
Locally Adapted and Organically Grown Landrace and Ancient Spring Cereals—A Unique Source of Minerals in the Human Diet



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Spring Cereal Material Produced and Used for the Study
2.2. Mineral Analyses
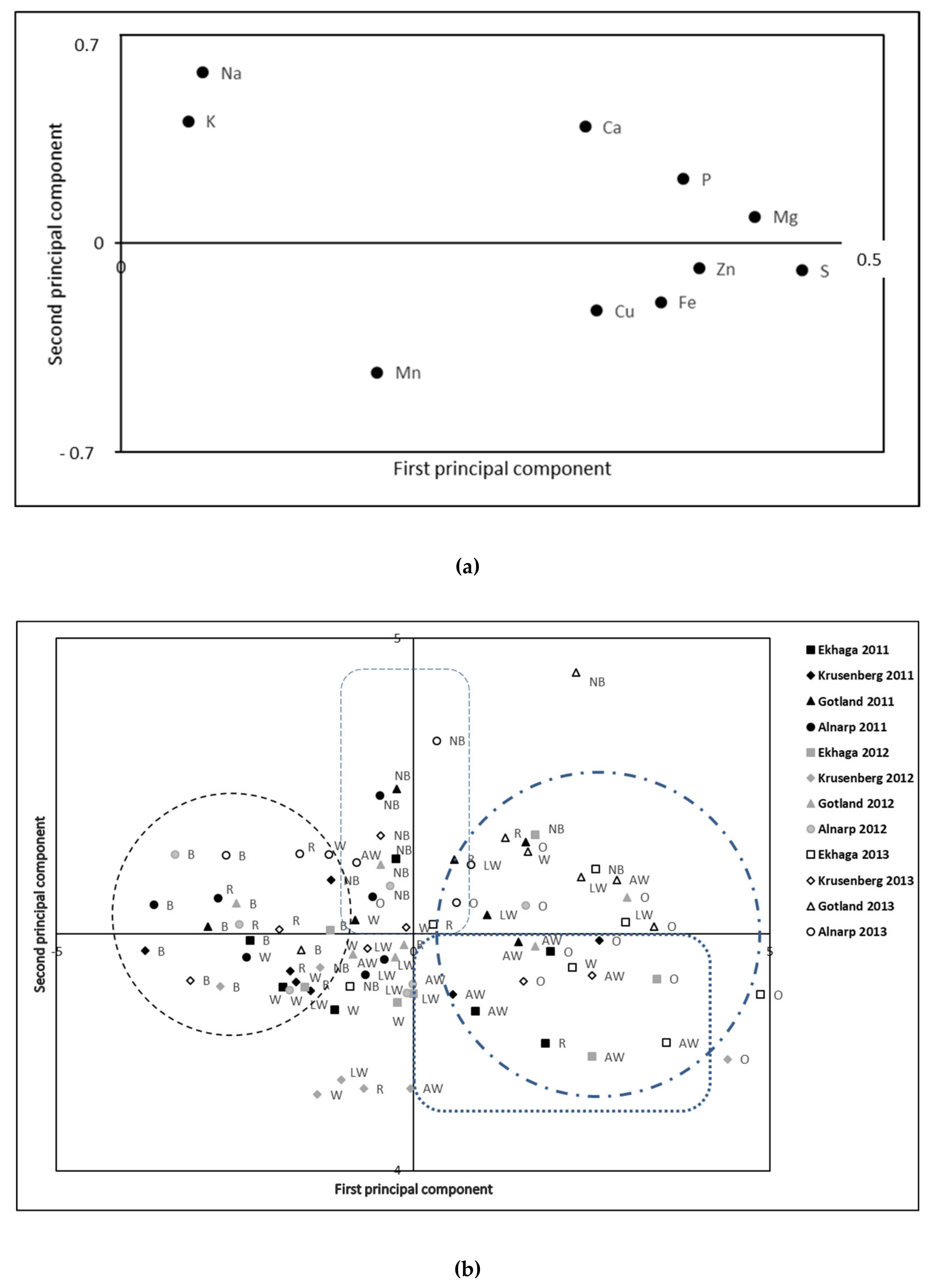
2.3. Statistical Analyses
2.4. Calculation of Nutritional Yield and Nutrient Density
3. Results
3.1. Variation of Mineral Contents in the Material
3.2. Minerals Variation by Genotype, Cultivation Location, Cultivation Year and Genotype Group
3.3. Combined Impact of Genotype/Genotype Group, Cultivation Location and Year on Minerals Content
3.4. Local Adaptation of the Genotypes
3.5. Nutritional Yield and Nutrient Density
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Sec. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- FAO Food and Agriculture Organization of the United Nations. FAOSTAT Database; FAO: Rome, Italy, 2019. [Google Scholar]
- Pena-Bautista, R.J.; Hernandez-Espinosa, N.; Jones, J.M.; Guzman, C.; Braun, H.J. Wheat-based foods: Their global and regional importance in the food supply, nutrition and health. Cereal Foods World 2017, 62, 231–249. [Google Scholar] [CrossRef]
- Andersen, V.; Bar, E.; Wirtanen, G. Nutritional and Health Aspects of Food in Nordic Countries; Elsevier, Academic Press: Amsterdam, The Netherlands, 2018; ISBN 978-0-12-809416-7. [Google Scholar]
- Peltonen-Sainio, P. Crop Production in a Northern Climate. FAO. Available online: www.fao.org/3/i3084e/i3084e15.pdf (accessed on 18 December 2020).
