From Polyclonal Sera to Recombinant Antibodies: A Review of Immunological Detection of Gluten in Foodstuff


Abstract
:1. Introduction
2. Gluten-Related Diseases
3. Gluten Content Labeling Legislation in Different Countries
4. Methods for Gluten Detection in Food Samples
- ELISA [41]: this is a quick, economic, versatile and robust method. ELISA presents a high sensitivity (in the low ppm range) and optical detection. However, there is a possibility for some false negatives, due to protein denaturalization, and there is some risk of false positives due to cross-reactions with similar but non-target proteins.
- Immunochromatographic assays [42]: these have a visual and simple result interpretation, and they are very simple to use by final operator. Their main weakness is that they cannot quantify.
- Western Blot [43]: this is a highly specific and sensitive method (low ppm), with additional strengths, like confirmatory values (molecular weights) and highly efficient insoluble protein detection. Nevertheless, it is a time-consuming method that must be performed by qualified personnel.
- Chromatography methods [44]: these are based on the separation and detection of peptides with a very high sensitivity. The main drawback is that they require complex and expensive instrumentation.
- Mass spectrometry [45]: this is a quick, reproducible, and very precise method of analysis that allows for species detection. However, it is not a quantitative method, and it requires complex and expensive instrumentation.
5. Immunological-Based Techniques for Gluten Detection in Food Samples
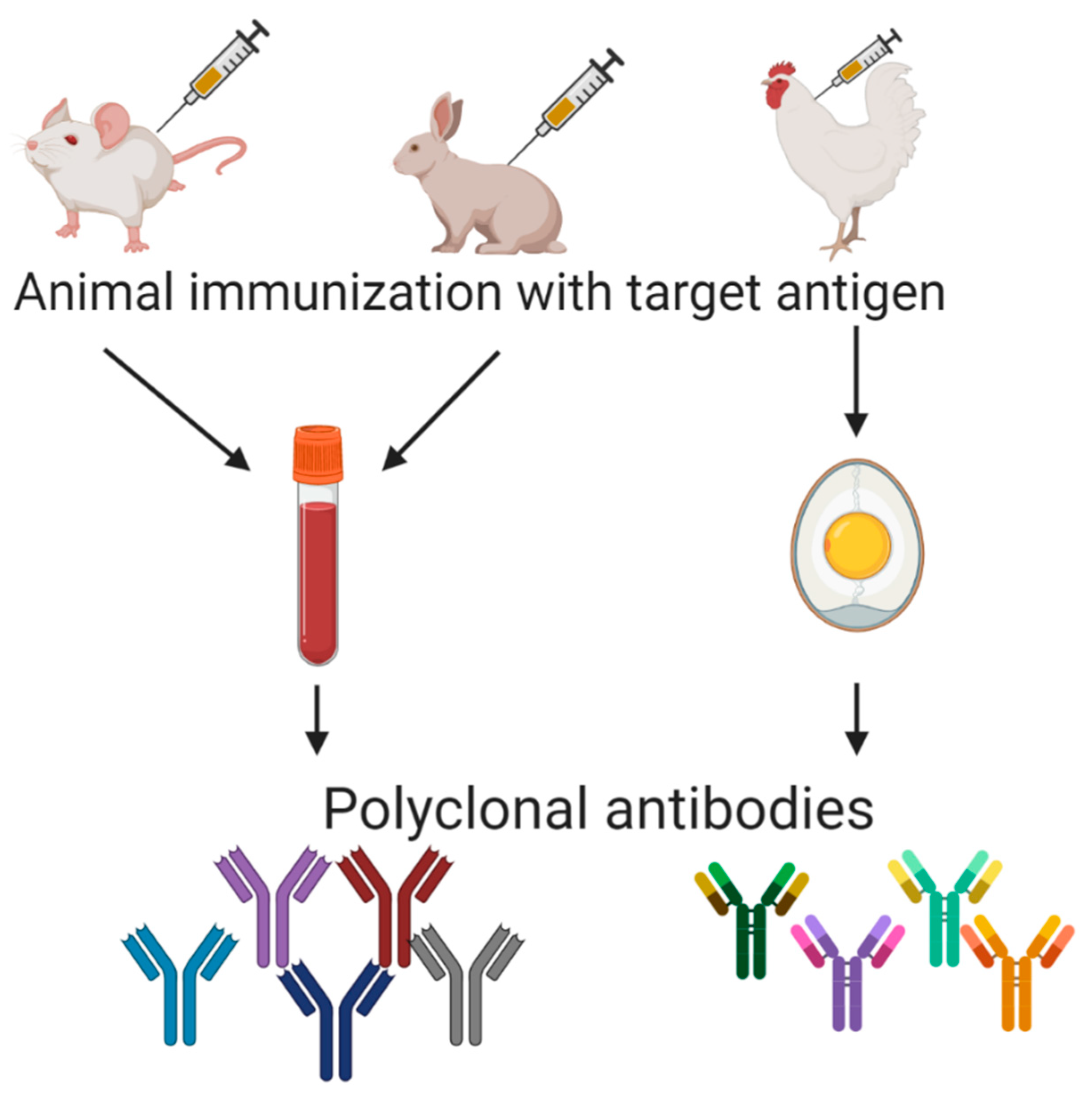
6. Polyclonal Antibodies
7. Hybridoma Secreted Monoclonal Antibodies
8. A New Era: Directed Evolution of Recombinant Antibodies
- Immune: constructed based on amplifications of variable (V) genes isolated from immunoglobulin-secreting plasma cells from immunized donors [86].
- Naïve: based on amplifications of V genes isolated from immunoglobulin-secreting plasma cells from non-immunized donors [87].
- Semi-synthetic: derived from unrearranged V genes from pre-B cells (germline cells) or a single antibody framework with genetically randomized complementarity determining regions (CDRs) [88].
- Synthetic: based on a human antibody framework with randomly integrated CDR cassettes [89].
