
Current Trends of Enterococci in Dairy Products: A Comprehensive Review of Their Multiple Roles

 , ,
, ,  and
and
Abstract
:1. Introduction
2. The Genus Enterococcus—A Bacterial Group That Has Evolved for Resistance
3. Raw Milk as a Source of Enterococci for Dairy Products
4. The Enterococcal Microbiota of Artisanal Cheeses
5. The Technological Roles and Potentialities of Enterococci in Artisanal Cheeses
6. Enterococci and Potential Health Benefits of Dairy Products
7. Enterococci as Opportunistic Pathogens
7.1. General Aspects
7.2. Virulence Factors of the Enterococci
7.3. Virulence Determinants in Enterococci from Foods
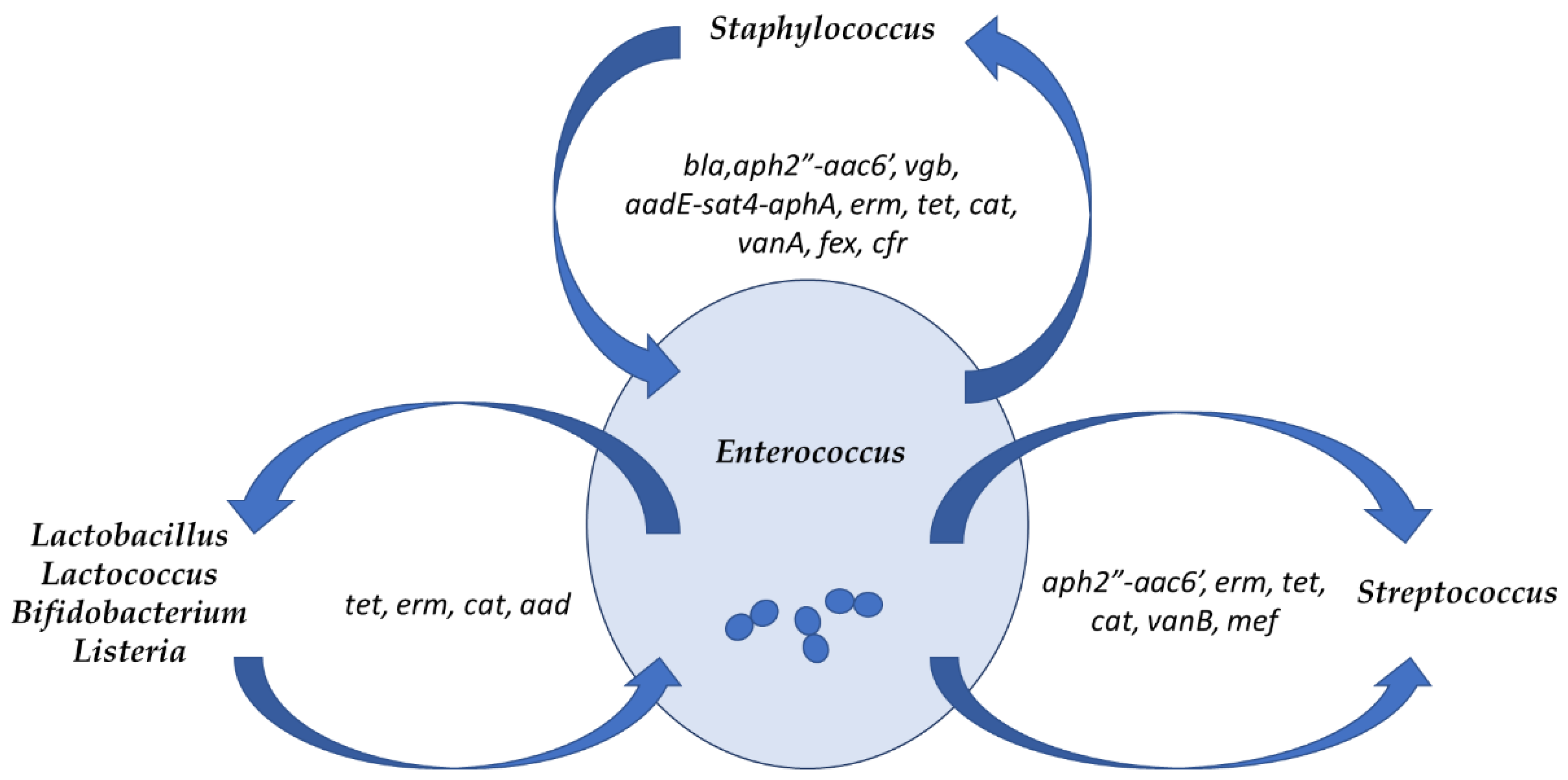
8. Enterococci as Reservoirs of Antibiotic Resistance
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Casey, M.G.; Häni, J.-P.; Gruskovnjak, J.; Schaeren, W.; Wechsler, D. Characterisation of the non-starter lactic acid bacteria (NSLAB) of Gruyère PDO cheese. Lait 2006, 86, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Fox, P.F.; Cogan, T.M.; Guinee, T.P. Chapter 52. Factors That Affect the Quality of Cheese. In Cheese: Chemistry, Physics and Microbiology, 4th ed.; McSweeney, P.L.H., Fox, P.F., Cotter, P.D., Everett, D.W., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 617–641. [Google Scholar]
- Franz, C.M.A.P.; Van Belkum, M.J.; Holzapfel, W.H.; Abriouel, H.; Gálvez, A. Diversity of enterococcal bacteriocins and their grouping in a new classification scheme. FEMS Microbiol. Rev. 2007, 31, 293–310. [Google Scholar] [CrossRef]
- Lebreton, F.; Willems, R.J.L.; Gilmore, M. Enterococcus diversity, origins in nature, and gut colonization. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014; pp. 5–63. [Google Scholar]
- Werner, G.; Coque, T.M.; Franz, C.M.; Grohmann, E.; Hegstad, K.; Jensen, L.; van Schaik, W.; Weaver, K. Antibiotic resistant enterococci–tales of a drug resistance gene trafficker. Int. J. Med. Microbiol. 2013, 303, 360–379. [Google Scholar] [CrossRef]
- Arias, C.; Murray, B. The rise of the Enterococcus: Beyond vancomycin resistance. Nat. Rev. Microbiol. 2012, 10, 266–278. [Google Scholar] [CrossRef] [Green Version]
- Graham, K.; Stack, H.; Rea, R. Safety, beneficial and technological properties of enterococci for use in functional food applications–a review. Crit. Rev. Food Sci. Nutr. 2020, 60, 3836–3861. [Google Scholar] [CrossRef] [PubMed]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The genus Enterococcus: Between probiotic potential and safety concerns–an update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef] [PubMed]
- Billroth, A.W. Untersuchungen über die Vegetationsformen von Coccobacteria Septica; Georg Reimer: Berlin, Germany, 1874. [Google Scholar]
- Rosenbach, F.J. Mikro-Organismen bei den Wund-Infections-Krankheiten des Menschen; Bergmann, J.F., Ed.; Verlag from J.F. Bergmann: Wiesbaden, Germany, 1884. [Google Scholar]
- Thiercelin, E.; Jouhaud, L. Reproduction de l’entérocoque; taches centrales; granulations peripheriques et microblastes. Comptes Rendus Seances Soc. Biol. Paris 1903, 55, 686–688. [Google Scholar]
- Andrews, F.W.; Horder, T.J. A study of streptococci pathogenic for man. Lancet 1906, 2, 708–713. [Google Scholar] [CrossRef] [Green Version]
- Lancefield, R.C. A serological differentiation of human and other groups of hemolytic streptococci. J. Exp. Med. 1933, 57, 571–595. [Google Scholar] [CrossRef]
- Sherman, J.M. The enterococci and related streptococci. J. Bacteriol 1937, 35, 81–93. [Google Scholar] [CrossRef] [Green Version]
- Schleifer, K.H.; Kilpper-Bälz, R. Transfer of Streptococcus faecalis and Streptococcus faecium to the genus Enterococcus nom. rev. as Enterococcus faecalis comb. nov. and Enterococcus faecium comb. nov. Int. J. Syst. Bacteriol. 1984, 34, 31–34. [Google Scholar]
- DSMZ. LPSN-List of Prokaryotic Names with Standing in Nomenclature. 2020. Available online: https://lpsn.dsmz.de/ (accessed on 10 February 2021).