- Spring Wheat, Lantmännen. Available online: https://www.lantmannenseed.com/varieties/spring-wheat (accessed on 18 December 2020).
- Shafie, F.A.; Rennie, D. Consumer perceptions towards organic food. Procedia Soc. Behav. Sci. 2012, 49, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Johansson, E.; Hussain, A.; Kuktaite, R.; Andersson, S.C.; Olsson, M.E. Contribution of organically grown crops to human health. Int. J. Environ. Res. Public Health 2014, 11, 3870–3893. [Google Scholar] [CrossRef]
- Zaccone, C.; Di Caterina, R.; Rotunno, T.; Quinto, M. Soil–farming system–food–health: Effect of conventional and organic fertilizers on heavy metal (Cd, Cr, Cu, Ni, Pb, Zn) content in semolina samples. Soil Till. Res. 2010, 107, 97–105. [Google Scholar] [CrossRef]
- Niederlie, P.; Schubert, M.N. How does veganism contribute to shape sustainable food systems? Practices, meanings and identities of vegan restaurants in Porto Alegre, Brazil. J. Rural Stud. 2020, 78, 304–313. [Google Scholar] [CrossRef]
- Reisch, L.; Eberle, U.; Lorek, S. Sustainable food consumption: An overview of contemporary issues and policies. Sust. Sci Pract. Ploicy 2013, 9, 7–25. [Google Scholar] [CrossRef]
- Pelletier, J.E.; Laska, M.N.; Neumark-Sztainer, D.; Story, M. Positive attitudes towards organic, local, and sustainable foods are associated with higher dietary quality among young adults. J. Acad. Nutr. Diet. 2013, 113, 127–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Larsson, H.; Kuktaite, R.; Johansson, E. Mineral composition of organically grown wheat genotypes: Contribution to daily minerals intake. Int. J. Environ. Res. Public Health 2010, 7, 3442–3456. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Ascarrunz, S.G.; Larsson, H.; Prieto-Linde, M.L.; Johansson, E. Mineral nutritional yield and nutrient density of locally adapted wheat genotypes under organic production. Foods 2016, 5, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Larsson, H.; Kuktaite, R.; Olsson, M.E.; Johansson, E. Carotenoid content in organically produced wheat: Relevance for human nutritional health on consumption. Int. J. Env. Res. Public Health 2015, 12, 14068–14083. [Google Scholar] [CrossRef] [Green Version]
- Zamaratskaia, G.; Gerhardt, K.; Wendin, K. Biocheical characteristics and potential applications of ancient cereals—An underexploited opportunity for sustainable production and consumption. Trends Food Sci. Technol. 2021, 107, 114–123. [Google Scholar] [CrossRef]
- Hamnér, K.; Kirchmann, H. Trace element concentrations in cereal grain of long-term field trials with organic fertilizer in Sweden. Nutr. Cycl. Agroecosyst. 2015, 103, 347–358. [Google Scholar] [CrossRef]
- Martinez-Ballesta, M.C.; Dominguez-Perles, R.; Moreno, D.A.; Muries, B.; Alcaraz-Lopez, C.; Bastias, E.; Garcia-Viguera, C.; Carvajal, M. Minerals in plant food: Effect of agricultural practices and role in human health. A review. Agron. Sustain. Dev. 2009, 30, 295–309. [Google Scholar] [CrossRef]
- Welch, R.M. The impact of mineral nutrients in food crops on global human health. Plant Soil 2002, 247, 83–90. [Google Scholar] [CrossRef]
- Grusak, M.A.; DellaPenna, D. Improving the nutrient composition of plants to enhance human nutrition and health. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 1999, 50, 133–161. [Google Scholar] [CrossRef]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyachoti, S.; Adebayo, S.; Godebo, T.R. Elemental composition of staple cereal crops in the Main Ethiopian Rift Valley. J. Food Comp. Anal. 2020, 103660. [Google Scholar] [CrossRef]
- Skendi, A.; Papageorgiou, M.; Irakli, M.; Katsantonis, D. Presence of mycotoxins, heavy metals and nitrate reisdues in organic commercial cereal-based foods sold in the Greek market. J. Cons. Prot. Food Saf. 2020, 15, 109–119. [Google Scholar] [CrossRef]
- Winiarska-Mieczan, A.; Kowalczuk-Vasilev, E.; Kwiatowska, K.; Kwiecien, M.; Baranowska-Wójcik, E.; Kiczorowska, B.; Klebanuik, R.; Samoli´nska, W. Dietary intake and content of Cu, Mn, Fe, and Zn in selected cereal products marketed in Poland. Biol. Trace Elem. Res. 2019, 187, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Giannenas, I.; Nisianakis, P.; Sarakatsianos, I.; Bonos, E.; Christaki, E.; Florou-Paneri, P. Trace and major elements content of cereal and proteinaceous feeds in Greece analyzed by inductively coupled plasma mass spectrometry. In Food Quality: Balancing Health and Disease, Handbook of Food Bioengineering; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: London, UK , 2018; pp. 197–223. [Google Scholar]