9. Final Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fasano, A.; Catassi, C. Clinical practice. Celiac disease. N. Engl. J. Med. 2012, 367, 2419–2426. [Google Scholar] [CrossRef]
- Codex Alimentarius. Codex Standard for Foods for Special Dietary Use for Persons Intolerant to Gluten. Revised 2008. CODEX Standard 118-1979. Available online: http://www.fao.org/fao-who-codexalimentarius/sh-proxy/es/?lnk=1&url=https%253A%252F%252Fworkspace.fao.org%252Fsites%252Fcodex%252FStandards%252FCXS%2B118-1979%252FCXS_118e_2015.pdf (accessed on 29 December 2020).
- Goesaert, H.; Brijs, K.; Veraverbeke, W.S.; Courtin, C.M.; Gebruers, K.; Delcour, J.A. Wheat flour constituents: How they impact bread quality, and how to impact their functionality. Trends Food Sci. Technol. 2005, 16, 12–30. [Google Scholar] [CrossRef]
- Martín, M.; Cacho, J.F.; Cepeda, A.; Martín, F.; Prieto, I. Informe del Comité Científico de la Agencia Española de Seguridad Alimentaria y Nutrición (AESAN) en Relación con la Enfermedad Celíaca y los Problemas que Plantean las Técnicas Analíticas para el Control del Contenido de Gluten en los Alimentos; AESAN: Madrid, Spain, 2010. [Google Scholar]
- Shewry, P.R.; Halford, N.G.; Tatham, A.S.; Popineau, Y.; Lafiandra, D.; Belton, P.S. The high molecular weight subunits of wheat glutenin and their role in determining wheat processing properties. Adv. Food Nutr. Res. 2003, 45, 219–302. [Google Scholar] [CrossRef] [PubMed]
- Cebolla, Á.; Moreno, M.; Coto, L.; Sousa, C. Gluten Immunogenic Peptides as Standard for the Evaluation of Potential Harmful Prolamin Content in Food and Human Specimen. Nutrients 2018, 10, 1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P. What Is Gluten—Why Is It Special? Front. Nutr. 2019, 6, 101. [Google Scholar] [CrossRef] [PubMed]
- Ozuna, C.V.; Iehisa, J.C.M.; Giménez, M.J.; Alvarez, J.B.; Sousa, C.; Barro, F. Diversification of the celiac disease α-gliadin complex in wheat: A 33-mer peptide with six overlapping epitopes, evolved following polyploidization. Plant J. 2015, 82, 794–805. [Google Scholar] [CrossRef] [PubMed]
- Schalk, K.; Lang, C.; Wieser, H.; Koehler, P.; Scherf, K.A. Quantitation of the immunodominant 33-mer peptide from α-gliadin in wheat flours by liquid chromatography tandem mass spectrometry. Sci. Rep. 2017, 7, 45092. [Google Scholar] [CrossRef]
- Balakireva, A.; Zamyatnin, A. Properties of Gluten Intolerance: Gluten Structure, Evolution, Pathogenicity and Detoxification Capabilities. Nutrients 2016, 8, 644. [Google Scholar] [CrossRef] [Green Version]
- Matsuo, H.; Yokooji, T.; Taogoshi, T. Common food allergens and their IgE-binding epitopes. Allergol. Int. 2015, 64, 332–343. [Google Scholar] [CrossRef] [Green Version]
- Caio, G.; Volta, U.; Sapone, A.; Leffler, D.A.; De Giorgio, R.; Catassi, C.; Fasano, A. Celiac disease: A comprehensive current review. BMC Med. 2019, 17, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapone, A.; Bai, J.C.; Ciacci, C.; Dolinsek, J.; Green, P.H.; Hadjivassiliou, M.; Kaukinen, K.; Rostami, K.; Sanders, D.S.; Schumann, M.; et al. Spectrum of gluten-related disorders: Consensus on new nomenclature and classification. BMC Med. 2012, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomares, O.; Akdis, M.; Martín-Fontecha, M.; Akdis, C.A. Mechanisms of immune regulation in allergic diseases: The role of regulatory T and B cells. Immunol. Rev. 2017, 278, 219–236. [Google Scholar] [CrossRef] [PubMed]
- Al Badri, F.M.; Baatjies, R.; Jeebhay, M.F. Assessing the health impact of interventions for baker’s allergy and asthma in supermarket bakeries: A group randomised trial. Int. Arch. Occup. Environ. Health 2020, 93, 589–599. [Google Scholar] [CrossRef]
- Sharma, N.; Bhatia, S.; Chunduri, V.; Kaur, S.; Sharma, S.; Kapoor, P.; Kumari, A.; Garg, M. Pathogenesis of Celiac Disease and Other Gluten Related Disorders in Wheat and Strategies for Mitigating Them. Front. Nutr. 2020, 7, 6. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Wen, L.; Li, H.; Wang, R.; Yin, J. Genetic variation at the interleukin-18 locus is associated with wheat-dependent exercise-induced anaphylaxis in the Han Chinese population. Gene 2020, 737, 144462. [Google Scholar] [CrossRef]
- Hollon, J.; Puppa, E.; Greenwald, B.; Goldberg, E.; Guerrerio, A.; Fasano, A. Effect of Gliadin on Permeability of Intestinal Biopsy Explants from Celiac Disease Patients and Patients with Non-Celiac Gluten Sensitivity. Nutrients 2015, 7, 1565–1576. [Google Scholar] [CrossRef]
- Kim, S.M.; Mayassi, T.; Jabri, B. Innate immunity: Actuating the gears of celiac disease pathogenesis. Best Pract. Res. Clin. Gastroenterol. 2015, 29, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Junker, Y.; Zeissig, S.; Kim, S.-J.; Barisani, D.; Wieser, H.; Leffler, D.A.; Zevallos, V.; Libermann, T.A.; Dillon, S.; Freitag, T.L.; et al. Wheat amylase trypsin inhibitors drive intestinal inflammation via activation of toll-like receptor 4. J. Exp. Med. 2012, 209, 2395–2408. [Google Scholar] [CrossRef]