- Lebreton, F.; Manson, A.L.; Saavedra, J.T.; Straub, T.J.; Earl, A.M.; Gilmore, M.S. Tracing the enterococci from Paleozoic origins to the hospital. Cell 2017, 169, 849–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mundt, J.O. Occurrence of enterococci in animals in a wild environment. J. Appl. Microbiol. 1963, 11, 136–140. [Google Scholar] [CrossRef]
- Martin, J.D.; Mundt, J.O. Enterococci in insects. J. Appl. Microbiol. 1972, 24, 575–580. [Google Scholar] [CrossRef]
- Silva, V.; Peixoto, F.; Igrejas, G.; Parelho, C.; Garcia, P.; Carvalho, I.; Sousa, M.; Pereira, J.E.; Rodrigues, A.; Poeta, P. First report on vanA-Enterococcus faecalis recovered from soils subjected to long-term livestock agricultural practices in Azores Archipelago. Int. J. Environ. Res. 2018, 12, 39–44. [Google Scholar] [CrossRef]
- Ben Said, L.B.; Klibi, N.; Dziri, R.; Borgo, F.; Boubadous, A.; Ben Slama, K.; Torres, C. Prevalence, antimicrobial resistance and genetic lineages of Enterococcus spp. from vegetable food, soil and irrigation water in farm environments in Tunisia. J. Sci. Food Agric. 2016, 96, 1627–1633. [Google Scholar] [CrossRef]
- Moore, D.F.; Guzman, J.A.; McGee, C. Species distribution and antimicrobial resistance of enterococci isolated from surface and ocean water. J. Appl. Microbiol. 2008, 105, 1017–1025. [Google Scholar] [CrossRef]
- Müller, T.; Ulrich, A.; Ott, E.-M.; Müller, M. Identification of plant-associated enterococci. J. Appl. Microbiol. 2001, 91, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Howden, B.P.; Stinear, T.P. Evolution of virulence in Enterococcus faecium, a hospital-adapted opportunistic pathogen. Curr. Opin. Microbiol. 2018, 41, 76–82. [Google Scholar] [CrossRef]
- Ben Braiek, O.; Smaoui, S. Enterococci: Between Emerging Pathogens and Potential Probiotics. Biomed. Res. Int. 2019, 2019, 5938210. [Google Scholar] [CrossRef]
- Novais, C.; Campos, J.; Freitas, A.R.; Barros, M.; Silveira, E.; Coque, T.M.; Antunes, P.; Peixe, L. Water supply and feed as sources of antimicrobial-resistant Enterococcus spp. in aquacultures of rainbow trout (Oncorhyncus mykiss), Portugal. Sci. Total Environ. 2018, 625, 1102–1112. [Google Scholar] [CrossRef] [PubMed]
- Channaiah, L.H.; Subramanyam, B.; Zurek, L. Molecular characterization of antibiotic resistant and potentially virulent enterococci isolated from swine farms and feed mills. J. Stored Prod. Res. 2018, 77, 189–196. [Google Scholar] [CrossRef]
- Lleò, M.M.; Bonato, B.; Benedetti, D.; Canepari, P. Survival of enterococcal species in aquatic environments. FEMS Microbiol. Ecol. 2005, 54, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Kwok, L.-Y.; Hou, Q.; Sun, Y.; Li, W.; Zhang, L.; Sun, Z. Comparative genomic analysis revealed great plasticity and environmental adaptation of the genomes of Enterococcus faecium. BMC Genom. 2019, 20, 60. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, O.; Stanton, C.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. The complex microbiota of raw milk. FEMS Microbiol. Rev. 2013, 37, 664–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perin, L.M.; Miranda, R.O.; Todorov, S.D.; Franco, B.D.G.; Nero, L.A. Virulence, antibiotic resistance and biogenic amines of bacteriocinogenic lactococci and enterococci isolated from goat milk. Int. J. Food Microbiol. 2014, 185, 121–126. [Google Scholar] [CrossRef]
- Giraffa, G. Enterococci from foods. FEMS Microbiol. Rev. 2003, 26, 163–171. [Google Scholar] [CrossRef]
- Garg, S.K.; Mital, B.K. Enterococci in milk and milk products. Crit. Rev. Microbiol. 1991, 18, 15–45. [Google Scholar] [CrossRef]
- Gelsomino, R.; Vancanneyt, M.; Cogan, T.M.; Condon, S.; Swings, J. Source of enterococci in a farmhouse raw-milk cheese. Appl. Environ. Microbiol. 2002, 68, 3560–3565. [Google Scholar] [CrossRef] [Green Version]
- Kakgli, D.M.; Vancanneyt, M.; Vandamme, P.; Hill, C.; Cogan, T.M. Contamination of milk by enterococci and coliforms from bovine faeces. J. Appl. Microbiol. 2007, 103, 1393–1405. [Google Scholar] [CrossRef]
- Kakgli, D.M.; Vancanneyt, M.; Hill, C.; Vandamme, P.; Cogan, T.M. Enterococcus and Lactobacillus contamination of raw milk in a farm dairy environment. Int. J. Food Microbiol. 2007, 114, 243–251. [Google Scholar] [CrossRef]
- Ortigosa, M.; Irigoyen, A.; Urdin, M.; García, S.; Ibañez, F.C.; Torre, P. Sources of enterococci in Idiazábal-type cheese. Int. J. Food Microbiol. 2008, 125, 146–152. [Google Scholar] [CrossRef]
- Wu, X.; Hou, S.; Zhang, Q.; Ma, Y.; Zhang, Y.; Kan, W.; Zhao, X. Prevalence of virulence and resistance to antibiotics in pathogenic enterococci isolated from mastitic cows. J. Vet. Med. Sci. 2016, 78, 1663–1668. [Google Scholar] [CrossRef] [Green Version]
- McAuley, C.M.; Britz, M.L.; Gobius, K.S.; Craven, H. Prevalence, seasonality, and growth of enterococci in raw and pasteurized milk in Victoria, Australia. J. Dairy Sci. 2015, 98, 8348–8358. [Google Scholar] [CrossRef] [Green Version]
- Giraffa, G.; Carminati, D.; Neviani, E. Enterococci isolated from dairy products: A review of risks and potential technological use. J. Food Prot. 1997, 60, 732–738. [Google Scholar] [CrossRef]
- McAuley, C.; Gobius, K.S.; Britz, M.L.; Craven, H.M. Heat resistance of thermoduric enterococci isolated from milk. Int. J. Food Microbiol. 2012, 154, 162–168. [Google Scholar] [CrossRef]
- Martínez, S.; López, M.; Bernardo, A. Thermal inactivation of Enterococcus faecium: Effect of growth temperature and physiological state of microbial cells. Lett. Appl. Microbiol. 2003, 37, 475–481. [Google Scholar] [CrossRef]
- Didienne, R.; Defargues, C.; Callon, C.; Meylheuc, T.; Hulin, S.; Montel, M.-C. Characteristics of microbial biofilm on wooden vats (‘gerles’) in PDO Salers cheese. Int. J. Food Microbiol. 2012, 156, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Jamet, E.; Akary, E.; Poisson, M.-A.; Chamba, J.-F.; Bertrand, X.; Serror, P. Prevalence and characterization of antibiotic resistant Enterococcus faecalis in French cheeses. Food Microbiol. 2012, 31, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Bouymajane, A.; Filali, F.R.; Oulghazi, S.; Ed-dra, A.; Benhallam, F.; El Allaoui, A.; Anissi, K.; Ouhmidou, B.; Moumni, M. Occurrence, molecular and antimicrobial resistance of Enterococcus spp. isolated from raw cow’s milk trade by street trading in Meknes city, Morocco. Germs 2018, 9, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Garroni, E.; Doulgeraki, A.I.; Pavli, F.; Spiteri, D.; Valdramidis, V.P. Characterization of indigenous lactic acid bacteria in cow milk of the Maltese Islands: A geographical and seasonal assessment. Microorganisms 2020, 8, 812. [Google Scholar] [CrossRef]
- Montel, M.-C.; Buchin, S.; Mallet, A.; Delbes-Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef] [PubMed]
- Giannino, M.L.; Marzotto, M.; Dellaglio, F.; Feligni, M. Study of microbial diversity in raw milk and fresh curd used for Fontina cheese production by culture-independent methods. Int. J. Food Microbiol. 2009, 130, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Masoud, W.; Vogensen, F.K.; Lillevang, S.; Al-Soud, W.A.; Sørensen, S.J.; Jakobsen, M. The fate of indigenous microbiota, starter cultures, Escherichia coli, Listeria innocua and Staphylococcus aureus in Danish raw milk and cheeses determined by pyrosequencing and quantitative real time (qRT)-PCR. Int. J. Food Microbiol. 2012, 153, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Von Neubeck, M.; Baur, C.; Krewinkel, M.; Stoeckel, M.; Kranz, B.; Stressler, T.; Fischer, L.; Hinrichs, J.; Scherer, S.; Wenning, M. Biodiversity of refrigerated raw milk microbiota and their enzymatic spoilage potential. Int. J. Food Microbiol. 2015, 211, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.I.; Martins, M.P.; Semedo, T.; Marques, J.J.F.; Tenreiro, R.; Crespo, M.T.B. Comparison of phenotypic and genotypic taxonomic methods for the identification of dairy enterococci. Antonie van Leeuwenhoek 2003, 85, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Citak, S.; Yucel, N.; Mendi, A. Antibiotic resistance of enterococcal isolates in raw milk. J. Food Process. Preserv. 2015, 29, 183–195. [Google Scholar] [CrossRef]
- Fricker, M.; Skånseng, B.; Rudi, K.; Stessl, B.; Ehling-Schulz, M. Shift from farm to dairy tank milk microbiota revealed by a polyphasic approach is independent from geographical origin. Int. J. Food Microbiol. 2011, 145, S24–S30. [Google Scholar] [CrossRef]
- Jiménez, E.; Ladero, V.; Chico, I.; Maldonado-Barragán, A.; López, M.; Martín, V.; Fernández, L.; Fernández, M.; Álvarez, M.A.; Torres, C.; et al. Antibiotic resistance, virulence determinants and production of biogenic amines among enterococci from ovine, feline, canine, porcine and human milk. BMC Microbiol. 2013, 13, 288. [Google Scholar] [CrossRef] [Green Version]
- Garnica, M.L.; Sáez-Nieto, J.A.; González, R.; Santos, J.A.; Gonzalo, C. Diversity of gram-positive catalase-negative cocci in sheep bulk tank milk by comparative 16S rDNA sequence analysis. Int. Dairy J. 2014, 34, 142–145. [Google Scholar] [CrossRef]
- Ariznabarreta, A.; Gonzalo, C.; San Primitivo, F. Microbiological quality and somatic cell count of ewe milk with special reference to staphylococci. J. Dairy Sci. 2002, 85, 1370–1375. [Google Scholar] [CrossRef]
- Ruiz, P.; Pérez-Martín, F.; Seseña, S.; Palop, M.L. Seasonal diversity and safety evaluation of enterococci population from goat milk in a farm. Dairy Sci. Technol. 2016, 96, 359–375. [Google Scholar] [CrossRef] [Green Version]