- Brizio, P.; Benedetto, A.; Squadrone, S.; Curcio, A.; Pellegrino, M.; Ferrero, M.; Abete, M.C. Heavy metals and essential elements in Italian cereals. Food Add. Contamin. Part B 2016, 9, 261–267. [Google Scholar] [CrossRef]
- Teklic, T.; Loncaric, Z.; Kovacevic, V.; Singh, B.R. Metallic trace elements in cereal grain—A review: How much metal do we eat? Food Energy Sec. 2013, 2, 81–95. [Google Scholar] [CrossRef]
- Velu, G.; Singh, R.; Huerta-Espino, J.; Pena, J.; Ortiz-Monasterio, I. Breeding for enhanced Zinc and Iron concentration in CIMMYT spring wheat germplasm. Czech J. Plant Breed. 2011, 47, S174–S177. [Google Scholar] [CrossRef] [Green Version]
- Velu, G.; Sing, R.P.; Crespo-Herrera, L.; Juliana, P.; Dreisigacker, S.; Valluru, R.; Stangoulis, J.; Sohu, V.S.; Mavi, G.V.; Mishra, V.K.; et al. Genetic dissection of grain zinc concentration in spring wheat for mainstreaming biofortification in CIMMYT wheat. Sci. Rep. 2018, 8, 13526. [Google Scholar] [CrossRef]
- Velu, G.; Singh, R.P.; Huerta-Espino, J.; Pena, R.J.; Arun, B.; Mahendru-Singh, A.; Mujahid, M.J.; Sohu, V.S.; Mavi, G.S.; Crossa, J.; et al. Performance of biofortified spring wheat genotypes in target environments for grain zinc and iron concentration. Field Crops Res. 2012, 137, 261–267. [Google Scholar] [CrossRef]
- Velu, G.; Herrera, L.C.; Guzman, C.; Huerta, J.; Payne, T.; Singh, R.P. Assesing genetic diversity to breed competitive biofortified wheat with enhanced grain Zn and Fe concentrations. Front. Plant Sci. 2019, 9, 1971. [Google Scholar] [CrossRef] [PubMed]
- Johansson, E.; Branlard, G.; Cuniberti, M.; Flagella, Z.; Hüsken, A.; Nurit, E.; Peña, R.J.; Sissons, M.; Vazquez, D. Genotypic and environmental effects on wheat technological and nutritional quality. In Wheat Quality for Improving Processing and Human Health; Igrejas, G., Ikeda, T., Guzmán, C., Eds.; Springer: Cham, Switzerland, 2020; pp. 171–204. [Google Scholar]
- Rubene, D.; Kuka, P. Minerals in oats, barley and wheat grains. In Research for Rural Development; Latvia University of Agriculture: Jelgava, Latvia, 2007; 319p. [Google Scholar]
- Rodelhutscord, M.; Rückert, C.; Maurer, H.P.; Schenkel, H.; Schipprack, W.; Knudsen, K.E.B.; Schollenberger, M.; Laux, M.; Eklund, M.; Siegert, W.; et al. Variation in chemical composition and physical characteristics of cereal grains from different genotypes. Arch. Anim. Nutr. 2016, 70, 87–107. [Google Scholar] [CrossRef]
- Mithril, C.; Dragsted, L.O.; Meyer, C.; Tetens, I.; Biltoft-Jensen, A.; Astru, A. Guidelines for the New Nordic Diet. Public Health Nutr. 2012, 15, 1941–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mithril, C.; Dragsted, L.O.; Meyer, C.; Tetens, I.; Biltoft-Jensen, A.; Astru, A. Dietary composition and nutrient content of the New Nordic Diet. Public Health Nutr. 2013, 16, 777–785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, A.; Egeberg, R.; Christensen, J.; Overvad, K.; Tjonneland, A. Healthy aspects of the Nordic Diet are related to lower total mortality. J. Nutr. 2011, 141, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Adamsson, V.; Reumark, A.; Fredriksson, I.-B.; Hammarström, E.; Vessby, B.; Johansson, G.; Risérus, U. Effects of a healthy Nordic diet on cardiovascular factors in hypercholesterolaemic subjects: A randomized controlled trial (NORDIET). J. Intern. Med. 2011, 269, 150–159. [Google Scholar] [CrossRef]
- Delin, S.; Soderstrom, M. Performance of soil electrical conductivity and different methods for mapping soil data from a small dataset. Acta Agr. Scand. Sect. B Soil Plant 2003, 52, 127–135. [Google Scholar] [CrossRef]
- Malik, A.H.; Kuktaite, R.; Johansson, E. Combined effect of genetic and environmental factors on the accumulation of proteins in the wheat grain and their relationship to bread-making quality. J. Cereal Sci. 2013, 57, 170–174. [Google Scholar] [CrossRef]
- Vazquez, D.; Berger, A.; Prieto-Linde, M.L.; Johansson, E. Can nitrogen fertilization be used to modulate yield, protein content and bread-making quality in Uruguayan wheat? J. Cereal Sci. 2019, 85, 153–161. [Google Scholar] [CrossRef]
- Mukamuhirwa, A.; Hovmalm, H.P.; Ortiz, R.; Nyamangyoku, O.; Prieto-Linde, M.L.; Ekholm, A.; Johansson, E. Effect of intermittent drought on grain yield and quality of rice (Oryza sativa L.) grown in Rwanda. J. Agron. Crop Sci. 2020, 206, 252–262. [Google Scholar] [CrossRef]
