, D.B. Metal-selective DNA-binding response of Escherichia coli NikR. Biochemistry 2004, 43, 10029–10038. [Google Scholar] [CrossRef]
- Chivers, P.T.; Sauer, R.T. Regulation of high affinity nickel uptake in bacteria. Ni2+-dependent interaction of NikR with wild-type and mutant operator sites. J. Biol. Chem. 2000, 275, 19735–19741. [Google Scholar] [CrossRef]
- Budnick, J.A.; Prado-Sanchez, E.; Caswell, C.C. Defining the regulatory mechanism of NikR, a nickel-responsive transcriptional regulator, in Brucella abortus. Microbiology 2018, 164, 1320–1325. [Google Scholar] [CrossRef]
- Fabini, E.; Zambelli, B.; Mazzei, L.; Ciurli, S.; Bertucci, C. Surface plasmon resonance and isothermal titration calorimetry to monitor the Ni(II)-dependent binding of Helicobacter pylori NikR to DNA. Anal. Bioanal. Chem. 2016, 408, 7971–7980. [Google Scholar] [CrossRef]
- Contreras, M.; Thiberge, J.M.; Mandrand-Berthelot, M.A.; Labigne, A. Characterization of the roles of NikR, a nickel-responsive pleiotropic autoregulator of Helicobacter pylori. Mol. Microbiol. 2003, 49, 947–963. [Google Scholar] [CrossRef]
- Vannini, A.; Pinatel, E.; Costantini, P.E.; Pelliciari, S.; Roncarati, D.; Puccio, S.; De Bellis, G.; Peano, C.; Danielli, A. Comprehensive mapping of the Helicobacter pylori NikR regulon provides new insights in bacterial nickel responses. Sci. Rep. 2017, 7, 45458. [Google Scholar] [CrossRef]
- Schreiter, E.R.; Sintchak, M.D.; Guo, Y.; Chivers, P.T.; Sauer, R.T.; Drennan, C.L. Crystal structure of the nickel-responsive transcription factor NikR. Nat. Struct. Biol. 2003, 10, 794–799. [Google Scholar] [CrossRef]
- Schreiter, E.R.; Wang, S.C.; Zamble, D.B.; Drennan, C.L. NikR-operator complex structure and the mechanism of repressor activation by metal ions. Proc. Natl. Acad. Sci. USA 2006, 103, 13676–13681. [Google Scholar] [CrossRef]
- Phillips, C.M.; Schreiter, E.R.; Guo, Y.; Wang, S.C.; Zamble, D.B.; Drennan, C.L. Structural basis of the metal specificity for nickel regulatory protein NikR. Biochemistry 2008, 47, 1938–1946. [Google Scholar] [CrossRef]
- Phillips, C.M.; Schreiter, E.R.; Stultz, C.M.; Drennan, C.L. Structural basis of low-affinity nickel binding to the nickel-responsive transcription factor NikR from Escherichia coli. Biochemistry 2010, 49, 7830–7838. [Google Scholar] [CrossRef]
- Chivers, P.T.; Tahirov, T.H. Structure of Pyrococcus horikoshii NikR: Nickel sensing and implications for the regulation of DNA recognition. J. Mol. Biol. 2005, 348, 597–607. [Google Scholar] [CrossRef]
- Bahlawane, C.; Dian, C.; Muller, C.; Round, A.; Fauquant, C.; Schauer, K.; de Reuse, H.; Terradot, L.; Michaud-Soret, I. Structural and mechanistic insights into Helicobacter pylori NikR activation. Nucleic Acids Res. 2010, 38, 3106–3118. [Google Scholar] [CrossRef]
- Benini, S.; Cianci, M.; Ciurli, S. Holo-Ni2+ Helicobacter pylori NikR contains four square-planar nickel-binding sites at physiological pH. Dalt. Trans. 2011, 40, 7831–7833. [Google Scholar] [CrossRef]
- West, A.L.; St John, F.; Lopes, P.E.; MacKerell, A.D., Jr.; Pozharski, E.; Michel, S.L. Holo-Ni(II)HpNikR is an asymmetric tetramer containing two different nickel-binding sites. J. Am. Chem. Soc. 2010, 132, 14447–14456. [Google Scholar] [CrossRef]
- West, A.L.; Evans, S.E.; Gonzalez, J.M.; Carter, L.G.; Tsuruta, H.; Pozharski, E.; Michel, S.L.J. Ni(II) coordination to mixed sites modulates DNA binding of HpNikR via a long-range effect. Proc. Natl. Acad. Sci. USA 2012, 109, 5633–5638. [Google Scholar] [CrossRef]
- Dian, C.; Schauer, K.; Kapp, U.; McSweeney, S.M.; Labigne, A.; Terradot, L. Structural basis of the nickel response in Helicobacter pylori: Crystal structures of HpNikR in apo and nickel-bound states. J. Mol. Biol. 2006, 361, 715–730. [Google Scholar] [CrossRef]
- Chivers, P.T.; Sauer, R.T. NikR is a ribbon-helix-helix DNA-binding protein. Protein Sci. 1999, 8, 2494–2500. [Google Scholar] [CrossRef]