- Stamnaes, J.; Sollid, L.M. Celiac disease: Autoimmunity in response to food antigen. Semin. Immunol. 2015, 27, 343–352. [Google Scholar] [CrossRef]
- Pagliari, D.; Urgesi, R.; Frosali, S.; Riccioni, M.E.; Newton, E.E.; Landolfi, R.; Pandolfi, F.; Cianci, R. The Interaction among Microbiota, Immunity, and Genetic and Dietary Factors Is the Condicio Sine Qua Non Celiac Disease Can Develop. J. Immunol. Res. 2015, 2015, 123653. [Google Scholar] [CrossRef] [PubMed]
- Schumann, M.; Richter, J.F.; Wedell, I.; Moos, V.; Zimmermann-Kordmann, M.; Schneider, T.; Daum, S.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Mechanisms of epithelial translocation of the alpha(2)-gliadin-33mer in coeliac sprue. Gut 2008, 57, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Hadjivassiliou, M.; Sanders, D.S.; Woodroofe, N.; Williamson, C.; Grünewald, R.A. Gluten ataxia. Cerebellum 2008, 7, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Volta, U.; De Giorgio, R.; Caio, G.; Uhde, M.; Manfredini, R.; Alaedini, A. Nonceliac Wheat Sensitivity: An Immune-Mediated Condition with Systemic Manifestations. Gastroenterol. Clin. N. Am. 2019, 48, 165–182. [Google Scholar] [CrossRef]
- Roszkowska, A.; Pawlicka, M.; Mroczek, A.; Bałabuszek, K.; Nieradko-Iwanicka, B. Non-Celiac Gluten Sensitivity: A Review. Medicina 2019, 55, 222. [Google Scholar] [CrossRef] [Green Version]
- Kurada, S.; Yadav, A.; Leffler, D.A. Current and novel therapeutic strategies in celiac disease. Expert Rev. Clin. Pharmacol. 2016, 9, 1211–1223. [Google Scholar] [CrossRef]
- Itzlinger, A.; Branchi, F.; Elli, L.; Schumann, M. Gluten-Free Diet in Celiac Disease—Forever and for All? Nutrients 2018, 10, 1796. [Google Scholar] [CrossRef] [Green Version]
- Ludvigsson, J.F.; Bai, J.C.; Biagi, F.; Card, T.R.; Ciacci, C.; Ciclitira, P.J.; Green, P.H.R.; Hadjivassiliou, M.; Holdoway, A.; Van Heel, D.A.; et al. Diagnosis and management of adult coeliac disease: Guidelines from the British Society of Gastroenterology. Gut 2014, 63, 1210–1228. [Google Scholar] [CrossRef]
- Commission Implementing Regulation (EU) No. 828/2014 of 30 July on the Requirements for the Provision of Information to Consumers on the Absence or Reduced Presence of Gluten in Food OJ L 228, 31.7.2014. pp. 5–8. Available online: https://eur-lex.europa.eu/eli/reg_impl/2014/828/oj (accessed on 29 December 2020).
- Food Allergen Labeling and Consumer Protection Act. Sec. 206 Regulations 21 USC 343. Title II of Public Law 108-282, Enacted on August 2, 2004. Available online: https://www.fda.gov/food/food-allergensgluten-free-guidance-documents-regulatory-information/food-allergen-labeling-and-consumer-protection-act-2004-falcpa (accessed on 29 December 2020).
- Scherf, K.A.; Poms, R.E. Recent developments in analytical methods for tracing gluten. J. Cereal Sci. 2016, 67, 112–122. [Google Scholar] [CrossRef]
- Australia New Zealand Food Standards Code–Standard 1.2.7–Nutrition, Health and Related Claims. F2018C00942. Available online: https://www.legislation.gov.au/Details/F2018C00942 (accessed on 29 December 2020).
- MODIFICACIÓN a la Norma Oficial Mexicana NOM-051-SCFI/SSA1-2010, Especificaciones Generales de Etiquetado para Alimentos y Bebidas no Alcohólicas Preenvasados-Información Comercial y Sanitaria, Publicada el 5 de abril de 2010. Diario Oficial de la Federación, 27 de Marzo de 2020. Available online: http://www.dof.gob.mx/normasOficiales/8150/seeco11_C/seeco11_C.html (accessed on 29 December 2020).
- Norma Oficial Mexicana NOM-247-SSA1-2008, Productos y Servicios. Cereales y sus Productos. Cereales, Harinas de Cereales, Sémolas o Semolinas. Alimentos a base de: Cereales, Semillas Comestibles, de Harinas, Sémolas o Semolinas o sus Mezclas. Productos de Panificación. Disposiciones y Especificaciones Sanitarias y Nutrimentales. Métodos de Prueba. Diario Oficial de la Federación, 27 de julio de 2009. Available online: http://dof.gob.mx/nota_detalle.php?codigo=5188649&fecha=10/05/2011 (accessed on 29 December 2020).
- Ley 27196. Enfermedad Celíaca Modificación Ley 26588. Boletín Oficial de la República Argentina No. 33259. Available online: https://www.boletinoficial.gob.ar/detalleAviso/primera/136317/20151118 (accessed on 29 December 2020).
- Lei No. 10674 de 16 de maio de 2003 Obriga a que os Produtos Aliementicios Comercializados Informen sobre a Presença de Gluten, como Medida Preventiva e de Controle da Doença Celiaca. Diário Oficial da União 19/5/2003. Available online: https://www.camara.leg.br/proposicoesWeb/prop_mostrarintegra?codteor=440852&filename=Legislacao (accessed on 29 December 2020).
- The China Food Law Blog. Available online: https://chinafoodlaw.blog/2018/10/09/gluten-free-in-china/ (accessed on 16 July 2020).