- Peleg, Z.; Cakmak, I.; Ozturk, L.; Yazici, A.; Jun, Y.; Budak, H.; Korol, A.B.; Fahima, T.; Saranga, Y. Quantitative trait loci conferring grain mineral nutrient concentrations in durum wheat x wild emmer wheat RIL population. Theor. Appl. Genet. 2009, 119, 353–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, V.K.; Rawat, N.; Chhuneja, P.; Neelam, K.; Aggarwal, R.; Randhawa, G.S.; Dhaliwal, H.S.; Keller, B.; Singh, K. Mapping of quantitative trait loci for grain iron and zink concentration in diploid A genom wheat. J. Hered. 2009, 6, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.K.; Crossa, J.; Arun, B.; Chand, R.; Trethovan, R.; Vargas, M.; Ortiz-Monasterio, I. Gentype x environment interaction for zinc and iron concentration of wheat grain in eastern Gangetic plains of India. Field Crops Res. 2010, 116, 268–277. [Google Scholar] [CrossRef]
- Hristov, N.; Mladenov, N.; Djuric, V.; Konkic-Spika, A.; Marjanovic-Jeromela, A.; Simic, D. Genotype by environment interactions in wheat quality breeding programs in southeast Europe. Euphytica 2012, 174, 315–324. [Google Scholar] [CrossRef]
- Johansson, E.; Svensson, G.; Tsegaye, S. Genotype and environment effects on bread-making quality of Swedish grown wheat cultivars containing HMW glutenin subunits 2 + 12 or 5 + 10. Acta Agric. Scand. 2000, 49, 225–233. [Google Scholar]
- Koppel, R.; Ingver, A. Stability and predictability of baking quality of winter wheat. Agron. Res. 2010, 8, 637–644. [Google Scholar]
- Malik, A.H.; Prieto-Linde, M.L.; Kuktaite, R.; Andersson, A.; Johansson, E. Individual and interactive effects of cultivar maturation time, nitrogen regime and temperature level on accumulation of wheat grain proteins. J. Sci. Food Agric. 2011, 91, 2192–2200. [Google Scholar] [CrossRef] [PubMed]
- Rozbicki, J.; Ceglińska, A.; Gozdowski, D.; Jakubczak, M.; Cacak-Pietrzak, G.; Mądry, W.; Golba, J.; Piechociński, M.; Sobczyński, G.; Studnicki, M.; et al. Influence of the cultivar, environment and management on the grain yield and bread-making quality in winter wheat. J. Cereal Sci. 2015, 61, 126–132. [Google Scholar] [CrossRef]
- Johansson, E.; Nilsson, H.; Mazhar, H.; Skerritt, J.; MacRitchie, F.; Svensson, G. Seasonal effects on storage proteins and gluten strength in four Swedish wheat cultivars. J. Sci. Food Agric. 2002, 82, 1305–1311. [Google Scholar] [CrossRef]
- Andersson, S.C.; Rumpunen, K.; Johansson, E.; Olsson, M.E. Tocopherols and tocotrienols in Sea Buchthorn (Hippophae rhamnoides L.) berries during ripening. J. Agric. Food Chem. 2008, 56, 6701–6706. [Google Scholar] [CrossRef]
- Baye, T.M.; Abebe, T.; Wilke, R.A. Genotype-environment interactions and their translational implications. Per. Med. 2011, 8, 59–70. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Maximino, J.V.; Barros, L.M.; Pereira, R.M.; de Santi, I.I.; Aranha, B.C.; Busanello, C.; Viana, V.E.; Freitag, R.A.; Batista, L.; de Oliveira, A.C.; et al. Mineral and fatty acid content variation in white oat genotypes grown in Brazil. Biol. Trace Elem. Res. 2020, 199, 1194–1206. [Google Scholar] [CrossRef]
- Bityutskii, N.; Yakkonen, K.; Loskutov, I. Content of iron, zinc and manganese in grains of Triticum aestivum, Secale cereal, Hordeum vulgare and Avena sativa cultivars registered in Russia. Genet. Resour. Crop Evol. 2017, 64, 1955–1961. [Google Scholar] [CrossRef]
- Jãkobsone, I.; Kantãne, I.; Zute, S.; Jansone, I.; Bartkevičs, V. Macroelements and trace elements in cereal grains cultivated in Latvia. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2015, 69, 152–157. [Google Scholar]
- DeFries, R.; Fanzo, J.; Remans, R.; Palm, C.; Wood, S.; Anderman, T.L. Metrics for land-scarce agriculture. Science 2015, 349, 238–240. [Google Scholar] [CrossRef]
- Oury, F.-X.; Leenhardt, F.; Rémésy, C.; Chanliaud, E.; Duperrier, B.; Balfourier, F.; Charmet, G. Genetic variability and stability of grain Mg, Zn and Fe concentrations in bread wheat. Eur. J. Agron. 2006, 25, 177–185. [Google Scholar] [CrossRef]
- Lundegårdh, B.; Jastrebova, J.; Zhokhov, S.; Mårtensson, A.; Öborn, I. Effects of Growing Location and Variety on Free Tryptophan and Mineral Nutrient Content in Wheat; Latvia University of Agriculture, Faculty of Agriculture: Jelgava, Latvia, 2009; pp. 21–27. [Google Scholar]
- Meltzer, H.M.; Brantesaeter, A.L.; Trolle, E.; Eneroth, H.; Fogelholm, M.; Ydersbond, T.A.; Birgisdottir, B.E. Environmental sustainability perspectives of the Nordic diet. Nutrients 2009, 11, 2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Statistik Från Jordbruksverket. Livsmedelskonsumtionen 1960–2006 (Consumption of Food 1960–2006). Statistikrapport 2009:2. (In Swedish). Available online: https://djur.jordbruksverket.se/webdav/files/SJV/Amnesomraden/Statistik,%20fakta/Livsmedel/2009_2/20092_amk_ihopb.pdf (accessed on 30 December 2020).