- Musiani, F.; Bertosa, B.; Magistrato, A.; Zambelli, B.; Turano, P.; Losasso, V.; Micheletti, C.; Ciurli, S.; Carloni, P. Computational study of the DNA-binding protein Helicobacter pylori NikR: The role of Ni2+. J. Chem. Theory Comput. 2010, 6, 3503–3515. [Google Scholar] [CrossRef]
- Leitch, S.; Bradley, M.J.; Rowe, J.L.; Chivers, P.T.; Maroney, M.J. Nickel-specific response in the transcriptional regulator, Escherichia coli NikR. J. Am. Chem. Soc. 2007, 129, 5085–5095. [Google Scholar] [CrossRef]
- Zambelli, B.; Bellucci, M.; Danielli, A.; Scarlato, V.; Ciurli, S. The Ni2+ binding properties of Helicobacter pylori NikR. Chem. Commun. 2007, 35, 3649–3651. [Google Scholar] [CrossRef]
- Wang, S.C.; Dias, A.V.; Bloom, S.L.; Zamble, D.B. Selectivity of metal binding and metal-induced stability of Escherichia coli NikR. Biochemistry 2004, 43, 10018–10028. [Google Scholar] [CrossRef]
- Ahn, B.E.; Cha, J.; Lee, E.J.; Han, A.R.; Thompson, C.J.; Roe, J.H. Nur, a nickel-responsive regulator of the Fur family, regulates superoxide dismutases and nickel transport in Streptomyces coelicolor. Mol. Microbiol. 2006, 59, 1848–1858. [Google Scholar] [CrossRef]
- Kim, H.M.; Shin, J.-H.; Cho, Y.-B.; Roe, J.-H. Inverse regulation of Fe- and Ni-containing SOD genes by a Fur family regulator Nur through small RNA processed from 3′UTR of the sodF mRNA. Nucleic Acids Res. 2014, 42, 2003–2014. [Google Scholar] [CrossRef]
- An, Y.J.; Ahn, B.E.; Han, A.R.; Kim, H.M.; Chung, K.M.; Shin, J.H.; Cho, Y.B.; Roe, J.H.; Cha, S.S. Structural basis for the specialization of Nur, a nickel-specific Fur homolog, in metal sensing and DNA recognition. Nucleic Acids Res. 2009, 37, 3442–3451. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Ramesh, A.; Ma, Z.; Ward, S.K.; Zhang, L.; George, G.N.; Talaat, A.M.; Sacchettini, J.C.; Giedroc, D.P. CsoR is a novel Mycobacterium tuberculosis copper-sensing transcriptional regulator. Nat. Chem. Biol. 2007, 3, 60–68. [Google Scholar] [CrossRef]
- Foster, A.W.; Pernil, R.; Patterson, C.J.; Scott, A.J.P.; Pålsson, L.O.; Pal, R.; Cummins, I.; Chivers, P.T.; Pohl, E.; Robinson, N.J. A tight tunable range for Ni(II) sensing and buffering in cells. Nat. Chem. Biol. 2017, 13, 409–414. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Cowart, D.M.; Ward, B.P.; Arnold, R.J.; DiMarchi, R.D.; Zhang, L.; George, G.N.; Scott, R.A.; Giedroc, D.P. Unnatural amino acid substitution as a probe of the allosteric coupling pathway in a mycobacterial Cu(I) sensor. J. Am. Chem. Soc. 2009, 131, 18044–18045. [Google Scholar] [CrossRef]
- Rodrigue, A.; Effantin, G.; Mandrand-Berthelot, M.A. Identification of rcnA (yohM), a nickel and cobalt resistance gene in Escherichia coli. J. Bacteriol. 2005, 187, 2912–2916. [Google Scholar] [CrossRef]
- Blaha, D.; Arous, S.; Bleriot, C.; Dorel, C.; Mandrand-Berthelot, M.A.; Rodrigue, A. The Escherichia coli metallo-regulator RcnR represses rcnA and rcnR transcription through binding on a shared operator site: Insights into regulatory specificity towards nickel and cobalt. Biochimie 2011, 93, 434–439. [Google Scholar] [CrossRef]
- Iwig, J.S.; Chivers, P.T. DNA recognition and wrapping by Escherichia coli RcnR. J. Mol. Biol. 2009, 393, 514–526. [Google Scholar] [CrossRef]
- Iwig, J.S.; Leitch, S.; Herbst, R.W.; Maroney, M.J.; Chivers, P.T. Ni(II) and Co(II) sensing by Escherichia coli RcnR. J. Am. Chem. Soc. 2008, 130, 7592–7606. [Google Scholar] [CrossRef]
- Higgins, K.A.; Chivers, P.T.; Maroney, M.J. Role of the N-terminus in determining metal-specific responses in the E. coli Ni- and Co-responsive metalloregulator, RcnR. J. Am. Chem. Soc. 2012, 134, 7081–7093. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.; Potter, S.C.; Lopez, R. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Higgins, K.A.; Hu, H.Q.; Chivers, P.T.; Maroney, M.J. Effects of select histidine to cysteine mutations on transcriptional regulation by Escherichia coli RcnR. Biochemistry 2013, 52, 84–97. [Google Scholar] [CrossRef]