- Akiyama, H.; Imai, T.; Ebisawa, M. Japan Food Allergen Labeling Regulation—History and Evaluation; Elsevier: Amsterdam, The Netherlands, 2011; pp. 139–171. [Google Scholar] [CrossRef]
- Panda, R.; Garber, E.A.E. Detection and Quantitation of Gluten in Fermented-Hydrolyzed Foods by Antibody-Based Methods: Challenges, Progress, and a Potential Path Forward. Front. Nutr. 2019, 6, 97. [Google Scholar] [CrossRef]
- Ferre-lópez, S.; Ribes-koninckx, C.; Genzor, C.; Gamen, S.; Peña, L.; Ortigosa, L.; Méndez, E. Immunochromatographic sticks for tissue transglutaminase and antigliadin antibody screening in celiac disease. Clin. Gastroenterol. Hepatol. 2004, 2, 480–484. [Google Scholar] [CrossRef]
- Panda, R.; Garber, E.A.E. Western blot analysis of fermented-hydrolyzed foods utilizing gluten-specific antibodies employed in a novel multiplex competitive ELISA. Anal. Bioanal. Chem. 2019, 411, 5159–5174. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bose, U.; Stockwell, S.; Howitt, C.A.; Colgrave, M. Assessing the Utility of Multiplexed Liquid Chromatography-Mass Spectrometry for Gluten Detection in Australian Breakfast Food Products. Molecules 2019, 24, 3665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boukid, F.; Prandi, B.; Faccini, A.; Sforza, S. A Complete Mass Spectrometry (MS)-Based Peptidomic Description of Gluten Peptides Generated During In Vitro Gastrointestinal Digestion of Durum Wheat: Implication for Celiac Disease. J. Am. Soc. Mass Spectrom. 2019, 30, 1481–1490. [Google Scholar] [CrossRef]
- Martín-Fernández, B.; Costa, J.; Oliveira, M.B.P.P.; López-Ruiz, B.; Mafra, I. Screening new gene markers for gluten detection in foods. Food Control 2015, 56, 57–63. [Google Scholar] [CrossRef]
- Martín-Fernández, B.; Costa, J.; Oliveira, M.B.P.P.; López-Ruiz, B.; Mafra, I. Combined effects of matrix and gene marker on the real-time PCR detection of wheat. Int. J. Food Sci. Technol. 2016, 51, 1680–1688. [Google Scholar] [CrossRef]
- García-García, A.; Madrid, R.; Sohrabi, H.; De La Cruz, S.; García, T.; Martín, R.; González, I. A sensitive and specific real-time PCR targeting DNA from wheat, barley and rye to track gluten contamination in marketed foods. LWT Food Sci. Technol. 2019, 114, 108378. [Google Scholar] [CrossRef]
- Burton, D.R. Structure and function of antibodies. In New Comprehensive Biochemistry; Calabi, F., Neuberger, M.S., Eds.; Elsevier: Amsterdam, The Netherlands, 1987; Volume 17, pp. 1–50. [Google Scholar]
- Lee, W.; Syed Atif, A.; Tan, S.C.; Leow, C.H. Insights into the chicken IgY with emphasis on the generation and applications of chicken recombinant monoclonal antibodies. J. Immunol. Methods 2017, 447, 71–85. [Google Scholar] [CrossRef]
- Frenzel, A.; Hust, M.; Schirrmann, T. Expression of Recombinant Antibodies. Front. Immunol. 2013, 4, 217. [Google Scholar] [CrossRef] [Green Version]
- Muyldermans, S. Nanobodies: Natural Single-Domain Antibodies. Annu. Rev. Biochem. 2013, 82, 775–797. [Google Scholar] [CrossRef] [Green Version]
- Kohler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef] [PubMed]
- Köhler, G.; Milstein, C. Derivation of specific antibody-producing tissue culture and tumor lines by cell fusion. Eur. J. Immunol. 1976, 6, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Manohar, R.S. Development of Enzyme-Linked Immunosorbent Assay for Evaluation of Chapati-Making Quality of Wheat Varieties. J. Agric. Food Chem. 2002, 50, 7455–7460. [Google Scholar] [CrossRef] [PubMed]
- Gujral, N.; Suh, J.W.; Sunwoo, H.H. Effect of anti-gliadin IgY antibody on epithelial intestinal integrity and inflammatory response induced by gliadin. BMC Immunol. 2015, 16, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skerritt, J.; Underwood, P.A. Specificity characteristics of monoclonal antibodies to wheat grain storage proteins. Biochim. Biophys. Acta-Protein Struct. Mol. Enzymol. 1986, 874, 245–254. [Google Scholar] [CrossRef]
- Galfre, G.; Howe, S.C.; Milstein, C.; Butcher, G.W.; Howard, J.C. Antibodies to major histocompatibility antigens produced by hybrid cell lines. Nature 1977, 266, 550–552. [Google Scholar] [CrossRef]
- Skerritt, J.H.; Hill, A.S. Monoclonal antibody sandwich enzyme immunoassays for determination of gluten in foods. J. Agric. Food Chem. 1990, 38, 1771–1778. [Google Scholar] [CrossRef]
- Sorell, L.; López, J.A.; Valdés, I.; Alfonso, P.; Camafeita, E.; Acevedo, B.; Chirdo, F.; Gavilondo, J.; Méndez, E. An innovative sandwich ELISA system based on an antibody cocktail for gluten analysis. Fed. Eur. Biochem. Soc. Lett. 1998, 439, 46–50. [Google Scholar] [CrossRef]
- Valdés, I.; García, E.; Llorente, M.; Méndez, E. Innovative approach to low-level gluten determination in foods using a novel sandwich enzyme-linked immunosorbent assay protocol. Eur. J. Gastroenterol. Hepatol. 2003, 15, 465–747. [Google Scholar] [CrossRef]
- Garcia, E.; Llorente, M.; Hernando, A.; Kieffer, R.; Wieser, H.; Mendez, E. Development of a general procedure for complete extraction of gliadins for heat processed and unheated foods. Eur. J. Gastroenterol. Hepatol. 2005, 17, 529–539. [Google Scholar] [CrossRef]