- Jiang, S.L.; Wu, J.G.; Thang, N.B.; Feng, Y.; Yang, X.E.; Shi, C.H. Genotypic variation of mineral elements contents in rice (Oryza sativa L.). Eur. Food. Res. Technol. 2008, 228, 115. [Google Scholar] [CrossRef]
- Belaggia, R.; Fragasso, M.; Miglietta, F.; Cattivelli, L.; Menga, V.; Nigro, F.; Pecchioni, N.; Fares, C. Mineral composition of durum wheat grain and pasta under increasing atmospheric CO2 concentrations. Food Chem. 2018, 242, 53–61. [Google Scholar] [CrossRef]
- Vignola, M.B.; Moiraghi, M.; Salvucci, E.; Baroni, V.; Pérez, G.T. Whole meal and white flour from Argentine wheat genotypes: Mineral and arabinoxylan differences. J. Cereal Sci. 2016, 71, 217–223. [Google Scholar] [CrossRef]
- Nordic Council of Ministers. Nordic Nutrition Recommendations 2012: Integrating Nutrition and Physical Activity, 5th ed.; Nordic Council of Ministers: Copenhagen, Denmark, 2012.


{kind=link}
{kind=link}
| Genotype | Type | Origin | Place 1 |
|---|---|---|---|
| Algot | Wheat | Cultivar 1953 | E, K, A |
| Alva | Barley | Cultivar 1977 | A |
| Argus | Black oats | Cultivar 1926 | G |
| Atle | Wheat | Cultivar 1953 | G A |
| Atson | Wheat | Cultivar 1954 | A |
| Aurore | Wheat | Cultivar 1929 | K |
| Balder | Barley | Cultivar 1945 | E, K |
| Bambu | White oats | Cultivar 1934 | E |
| Blenda | White oats | Cultivar 1950 | G |
| Dacke | Wheat | Cultivar 1990 | E, K |
| Diamant brun | Wheat | Cultivar 1928 | E, K, G, A |
| Domen | Barley | Cultivar 1959 | E |
| Dragon | Wheat | Cultivar 1988 | G, A |
| Ella | Wheat | Cultivar 1950 | E, K, G, A |
| Emmer Gotland | Emmer wheat | Primitive | E, K, G, A |
| Engelbrekt | Black oats | Cultivar 1924 | E, K, G, A |
| Extra Klock | Black oats | Cultivar 1955 | K |
| Gotlandskorn | Barley | Cultivar 1915 | G, A |
| Gullkorn | Barley | Cultivar 1913 | G |
| Hulless 6row barley | Hulless barley | Genebank | E, K, G, A |
| Hulless 2row barley | Hulless barley | Genebank | E, K, G, A |
| Hulless oats | Hulless oats | Genebank | E, K, G, A |
| Ingrid | Barley | Cultivar 1958 | E, K, G, A |
| Jusso | Rye | Landrace | E, K, G, A |
| Kajsa | Barley | Cultivar 1977 | E, K |
| Klock | Black oats | Cultivar 1917 | A |
| Kärn | Wheat | Cultivar 1946 | E, K |
| Landrace Dalarna | Wheat | Landrace | E, K, G |
| Landrace Halland | Wheat | Landrace | E, K |
| Lina | Barley | Cultivar 1982 | A |
| Orion | Black oats | Cultivar 1920 | E, K, G |
| Osmo | Back oats | Cultivar 1921 | A |
| Palu | White oats | Cultivar 1945 | A |
| Prins | Wheat | Cultivar 1965 | E, G |
| Rika | Barley | Cultivar 1949 | G |
| Seger | White oats | Cultivar 1908 | K, A |
| Selma | White oats | Cultivar 1970 | A |
| Sisu | White oats | Cultivar 1953 | E |
| Sol | White oats | Cultivar 1950 | E, K |
| Spelt wheat Gotland | Spelt wheat | Spelt | E, K, G, A |
| Spelt wheat Gotland d | Spelt wheat | Spelt | G |
| Summer oats | White oats | Landrace | G |
| Svanhals | Barley | Cultivar 1903 | K |
| Ur Gotland | Black oats | Landrace | G |
| Walter | Wheat | Cultivar 1972 | A |
| Virma | White oats | Cultivar 1988 | E, K, G, A |
| Öland | Wheat | Landrace | E, K, G, A |
| Location | pH a | Organic Matter (%) | Clay (%) | P-Al b (mg 100 g−1) | K-Al b (mg 100 g−1) | FYM c | Organic Since |
|---|---|---|---|---|---|---|---|
| Ekhaga | 5.7–6.1 | 7.2–9.8 | 35–38 | 6.2–7.1 | 25–27.8 | No | 1987 |
| Krusenberg d | 5.8 | 2.0 | 9.0 | 8.0 | 5.0 | No | 2001 |