- Carr, C.E.; Musiani, F.; Huang, H.; Chivers, P.T.; Ciurli, S.; Maroney, M.J. Glutamate ligation in the Ni(II)- and Co(II)-responsive Escherichia coli transcriptional regulator, RcnR. Inorg. Chem. 2017, 56, 6459–6476. [Google Scholar] [CrossRef]
- Chang, F.M.J.; Martin, J.E.; Giedroc, D.P. Electrostatic occlusion and quaternary structural ion pairing are key determinants of Cu(I)-mediated allostery in the copper-sensing operon repressor (CsoR). Biochemistry 2015, 54, 2463–2472. [Google Scholar] [CrossRef]
- Denby, K.J.; Iwig, J.; Bisson, C.; Westwood, J.; Rolfe, M.D.; Sedelnikova, S.E.; Higgins, K.; Maroney, M.J.; Baker, P.J.; Chivers, P.T.; et al. The mechanism of a formaldehyde-sensing transcriptional regulator. Sci. Rep. 2016, 6, 38879. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.-T.; Bobst, C.E.; Iwig, J.S.; Chivers, P.T.; Kaltashov, I.A.; Maroney, M.J. Co(II) and Ni(II) binding of the Escherichia coli transcriptional repressor RcnR orders its N terminus, alters helix dynamics, and reduces DNA affinity. J. Biol. Chem. 2018, 293, 324–332. [Google Scholar] [CrossRef]
- Foster, A.W.; Patterson, C.J.; Pernil, R.; Hess, C.R.; Robinson, N.J. Cytosolic Ni(II) sensor in cyanobacterium. J. Biol. Chem. 2012, 287, 12142–12151. [Google Scholar] [CrossRef]
- Foster, A.W.; Pernil, R.; Patterson, C.J.; Robinson, N.J. Metal specificity of cyanobacterial nickel-responsive repressor InrS: Cells maintain zinc and copper below the detection threshold for InrS. Mol. Microbiol. 2014, 92, 797–812. [Google Scholar] [CrossRef]
- Carr, C.E.; Foster, A.W.; Maroney, M.J. An XAS investigation of the nickel site structure in the transcriptional regulator InrS. J. Inorg. Biochem. 2017, 177, 352–358. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Cowart, D.M.; Scott, R.A.; Giedroc, D.P. Molecular insights into the metal selectivity of the copper(I)-sensing repressor CsoR from Bacillus subtilis. Biochemistry 2009, 48, 3325–3334. [Google Scholar] [CrossRef]
- Rubio-Sanz, L.; Prieto, R.I.; Imperial, J.; Palacios, J.M.; Brito, B. Functional and expression analysis of the metal-inducible dmeRF System from Rhizobium leguminosarum bv. viciae. Appl. Environ. Microbiol. 2013, 79, 6414–6422. [Google Scholar] [CrossRef]
- Dokpikul, T.; Chaoprasid, P.; Saninjuk, K.; Sirirakphaisarn, S.; Johnrod, J.; Nookabkaew, S.; Sukchawalit, R.; Mongkolsuk, S. Regulation of the cobalt/nickel efflux operon dmeRF in Agrobacterium tumefaciens and a Link between the iron-Sensing regulator RirA and cobalt/nickel resistance. Appl. Environ. Microbiol. 2016, 82, 4732–4742. [Google Scholar] [CrossRef]
- Li, Z.; Song, X.; Wang, J.; Bai, X.; Gao, E.; Wei, G. Nickel and cobalt resistance properties of Sinorhizobium meliloti isolated from Medicago lupulina growing in gold mine tailing. PeerJ 2018, 6, e5202. [Google Scholar] [CrossRef]
- Cavet, J.S.; Meng, W.; Pennella, M.A.; Appelhoff, R.J.; Giedroc, D.P.; Robinson, N.J. A nickel-cobalt-sensing ArsR-SmtB family repressor. Contributions of cytosol and effector binding sites to metal selectivity. J. Biol. Chem. 2002, 277, 38441–38448. [Google Scholar] [CrossRef]
- Pennella, M.A.; Shokes, J.E.; Cosper, N.J.; Scott, R.A.; Giedroc, D.P. Structural elements of metal selectivity in metal sensor proteins. Proc. Natl. Acad. Sci. USA 2003, 100, 3713–3718. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Caballero, H.; Lee, C.W.; Giedroc, D.P. Mycobacterium tuberculosis NmtR harbors a nickel sensing site with parallels to Escherichia coli RcnR. Biochemistry 2011, 50, 7941–7952. [Google Scholar] [CrossRef]
- Lee, C.W.; Chakravorty, D.K.; Chang, F.-M.J.; Reyes-Caballero, H.; Ye, Y.; Merz, K.M.; Giedroc, D.P. Solution structure of Mycobacterium tuberculosis NmtR in the apo state: Insights into Ni(II)-mediated allostery. Biochemistry 2012, 51, 2619–2629. [Google Scholar] [CrossRef]