- Köhler, P.; Schwalb, T.; Immer, U.; Lacorn, M.; Wehling, P.; Don, C. AACCI Approved Methods Technical Committee Report: Collaborative Study on the Immunochemical Determination of Partially Hydrolyzed Gluten Using an R5 Competitive ELISA. Cereal Foods World 2013, 58, 113–117. [Google Scholar] [CrossRef]
- Méndez, E.; Vela, C.; Immer, U.; Janssen, F.W. Report of a collaborative trial to investigate the performance of the R5 enzyme linked immunoassay to determine gliadin in gluten-free food. Eur. J. Gastroenterol. Hepatol. 2005, 17, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Koerner, T.B.; Abbott, M.; Godefroy, S.B.; Popping, B.; Yeung, J.M.; Diaz-Amigo, C.; Roberts, J.; Taylor, S.L.; Baumert, J.L.; Ulberth, F.; et al. Validation Procedures for Quantitative Gluten ELISA Methods: AOAC Allergen Community Guidance and Best Practices. J. AOAC Int. 2013, 96, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Morón, B.; Cebolla, Á.; Manyani, H.; Álvarez-Maqueda, M.; Megías, M.; Thomas, M.D.C.; López, M.C.; Sousa, C. Sensitive detection of cereal fractions that are toxic to celiac disease patients by using monoclonal antibodies to a main immunogenic wheat peptide. Am. J. Clin. Nutr. 2008, 87, 405–414. [Google Scholar] [CrossRef] [Green Version]
- Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.; Sollid, L.; Khosla, C. Structural Basis for Gluten Intolerance in Celiac Sprue. Science 2002, 297, 2275–2279. [Google Scholar] [CrossRef] [Green Version]
- Morón, B.; Bethune, M.T.; Comino, I.; Manyani, H.; Ferragud, M.; López, M.C.; Cebolla, Á.; Khosla, C.; Sousa, C. Toward the Assessment of Food Toxicity for Celiac Patients: Characterization of Monoclonal Antibodies to a Main Immunogenic Gluten Peptide. PLoS ONE 2008, 3, e2294. [Google Scholar] [CrossRef] [Green Version]
- Cereals & Grains Association. Available online: http://methods.aaccnet.org/summaries/38-52-01.aspx (accessed on 18 October 2020).
- Official Methods of Analysis AOAC International. Available online: http://www.eoma.aoac.org/methods/info.asp?ID=50740 (accessed on 17 October 2020).
- Anderson, R.P.; Degano, P.; Godkin, A.J.; Jewell, D.P.; Hill, A.V.S. In vivo antigen challenge in celiac disease identifies a single transglutaminase-modified peptide as the dominant A-gliadin T-cell epitope. Nat. Med. 2000, 6, 337–342. [Google Scholar] [CrossRef]
- Arentz-Hansen, H.; Körner, R.; Molberg, Ø.; Quarsten, H.; Vader, W.; Kooy, Y.M.C.; Lundin, K.E.A.; Koning, F.; Roepstorff, P.; Sollid, L.M.; et al. The Intestinal T Cell Response to α-Gliadin in Adult Celiac Disease Is Focused on a Single Deamidated Glutamine Targeted by Tissue Transglutaminase. J. Exp. Med. 2000, 191, 603–612. [Google Scholar] [CrossRef]
- Stern, M.; Ciclitira, P.J.; van Eckert, R.; Feighery, C.; Janssen, F.W.; Méndez, E.; Mothes, T.; Troncone, R.; Wieser, H. Analysis and clinical effects of gluten in coeliac disease. Eur. J. Gastroenterol. Hepatol. 2001, 13, 741–747. [Google Scholar] [CrossRef]
- Mitea, C.; Kooy-Winkelaar, Y.; Van Veelen, P.; De Ru, A.; Drijfhout, J.W.; Koning, F.; Dekking, L. Fine specificity of monoclonal antibodies against celiac disease–inducing peptides in the gluteome. Am. J. Clin. Nutr. 2008, 88, 1057–1066. [Google Scholar] [CrossRef] [Green Version]
- Mujico, J.R.; Dekking, L.; Kooy-Winkelaar, Y.; Verheijen, R.; Wichen, P.V.; Streppel, L.; Sajic, N.; Drijfhout, J.W.; Koning, F. Validation of a New Enzyme-Linked Immunosorbent Assay to Detect the Triggering Proteins and Peptides for Celiac Disease: Interlaboratory Study. J. AOAC Int. 2019, 95, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Tranquet, O.; Larré, C.; Denery-Papini, S. Allergic reactions to hydrolysed wheat proteins: Clinical aspects and molecular structures of the allergens involved. Crit. Rev. Food Sci. Nutr. 2020, 60, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Tranquet, O.; Lupi, R.; Echasserieau-Laporte, V.; Pietri, M.; Larré, C.; Denery-Papini, S. Characterization of Antibodies and Development of an Indirect Competitive Immunoassay for Detection of Deamidated Gluten. J. Agric. Food Chem. 2015, 63, 5403–5409. [Google Scholar] [CrossRef]
- Tranquet, O.; Gaudin, J.-C.; Patil, S.; Steinbrecher, J.; Matsunaga, K.; Teshima, R.; Sakai, S.; Larré, C.; Denery-Papini, S. A chimeric IgE that mimics IgE from patients allergic to acid-hydrolyzed wheat proteins is a novel tool for in vitro allergenicity assessment of functionalized glutens. PLoS ONE 2017, 12, e0187415. [Google Scholar] [CrossRef]
- Winter, G. Harnessing Evolution to Make Medicines (Nobel Lecture). Angew. Chem. Int. Ed. 2019, 58, 14438–14445. [Google Scholar] [CrossRef] [Green Version]
- Meigs, L. Animal testing and its alternatives–the most important omics is economics. Altern. Anim. Exp. 2018, 35, 275–305. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.P.; Petrenko, V.A. Phage Display. Chem. Rev. 1997, 97, 391–410. [Google Scholar] [CrossRef] [PubMed]