- Cocolin, L.; Foschino, R.; Comi, G.; Fortina, M.G. Description of the bacteriocins produced by two strains of Enterococcus faecium isolated from Italian goat milk. Food Microbiol. 2007, 24, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Perin, L.M.; Todorov, S.D.; Nero, L.A. Investigation of genes involved in nisin production in Enterococcus spp. strains isolated from raw goat milk. Antonie van Leeuwenhoek 2016, 109, 1271–1280. [Google Scholar] [CrossRef]
- Perin, L.M.; Nero, L.A. Antagonistic lactic acid bacteria isolated from goat milk and identification of a novel nisin variant Lactococcus lactis. BMC Microbiol. 2014, 14, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Júnior, W.L.G.A.; Ferrari, I.S.; Souza, J.V.; Silva, C.D.A.; Costa, M.M.; Dias, F.S. Characterization and evaluation of lactic acid bacteria isolated from goat milk. Food Control 2015, 53, 96–103. [Google Scholar] [CrossRef]
- Sarkar, S.L.; Hossain, M.I.; Monika, S.A.; Sanyal, S.K.; Roy, P.C.; Hossain, M.A.; Jahid, I.K. Probiotic potential of Pediococcus acidilactici and Enterococcus faecium isolated from indigenous yogurt and raw goat milk. Microbiol. Biotechnol. Lett. 2020, 48, 276–286. [Google Scholar] [CrossRef]
- Achemchem, F.; Cebrián, R.; Abrini, J.; Martínez-Bueno, M.; Valdivia, E.; Maqueda, M. Antimicrobial characterization and safety aspects of the bacteriocinogenic Enterococcus hirae F420 isolated from Moroccan raw goat milk. Can. J. Microbiol. 2012, 58, 596–604. [Google Scholar] [CrossRef]
- Callon, C.; Duthoit, F.; Delbès, C.; Ferrand, M.; Le Frileux, Y.; De Crémoux, R.; Montel, M.-C. Stability of microbial communities in goat milk during a lactation year: Molecular approaches. Syst. Appl. Microbiol. 2007, 30, 547–560. [Google Scholar] [CrossRef] [PubMed]
- Perin, L.M.; Pereira, J.G.; Bersot, L.S.; Nero, L.A. The microbiology of raw milk. In Raw Milk: Balance between Hazards and Benefits; Nero, L.A., Carvalho, A.F., Eds.; Academic Press: Chennai, India, 2019; pp. 45–64. [Google Scholar]
- Franciosi, E.; Settanni, L.; Cavazza, A.; Poznanski, E. Biodiversity and technological potential of wild lactic acid bacteria from raw cow’s milk. Int. Dairy J. 2009, 19, 3–11. [Google Scholar] [CrossRef]
- Hantsis-Zacharov, E.; Halpern, M. Culturable psychrotrophic bacterial communities in raw milk and their proteolytic and lipolytic traits. Appl. Environ. Microbiol. 2007, 73, 7162–7168. [Google Scholar] [CrossRef] [Green Version]
- Banwo, K.; Sanni, A.; Tan, H. Technological properties and probiotic potential of Enterococcus faecium strains isolated from cow milk. J. Appl. Microbiol. 2012, 114, 229–241. [Google Scholar] [CrossRef]
- Elkenany, R.M.; Elsayed, M.M.; Eltaysh, R.A.; Zakaria, A.I.; El-Baz, A.H. In vitro probiotic characteristics of Enterococcus species isolated from raw cow milk. Int. J. Probiotics Prebiotics 2018, 13, 117. [Google Scholar]
- Bouton, Y.; Guyot, P.; Grappin, R. Preliminary characterization of microflora of Comté cheese. J. Appl. Microbiol. 1998, 85, 123–131. [Google Scholar] [CrossRef]
- Pappa, E.C.; Kondyli, E.; Samelis, J. Microbiological and biochemical characteristics of Kashkaval cheese produced using pasteurized or raw milk. Int. Dairy J. 2019, 89, 60–67. [Google Scholar] [CrossRef]
- Dal Bello, B.; Rantsiou, K.; Bellio, A.; Zeppa, G.; Ambrosoli, R.; Civera, T.; Cocolin, L. Microbial ecology of artisanal products from North West of Italy and antimicrobial activity of the autochthonous populations. LWT Food Sci. Technol. 2010, 43, 1151–1159. [Google Scholar] [CrossRef]
- Morandi, S.; Brasca, M.; Lodi, R. Technological, phenotypic and genotypic charaterisation of wild lactic acid bacteria involved in the production of Bitto PDO Italian cheese. Dairy Sci. Technol. 2011, 91, 341–359. [Google Scholar] [CrossRef]
- Calasso, M.; Ercolini, D.; Mancini, L.; Stellato, G.; Minervini, F.; Di Cagno, R.; De Angelis, M.; Gobetti, M. Relationships among house, rind, and core microbiotas during manufacture of traditional Italian cheeses at the same dairy plant. Food Microbiol. 2016, 54, 115–126. [Google Scholar] [CrossRef]
- Di Grigoli, A.; Francesca, N.; Gaglio, R.; Guarrasi, V.; Moschetti, M.; Scatassa, M.L.; Settanni, L.; Bonanno, A. The influence of the wooden equipment employed for cheese manufacture on the characteristics of a traditional stretched cheese during ripening. Food Microbiol. 2015, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casalta, E.; Sorba, J.-M.; Aigle, M.; Ogier, J.-C. Diversity and dynamics of the microbial community during the manufacture of Calenzana, an artisanal Corsican cheese. Int. J. Food Microbiol. 2009, 133, 243–251. [Google Scholar] [CrossRef]
- Pisano, M.B.; Fadda, M.E.; Deplano, M.; Corda, A.; Cosentino, S. Microbiological and chemical characterization of Fiore Sardo, a traditional Sardinian cheese made from ewe’s milk. Int. J. Dairy Technol. 2006, 59, 171–179. [Google Scholar] [CrossRef]
- Carraro, L.; Maifreni, M.; Bartolomeoli, I.; Martino, M.E.; Novelli, E.; Frigo, F.; Marino, M.; Cardazzo, B. Comparison of culture-dependent and -independent methods for bacterial community monitoring during Montasio cheese manufacturing. Res. Microbiol. 2011, 162, 231–239. [Google Scholar] [CrossRef]
- Marino, M.; Maifreni, M.; Rondinini, G. Microbiological characterization of artisanal Montasio cheese: Analysis of its indigenous lactic acid bacteria. FEMS Microbiol. Lett 2003, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Franciosi, E.; Carafa, I.; Nardin, T.; Schiavon, S.; Poznanski, E.; Cavazza, A.; Larcher, R.; Tuohy, K.M. Biodiversity and γ-aminobutyric acid production by lactic acid bacteria isolated from traditional alpine raw cow’s milk cheeses. BioMed Res. Int. 2014, 2015, 625740. [Google Scholar] [CrossRef] [Green Version]
- Poznanski, E.; Cavazza, A.; Cappa, F.; Cocconcelli, P.S. Indigenous raw milk microbiota influences the bacterial development in traditional cheese from an alpine natural park. Int. J. Food Microbiol. 2004, 92, 141–151. [Google Scholar] [CrossRef]
- Randazzo, C.; Vaughan, E.E.; Caggia, C. Artisanal and experimental Pecorino Siciliano cheese: Microbial dynamics during manufacture assessed by culturing and PCR-DGGE analyses. Int. J. Food Microbiol. 2006, 109, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aponte, M.; Fusco, V.; Andolfi, R.; Coppola, S. Lactic acid bacteria occurring during manufacture and ripening of Provolone del Monaco cheese: Detection by different analytical approaches. Int. Dairy J. 2008, 18, 403–413. [Google Scholar] [CrossRef] [Green Version]
- Dolci, P.; Alessandria, V.; Zeppa, G.; Rantsiou, K.; Cocolin, L. Microbiological characterization of artisanal Raschera PDO cheese: Analysis of its indigenous lactic acid bacteria. Food Microbiol. 2008, 25, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Suzzi, G.; Caruso, M.; Gardini, F.; Lombardi, A.; Vannini, L.; Guerzoni, M.E.; Andrighetto, C.; Lanorte, M.T. A survey of the enterococci isolated from an artisanal Italian goat’s cheese (semicotto caprino). J. Appl. Microbiol. 2000, 89, 267–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombo, E.; Franzetti, L.; Frusca, M.; Scarpellini, M. Phenotypic and genotypic characterization of lactic acid bacteria isolated from artisanal Italian goat cheese. J. Food Prot. 2010, 73, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Gaglio, R.; Cruciata, M.; Scatassa, M.L.; Tolone, M.; Mancuso, I.; Cardamone, C.; Corona, O.; Todaro, M.; Settanni, L. Influence of the early bacterial biofilms developed on vats made with seven wood types on PDO Vastedda della valle del Belice cheese characteristics. Int. J. Food Microbiol. 2019, 291, 91–103. [Google Scholar] [CrossRef]
- Tavaria, F.; Malcata, F.X. Microbiological characterization of Serra da Estrela cheese throughout its Appelation d’Origine Protégée Region. J. Food Prot. 1998, 61, 601–607. [Google Scholar] [CrossRef] [PubMed]
- Macedo, A.C.; Tavares, T.G.; Malcata, F.X. Influence of native lactic acid bacteria on the microbiological, biochemical and sensory profiles of Serra da Estrela cheese. Food Microbiol. 2004, 21, 233–240. [Google Scholar] [CrossRef]
- Pinho, O.; Pintado, A.I.E.; Gomes, A.M.P.; Pintado, M.E.; Malcata, F.X.; Ferreira, I.M.P.L.V.O. Interrelationships among microbiological, physicochemical, and biochemical properties of Terrincho cheese, with emphasis on biogenic amines. J. Food Prot. 2004, 67, 2779–2785. [Google Scholar] [CrossRef]
- Alegría, A.; Álvarez-Martín, P.; Sacristán, N.; Fernández, E.; Delgado, S.; Mayo, B. Diversity and evolution of the microbial populations during manufacture and ripening of Casín, a traditional Spanish, starter-free cheese made from cow’s milk. Int. J. Food Microbiol. 2009, 136, 44–51. [Google Scholar] [CrossRef] [Green Version]
- Martín-Platero, A.M.; Valdivia, E.; Maqueda, M.; Martín-Sánchez, I.; Martínez-Bueno, M. Polyphasic approach to bacterial dynamics during the ripening of Spanish farmhouse cheese, using culture-dependent and -independent methods. Appl. Environ. Microbiol. 2008, 74, 5662–5673. [Google Scholar] [CrossRef] [Green Version]
- Picon, A.; Garde, S.; Ávila, M.; Nuñez, M. Microbiota dynamics and lactic acid bacteria biodiversity in raw goat milk cheeses. Int. Dairy J. 2016, 58, 14–22. [Google Scholar] [CrossRef]
- Diezhandino, I.; Fernández, D.; González, L.; McSweeney, P.L.H.; Fresno, J.M. Microbiological, physico-chemical and proteolytic changes in a Spanish blue cheese during ripening (Valdeón cheese). Food Chem. 2015, 168, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Dráb, V.; Kavková, M.; Markvartová, M.; Hanáková, J.; Roubal, P. Microbial diversity of Livanjski cheese with the emphasis on lactic acid bacteria based on culture-dependent and sequencing method. Int. J. Dairy Technol. 2020, 73, 202–214. [Google Scholar] [CrossRef]