- Kim, J.-S.; Kang, S.-O.; Lee, J.K. The protein complex composed of nickel-binding SrnQ and DNA binding motif-bearing SrnR of Streptomyces griseus represses sodF transcription in the presence of nickel. J. Biol. Chem. 2003, 278, 18455–18463. [Google Scholar] [CrossRef]
- Campbell, D.R.; Chapman, K.E.; Waldron, K.J.; Tottey, S.; Kendall, S.; Cavallaro, G.; Andreini, C.; Hinds, J.; Stoker, N.G.; Robinson, N.J.; et al. Mycobacterial cells have dual nickel-cobalt sensors: Sequence relationships and metal sites of metal-responsive repressors are not congruent. J. Biol. Chem. 2007, 282, 32298–32310. [Google Scholar] [CrossRef]
- Zhu, T.; Tian, J.; Zhang, S.; Wu, N.; Fan, Y. Identification of the transcriptional regulator NcrB in the nickel resistance determinant of Leptospirillum ferriphilum UBK03. PLoS ONE 2011, 6, e17367. [Google Scholar] [CrossRef]
- Benoit, S.L.; Maier, R.J. Mua (HP0868) is a nickel-binding protein that modulates urease activity in Helicobacter pylori. MBio 2011, 2, e00039-11. [Google Scholar] [CrossRef]
- Kidd, S.P.; Djoko, K.Y.; Ng, J.; Argente, M.P.; Jennings, M.P.; McEwan, A.G. A novel nickel responsive MerR-like regulator, NimR, from Haemophilus influenzae. Metallomics 2011, 3, 1009–1018. [Google Scholar] [CrossRef]
- Blokesch, M.; Rohrmoser, M.; Rode, S.; Böck, A. HybF, a zinc-containing protein involved in NiFe hydrogenase maturation. J. Bacteriol. 2004, 186, 2603–2611. [Google Scholar] [CrossRef]
- Atanassova, A.; Zamble, D.B. Escherichia coli HypA is a zinc metalloprotein with a weak affinity for nickel. J. Bacteriol. 2005, 187, 4689–4697. [Google Scholar] [CrossRef]
- Mehta, N.; Olson, J.W.; Maier, R.J. Characterization of Helicobacter pylori nickel metabolism accessory proteins needed for maturation of both urease and hydrogenase. J. Bacteriol. 2003, 185, 726–734. [Google Scholar] [CrossRef]
- Hu, H.Q.; Johnson, R.C.; Merrell, D.S.; Maroney, M.J. Nickel ligation of the N-terminal amine of HypA is required for urease maturation in Helicobacter pylori. Biochemistry 2017, 56, 1105–1116. [Google Scholar] [CrossRef]
- Herbst, R.W.; Perovic, I.; Martin-Diaconescu, V.; O’Brien, K.; Chivers, P.T.; Pochapsky, S.S.; Pochapsky, T.C.; Maroney, M.J. Communication between the zinc and nickel sites in dimeric HypA: Metal recognition and pH sensing. J. Am. Chem. Soc. 2010, 132, 10338–10351. [Google Scholar] [CrossRef]
- Xia, W.; Li, H.; Sze, K.-H.; Sun, H. Structure of a nickel chaperone, HypA, from Helicobacter pylori reveals two distinct metal binding sites. J. Am. Chem. Soc. 2009, 131, 10031–10040. [Google Scholar] [CrossRef]
- Watanabe, S.; Arai, T.; Matsumi, R.; Atomi, H.; Imanaka, T.; Miki, K. Crystal structure of HypA, a nickel-binding metallochaperone for [NiFe] hydrogenase maturation. J. Mol. Biol. 2009, 394, 448–459. [Google Scholar] [CrossRef]
- Spronk, C.A.E.M.; Żerko, S.; Górka, M.; Koźmiński, W.; Bardiaux, B.; Zambelli, B.; Musiani, F.; Piccioli, M.; Basak, P.; Blum, F.C.; et al. Structure and dynamics of Helicobacter pylori nickel-chaperone HypA: An integrated approach using NMR spectroscopy, functional assays and computational tools. J. Biol. Inorg. Chem. 2018, 23, 1309–1330. [Google Scholar] [CrossRef]
- Watanabe, S.; Kawashima, T.; Nishitani, Y.; Kanai, T.; Wada, T.; Inaba, K.; Atomi, H.; Imanaka, T.; Miki, K. Structural basis of a Ni acquisition cycle for [NiFe] hydrogenase by Ni-metallochaperone HypA and its enhancer. Proc. Natl. Acad. Sci. USA 2015, 112, 7701–7706. [Google Scholar] [CrossRef] [Green Version]
- Douglas, C.D.; Ngu, T.T.; Kaluarachchi, H.; Zamble, D.B. Metal Transfer within the Escherichia coli HypB–HypA Complex of Hydrogenase Accessory Proteins. Biochemistry 2013, 52, 6030–6039. [Google Scholar] [CrossRef]
- Hu, H.Q.; Huang, H.; Maroney, M.J. The Helicobacter pylori HypA·UreE2 complex contains a novel high-affinity Ni(II)-binding site. Biochemistry 2018, 57, 2932–2942. [Google Scholar] [CrossRef]