- Arnold, F.H. Innovation by Evolution: Bringing New Chemistry to Life (Nobel Lecture). Angew. Chem. Int. Ed. 2019, 58, 14420–14426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyce, C.; Burton, D.R.; Briney, B. Comparisons of the antibody repertoires of a humanized rodent and humans by high throughput sequencing. Sci. Rep. 2020, 10, 1120. [Google Scholar] [CrossRef] [Green Version]
- Hozumi, N.; Tonegawa, S. Evidence for somatic rearrangement of immunoglobulin genes coding for variable and constant regions. Proc. Natl. Acad. Sci. USA 1976, 73, 3628–3632. [Google Scholar] [CrossRef] [Green Version]
- Hust, M. Mating antibody phage display with proteomics. Trends Biotechnol. 2004, 22, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Clackson, T.; Hoogenboom, H.R.; Griffiths, A.D.; Winter, G. Making antibody fragments using phage display libraries. Nature 1991, 352, 624–628. [Google Scholar] [CrossRef] [PubMed]
- De Haard, H.J.; Van Neer, N.; Reurs, A.; Hufton, S.E.; Roovers, R.C.; Henderikx, P.; De BruїNe, A.P.; Arends, J.-W.; Hoogenboom, H.R. A Large Non-immunized Human Fab Fragment Phage Library That Permits Rapid Isolation and Kinetic Analysis of High Affinity Antibodies. J. Biol. Chem. 1999, 274, 18218–18230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pini, A.; Viti, F.; Santucci, A.; Carnemolla, B.; Zardi, L.; Neri, P.; Neri, D. Design and Use of a Phage Display Library. J. Biol. Chem. 1998, 273, 21769–21776. [Google Scholar] [CrossRef] [Green Version]
- Wagner, H.; Wehrle, S.; Weiss, E.; Cavallari, M.; Weber, W. A Two-Step Approach for the Design and Generation of Nanobodies. Int. J. Mol. Sci. 2018, 19, 3444. [Google Scholar] [CrossRef] [Green Version]
- Böldicke, T. Single domain antibodies for the knockdown of cytosolic and nuclear proteins. Protein Sci. 2017, 26, 925–945. [Google Scholar] [CrossRef] [Green Version]
- Hoogenboom, H. Designing and optimizing library selection strategies for generating high-affinity antibodies. Trends Biotechnol. 1997, 15, 62–70. [Google Scholar] [CrossRef]
- Leow, C.H.; Fischer, K.; Leow, C.Y.; Cheng, Q.; Chuah, C.; McCarthy, J. Single Domain Antibodies as New Biomarker Detectors. Diagnostics 2017, 7, 52. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.P. Phage Display: Simple Evolution in a Petri Dish (Nobel Lecture). Angew. Chem. Int. Ed. 2019, 58, 14428–14437. [Google Scholar] [CrossRef] [Green Version]
- Plückthun, A. Ribosome Display: A Perspective; Springer: New York, NY, USA, 2012; pp. 3–28. [Google Scholar] [CrossRef]
- Yamaguchi, J.; Naimuddin, M.; Biyani, M.; Sasaki, T.; Machida, M.; Kubo, T.; Funatsu, T.; Husimi, Y.; Nemoto, N. cDNA display: A novel screening method for functional disulfide-rich peptides by solid-phase synthesis and stabilization of mRNA-protein fusions. Nucleic Acids Res. 2009, 37, e108. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Choi, J.H.; Xu, Z. Microbial cell-surface display. Trends Biotechnol. 2003, 21, 45–52. [Google Scholar] [CrossRef]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef] [PubMed]
- Parmley, S.F.; Smith, G.P. Antibody-selectable filamentous fd phage vectors: Affinity purification of target genes. Gene 1988, 73, 305–318. [Google Scholar] [CrossRef]
- De la Cruz, V.F.; Lal, A.A.; McCutchan, T.F. Immunogenicity and epitope mapping of foreign sequences via genetically engineered filamentous phage. J. Biol. Chem. 1988, 263, 4318–4322. [Google Scholar]
- Smith, G.P. Surface display and peptide libraries. Gene 1993, 128, 1–2. [Google Scholar] [CrossRef]
- Armstrong, N.; Adey, N.B.; McConnell, S.J.; Kay, B.K. CHAPTER 3-Vectors for Phage Display. In Phage Display of Peptides and Proteins; Kay, B.K., Winter, J., McCafferty, J., Eds.; Academic Press: Burlington, MA, USA, 1996; pp. 35–53. [Google Scholar] [CrossRef]
- Makowski, L. Structural constraints on the display of foreign peptides on filamentous bacteriophages. Gene 1993, 128, 5–11. [Google Scholar] [CrossRef]
- Lipovsek, D.; Plückthun, A. In-vitro protein evolution by ribosome display and mRNA display. J. Immunol. Methods 2004, 290, 51–67. [Google Scholar] [CrossRef]
- Hoischen, C.; Fritsche, C.; Gumpert, J.; Westermann, M.; Gura, K.; Fahnert, B. Novel Bacterial Membrane Surface Display System Using Cell Wall-Less L-Forms of Proteus mirabilis and Escherichia coli. Appl. Environ. Microbiol. 2002, 68, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Mesnage, S.P.; Weber-Levy, M.; Haustant, M.; Mock, M.L.; Fouet, A.S. Cell Surface-Exposed Tetanus Toxin Fragment C Produced by Recombinant Bacillus anthracis Protects against Tetanus Toxin. Infect. Immun. 1999, 67, 4847–4850. [Google Scholar] [CrossRef] [Green Version]
- Chao, G.; Lau, W.L.; Hackel, B.J.; Sazinsky, S.L.; Lippow, S.M.; Wittrup, K.D. Isolating and engineering human antibodies using yeast surface display. Nat. Protoc. 2006, 1, 755–768. [Google Scholar] [CrossRef]