- De Freitas, I.; Pinon, N.; Thierry, A.; Lopez, C.; Maubois, J.-L.; Lortal, S. In depth dynamic characterization of French PDO Cantal cheese made from raw milk. Lait 2007, 87, 97–117. [Google Scholar] [CrossRef]
- Asteri, I.-A.; Robertson, N.; Kagkli, D.-M.; Andrewes, P.; Nychas, G.; Coolbear, T.; Holland, R.; Crow, V.; Tsakalidou, E. Technological and flavour potential of cultures isolated from traditional Greek cheeses—A pool of novel species and starters. Int. Dairy J. 2009, 19, 595–604. [Google Scholar] [CrossRef]
- Samelis, J.; Kakouri, A. Major technological differences between and industrial-type and five artisan-type Greek PDO Galotyri market cheeses as revealed by great variations in their lactic acid microbiota. AIMS Agric. Food 2019, 4, 685–710. [Google Scholar] [CrossRef]
- Giannou, E.; Lianou, A.; Kakouri, A.; Kallimanis, A.; Drainas, C.; Samelis, J. Identification and biopreservation potential of Enterococcus spp. isolated from fully ripened Graviera, a traditional hard Greek cheese. Ital. J. Food Sci. 2009, 21, 135–148. [Google Scholar]
- Vandera, E.; Kakouri, A.; Koukkou, A.-I.; Samelis, J. Major ecological shifts within the dominant nonstarter lactic acid bacteria in mature Greek Graviera cheese as affected by the starter culture type. Int. J. Food Microbiol. 2019, 290, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Zoumpopoulou, G.; Papadimitriou, K.; Alexandraki, V.; Mavrogonatou, E.; Alexopoulou, K.; Anastasiou, R.; Gerogalaki, M.; Kletsas, D.; Tsakalidou, E.; Giaouris, E. The microbiota of Kalathaki and Melichloro Greek artisanal cheeses comprises functional lactic acid bacteria. LWT Food Sci. Technol. 2020, 130, 109570. [Google Scholar] [CrossRef]
- Serio, A.; Paparella, A.; Chaves-López, C.; Corsetti, A.; Suzzi, G. Enterococcus populations in Pecorino Abruzzese cheese: Biodiversity and safety aspects. J. Food Prot. 2007, 70, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Campolo, O.; Romeo, F.V.; Attinà, A.; Zappalà, L.; Palmeri, V. Hygienic and physicochemical quality characterization of artisanal and industrial Pecorino Calabrese cheese. Int. J. Dairy Technol. 2013, 66, 595–603. [Google Scholar] [CrossRef]
- Bonomo, M.G.; Salzano, G. Microbial diversity and dynamics of Pecorino di Filiano PDO, a traditional cheese of Basilicata region (Southern Italy). Int. J. Dairy Technol. 2012, 65, 1–11. [Google Scholar] [CrossRef]
- Mormile, A.; Barile, M.; Mercogliano, R.; Johansson, P.; Björkroth, K.J.; Aponte, M.; Murru, N. Dynamics of lactic acid bacteria in “Pecorino di Tramonti”–a ewe’s milk cheese–with particular emphasis on enterococci: A preliminary study. Ann. Microbiol. 2016, 66, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Todaro, M.; Francesca, N.; Reale, S.; Moschetti, G.; Vitale, F.; Settanni, L. Effect of different salting technologies on the chemical and microbiological characteristics of PDO Pecorino Siciliano cheese. Eur. Food Res. Technol. 2011, 233, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Todaro, M.; Palmeri, M.; Settanni, L.; Scatassa, M.L. Effect of refrigerated storage on microbiological, chemical and sensory characteristics of a ewe’s raw milk stretched cheese. Food Packag. Shelf Life 2017, 11, 67–73. [Google Scholar] [CrossRef] [Green Version]
- Casalta, E.; Zennaro, R. Effect of specific starters on microbiological, biochemical and sensory characteristics of Venaco, a Corsican soft cheese. Sci. Aliment. 1997, 17, 79–94. [Google Scholar]
- Freitas, A.C.; Malcata, F.X. Influence of milk type, coagulant, salting procedure and ripening time on the final characteristics of Picante cheese. Int. Dairy J. 1996, 6, 109–1116. [Google Scholar] [CrossRef]
- Freitas, A.C.; Malcata, F.X. Technological optimization of Picante cheese using microbiological, chemical and physical criteria. J. Food Eng. 1999, 41, 163–175. [Google Scholar] [CrossRef]
- Gonçalves, M.T.P.; Benito, M.J.; Córdoba, M.G.; Egas, C.; Merchán, A.V.; Galván, A.I.; Ruiz-Moyano, S. Bacterial communities in Serpa cheese by culture dependent techniques, 16S rRNA gene senquencing and high-throughput sequencing analysis. J. Food Sci. 2018, 83, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Dahl, S.; Tavaria, F.K.; Malcata, F.X. Relationships between flavour and microbiological profiles in Serra da Estrela cheese throughout ripening. Int. Dairy J. 2000, 10, 255–262. [Google Scholar] [CrossRef]
- Tavaria, F.K.; Malcata, F.X. On the microbiology of Serra da Estrela cheese: Geographical and chronological considerations. Food Microbiol. 2000, 17, 293–304. [Google Scholar] [CrossRef]
- Macedo, A.C.; Malcata, F.X. Technological optimization of the manufacture of Serra cheese. J. Food Eng. 1997, 31, 433–447. [Google Scholar] [CrossRef]
- Pintado, A.I.; Pinho, O.; Ferreira, I.M.; Pintado, M.M.; Gomes, A.M.; Malcata, F.X. Microbiological, biochemical and biogenic amine profiles of Terrincho cheese manufactured in several dairy farms. Int. Dairy J. 2008, 18, 631–640. [Google Scholar] [CrossRef]
- Muruzović, M.Ž.; Mladenović, K.G.; Žugić-Petrović, T.D.; Čomić, L.R. Characterization of lactic acid bacteria isolated from traditionally made Serbian Cheese and evaluation of their antagonistic potential against Enterobacteriaceae. J. Food Process. Preserv. 2018, 42, e13577. [Google Scholar] [CrossRef]
- Abriouel, H.; Martín-Platero, A.; Maqueda, M.; Valdivia, E.; Martínez-Bueno, M. Biodiversity of the microbial community in a Spanish farmhouse cheese as revealed by culture-dependent and culture-independent methods. Int. J. Food Microbiol. 2008, 127, 200–208. [Google Scholar] [CrossRef]
- Martín-Platero, A.M.; Maqueda, M.; Valdivia, E.; Purswani, J.; Martínez-Bueno, M. Polyphasic study of microbial communities of two Spanish farmhouse goats’ milk cheeses from Sierra de Aracena. Food Microbiol. 2009, 26, 294–304. [Google Scholar] [CrossRef]
- Ordiales, E.; Martín, A.; Benito, M.J.; Hernández, A.; Ruiz-Moyano, S.; Córdoba, M.G. Role of the microbial population on the flavor of the soft-bodied cheese Torta del Casar. J. Dairy Sci. 2013, 96, 5477–5486. [Google Scholar] [CrossRef]
- Yunita, D.; Dodd, C.E.R. Microbial community dynamics of a blue-veined raw milk cheese from the United Kingdom. J. Dairy Sci. 2017, 101, 4923–4935. [Google Scholar] [CrossRef]
- Van Hoorde, K.; Verstraete, T.; Vandamme, P.; Huys, G. Diversity of lactic acid bacteria in two Flemish artisan raw milk Gouda-type cheeses. Food Microbiol. 2008, 25, 929–935. [Google Scholar] [CrossRef]
- Delgado, S.; Mayo, B. Phenotypic and genetic diversity of Lactococcus lactis and Enterococcus spp. strains isolated from Northern Spain starter-free farmhouse cheeses. Int. J. Food Microbiol. 2004, 90, 309–319. [Google Scholar] [CrossRef]
- Colombo, F.; Borgo, F.; Fortina, M.G. Genotypic characterization of non-starter lactic acid bacteria involved in the ripening of artisanal Bitto PDO cheese. J. Basic Microbiol. 2009, 49, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Jurkovic, D.; Križková, L.; Sojka, M.; Belicová, A.; Dušinský, R.; Krajčovič, J.; Snauwaert, C.; Naser, S.; Vandamme, P.; Vancanneyt, M. Molecular identification and diversity of enterococci isolated from Slovak Bryndza cheese. J. Gen. Appl. Microbiol. 2006, 52, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurkovic, D.; Križková, L.; Sojka, M.; Takáčova, M.; Dušinský, R.; Krajčovič, J.; Vandamme, P.; Vancanneyt, M. Genetic diversity of Enterococcus faecium isolated from Bryndza cheese. Int. J. Food Microbiol. 2007, 116, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Pangallo, D.; Šaková, N.; Koreňová, J.; Puškárová, A.; Kraková, L.; Valík, L.; Kuchta, T. Microbial diversity and dynamics during the production of May bryndza cheese. Int J. Food Microbiol. 2014, 170, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Guarrasi, V.; Sannino, C.; Moschetti, M.; Bonanno, A.; Di Grigoli, A.; Settani, L. The individual contribution of starter and non-starter lactic acid bacteria to the volatile organic compound composition of Caciocavallo Palermitano cheese. Int. J. Food Microbiol. 2017, 259, 35–42. [Google Scholar] [CrossRef]
- Aquilanti, L.; Dell’Aquila, L.; Zannini, E.; Zocchetti, A.; Clementi, F. Resident lactic acid bacteria in raw milk Canestrato Pugliese cheese. Lett Appl. Microbiol. 2006, 43, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Fontana, C.; Cappa, F.; Rebecchi, A.; Cocconcelli, P.S. Surface microbiota analysis of Taleggio, Gorgonzolla, Casera, Scimudin and Fromaggio di Fossa Italian cheeses. Int. J. Food Microbiol. 2010, 138, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Morandi, S.; Silvetti, T.; Lopez, M.L.; Brasca, M. Antimicrobial activity, antibiotic resistance and the safety of lactic acid bacteria in raw milk Valtellina Casera cheese. J. Food Saf. 2015, 35, 193–205. [Google Scholar] [CrossRef]
- Mangia, N.P.; Fancello, F.; Deiana, P. Microbiological characterization using combined culture dependent and independent approaches of Casizolu pasta filata cheese. J. Appl. Microbiol 2015, 120, 329–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryssel, M.; Johansen, P.; Al-Soud, W.A.; Sørensen, S.; Arneborg, N.; Jespersen, L. Microbial diversity and dynamics throughout manufacturing and ripening of surface ripened semi-hard Danish Danbo cheeses investigated by culture-independent techniques. Int. J. Food Microbiol. 2015, 215, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Bozoudi, D.; Torriani, S.; Zdragas, A.; Litopoulou-Tzanetaki, E. Assessment of microbial diversity of the dominant microbiota in fresh and mature PDO Feta cheese made at three mountainous areas of Greece. LWT Food Sci. Technol. 2016, 72, 525–533. [Google Scholar] [CrossRef]