- Kennedy, D.C.; Herbst, R.W.; Iwig, J.S.; Chivers, P.T.; Maroney, M.J. A dynamic Zn site in Helicobacter pylori HypA: A potential mechanism for metal-specific protein activity. J. Am. Chem. Soc. 2007, 129, 16–17. [Google Scholar] [CrossRef]
- Blum, F.C.; Hu, H.Q.; Servetas, S.L.; Benoit, S.L.; Maier, R.J.; Maroney, M.J.; Merrell, D.S. Structure-function analyses of metal-binding sites of HypA reveal residues important for hydrogenase maturation in Helicobacter pylori. PLoS ONE 2017, 12, e0183260. [Google Scholar] [CrossRef]
- Xia, W.; Li, H.; Yang, X.; Wong, K.B.; Sun, H. Metallo-GTPase HypB from Helicobacter pylori and its interaction with nickel chaperone protein HypA. J. Biol. Chem. 2012, 287, 6753–6763. [Google Scholar] [CrossRef]
- Kwon, S.; Watanabe, S.; Nishitani, Y.; Kawashima, T.; Kanai, T.; Atomi, H.; Miki, K. Crystal structures of a [NiFe] hydrogenase large subunit HyhL in an immature state in complex with a Ni chaperone HypA. Proc. Natl. Acad. Sci. USA 2018, 115, 7045–7050. [Google Scholar] [CrossRef] [Green Version]
- Leipe, D.D.; Wolf, Y.I.; Koonin, E.V.; Aravind, L. Classification and evolution of P-loop GTPases and related ATPases. J. Mol. Biol. 2002, 317, 41–72. [Google Scholar] [CrossRef] [Green Version]
- Leach, M.R.; Sandal, S.; Sun, H.; Zamble, D.B. Metal binding activity of the Escherichia coli hydrogenase maturation factor HypB. Biochemistry 2005, 44, 12229–12238. [Google Scholar] [CrossRef]
- Dias, A.V.; Mulvihill, C.M.; Leach, M.R.; Pickering, I.J.; George, G.N.; Zamble, D.B. Structural and biological analysis of the metal sites of Escherichia coli hydrogenase accessory protein HypB. Biochemistry 2008, 47, 11981–11991. [Google Scholar] [CrossRef]
- Sydor, A.M.; Liu, J.; Zamble, D.B. Effects of metal on the biochemical properties of Helicobacter pylori HypB, a maturation factor of [NiFe]-hydrogenase and urease. J. Bacteriol. 2011, 193, 1359–1368. [Google Scholar] [CrossRef]
- Sasaki, D.; Watanabe, S.; Matsumi, R.; Shoji, T.; Yasukochi, A.; Tagashira, K.; Fukuda, W.; Kanai, T.; Atomi, H.; Imanaka, T.; et al. Identification and structure of a novel archaeal HypB for [NiFe] hydrogenase maturation. J. Mol. Biol. 2013, 425, 1627–1640. [Google Scholar] [CrossRef]
- Gasper, R.; Scrima, A.; Wittinghofer, A. Structural insights into HypB, a GTP-binding protein that regulates metal binding. J. Biol. Chem. 2006, 281, 27492–27502. [Google Scholar] [CrossRef]
- Sydor, A.M.; Lebrette, H.; Ariyakumaran, R.; Cavazza, C.; Zamble, D.B. Relationship between Ni(II) and Zn(II) coordination and nucleotide binding by the Helicobacter pylori [NiFe]-hydrogenase and urease maturation factor HypB. J. Biol. Chem. 2014, 289, 3828–3841. [Google Scholar] [CrossRef]
- Maier, T.; Lottspeich, F.; Böck, A. GTP hydrolysis by HypB is essential for nickel insertion into hydrogenases of Escherichia coli. Eur. J. Biochem. 1995, 230, 133–138. [Google Scholar] [CrossRef]
- Maier, T.; Jacobi, A.; Sauter, M.; Böck, A. The product of the hypB gene, which is required for nickel incorporation into hydrogenases, is a novel guanine nucleotide-binding protein. J. Bacteriol. 1993, 175, 630–635. [Google Scholar] [CrossRef]
- Lacasse, M.J.; Douglas, C.D.; Zamble, D.B. Mechanism of selective nickel transfer from HypB to HypA, Escherichia coli [NiFe]-hydrogenase accessory proteins. Biochemistry 2016, 55, 6821–6831. [Google Scholar] [CrossRef]
- Chan Chung, K.C.; Cao, L.; Dias, A.V.; Pickering, I.J.; George, G.N.; Zamble, D.B. A high-affinity metal-binding peptide from Escherichia coli HypB. J. Am. Chem. Soc. 2008, 130, 14056–14057. [Google Scholar] [CrossRef]
- Chan, K.-H.; Li, T.; Wong, C.-O.; Wong, K.-B. Structural basis for GTP-dependent dimerization of hydrogenase maturation factor HypB. PLoS ONE 2012, 7, e30547. [Google Scholar] [CrossRef]
- Cai, F.; Ngu, T.T.; Kaluarachchi, H.; Zamble, D.B. Relationship between the GTPase, metal-binding, and dimerization activities of E. coli HypB. J. Biol. Inorg. Chem. 2011, 16, 857–868. [Google Scholar] [CrossRef]