| Gotland | 7.5–8.3 | 2.5–3.9 | 18–20 | 5.2–9.8 | 9.1–10.4 | Applied | 1987 |
| Alnarp | 7.3–7.8 | 3.1–4.5 | 18–22 | 7.7–26.7 | 10.2–18.7 | No | 1992 |
| Source | Df | Zn (103) | S (105) | P (106) | Na (103) | Mn (103) | Mg (105) | K (106) | Fe (102) | Cu | Ca (105) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 12 genotypes over four locations and three years | |||||||||||
| Genotype | 11 | 0.32 *** | 4.57 *** | 2.57 *** | 2.32 *** | 1.07 *** | 1.80 *** | 1.12 *** | 2.13 *** | 2.34 *** | 1.47 *** |
| Locality | 3 | 1.18 *** | 4.82 *** | 1.19 *** | 1.09 *** | 4.16 *** | 1.20 *** | 0.06 | 6.72 *** | 22.7 *** | 0.28 *** |
| Year | 2 | 0.08 *** | 2.32 * | 0.10 | 1.92 *** | 0.34 *** | 0.70 *** | 4.26 *** | 1.57 *** | 21.7 *** | 2.47 *** |
| Ge*L | 33 | 0.03 | 0.20 | 0.13 | 0.21 | 0.10 *** | 0.15 * | 0.08 | 0.54 | 0.36 | 0.03 |
| Ge*Y | 21 | 0.03 | 0.23 | 0.21 ** | 0.36 *** | 0.07 * | 0.13 | 0.10 * | 0.60 | 0.40 | 0.12 *** |
| L*Y | 6 | 0.14 *** | 2.03 *** | 0.66 *** | 0.82 *** | 0.17 *** | 0.31 *** | 0.31 *** | 3.12 *** | 4.01 *** | 0.07 *** |
| Error | 182 | 0.02 | 0.17 | 0.09 | 0.14 | 0.03 | 0.08 | 0.05 | 0.56 | 0.24 | 0.01 |
| 7 genotype groups over four locations and three years | |||||||||||
| Group | 6 | 0.79 *** | 31.4 *** | 15.1 *** | 5.33 *** | 4.84 *** | 8.82 *** | 1.68 *** | 10.3 *** | 6.98 *** | 6.26 *** |
| Locality | 3 | 1.93 *** | 11.1 *** | 3.20 *** | 2.87 *** | 9.84 *** | 2.86 *** | 0.10 | 9.28 *** | 50.3 *** | 0.85 *** |
| Year | 2 | 2.33 *** | 4.65 *** | 0.73 *** | 3.87 *** | 1.25 *** | 1.17 *** | 6.44 *** | 10.2 *** | 54.7 *** | 3.81 *** |
| Gr*L | 18 | 0.09 *** | 0.28 | 0.15 | 0.34 *** | 0.35 *** | 0.41 *** | 0.34 *** | 1.23 *** | 0.93 *** | 0.10 *** |
| Gr*Y | 12 | 0.06 *** | 0.54 *** | 0.48 *** | 1.14 *** | 0.15 *** | 0.33 *** | 0.24 *** | 1.61 *** | 1.29 *** | 0.59 *** |
| L*Y | 6 | 0.21 *** | 5.24 *** | 1.28 *** | 1.06 *** | 0.44 *** | 0.49 *** | 0.52 *** | 5.94 *** | 8.66 *** | 0.26 *** |
| Error | 84 | 0.03 | 0.18 | 0.13 | 0.12 | 0.03 | 0.09 | 0.05 | 0.51 | 0.44 | 0.03 |
| Source | Zn | S | P | Na | Mn | Mg | K | Fe | Cu | Ca |
|---|---|---|---|---|---|---|---|---|---|---|
| Genotype | 25.1 | 42.3 | 50.6 | 28.2 | 29.8 | 43.8 | 36.8 | 14.0 | 17.0 | 55.9 |
| Locality | 29.3 | 15.0 | 8.58 | 6.20 | 39.1 | 8.32 | 0.94 | 10.2 | 25.9 | 3.86 |
| Year | 9.32 | 1.90 | 0.47 | 7.33 | 2.52 | 1.20 | 19.2 | 2.18 | 22.0 | 15.1 |
| Group | 5.16 | 53.0 | 59.6 | 19.2 | 35.6 | 51.1 | 18.7 | 6.24 | 6.53 | 54.5 |
| Locality | 23.2 | 9.45 | 6.19 | 5.74 | 37.5 | 7.19 | 0.25 | 8.70 | 29.0 | 3.54 |
| Year | 16.1 | 3.14 | 0.46 | 7.11 | 2.44 | 1.64 | 22.9 | 5.79 | 19.0 | 11.1 |
| Source | Zn (101) | S (103) | P (103) | Na (101) | Mn (101) | Mg (103) | K (103) | Fe (101) | Cu | Ca (102) |
|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | ||||||||||
| Diamant brun | 4.60 a,b | 1.57 b,c,d,e | 4.36 c,d | 1.91 b | 3.41 a,b | 1.30 c,d | 3.60 c | 4.65 a | 5.32 a,b,c,d | 5.57 d |
| Ella | 4.01 b,c | 1.50 d,e | 4.27 d | 2.11 b | 3.01 b,c | 1.30 c,d | 3.69 c | 4.08 a,b | 5.45 a,b,c,d | 4.95 e,f |
| Emmer Gotland | 4.82 a | 1.61 b,c,d | 5.00 a,b | 2.28 b | 3.09 b,c | 1.46 a,b | 4.15 a,b | 4.38 a | 6.13 a,b | 4.01 g |
| Engelbrekt | 3.93 b,c | 1.95 a | 5.41 a | 2.82 b | 3.78 a,b | 1.47 a | 3.65 c | 4.89 a | 5.23 b,c,d | 6.16 c |
| Hulless 6row barley | 3.79 c | 1.53 c,d,e | 4.57 b,c,d | 5.69 a | 1.55 e,f | 1.30 b,c | 4.22 a,b | 4.32 a,b | 4.86 c,d | 5.48 d |