- Dolci, P.; De Filippis, F.; La Storia, A.; Ercolini, D.; Cocolin, L. rRNA-based monitoring of the microbiota involved in Fontina PDO cheese production in relation to different stages of cow lactation. Int. J. Food Microbiol. 2014, 185, 127–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senini, L.; Cappa, F.; Cocconcelli, P.S. Use of rRNA-targeted oligonucleotide probes for the characterization of the microflora from fermentation of Fontina cheese. Food Microbiol. 1997, 14, 469–476. [Google Scholar] [CrossRef]
- Barbieri, E.; Schiavano, G.F.; De Santi, M.; Vallorani, L.; Casadei, L.; Guescini, M.; Gioacchini, A.M.; Rinaldi, L.; Stocchi, V.; Brandi, G. Bacterial diversity of traditional Fossa (pit) cheese and its ripening environment. Int. Dairy J. 2012, 23, 62–67. [Google Scholar] [CrossRef]
- Pavunc, A.L.; Beganović Kos, B.; Uroić, K.; Blažić, M.; Šušković, J. Characterization and application of autochthonous starter cultures for fresh cheese production. Food Technol. Biotechnol. 2012, 50, 141–151. [Google Scholar]
- Gonzalez, L.; Sacristán, N.; Arenas, R.; Fresno, J.M.; Tornadijo, M.E. Enzymatic activity of lactic acid bacteria (with antimicrobial properties) isolated from a traditional Spanish cheese. Food Microbiol. 2010, 27, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Van Hoorde, K.; Heyndrickx, M.; Vandamme, P.; Huys, G. Influence of pasteurization, brining conditions and production environment on the microbiota of artisan Gouda-type cheeses. Food Microbiol. 2010, 27, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Delcenserie, V.; Taminiau, B.; Delhalle, L.; Nezer, C.; Doyen, P.; Crevecoeur, S.; Roussey, D.; Korsak, N.; Daube, G. Microbiota characterization of a Belgian protected designation of origin cheese, Herve cheese, using metagenomic analysis. J. Dairy Sci. 2014, 97, 6046–6056. [Google Scholar] [CrossRef] [Green Version]
- Fuka, M.M.; Engel, M.; Skelin, M.; Redžepović, S.; Schloter, M. Bacterial communities associated with the production of artisanal Istrian cheese. Int. J. Food Microbiol. 2010, 142, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Nieto-Arribas, P.; Seseña, S.; Poveda, J.M.; Chicón, R.; Cabezas, L.; Palop, L. Enterococcus populations in artisanal Manchego cheese: Biodiversity, technological and safety aspects. Food Microbiol. 2011, 28, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Nacef, M.; Chevalier, M.; Chollet, S.; Drider, D.; Flahaut, C. MALDI-TOF mass spectrometry for the identification of lactic acid bacteria isolated from a French cheese: The Maroilles. Int. J. Food Microbiol. 2017, 247, 2–8. [Google Scholar] [CrossRef]
- Morea, M.; Baruzzi, F.; Cocconcelli, P.S. Molecular and physiological characterization of dominant bacterial populations in traditional Mozzarella cheese processing. J. Appl. Microbiol. 1999, 87, 574–582. [Google Scholar] [CrossRef]
- Alegría, Á.; Szczesny, P.; Mayo, N.; Barodowski, J.; Kowalczyk, M. Biodiversity in Oscypek, a traditional Polish cheese, determined by culture-dependent and -independent approaches. Appl. Environ. Microbiol. 2012, 78, 1890–1898. [Google Scholar] [CrossRef] [Green Version]
- Aquilanti, L.; Silvestri, G.; Zannini, E.; Osimani, A.; Santarelli, S.; Clementi, F. Phenotypic, genotypic and technological characterization of predominant lactic acid bacteria in Pecorino cheese from central Italy. J. Appl. Microbiol. 2007, 103, 948–960. [Google Scholar] [CrossRef]
- De Pasquale, I.; Di Cagno, R.; Buchin, S.; De Angelis, M.; Gobbetti, M. Use of autochthonous mesophilic lactic acid bacteria as starter cultures for making Pecorino Crotonese cheese: Effect on compositional, microbiological and biochemical attributes. Food Res. Int. 2019, 116, 1344–1356. [Google Scholar] [CrossRef] [PubMed]
- Russo, N.; Caggia, C.; Pino, A.; Coque, T.M.; Aroli, S.; Randazzo, C.L. Enterococcus spp. in Ragusano PDO and Pecorino Siciliano cheese types: A snapshot of their antibiotic resistance distribution. Food Chem. Toxicol. 2018, 120, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.C.; Pais, C.; Malcata, F.X.; Hogg, T.A. Microbiological characterization of Picante da Beira Baixa cheese. J. Food Prot. 1996, 59, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.C.; Pintado, A.E.; Manuela, E.P.; Malcata, F.X. Role of dominant microflora of Picante cheese on proteolysis and lipolysis. Int. Dairy J. 1999, 9, 593–603. [Google Scholar] [CrossRef]
- Freitas, A.C.; Fresno, J.M.; Prieto, B.; Franco, I.; Malcata, F.X.; Carballo, J. How milk type, coagulant, salting procedure and ripening time affect the profile of free amino acids in Picante da Beira Baixa cheese. J. Sci. Food Agric. 1999, 79, 611–618. [Google Scholar] [CrossRef]
- Câmara, S.P.A.; Dapkevicius, A.; Silva, C.C.G.; Malcata, F.X.; Dapkevicius, M.L.N.E. Artisanal Pico cheese as reservoir of Enterococcus species possessing virulence and antibiotic resistance properties: Implications for food safety. Food Biotechnol. 2020, 34, 25–41. [Google Scholar] [CrossRef]
- Domingos-Lopes, M.F.P.; Stanton, C.; Ross, P.R.; Dapkevicius, M.L.E.; Silva, C.C.G. Genetic diversity, safety and technological characterization of lactic acid bacteria isolated from artisanal Pico cheese. Food Microbiol. 2017, 63, 178–190. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Coelho, M.C.; Todorov, S.D.; Franco, B.D.G.M.; Dapkevicius, M.L.E.; Silva, C.C.G. Technological properties of bacteriocin-producing lactic acid bacteria isolated from Pico cheese, an artisanal cow’s milk cheese. J. Appl. Microbiol. 2014, 116, 573–585. [Google Scholar] [CrossRef]
- Kongo, J.M.; Gomes, A.M.; Malcata, F.X.; McSweeney, P.H.L. Microbiological, biochemical and compositional changes during ripening of São Jorge–a raw milk cheese from the Azores (Portugal). Food Chem. 2009, 112, 131–138. [Google Scholar] [CrossRef]
- Delbès, C.; Ali-Mandjee, L.; Montel, M.-C. Monitoring bacterial communities in raw-milk and cheese by culture-dependent and -independent 16S rRNA gene-based analyses. Appl. Environ. Microbiol. 2007, 73, 1882–1891. [Google Scholar] [CrossRef] [Green Version]
- Duthoit, F.; Godon, J.-J.; Montel, M.C. Bacterial community dynamics during production of registered designation of origin Salers cheese as evaluated by 16S rRNA gene single-strand conformation polymorphism analysis. Appl. Environ. Microbiol. 2003, 69, 3840–3848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duthoit, F.; Tessier, L.; Montel, M.C. Diversity, dynamics and activity of bacterial populations in “Registered Designation of Origin” Salers cheese by single-stranded conformation polymorphism analysis of 16S rRNA genes. J. Appl. Microbiol. 2005, 98, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Duthoit, F.; Callon, C.; Tessier, L.; Montel, M.C. Relationship between sensorial characteristics and microbial dynamics in “Registered Designation of Origin” Salers cheese. Int. J. Food Microbiol. 2005, 103, 259–270. [Google Scholar] [CrossRef] [PubMed]
- González, L.; Cuadrillero, A.F.; Castro, J.M.; Bernardo, A.; Tornadijo, M.E. Selection of lactic acid bacteria isolated from San Simón da Costa cheese (PDO) in order to develop an autochthonous starter culture. Adv. Microbiol. 2015, 5, 748–759. [Google Scholar] [CrossRef] [Green Version]
- Ledina, T.; Golob, M.; Djordjević, J.; Magas, V.; Colovic, S.; Bulajic, S. MALDI-TOF mass spectrometry for the identification of Serbian artisanal cheeses microbiota. J. Consum. Prot. Food S 2018, 13, 309–314. [Google Scholar] [CrossRef]
- Macedo, A.C.; Malcata, F.X.; Hogg, T.A. Microbiological profile in Serra ewes’ cheese during ripening. J. Appl. Microbiol. 1995, 79, 1–11. [Google Scholar] [CrossRef]
- Tavaria, F.K.; Tavares, T.G.; Silva-Ferreira, A.C.; Malcata, F.X. Contribution of coagulant and native microflora to the volatile fatty acid profile of an artisanal cheese. Int. Dairy J. 2006, 16, 886–894. [Google Scholar] [CrossRef]
- Ercolini, D.; Hill, P.J.; Dodd, C.E.R. Bacterial community structure and location in Stilton cheese. Appl. Environ. Microbiol. 2003, 69, 3540–3548. [Google Scholar] [CrossRef] [Green Version]
- Pimentel, L.L.; Semedo, T.; Tenreiro, R.; Crespo, M.T.B.; Pintado, M.M.; Malcata, F.X. Assessment of safety of enterococci isolated throughout traditional Terrincho cheesemaking: Virulence factors and antibiotic susceptibility. J. Food Prot. 2007, 70, 2161–2167. [Google Scholar] [CrossRef]
- Majhenic, A.Č.; Rogelj, I.; Perko, B. Enterococi from Tolminc cheese: Population structure, antibiotic susceptibility and incidence of virulence determinants. Int. J. Food Microbiol. 2005, 102, 239–244. [Google Scholar] [CrossRef]
- Fortina, M.G.; Ricci, G.; Acquati, A.; Zeppa, G.; Gandini, A.; Manachini, P.L. Genetic characterization of some lactic acid bacteria occurring in an artisanal protected denomination origin (PDO) Italian cheese, the Toma piemontese. Food Microbiol. 2003, 20, 397–404. [Google Scholar] [CrossRef]
- Terzic-Vidojevic, A.; Tolinacki, M.; Nikolic, M.; Vejovic, K.; Jovanovic, S.; Macej, O.; Topisirovic, L. Artisanal Vlasina raw goat’s milk cheese: Evaluation and selection of autochthonous lactic acid bacteria as starter cultures. Food Technol. Biotechnol. 2013, 51, 554–563. [Google Scholar]