- Fu, C.; Olson, J.W.; Maier, R.J. HypB protein of Bradyrhizobium japonicum is a metal-binding GTPase capable of binding 18 divalent nickel ions per dimer. Proc. Natl. Acad. Sci. USA 1995, 92, 2333–2337. [Google Scholar] [CrossRef]
- Olson, J.W.; Fu, C.; Maier, R.J. The HypB protein from Bradyrhizobium japonicum can store nickel and is required for the nickel-dependent transcriptional regulation of hydrogenase. Mol. Microbiol. 1997, 24, 119–128. [Google Scholar] [CrossRef]
- Rey, L.; Imperial, J.; Palacios, J.M.; Ruiz-Argüeso, T. Purification of Rhizobium leguminosarum HypB, a nickel-binding protein required for hydrogenase synthesis. J. Bacteriol. 1994, 176, 6066–6073. [Google Scholar] [CrossRef]
- Olson, J.W.; Maier, R.J. Dual roles of Bradyrhizobium japonicum nickel in protein in nickel storage and GTP-dependent Ni mobilization. J. Bacteriol. 2000, 182, 1702–1705. [Google Scholar] [CrossRef]
- Kovermann, M.; Schmid, F.X.; Balbach, J. Molecular function of the prolyl cis/trans isomerase and metallochaperone SlyD. Biol. Chem. 2013, 394, 965–975. [Google Scholar] [CrossRef]
- Martino, L.; He, Y.; Hands-Taylor, K.L.; Valentine, E.R.; Kelly, G.; Giancola, C.; Conte, M.R. The interaction of the Escherichia coli protein SlyD with nickel ions illuminates the mechanism of regulation of its peptidyl-prolyl isomerase activity. FEBS J. 2009, 276, 4529–4544. [Google Scholar] [CrossRef]
- Weininger, U.; Haupt, C.; Schweimer, K.; Graubner, W.; Kovermann, M.; Bruser, T.; Scholz, C.; Schaarschmidt, P.; Zoldak, G.; Schmid, F.X.; et al. NMR solution structure of SlyD from Escherichia coli: Spatial separation of prolyl isomerase and chaperone function. J. Mol. Biol. 2009, 387, 295–305. [Google Scholar] [CrossRef]
- Schmidpeter, P.A.; Ries, L.K.; Theer, T.; Schmid, F.X. Prolyl isomerization and its catalysis in protein folding and protein function. J. Mol. Biol. 2015, 427, 1609–1631. [Google Scholar] [CrossRef]
- Kaluarachchi, H.; Altenstein, M.; Sugumar, S.R.; Balbach, J.; Zamble, D.B.; Haupt, C. Nickel binding and [NiFe]-hydrogenase maturation by the metallochaperone SlyD with a single metal-binding site in Escherichia coli. J. Mol. Biol. 2012, 417, 28–35. [Google Scholar] [CrossRef]
- Löw, C.; Neumann, P.; Tidow, H.; Weininger, U.; Haupt, C.; Friedrich-Epler, B.; Scholz, C.; Stubbs, M.T.; Balbach, J. Crystal structure determination and functional characterization of the metallochaperone SlyD from Thermus thermophilus. J. Mol. Biol. 2010, 398, 375–390. [Google Scholar] [CrossRef]
- Quistgaard, E.M.; Weininger, U.; Ural-Blimke, Y.; Modig, K.; Nordlund, P.; Akke, M.; Löw, C. Molecular insights into substrate recognition and catalytic mechanism of the chaperone and FKBP peptidyl-prolyl isomerase SlyD. BMC Biol. 2016, 14, 82. [Google Scholar] [CrossRef]
- Cheng, T.; Li, H.; Xia, W.; Sun, H. Multifaceted SlyD from Helicobacter pylori: Implication in [NiFe] hydrogenase maturation. J. Biol. Inorg. Chem. 2012, 17, 331–343. [Google Scholar] [CrossRef]
- Kaluarachchi, H.; Sutherland, D.E.; Young, A.; Pickering, I.J.; Stillman, M.J.; Zamble, D.B. The Ni(II)-binding properties of the metallochaperone SlyD. J. Am. Chem. Soc. 2009, 131, 18489–18500. [Google Scholar] [CrossRef]
- Hottenrott, S.; Schumann, T.; Plückthun, A.; Fischer, G.; Rahfeld, J.U. The Escherichia coli SlyD is a metal ion-regulated peptidyl-prolyl cis/trans-isomerase. J. Biol. Chem. 1997, 272, 15697–15701. [Google Scholar] [CrossRef]
- Kaluarachchi, H.; Zhang, J.W.; Zamble, D.B. Escherichia coli SlyD, more than a Ni(II) reservoir. Biochemistry 2011, 50, 10761–10763. [Google Scholar] [CrossRef]
- Lee, M.H.; Pankratz, H.S.; Wang, S.; Scott, R.A.; Finnegan, M.G.; Johnson, M.K.; Ippolito, J.A.; Christianson, D.W.; Hausinger, R.P. Purification and characterization of Klebsiella aerogenes UreE protein: A nickel-binding protein that functions in urease metallocenter assembly. Protein Sci. 1993, 2, 1042–1052. [Google Scholar] [CrossRef]