| Hulless 2row barley | 3.79 c | 1.60 b,c,d,e | 5.03 a,b | 5.96 a | 1.64 d,e | 1.36 a,b,c | 4.34 a | 4.61 a | 5.58 a,b,c | 5.59 d |
| Hulless oats | 3.47 c,d | 1.84 a,b | 4.96 a,b,c | 2.16 b | 3.90 a,b | 1.40 a,b,c | 3.66 c | 4.30 a,b | 4.49 c,d | 7.93 a |
| Ingrid | 2.70 d | 1.34 e | 3.70 e | 3.26 b | 0.80 f | 1.08 e | 3.52 c | 3.09 b | 4.46 d | 3.22 g |
| Jusso | 4.17 a,b,c | 1.50 d,e | 4.22 d,e | 2.00 b | 2.40 c,d | 1.15 d,e | 4.43 a | 4.33 a,b | 5.38 a,b,c,d | 4.69 f |
| Spelt wheat Gotland | 4.41 a,b,c | 1.80 a,b,c | 4.90 a,b | 2.18 b | 2.86 a,b | 1.47 a | 3.72 b,c | 4.27 a | 6.76 a | 3.34 g |
| Virma | 3.90 b,c | 2.00 a | 5.12 a,b | 1.88 b | 4.08 a | 1.41 a,b,c | 3.89 b,c | 4.55 a | 4.97 c,d | 6.62 b |
| Öland | 4.15 a,b,c | 1.60 b,c,d,e | 4.68 b,c,d | 1.54 b | 3.16 b,c | 1.40 a,b,c | 3.70 c | 4.92 a | 5.27 a,b,c,d | 5.19 d,e |
| Localities | ||||||||||
| Ekhaga | 4.88 a | 1.76 a | 4.68 b | 3.14 a | 3.41 b | 1.37 a | 3.87 a | 5.01 a | 6.41 a | 5.18 b |
| Krusenberg | 3.91 b | 1.60 b,c | 4.42 c | 1.82 b | 4.13 a | 1.29 b | 3.84 a | 4.27 b | 4.89 b,c | 4.71 c |
| Gotland | 4.14 b | 1.72 a,b | 4.97 a | 3.30 a | 1.43 d | 1.42 a | 3.96 a | 4.20 b | 5.30 b | 5.59 a |
| Alnarp | 3.23 c | 1.49 c | 4.64 b,c | 3.05 a | 2.25 c | 1.28 b | 3.94 a | 4.06 b | 4.56 c | 5.24 b |
| Years | ||||||||||
| 2011 | 3.79 b | 1.60 a | 4.68 a | 2.26 b | 3.18 a | 1.31 a | 4.11 a | 4.20 a | 4.53 b | 5.17 b |
| 2012 | 3.76 b | 1.64 a | 4.71 a | 2.63 b | 2.61 b | 1.35 a | 3.55 b | 4.51 a | 5.83 a | 4.50 c |
| 2013 | 4.56 a | 1.70 a | 4.63 a | 3.80 a | 2.62 b | 1.36 a | 4.08 a | 4.49 a | 5.63 a | 6.05 a |
| Groups | ||||||||||
| Rye | 4.17 a,b | 1.50 b,c,d | 4.22 d,e | 2.00 b,c | 2.40 b,c | 1.15 c | 4.43 a | 4.33 a,b | 5.38 a,b | 4.69 b,c |
| Oats | 3.98 b | 2.03 a | 5.40 a | 2.16 c | 3.84 a | 1.49 a | 3.85 c,d | 4.88 a | 5.05 b | 6.69 a |
| Wheat | 3.84 b | 1.50 c | 4.20 d,e | 2.20 c | 2.89 b | 1.28 b | 3.80 c,d | 4.02 b,c | 5.10 b | 4.94 b |
| Ancient wheat | 4.68 a | 1.72 b | 5.04 b | 2.20 c | 3.04 a,b | 1.49 a | 3.98 b,c | 4.34 a,b | 6.36 a | 3.74 c,d |
| Naked barley | 3.79 b | 1.57 b,c | 4.79 b,c | 5.82 a | 1.59 c,d | 1.33 b | 4.28 a,b | 4.46 a,b | 5.21 b | 5.54 b |
| Barley | 3.09 c | 1.34 d | 3.98 e | 3.50 b | 1.01 d | 1.12 c | 3.67 d | 3.54 c | 4.82 b | 3.54 d |
| Landrace wheat | 4.07 a,b | 1.57 b,c | 4.45 c,d | 1.98 c | 3.40 a,b | 1.32 b | 3.89 c,d | 4.38 a,b | 5.35 b | 4.97 b |
| Location and Genotype | Genotype Group | All Ten Minerals | Four Selected Minerals | ||||
|---|---|---|---|---|---|---|---|
| 2011 | 2012 | 2013 | 2011 | 2012 | 2013 | ||
| Ekhaga | |||||||
| Bambu | Oat | 1.67 | 3.71 | 3.30 | |||
| Emmer Gotland | Ancient | 0.05 | 2.60 | 1.93 | |||
| Engelbrekt | Oat | 0.10 | 2.80 | 1.76 | |||
| Orion | Oat | 2.83 | 4.28 | 3.01 | 0.85 | 2.43 | 2.18 |
| Sisu | Oat | 0.14 | 2.14 | 2.00 | |||
| Sol | Oat | 0.31 | 3.64 | 0.79 | |||
| Spelt Gotland | Ancient | 0.06 | 1.11 | X | 0.03 | 2.38 | X |
| Virma | Oat | 1.07 | 3.34 | 2.77 | |||
| Krusenberg | |||||||
| Emmer Gotland | Ancient | 1.56 | 1.43 | 0.66 | |||
| Engelbrekt | Oat | 2.83 | 1.14 | 2.22 | 1.71 | 0.61 | 2.29 |
| Extra klock | Oat | 2.16 | 2.15 | 3.32 | 0.17 | 0.49 | 3.09 |
| Orion | Oat | 3.32 | 2.05 | 5.13 | 1.57 | 1.12 | 4.15 |
| Seger | Oat | 2.91 | 3.54 | 5.22 | 1.22 | 1.86 | 393 |
| Sol | Oat | 2.51 | 1.93 | 4.21 | 0.70 | 0.80 | 3.04 |