- Golić, N.; Čadež Terzić-Vidojević, A.; Šuranská, H.; Begnović, J.; Lozo, J.; Kos, B.; Šušković, J.; Raspor, P.; Topisirović, L. Evaluation of lactic acid bacteria and yeast diversity in traditional white pickled and fresh soft cheeses from the mountain regions of Serbia and lowland regions of Croatia. Int. J. Food Microbiol. 2013, 166, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Veljovic, K.; Terzic-Vidojevic, A.; Vukasinovic, M.; Strahinic, I.; Begovic, J.; Lozo, J.; Ostojic, M.; Topisirovic, L. Preliminary characterization of lactic acid bacteria isolated from Zlatar cheese. J. Appl. Microbiol. 2007, 103, 2141–2152. [Google Scholar] [CrossRef]
- Yerlikaya, O.; Akbulut, N. Potential use of probiotic Enterococcus faecium and Enterococcus durans strains in Izmir Tulum cheese as adjunct culture. J. Food Sci. Technol. 2019, 56, 2175–2185. [Google Scholar] [CrossRef] [PubMed]
- Malek, R.; El-Attar, A.; Mohamed, M.; Anwar, S.; El-Soda, M.; Béal, C. Technological and safety properties display biodiversity among enterococci isolated from two Egyptian cheeses, “Ras” and “Domiati”. Int. J. Food Microbiol. 2012, 153, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Mc Sweeney, P.L.H. Biochemistry of cheese ripening. Int. J. Dairy Technol 2004, 57, 127–144. [Google Scholar] [CrossRef]
- Câmara, S.P.A.; Dapkevicius, A.; Riquelme, C.; Elias, R.B.; Silva, C.C.G.; Malcata, F.X.; Dapkevicius, M.L.N.E. Potential of lactic acid bacteria from Pico cheese for starter culture development. Food Sci. Technol. Int. 2019, 25, 303–317. [Google Scholar] [CrossRef]
- Serio, A.; Chaves-López, C.; Paparella, A.; Suzzi, G. Evaluation of metabolic activities of enterococci isolated from Pecorino Abruzzese cheese. Int. Dairy J. 2010, 20, 459–464. [Google Scholar] [CrossRef]
- Andrighetto, C.; Knijff, E.; Lombardi, A.; Torriani, S.; Vancanneyt, M.; Kersters, K.; Swings, J.; Dellaglio, F. Phenotypic and genetic diversity of enterococci isolated from Italian cheeses. J. Dairy Res. 2001, 68, 303–316. [Google Scholar] [CrossRef]
- Durlu-Ozkaya, F.; Xanthopoulous, V.; Tunail, N.; Litopoulou-Tzanetaki, E. Technologically important properties of lactic acid bacteria isolates from Beyaz cheese made from raw ewes’ milk. J. Appl. Microbiol. 2001, 91, 861–870. [Google Scholar] [CrossRef]
- Sarantinopoulos, P.; Andrighetto, C.; Gerogalaki, M.D.; Rea, M.C.; Lombardi, A.; Cogan, T.M.; Kalantzopoulos, G.; Tsakalidou, E. Biochemical properties of enterococci relevant to their technological performance. Int. Dairy J. 2001, 11, 621–647. [Google Scholar] [CrossRef]
- Morandi, S.; Brasca, M.; Andrighetto, C.; Lombardi, A.; Lodi, R. Technological and molecular characterization of enterococci isolated from north-west Italian dairy products. Int. Dairy J. 2006, 16, 867–875. [Google Scholar] [CrossRef]
- Margalho, L.P.; Schalkwijk, S.; Bachmann, H.; Sant’Ana, A.A. Enterococcus spp. in Brazilian artisanal cheeses: Occurrence and assessment of phenotypic and safety properties of a large set of strains through the use of high throughput tools combined with multivariate statistics. Food Control 2020, 118, 107425. [Google Scholar] [CrossRef]
- Dagdemir, E.; Ozdemir, S. Technological characterization of the natural lactic acid bacteria of artisanal Turkish White Pickled cheese. Int. J. Dairy Technol. 2008, 61, 133–140. [Google Scholar] [CrossRef]
- Tuncer, Y. Some technological properties of phenotypically identified enterococci strains isolated from Turkish Tulum cheese. Afr. J. Biotechnol. 2009, 8, 7008–7016. [Google Scholar]
- Gútiez, L.; Gómez-Sala, B.; Recio, I.; Del Campo, R.; Cintas, L.M.; Herranz, C.; Hernández, P.E. Enterococcus faecalis strains from food, environmental and clinical origin produce ACE-inhibitory peptides and other bioactive peptides during growth in bovine skim milk. Int. J. Food Microbiol. 2013, 166, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Van Tyne, D.; Gilmore, M.S. Friend turned foe: Evolution of enterococcal virulence and antibiotic resistance. Ann. Rev. Microbiol. 2014, 68, 337–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christensen, J.E.; Dudley, E.G.; Pederson, A.; Steele, J.L. Peptidases and amino acid catabolism in lactic acid bacteria. Antonie van Leeuwenhoek 1999, 76, 217–246. [Google Scholar] [CrossRef]
- Olvera-García, M.; Sanchez-Flores, A.; Baruch, M.Q. Genomic and functional characterization of two Enterococcus strains isolated from Cotija cheese and their potential role in ripening. Appl. Microbiol. Biotechnol. 2018, 102, 2251–2267. [Google Scholar] [CrossRef]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic amine production by lactic acid bacteria: A review. Foods 2019, 8, 17. [Google Scholar] [CrossRef] [Green Version]
- Mc Donnell, M.; Fitzgerald, R.; Fhaoláin, I.N.; Jennings, V.; O’Cuinn, G. Purification and characterization of aminopeptidase P from Lactococcus lactis subsp. cremoris. J. Dairy Res. 1997, 64, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Perin, L.M.; Belviso, S.; Dal Bello, B.; Nero, L.A.; Cocolin, L. Technological properties and biogenic amines production by bacteriocinogenic lactococci and enterococci strains isolated from raw goat’s milk. J. Food Prot. 2017, 80, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Lortal, S.; Chapot-Chartier, M.-P. Role, mechanisms and control of lactic acid bacteria lysis in cheese. Int. Dairy J. 2005, 15, 857–871. [Google Scholar] [CrossRef] [Green Version]
- El-Din, B.B.; El-Soda, M.; Ezzat, N. Proteolytic, lipolytic and autolytic activities of enterococci strains isolated from Egyptian dairy products. Lait 2002, 82, 289–304. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, G.E.; Ross, P.R.; Wallace, J.M.; Scanlan, F.P.; Jägers, P.P.J.M.; Fitzgerald, F.F.; Collins, J.K.; Stanton, C. Influence of a probiotic adjunct culture of Enterococcus faecium on the quality of Cheddar cheese. J. Agric. Food Chem. 1999, 47, 4907–4916. [Google Scholar] [CrossRef]
- Fortina, M.G.; Ricci, G.; Borgo, F.; Manachini, P.L.; Arends, K.; Schiwon, K.; Abajy, M.Y.; Grohmann, E. A survey on biotechnological potential and safety of the novel Enterococcus species of dairy origin, E. italicus. Int. J. Food Microbiol. 2008, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Irlinger, F.; Helinck, S.; Jany, J.L. Chapter 11—Secondary and adjunct cultures. In Cheese, 4th ed.; McSweeney, P.L.H., Fox, P.F.F., Cotter, P.D., Everett, D.W., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 273–300. [Google Scholar]
- Lynch, K.M.; Zannini, E.; Coffey, A.; Arendt, E.K. Lactic acid bacteria exopolysaccharides in foods and beverages: Isolation, properties, characterization and health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 155–176. [Google Scholar] [CrossRef]
- Quintans, N.G.; Blancato, V.; Repizo, G.; Magni, C.; López, P. Citrate metabolism and aroma compound production in lactic acid bacteria. In Molecular Aspects of Lactic Acid Bacteria for Traditional and New Applications; Mayo, B., López, P., Pérez-Martínez, G., Eds.; Research Signpost: Kerala, India, 2008; pp. 65–88. [Google Scholar]
- Sarantinopoulos, P.; Kalantzpoulos, G.; Tsakalidou, E. Effect of Enterococcus faecium on microbiological, physicochemical and sensory characteristics of Greek Feta cheese. Int. J. Food Microbiol. 2002, 76, 93–105. [Google Scholar] [CrossRef]
- Abeijón, M.C.; Medina, R.B.; Katz, M.B.; González, S.N. Technological properties of Enterococcus faecium isolated from ewe’s milk and cheese with importance for flavour development. Can. J. Microbiol. 2006, 52, 237–245. [Google Scholar] [CrossRef]
- Martino, G.P.; Espariz, M.; Nizo, G.G.; Esteban, L.; Blancato, V.S.; Magni, C. Safety assessment and functional properties of four enterococci strains isolated from regional Argentinean cheese. Int. J. Food Microbiol. 2018, 277, 1–9. [Google Scholar] [CrossRef]
- Arena, M.P.; Capozzi, V.; Spano, G.; Fiocco, D. The potential of lactic acid bacteria to colonize biotic and abiotic surfaces and the investigation of their interactions and mechanisms. Appl. Microbiol. Biotechnol. 2017, 101, 2641–2657. [Google Scholar] [CrossRef]
- Mende, S.; Rohm, H.; Jaros, D. Influence of exopolysaccharides on the structure, texture, stability and sensory properties of yoghurt and related products. Int. Dairy J. 2016, 52, 57–71. [Google Scholar] [CrossRef]
- Mozzi, F.; Vaningelgem, F.; Hébert, E.M.; Van der Meulen, R.; Foulquié Moreno, M.R.; Valdez, G.F.; De Vuyst, L. Diversity of heteropolysaccharide-producing lactic acid bacterium strains and their biopolymers. Appl. Environ. Microbiol. 2006, 72, 4431–4435. [Google Scholar] [CrossRef] [Green Version]
- Özkan, E.R.; Demirci, T.; Akin, N. In vitro assessment of probiotic and virulence potential of Enterococcus faecium strains derived from artisanal goatskin casing Tulum cheeses produced in central Taurus Mountains of Turkey. LWT Food Sci. Technol. 2021, 141, 110908. [Google Scholar] [CrossRef]
- Constantino, G.; Calasso, M.; Minervini, F.; De Angelis, M. Use of exopolysaccharide-synthesizing lactic acid bacteria and fat replacers for manufacturing reduced-fat Burrata cheese: Microbiological aspects and sensory evaluation. Microorganisms 2020, 8, 1618. [Google Scholar] [CrossRef] [PubMed]
- Yerlikaya, O.; Akpinar, A.; Saygili, D. Analysis of some physicochemical, rheological, sensorial properties, and probiotic viability of fermented milks containing Enterococcus faecium and Enterococcus durans strains. J. Food Process. Preserv. 2020, 44, e14553. [Google Scholar] [CrossRef]
- Cleveland, J.; Montville, T.J.; Nes, I.F.; Chikindas, M.L. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol. 2001, 71, 1–20. [Google Scholar] [CrossRef]