- Mulrooney, S.B.; Hausinger, R.P. Sequence of the Klebsiella aerogenes urease genes and evidence for accessory proteins facilitating nickel incorporation. J. Bacteriol. 1990, 172, 5837–5843. [Google Scholar] [CrossRef]
- Zambelli, B.; Banaszak, K.; Merloni, A.; Kiliszek, A.; Rypniewski, W.; Ciurli, S. Selectivity of Ni(II) and Zn(II) binding to Sporosarcina pasteurii UreE, a metallochaperone in the urease assembly: A calorimetric and crystallographic study. J. Biol. Inorg. Chem. 2013, 18, 1005–1017. [Google Scholar] [CrossRef]
- Benoit, S.; Maier, R.J. Dependence of Helicobacter pylori urease activity on the nickel-sequestering ability of the UreE accessory protein. J. Bacteriol. 2003, 185, 4787–4795. [Google Scholar] [CrossRef]
- Bellucci, M.; Zambelli, B.; Musiani, F.; Turano, P.; Ciurli, S. Helicobacter pylori UreE, a urease accessory protein: Specific Ni2+- and Zn2+-binding properties and interaction with its cognate UreG. Biochem. J. 2009, 422, 91–100. [Google Scholar] [CrossRef]
- Banaszak, K.; Martin-Diaconescu, V.; Bellucci, M.; Zambelli, B.; Rypniewski, W.; Maroney, M.J.; Ciurli, S. Crystallographic and X-ray absorption spectroscopic characterization of Helicobacter pylori UreE bound to Ni2+ and Zn2+ reveals a role for the disordered C-terminal arm in metal trafficking. Biochem. J. 2012, 441, 1017–1035. [Google Scholar] [CrossRef]
- Stola, M.; Musiani, F.; Mangani, S.; Turano, P.; Safarov, N.; Zambelli, B.; Ciurli, S. The nickel site of Bacillus pasteurii UreE, a urease metallo-chaperone, as revealed by metal-binding studies and X-ray absorption spectroscopy. Biochemistry 2006, 45, 6495–6509. [Google Scholar] [CrossRef]
- Ciurli, S.; Safarov, N.; Miletti, S.; Dikiy, A.; Christensen, S.K.; Kornetzky, K.; Bryant, D.A.; Vandenberghe, I.; Devreese, B.; Samyn, B.; et al. Molecular characterization of Bacillus pasteurii UreE, a metal-binding chaperone for the assembly of the urease active site. J. Biol. Inorg. Chem. 2002, 7, 623–631. [Google Scholar] [CrossRef]
- Brayman, T.G.; Hausinger, R.P. Purification, characterization, and functional analysis of a truncated Klebsiella aerogenes UreE urease accessory protein lacking the histidine-rich carboxyl terminus. J. Bacteriol. 1996, 178, 5410–5416. [Google Scholar] [CrossRef]
- Colpas, G.J.; Hausinger, R.P. In vivo and in vitro kinetics of metal transfer by the Klebsiella aerogenes urease nickel metallochaperone, UreE. J. Biol. Chem. 2000, 275, 10731–10737. [Google Scholar] [CrossRef]
- Colpas, G.J.; Brayman, T.G.; McCracken, J.; Pressler, M.A.; Babcock, G.T.; Ming, L.J.; Colangelo, C.M.; Scott, R.A.; Hausinger, R.P. Spectroscopic characterization of metal binding by Klebsiella aerogenes UreE urease accessory protein. J. Biol. Inorg. Chem. 1998, 3, 150–160. [Google Scholar] [CrossRef]
- Colpas, G.J.; Brayman, T.G.; Ming, L.J.; Hausinger, R.P. Identification of metal-binding residues in the Klebsiella aerogenes urease nickel metallochaperone, UreE. Biochemistry 1999, 38, 4078–4088. [Google Scholar] [CrossRef]
- Grossoehme, N.E.; Mulrooney, S.B.; Hausinger, R.P.; Wilcox, D.E. Thermodynamics of Ni2+, Cu2+, and Zn2+ binding to the urease metallochaperone UreE. Biochemistry 2007, 46, 10506–10516. [Google Scholar] [CrossRef]
- Soriano, A.; Colpas, G.J.; Hausinger, R.P. UreE stimulation of GTP-dependent urease activation in the UreD-UreF-UreG-urease apoprotein complex. Biochemistry 2000, 39, 12435–12440. [Google Scholar] [CrossRef]
- Song, H.K.; Mulrooney, S.B.; Huber, R.; Hausinger, R.P. Crystal structure of Klebsiella aerogenes UreE, a bickel-binding metallochaperone for urease activation. J. Biol. Chem. 2001, 276, 49359–49364. [Google Scholar] [CrossRef]
- Remaut, H.; Safarov, N.; Ciurli, S.; Van Beeumen, J. Structural Basis for Ni2+ transport and assembly of the urease active site by the metallochaperone UreE from Bacillus pasteurii. J. Biol. Chem. 2001, 276, 49365–49370. [Google Scholar] [CrossRef]