- Bowers, P.M.; Horlick, R.A.; Kehry, M.R.; Neben, T.Y.; Tomlinson, G.L.; Altobell, L.; Zhang, X.; Macomber, J.L.; Krapf, I.P.; Wu, B.F.; et al. Mammalian cell display for the discovery and optimization of antibody therapeutics. Methods 2014, 65, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Dieterich, W.; Ehnis, T.; Bauer, M.; Donner, P.; Volta, U.; Riecken, E.O.; Schuppan, D. Identification of tissue transglutaminase as the autoantigen of celiac disease. Nat. Med. 1997, 3, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Sblattero, D.; Florian, F.; Not, T.; Ventura, A.; Bradbury, A.; Marzari, R. Analyzing the peripheral blood antibody repertoire of a celiac disease patient using phage antibody libraries. Hum. Antibodies 2000, 9, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Marzari, R.; Sblattero, D.; Florian, F.; Tongiorgi, E.; Not, T.; Tommasini, A.; Ventura, A.; Bradbury, A. Molecular Dissection of the Tissue Transglutaminase Autoantibody Response in Celiac Disease. J. Immunol. 2001, 166, 4170–4176. [Google Scholar] [CrossRef] [Green Version]
- Sblattero, D.; Florian, F.; Azzoni, E.; Ziberna, F.; Tommasini, A.; Not, T.; Ventura, A.; Bradbury, A.; Marzari, R. One-step cloning of anti tissue transglutaminase scFv from subjects with celiac disease. J. Autoimmun. 2004, 22, 65–72. [Google Scholar] [CrossRef]
- Not, T.; Ziberna, F.; Vatta, S.; Quaglia, S.; Martelossi, S.; Villanacci, V.; Marzari, R.; Florian, F.; Vecchiet, M.; Sulic, A.-M.; et al. Cryptic genetic gluten intolerance revealed by intestinal antitransglutaminase antibodies and response to gluten-free diet. Gut 2011, 60, 1487–1493. [Google Scholar] [CrossRef]
- Høydahl, L.S.; Richter, L.; Frick, R.; Snir, O.; Gunnarsen, K.S.; Landsverk, O.J.B.; Iversen, R.; Jeliazkov, J.R.; Gray, J.J.; Bergseng, E.; et al. Plasma Cells Are the Most Abundant Gluten Peptide MHC-expressing Cells in Inflamed Intestinal Tissues From Patients With Celiac Disease. Gastroenterology 2019, 156, 1428–1439.e10. [Google Scholar] [CrossRef] [Green Version]
- Rhyner, C.; Weichel, M.; Hubner, P.; Achatz, G.; Blaser, K.; Crameri, R. Phage display of human antibodies from a patient suffering from coeliac disease and selection of isotype-specific scFv against gliadin. Immunology 2003, 110, 269–274. [Google Scholar] [CrossRef]
- Chen, T.; Hoffmann, K.; Östman, S.; Sandberg, A.-S.; Olsson, O. Identification of gliadin-binding peptides by phage display. BMC Biotechnol. 2011, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Doña, V.; Urrutia, M.; Bayardo, M.; Alzogaray, V.; Goldbaum, F.A.; Chirdo, F.G. Single Domain Antibodies Are Specially Suited for Quantitative Determination of Gliadins under Denaturing Conditions. J. Agric. Food Chem. 2010, 58, 918–926. [Google Scholar] [CrossRef]
- García-García, A.; Madrid, R.; González, I.; García, T.; Martín, R. A novel approach to produce phage single domain antibody fragments for the detection of gluten in foods. Food Chem. 2020, 321, 126685. [Google Scholar] [CrossRef] [PubMed]
- Jayathilake, C.; Kumachi, S.; Arai, H.; Motohashi, M.; Terai, T.; Murakami, A.; Nemoto, N. In vitro selection of anti-gliadin single-domain antibodies from a naïve library for cDNA-display mediated immuno-PCR. Anal. Biochem. 2020, 589, 113490. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.Y.; Iorno, N.; Sierro, F.; Christ, D. Selection of human antibody fragments by phage display. Nat. Protoc. 2007, 2, 3001–3008. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.M.; Rallabhandi, P.; Williams, K.M.; Pahlavan, A. Characterization of Antibodies for Grain-Specific Gluten Detection. J. Food Sci. 2016, 81, T810–T816. [Google Scholar] [CrossRef] [PubMed]
- Anzai, H.; Terai, T.; Jayathilake, C.; Suzuki, T.; Nemoto, N. A novel immuno-PCR method using cDNA display. Anal. Biochem. 2019, 578, 1–6. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Osborne Fraction | Wheat | Rye | Oats | Barley | Corn | |
|---|---|---|---|---|---|---|
| Globulin | Edestin | |||||
| Albumin | Leucosin | |||||
| Gluten | Prolamin | Gliadin | Secalin | Avenin | Hordein | Zein |
| Glutelin | Glutenin | Secalinin | Aveninin | Hordeinin | Zeinin | |
| Antibody | Company | Host | Isotype | Raised against |
|---|---|---|---|---|
| PAB29118 | Abnova | Chicken | IgY | Wheat flour protein extract |
| MBS617177 | MyBioSource | Rabbit | IgG | Wheat gluten |
| MBS838918 | MyBioSource | Rabbit | IgG | Wheat gliadin |
| MBS625849 | MyBioSource | Chicken | IgY | Wheat gluten |
| LS-C66756 | LifeSpanBiosciences | Rabbit | IgG | Wheat gluten |
| LS-C129350 | LifeSpanBiosciences | Chicken | IgY | Wheat gluten |
| LS-C750830 | LifeSpanBiosciences | Chicken | IgY | Wheat gluten |
| G8138-01 | USBiological | Rabbit | IgG | Wheat gluten |
| G8138-02 | USBiological | Chicken | IgY | Wheat flour protein extract |
| AS09 571 | Agrisera | Chicken | IgY | Wheat flour protein extract |
| PA5-97536 | Invitrogen | Rabbit | IgG | Wheat gliadin native protein |
| G9144 | Sigma-Aldrich (Merck) | Rabbit | IgG | Native and heat-treated wheat gliadin |
| Patent No. | Summarized Patents | Applicant |
|---|---|---|