- Javed, A.; Masud, T.; ul Ain, Q.; Imran, M.; Maqsood, S. Enterocins of Enterococcus faecium, emerging natural food preservatives. Ann. Microbiol. 2011, 61, 699–708. [Google Scholar] [CrossRef]
- Nilsen, T.; Nes, I.F.; Holo, H.; Enterolysin, A. A cell wall-degrading bacteriocin from Enterococcus faecalis LMG 2333. Appl. Environ. Microbiol. 2003, 69, 2975–2984. [Google Scholar] [CrossRef] [Green Version]
- Oumer, A.; Gaya, P.; Fernández-García, E.; Mariaca, R.; Garde, S.; Medina, M.; Nuñez, M. Proteolysis and formation of volatile compounds in cheese manufactured with a bacteriocin-producing adjunct culture. J. Dairy Res. 2001, 68, 117–129. [Google Scholar] [CrossRef]
- Ávila, M.; Garde, S.; Medina, M.; Nuñez, M. Effect of milk inoculation with bacteriocin-producing lactic acid bacteria on a Lactobacillus helveticus adjunct cheese culture. J. Food Prot. 2005, 78, 1026–1033. [Google Scholar] [CrossRef]
- Silva, C.C.G.; Silva, S.P.M.; Ribeiro, S.C. Application of bacteriocins and protective cultures in dairy food preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef] [PubMed]
- Aspri, M.; O’Connor, P.M.; Field, D.; Cotter, P.D.; Ross, P.; Hill, C.; Papademas, P. Application of bacteriocin-producing Enterococcus faecium isolated from donkey milk, in the bio-control of Listeria monocytogenes in fresh whey cheese. Int. Dairy J. 2017, 73, 1–9. [Google Scholar] [CrossRef]
- Vandera, E.; Lianou, A.; Kakouri, A.; Feng, J.; Kokkou, A.-I.; Samelis, J. Enhanced control of Listeria monocytogenes by Enterococcus faecium KE82, a multiple enterocin-producing strain, in different milk environments. J. Food Prot. 2017, 80, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.C.; Ross, R.P.; Stanton, C.; Silva, C.C.G. Characterization and application of antilisterial enterocins on model fresh cheese. J. Food Prot. 2017, 80, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, A.; Maqueda, M.; Gálvez, A.; Martínez-Bueno, M.; Rodríguez, A.; Valdivia, E. Biocontrol of psychrotrophic enterotoxigenic Bacillus cereus in a nonfat hard cheese by an enterococcal strain producing enterocin AS-48. J. Food Prot. 2004, 67, 1517–1521. [Google Scholar] [CrossRef]
- Golob, M.; Pate, M.; Kušar, D.; Dermota, U.; Avberšek, J.; Papić, B.; Zdvoc, I. Antimicrobial resistance and virulence genes in Enterococcus faecium and Enterococcus faecalis from humans and retail red meat. BioMed Res. Int. 2019, 2019, 2815279. [Google Scholar] [CrossRef] [Green Version]
- Cui, P.; Feng, L.; He, J.; An, T.; Fu, X.; Li, C.; Zhao, X.; Zhai, Y.; Li, H.; Yan, W.; et al. Antimicrobial resistance, virulence genes and biofilm formation capacity among Enterococcus species from Yaks in Aba Tibetan Autonomous Prefecture, China. Front. Microbiol. 2020, 11, 1250. [Google Scholar] [CrossRef]
- Rahmeh, R.; Akbar, A.; Kishk, M.; Al Onaizi, T.; Al-Shatti, A.; Shajan, A.; Akbar, B.; Al-Mutairi, S.; Yateem, A. Characterization of semipurified enterocins produced by Enterococcus faecium strains isolated from raw camel milk. J. Dairy Sci. 2018, 101, 4944–4952. [Google Scholar] [CrossRef]
- García-Solache, M.; Rice, L.B. The Enterococcus: A model of adaptability to its environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H. Milk-derived bioactive peptides: From science to applications. J. Funct. Foods 2009, 1, 177–187. [Google Scholar] [CrossRef]
- Gútiez, L.; Borrero, J.; Jiménez, J.J.; Gómez-Sala, B.; Recio, I.; Cintas, L.M.; Herranz, C.; Hernández, P.E. Genetic and biochemical evidence that recombinant Enterococcus spp. strains expressing gelatinase (GelE) produce bovine milk-derived hydrolysates with high angiotensin converting-enzyme-inhibitory activity. J. Agric. Food Chem. 2014, 62, 5555–5564. [Google Scholar] [CrossRef] [PubMed]
- Regazzo, D.; Da Dalt, L.; Lombardi, A.; Andrighetto, C.; Negro, A.; Gabai, G. Fermented milks from Enterococcus faecalis TH563 and Lactobacillus delbrueckii subsp. Bulgaricus LA2 manifest different degrees of ACE-inhibitory and immunomodulatory activities. Dairy Sci. Technol. 2010, 90, 469–476. [Google Scholar] [CrossRef]
- Muguerza, B.; Ramos, M.; Sánchez, E.; Manso, M.A.; Miguel, M.; Aleixandre, A.; Delgado, M.A.; Recio, I. Antihypertensive activity of milk fermented by Enterococcus faecalis strains isolated from raw milk. Int. Dairy J. 2006, 26, 61–69. [Google Scholar] [CrossRef]
- Quirós, A.; Ramos, M.; Muguerza, B.; Delgado, M.A.; Miguel, M.; Aleixandre, A.; Recio, I. Identification of novel antihypertensive peptides in milk fermented with Enterococcus faecalis. Int. Dairy J. 2007, 17, 33–41. [Google Scholar] [CrossRef]
- Torres-Llanez, M.J.; González-Córdova, A.F.; Hernandez-Mendoza, A.; Garcia, H.S. Angiotensin-converting enzyme inhibitory activity in Mexican Fresco cheese. J. Dairy Sci. 2011, 94, 3794–3800. [Google Scholar] [CrossRef] [PubMed]
- Nilsen, R.; Pripp, A.H.; Høstmark, A.T.; Haug, A.; Skeie, S. Is consumption of a cheese rich in angiotensin-converting enzyme-inhibiting peptides, such as the Norwegian cheese Gamalost, associated with reduced blood pressure? J. Dairy Sci. 2014, 97, 2662–2668. [Google Scholar] [CrossRef] [Green Version]
- Baptista, D.P.; Gigante, M.L. Bioactive peptides in ripened cheeses: Release during technological processes and resistance to the gastrointestinal tract. J. Sci. Food Agri. 2021. [Google Scholar] [CrossRef]
- Martini, S.; Conte, A.; Tagliazucchi, D. Effect of ripening and in vitro digestion on the evolution and fate of bioactive peptides in Parmigiano-Reggiano cheese. Int. Dairy J. 2020, 105, 104668. [Google Scholar] [CrossRef]
- Lu, Y.; Govindasamy-Lucey, S.; Lucey, J.A. Angiotensin-I-converting enzyme-inhibitory peptides in commercial Wisconsin Cheddar cheeses of different ages. J. Dairy Sci. 2016, 99, 41–52. [Google Scholar] [CrossRef]
- O’Shea, E.F.; Cotter, P.D.; Stanton, C.; Ross, P.R.; Hill, C. Production of bioactive substances by intestinal bacteria as a basis for explaining probiotic mechanisms: Bacteriocins and conjugated linoleic acid. Int. J. Food Microbiol. 2012, 152, 189–205. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, T.; Ud Din, A.; Hassan, A.; Wang, Y.; Wang, G. Beneficial effects of Enterococcus faecalis in hypercholesterolemic mice on cholesterol transportation and gut microbiota. Appl. Microbiol. Biotechnol. 2019, 103, 3181–3191. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhang, F.; Xu, D.; Zhang, Z.; Xu, F.; Tao, X.; Qiu, L.; Wei, H. Enterococcus faecium WEFA23 from infants lessens high-fat diet-induced hyperlipidemia via cholesterol 7-alpha-hydroxylase gene by altering the composition of gut microbiota in rats. J. Dairy Sci. 2018, 101, 7757–7767. [Google Scholar] [CrossRef] [PubMed]
- Albano, C.; Morandi, S.; Silvetti, T.; Casiraghi, M.C.; Manini, F.; Brasca, M. Lactic acid bacteria with cholesterol-lowering properties for dairy applications: In vitro and in situ activity. J. Dairy Sci. 2018, 101, 10807–10818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Qiu, L.; Liu, Z.; Zhan, H.; Tao, X.; Shah, N.; Wei, H. Beneficial effects of probiotic cholesterol-lowering strain of Enterococcus faecium WEFA23 from infants on diet-induced metabolic syndrome in rats. J. Dairy Sci. 2017, 100, 1618–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baccouri, O.; Boukerb, A.M.; Ben Farhat, L.; Zébré, A.; Zimmerman, K.; Domann, E.; Cambronel, M.; Barreau, M.; Maillot, O.; Rincé, I.; et al. Probiotic potential and safety evaluation of Enterococcus faecalis OB14 and OB15, isolated from traditional Tunisian Testouri cheese and Rigouta, using physiological and genomic analysis. Front. Microbiol. 2019, 10, 881. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.; Ascenção, K.; Gullón, P.; Henriques, S.M.S.; Pinto, J.M.S.; Rocha-Santos, T.A.P.; Freitas, A.C.; Gomes, A.M. Production of conjugated linoleic acid by food-grade bacteria: A review. Int. J. Dairy Technol. 2012, 65, 467–481. [Google Scholar] [CrossRef]
- Kishino, S.; Ogawa, J.; Omura, Y.; Matsumura, K.; Shimizu, S. Conjugated linoleic acid production from linoleic acid by lactic acid bacteria. J. Am. Oil Chem. Soc. 2002, 79, 159–163. [Google Scholar] [CrossRef]
- Ross, R.P.; Stanton, C.; Hill, C.; Fitzgeral, G.F.; Coffey, A. Novel cultures for cheese improvement. Trends Food Sci. Technol. 2000, 11, 96–104. [Google Scholar] [CrossRef]
- Del Toro-Barbosa, M.; Hurtado-Romero, A.; Garcia-Amezquita, L.E.; Carcía-Cayuela, T. Psychobiotics: Mechanisms of action, evaluation methods and effectiveness in applications with food products. Nutrients 2020, 12, 3896. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. 2012, 13, 701–710. [Google Scholar] [CrossRef]
- Santos-Espinosa, A.; Beltrán-Barrientos, L.M.; Reyes-Díaz, R.; Mazorra-Manzano, M.A.; Hernández-Mendoza, A.; González-Aguilar, G.A.; Sayago-Ayerdi, S.G.; Vallejo-Cordoba, B.; González-Cárdova, A.F. Gamma-aminobutyric acid (GABA) production in milk fermented by specific wild lactic acid bacteria strains isolated from artisanal Mexican cheeses. Ann. Microbiol. 2020, 70, 12. [Google Scholar] [CrossRef]
- Ribeiro, S.C.; Domingos-Lopes, M.F.P.; Stanton, C.; Ross, R.P.; Silva, C.C.G. Production of γ-aminobutyric acid (GABA) by Lactobacillus otakiensis and other Lactobacillus sp.isolated from traditional Pico cheese. Int. J. Dairy Technol. 2018, 70, 1–6. [Google Scholar] [CrossRef]
- Saavedra, L.; Taranto, M.P.; Sesma, F.; de Valdez, G.M. Homemade traditional cheeses for the isolation of probiotic Enterococcus faecium strains. Int. J. Food Microbiol. 2003, 241–245. [Google Scholar] [CrossRef]