- Shi, R.; Munger, C.; Asinas, A.; Benoit, S.L.; Miller, E.; Matte, A.; Maier, R.J.; Cygler, M. Crystal structures of apo and metal-bound forms of the UreE protein from Helicobacter pylori: Role of multiple metal binding sites. Biochemistry 2010, 49, 7080–7088. [Google Scholar] [CrossRef]
- Khorasani-Motlagh, M.; Noroozifar, M.; Kerman, K.; Zamble, D.B. Complex formation between the Escherichia coli [NiFe]-hydrogenase nickel maturation factors. BioMetals 2019, 32, 521–532. [Google Scholar] [CrossRef]
- Leach, M.R.; Zhang, J.W.; Zamble, D.B. The role of complex formation between the Escherichia coli hydrogenase accessory factors HypB and SlyD. J. Biol. Chem. 2007, 282, 16177–16186. [Google Scholar] [CrossRef]
- Stingl, K.; Schauer, K.; Ecobichon, C.; Labigne, A.; Lenormand, P.; Rousselle, J.-C.; Namane, A.; de Reuse, H. In vivo interactome of Helicobacter pylori urease revealed by tandem affinity purification. Mol. Cell. Proteomics 2008, 7, 2429–2441. [Google Scholar] [CrossRef]
- Cheng, T.; Li, H.; Yang, X.; Xia, W.; Sun, H. Interaction of SlyD with HypB of Helicobacter pylori facilitates nickel trafficking. Metallomics 2013, 5, 804–807. [Google Scholar] [CrossRef]
- Chan Chung, K.C.; Zamble, D.B. The Escherichia coli metal-binding chaperone SlyD interacts with the large subunit of [NiFe]-hydrogenase 3. FEBS Lett. 2011, 585, 291–294. [Google Scholar] [CrossRef]
- Chan Chung, K.C.; Zamble, D.B. Protein Interactions and localization of the Escherichia coli accessory protein HypA during nickel insertion to [NiFe] hydrogenase. J. Biol. Chem. 2011, 286, 43081–44390. [Google Scholar] [CrossRef]
- Yang, X.; Li, H.; Cheng, T.; Xia, W.; Lai, Y.-T.; Sun, H. Nickel translocation between metallochaperones HypA and UreE in Helicobacter pylori. Metallomics 2014, 6, 1731–1736. [Google Scholar] [CrossRef]
- Yang, X.; Li, H.; Lai, T.P.; Sun, H. UreE-UreG complex facilitates nickel transfer and preactivates GTPase of UreG in Helicobacter pylori. J. Biol. Chem. 2015, 290, 12474–12485. [Google Scholar] [CrossRef]
- Fong, Y.H.; Wong, H.C.; Yuen, M.H.; Lau, P.H.; Chen, Y.W.; Wong, K.-B. Structure of UreG/UreF/UreH complex reveals how urease accessory proteins facilitate maturation of Helicobacter pylori Urease. PLoS Biol. 2013, 11, e1001678. [Google Scholar] [CrossRef]
- Moncrief, M.B.; Hausinger, R.P. Purification and activation properties of UreD-UreF-urease apoprotein complexes. J. Bacteriol. 1996, 178, 5417–5421. [Google Scholar] [CrossRef] [Green Version]
- Soriano, A.; Hausinger, R.P. GTP-dependent activation of urease apoprotein in complex with the UreD, UreF, and UreG accessory proteins. Proc. Natl. Acad. Sci. USA 1999, 96, 11140–11144. [Google Scholar] [CrossRef] [Green Version]
- Giedroc, D.P.; Arunkumar, A.I. Metal sensor proteins: Nature’s metalloregulated allosteric switches. Dalt. Trans. 2007, 29, 3107–3120. [Google Scholar] [CrossRef]
- Eaton, K.A.; Krakowka, S. Effect of gastric pH on urease-dependent colonization of gnotobiotic piglets by Helicobacter pylori. Infect. Immun. 1994, 62, 3604–3607. [Google Scholar]
- Eaton, K.A.; Brooks, C.L.; Morgan, D.R.; Krakowka, S. Essential role of urease in pathogenesis of gastritis induced by Helicobacter pylori in gnotobiotic piglets. Infect. Immun. 1991, 59, 2470–2475. [Google Scholar]














© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Higgins, K. Nickel Metalloregulators and Chaperones. Inorganics 2019, 7, 104. https://doi.org/10.3390/inorganics7080104
Higgins K. Nickel Metalloregulators and Chaperones. Inorganics. 2019; 7(8):104. https://doi.org/10.3390/inorganics7080104
Chicago/Turabian StyleHiggins, Khadine. 2019. "Nickel Metalloregulators and Chaperones" Inorganics 7, no. 8: 104. https://doi.org/10.3390/inorganics7080104
APA StyleHiggins, K. (2019). Nickel Metalloregulators and Chaperones. Inorganics, 7(8), 104. https://doi.org/10.3390/inorganics7080104