| WO2006004394A2 | A method for the screening of basic ingredients, semi-manufactured ingredients, and food products that are intended to be used in a gluten-free diet, based on antibodies raised against T cell stimulatory peptides. | Academisch Ziekenhuis Leiden (The Netherlands) |
| WO2006051145A1 | Competitive ELISA for the detection of gluten hydrolysate based on the R5 monoclonal antibody. | Consejo Superior de Investigaciones Científicas (Spain) |
| WO2007104825A1 | Method for extracting gluten from processed (by heat) and unprocessed foods based on the use of ionic and non-ionic detergents as prior step for ELISA tests. | Consejo Superior de Investigaciones Científicas (Spain) |
| WO2014132204A1 | Monoclonal antibody that is capable of bonding to deamidated gluten proteins (related with celiac disease pathogenesis) and has no cross-reaction with non-deamidated gluten proteins. | Institut national de la recherche agronomique (France) |
| WO2015164615A1 | Isolated monoclonal antibodies and fragments that bind to 11 peptides that can be found in gluten proteins. | University of Chicago (USA) and University of Oslo (Norway) |
| WO2018071718A1 | Antibodies, fragments, or polypeptides in the detection of gliadin: heavy chain and light chain variable sequences, and associated sequences of complementarity-determining regions (CDRs). | Nima Labs Inc. (USA) |
| WO2019154559A1 | Immunoassay methods for the quantification of the total gluten content of grains in food samples. | R-Biopharm AG. (Germany) |
| ES2392412A1 | Solutions for the extraction and solubilization of gluten, composed of arginine and ethanol. | Biomedal S.L (Spain) |
| GB2207921A | Hybridoma cell line ATCC HB9798 that produces monoclonal antibodies directed against omega gliadin protein of wheat and related proteins in rye and barley. | Commonwealth Scientific and Industrial Research Organization (United Kingdom) |
| CN101698832A | Anti-gliadin monoclonal antibody and the hybridoma cell line obtained from it. | Quingdao Biomade Technology Company Ltd. (China) |
| CN107860918A | Colloidal gold immunochromatography test strip for the gluten allergen in food and the preparation method of colloidal gold immunochromatography test strip. | Biofront Technology Company Ltd. (China) |
| Type of Library | Antibody Format | Isotype VH Family Gene/s | Selection Driven by | References |
|---|---|---|---|---|
| Immune (peripheral blood lymphocytes (PBLs) from celiac patients) | Single-chain fragment variable antibody (ScFv) | IgG VH4 | α-gliadin human transglutaminase Other antigens | [109] |
| Immune (PBLs and intestinal lymphocytes (IBLs) from celiac patients) | ScFv | VH5 | Human transglutaminase α-gliadin | [110] |
| Immune (IBLs from celiac patients) | ScFv | VH5 | Human transglutaminase | [111] |
| Immune (IBLs from celiac patient relatives) | ScFv | VH51-1 | Human transglutaminase | [112] |
| Naïve | ScFv | NA | Gliadin HLA-presenting peptides | [113] |
| Immune | ScFv | IgA, IgG, and IgM | Gliadin | [114] |
| Random peptides | Random peptides | NA | Gliadin HLA-presenting cells | [115] |
| Technology Used | Type of Library | Antibody Format | Selection Driven by | Gluten Detection Method | References |
|---|---|---|---|---|---|
| Phage display | Immune | VHH | Gliadin | Capture ELISA | [116] |
| Phage display | Naïve | single-domain antibody (dAb) | Consensus gluten peptide | Phage ELISA | [117] |
| cDNA display | Naïve | VHH | Gliadin | cDNA display mediated immuno-PCR (cD-IPCR) | [118] |
| Technology | Market Available Test for Gluten Detection Based on These Technologies | Validated Tests for Gluten Detection Based on These Antibodies | Clinical Applications Developments with These Antibodies | Inter-Batch Evenness | Need Animal Experimentation for Its Development | Technical Readiness |
|---|---|---|---|---|---|---|
| Polyclonal antibodies | Yes | Yes | Yes | Variable | Yes | Mature |
| Monoclonal antibodies | Yes | Yes | Yes | Yes | Yes | Mature |
| Recombinant antibodies | Not yet | Not yet | Yes | Yes | Not all | Recent |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Calvo, E.; García-García, A.; Madrid, R.; Martin, R.; García, T. From Polyclonal Sera to Recombinant Antibodies: A Review of Immunological Detection of Gluten in Foodstuff. Foods 2021, 10, 66. https://doi.org/10.3390/foods10010066
Garcia-Calvo E, García-García A, Madrid R, Martin R, García T. From Polyclonal Sera to Recombinant Antibodies: A Review of Immunological Detection of Gluten in Foodstuff. Foods. 2021; 10(1):66. https://doi.org/10.3390/foods10010066
Chicago/Turabian StyleGarcia-Calvo, Eduardo, Aina García-García, Raquel Madrid, Rosario Martin, and Teresa García. 2021. "From Polyclonal Sera to Recombinant Antibodies: A Review of Immunological Detection of Gluten in Foodstuff" Foods 10, no. 1: 66. https://doi.org/10.3390/foods10010066
APA StyleGarcia-Calvo, E., García-García, A., Madrid, R., Martin, R., & García, T. (2021). From Polyclonal Sera to Recombinant Antibodies: A Review of Immunological Detection of Gluten in Foodstuff. Foods, 10(1), 66. https://doi.org/10.3390/foods10010066