| Spelt Gotland | Ancient | 0.30 | 2.36 | 1.34 | |||
| Virma | Oat | 2.41 | 0.36 | 2.34 | |||
| Gotland | |||||||
| Argus | Oat | 0.51 | 3.36 | X | |||
| Blenda | Oat | 0.85 | 1.26 | 1.72 | |||
| Emmer Gotland | Ancient | 1.29 | 1.07 | 1.28 | 1.62 | 0.62 | 2.45 |
| Engelbrekt | Oat | 0.61 | 2.28 | 0.83 | |||
| Hulless oats | Oat | 0.53 | 1.44 | X | |||
| Orion | Oat | 1.52 | 4.20 | 1.68 | |||
| Spelt wheat Gotland | Ancient | 1.02 | 0.73 | X | 1.61 | 1.29 | X |
| Spelt wheat Gotland d | Ancient | 0.08 | 1.21 | 3.52 | |||
| Sommarhavre | Oat | 2.45 | 3.08 | 2.39 | 0.02 | 0.86 | 2.91 |
| Ur Gotland | Oat | 2.98 | 4.17 | 2.50 | 0.31 | 2.07 | 2.28 |
| Virma | Oat | 1.13 | 0.86 | 1.69 | |||
| Alnarp | |||||||
| Diamant brun | Wheat | 0.72 | 1.72 | 0.88 | 0.81 | 1.99 | 1.12 |
| Engelbrekt | Oat | 0.67 | 3.75 | X | |||
| Emmer Gotland | Ancient | 0.96 | 1.93 | 0.62 | |||
| Hulless 2row barley | Naked barley | 0.20 | 0.92 | 0.65 | 0.84 | 1.08 | 1.24 |
| Klock | Oat | 1.51 | 3.81 | 2.09 | |||
| Palu | Oat | 1.07 | 3.32 | 1.87 | |||
| Seger | Oat | 1.33 | 4.64 | X | |||
| Spelt wheat Gotland | Ancient | 0.70 | 1.52 | X | 1.35 | 2.41 | X |
| Osmo | Oat | 2.21 | 5.57 | 2.62 | 0.11 | 3.32 | 1.20 |
| Virma | Oat | 0.67 | 1.95 | 1.51 | |||
| Öland | Landrace wheat | 0.61 | 1.52 | 1.93 | 0.81 | 1.71 | 2.18 |
| Source | Yield | Protein | Zn NY | Fe NY | Zn ND | Fe ND |
|---|---|---|---|---|---|---|
| Rye | 2330 b | 11.4 b,c,d | 22.3 a,b | 32.0 a,b | 197 b,c | 283 b,c |
| Oats | 2790 b | 11.6 c,d | 22.8 b | 27.8 b | 205 b | 254 c |
| Wheat | 3060 a,b | 12.2 a,b,c | 27.9 a | 49.8 a | 214 b | 303 b |
| Ancient wheat | 2830 b | 13.3 a | 20.9 b | 33.8 a,b | 175 c | 279 b,c |
| Naked barley | 2580 b | 13.1 a,b | 26.0 a,b | 33.3 a,b | 218 b | 277 b,c |
| Barley | 3550 a | 11.0 d | 21.5 b | 27.3 b | 270 a | 344 a |
| Landrace wheat | 3040 a,b | 12.3 a,b,c,d | 30.4 a | 41.9 a | 201 b,c | 277 b,c |
| Product | Minerals (mg) | |||
|---|---|---|---|---|
| Zn | Fe | Mg | Cu | |
| Rice [62] | 3–11 | 0.2–7 | 22–62 | 0.8–6 |
| Pasta [63] | 3–6 | 9–25 | 107–145 | 0.4–0.8 |
| White flour [64] | 1–3 | 2–3 | 44–65 | 0.4–0.7 |
| Conventional whole grain wheat [58,59] | 4.3–7.3 | 8–10 | 224–320 | 0.9–1.2 |
| Whole grain wheat (present study) | 9.6 | 10 | 320 | 1.3 |
| NND mix [present study] | 10 | 11 | 329 | 1.3 |
| DRI [65] | 8 | 12 | 315 | 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johansson, E.; Prieto-Linde, M.L.; Larsson, H. Locally Adapted and Organically Grown Landrace and Ancient Spring Cereals—A Unique Source of Minerals in the Human Diet. Foods 2021, 10, 393. https://doi.org/10.3390/foods10020393
Johansson E, Prieto-Linde ML, Larsson H. Locally Adapted and Organically Grown Landrace and Ancient Spring Cereals—A Unique Source of Minerals in the Human Diet. Foods. 2021; 10(2):393. https://doi.org/10.3390/foods10020393
Chicago/Turabian StyleJohansson, Eva, Maria Luisa Prieto-Linde, and Hans Larsson. 2021. "Locally Adapted and Organically Grown Landrace and Ancient Spring Cereals—A Unique Source of Minerals in the Human Diet" Foods 10, no. 2: 393. https://doi.org/10.3390/foods10020393
APA StyleJohansson, E., Prieto-Linde, M. L., & Larsson, H. (2021). Locally Adapted and Organically Grown Landrace and Ancient Spring Cereals—A Unique Source of Minerals in the Human Diet. Foods, 10(2), 393. https://doi.org/10.3390/foods10020393