- Nascimento, L.C.S.; Casarotti, S.N.; Todorov, S.D.; Penna, A.L.B. Probiotic potential and safety of enterococci strains. Ann. Microbiol. 2019, 69, 241–252. [Google Scholar] [CrossRef]
- Hermanns, G.; Funck, G.D.; Schmidt, J.T.; Pereira, J.Q.; Brandelli, A.; Richards, N.S.P.S. Evaluation of probiotic characteristics of lactic acid bacteria isolated from artisan cheese. J. Food Saf. 2014, 34, 380–387. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; Huycke, M.M. Risks associated with enterococci as probiotics. Food Res. Int. 2020, 129, 108788. [Google Scholar] [CrossRef]
- Homayouni, A.; Ansari, F.; Azizi, A.; Pourjafar, H.; Madadi, M. Cheese as a potential food carrier to deliver probiotic microorganisms into the human gut: A review. Curr. Nutr. Food Sci. 2018, 14, 1–13. [Google Scholar] [CrossRef]
- Castro, J.M.; Tornadijo, M.E.; Fresno, J.M.; Sandoval, H. Biocheese: A food probiotic carrier. BioMed Res. Int. 2015, 2015, 723056. [Google Scholar] [CrossRef] [PubMed]
- Kayser, F.H. Safety aspects of enterococci from the medical point of view. Int. J. Food Microbiol. 2003, 88, 255–262. [Google Scholar] [CrossRef]
- Mundy, L.M.; Sahm, D.F.; Gilmore, M. Relationships between enterococcal virulence and antimicrobial resistance. Clin. Microbiol. Rev. 2000, 13, 513–522. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.; Pardesi, K.R. Emerging strategies to combat ESKAPE pathogens in the era of antimicrobial resistance: A review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Łaniewska-Trokenheim, Ł. Virulence factors of Enterococcus species presented in foods. LWT Food Sci. Technol. 2017, 75, 670–676. [Google Scholar] [CrossRef]
- Lebreton, F.; Riboulet-Bisson, E.; Serror, P.; Sanguinetti, M.; Posteraro, B.; Torelli, R.; Hartke, A.; Auffray, Y.; Giard, J.-C. Ace, which encodes and adhesin in Enterococcus faecalis, is regulated by Ers and is involved in virulence. Infect. Immun. 2009, 77, 2832–2839. [Google Scholar] [CrossRef] [Green Version]
- Vanek, N.N.; Simon, S.I.; Jacques-Palaz, K.; Mariscalco, M.M.; Dunny, G.M.; Rakita, R.M. Enterococcus faecalis aggregation substance promotes opsonin-independent binding to human neutrophils via a complement receptor type 3-mediated mechanism. FEMS Immunol. Med. Microbiol. 1999, 26, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Kayaoglu, G.; Østarvik, D. Virulence factors of Enterococcus faecalis: Relationship to endodontic disease. Crit. Rev. Oral Biol. Med. 2004, 15, 308–320. [Google Scholar] [CrossRef] [Green Version]
- Süßmuth, S.D.; Muscholl-Silberhorn, A.; Wirth, R.; Susa, M.; Marre, R.; Rozdzinski, E. Aggregation substance promotes adherence, phagocytosis, and intracellular survival of Enterococcus faecalis within human macrophages and suppresses respiratory burst. Infect. Immun. 2000, 68, 4900–4906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duprè, I.; Zanetti, S.; Schito, A.M.; Fadda, G.; Sechi, L.A. Incidence of virulence determinants in clinical Enterococcus faecium and Enterococcus faecalis isolates collected in Sardinia (Italy). J. Med. Microbiol. 2003, 52, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Elsner, H.-A.; Sobottka, I.; Mack, D.; Claussen, M.; Laufs, R.; Wirth, R. Virulence factors of Enterococcus faecalis and Enterococcus faecium blood culture isolates. Eur. J. Clin. Microbiol. Infect. Dis. 2000, 19, 39–42. [Google Scholar] [CrossRef] [PubMed]
- Coque, T.M.; Patterson, J.E.; Steckelberg, J.M.; Murray, B. Incidence of hemolysin, gelatinase, and aggregation substance among enterococci isolated from patients with endocarditis and other infections and from feces of hospitalized and community-based persons. Infect. Immun. 2004, 72, 2445–2448. [Google Scholar] [CrossRef]
- Kim, S.-H.; Kim, D.-H.; Lim, H.-W.; Seo, K.-H. High prevalence of non-faecalis and non-faecium Enterococcus spp. in farmstead cheesehouse and their applicability as hygiene indicators. LWT Food Sci. Technol. 2020, 126, 109271. [Google Scholar] [CrossRef]
- Nallapareddy, S.R.; Singh, K.V.; Duh, R.-W.; Weinstock, G.; Murray, B.E. Diversity of ace, a gene encoding a microbial surface component recognizing adhesive matrix molecules from different strains of Enterococcus faecalis and evidence for production of Ace during human infections. Infect. Immun. 2000, 68, 5210–5217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, C.M.A.P.; Stiles, M.E.; Schleifer, K.H.; Holzapfel, W.H. Enterococci in foods, a conundrum for food safety. Int. J. Food Microbiol. 2003, 88, 105–122. [Google Scholar] [CrossRef]
- Eaton, T.J.; Gasson, M.J. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef] [Green Version]
- Toledo-Arana, A.; Valle, J.; Solano, C.; Arrizubieta, M.J.; Cucarella, C.; Lamata, M.; Amorena, B.; Leiva, J.; Penadés, J.R.; Lasa, I. The enterococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Appl. Environ. Microbiol. 2001, 67, 4538–4545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fux, C.A.; Costerton, J.W.; Stewart, P.S.; Stoodley, P. Survival strategies of infectious biofilms. Trends Microbiol. 2005, 13, 34–40. [Google Scholar] [CrossRef]
- Leavis, H.; Top, J.; Shankar, N.; Borgen, K.; Bonten, M.; van Embden, J.; Willems, R.J. A novel putative pathogenicity Island linked to esp virulence gene of Enterococcus faecium and associated with epidemicity. J. Bacteriol. 2004, 186, 672–682. [Google Scholar] [CrossRef] [Green Version]
- Ochoa, S.A.; Escalona, G.; Cruz-Cordova, A.; D’Ávila, L.B.; Saldana, Z.; Cázares-Domímguez, V.; Eslava, C.A.; López-Martínez, B.; Hernández-Castro, R.; Aquino-Jarquin, G.; et al. Molecular analysis and distribution of multidrugresistant Enterococcus faecium isolates belonging to clonal complex 17 in a tertiary care center in Mexico City. BMC Microbiol. 2013, 13, 291. [Google Scholar] [CrossRef] [Green Version]
- Oancea, C.; Klare, I.; Witte, W.; Werner, G. Conjugative transfer of the virulence gene, esp, among isolates of Enterococcus faecium and Enterococcus faecalis. J. Antimicrob. Chemother. 2004, 54, 232–235. [Google Scholar] [CrossRef] [Green Version]
- Ogier, J.-C.; Serror, P. Safety assessment of dairy microorganisms: The Enterococcus genus. Int. J. Food Microbiol. 2008, 126, 291–301. [Google Scholar] [CrossRef]
- Vankerckhoven, V.; Van Autgaerden, T.; Vael, C.; Lammens, C.; Chapelle, S.; Rossi, R.; Jabes, D.; Goossens, H. Development of a multiplex PCR for the detection of asa1, gelE, cylA, esp, and hyl genes in enterococci and survey for virulence determinants among European hospital isolates of Enterococcus faecium. J. Clin. Microbiol. 2004, 42, 4473–4479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahansepas, A.; Sharifi, Y.; Aghazadeh, M.; Rezaee, M.A. Comparative analysis of Enterococcus faecalis and Enterococcus faecium strains isolated from clinical samples and traditional cheese types in the Northwest of Iran: Antimicrobial susceptibility and virulence traits. Archiv. Microbiol. 2020, 202, 756–772. [Google Scholar] [CrossRef] [PubMed]
- Cariolato, D.; Andrighetto, C.; Lombardi, A. Occurrence of virulence factors and antibiotic resistances in Enterococcus faecalis and Enterococcus faecium collected from dairy and human samples in North Italy. Food Control 2008, 19, 886–892. [Google Scholar] [CrossRef]
- Gaglio, R.; Couto, N.; Marques, C.; Lopes, M.F.S.; Moschetti, G.; Pomba, C.; Settani, L. Evaluation of antimicrobial resistance and virulence of enterococci from equipment surfaces, raw materials, and traditional cheeses. Int. J. Food Microbiol. 2016, 236, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Hammad, A.M.; Hassan, H.A.; Shimamoto, T. Prevalence, antibiotic resistance and virulence of Enterococcus spp. in Egyptian fresh raw milk cheese. Food Control 2015, 50, 815–820. [Google Scholar] [CrossRef]
- İspirli, H.; Demirbaş, F.; Dertli, E. Characterization of functional properties of Enterococcus spp. Isolated from Turkish white cheese. LWT Food Sci. Technol. 2017, 75, 358–365. [Google Scholar] [CrossRef]
- Fuka, M.M.; Maksimovic, A.Z.; Tanuwidjaja, I.; Hulak, N.; Schloter, M. Characterization of enterococcal community isolated from an artisan Istrian raw milk cheese: Biotechnological and safety aspects. Food Technol. Biotechnol. 2017, 55, 368–380. [Google Scholar] [CrossRef]
- Cebrián, R.; Baños, A.; Valdivia, E.; Pérez-Pulido, R.; Martínez-Bueno, M.; Maqueda, M. Characterization of functional, safety, and probiotic properties of Enterococcus faecalis UGRA10, a new AS-48-producer strain. Food Microbiol. 2012, 30, 59–67. [Google Scholar] [CrossRef]
- Templer, S.P.; Baumgartner, A. Enterococci from Appenzeller and Schabziger raw milk cheese: Antibiotic resistance, virulence factors, and persistence of particular strains in the products. J. Food Prot. 2007, 70, 450–455. [Google Scholar] [CrossRef]
- Yuksel, F.N.; Akcelik, N.; Akcelik, M. Incidence of antibiotic resistance and virulence determinants in Enterococcus faecium and Enterococcus faecalis strains isolated from traditional cheeses in Turkey. Mol. Gen. Microbiol. Virol. 2015, 30, 206–215. [Google Scholar] [CrossRef]
- Popović, N.; Dinić, M.; Tolinački, M.; Mihaljović, S.; Terzić-Vidojević, A.; Bojić, S.; Djokić, J.; Golić, N.; Veljović, K. New insight into biofilm formation ability, the presence of virulence genes and probiotic potential of Enterococcus sp. dairy isolates. Front. Microbiol. 2018, 9, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chajęcka-Wierzchowska, W.; Zadernowska, A.; Zarzecka, U.; Zakrzewski, A.; Gajewska, J. Enterococci from ready-to-eat food-horizontal gene transfer of antibiotic resistance genes and genotypic characterization by PCR melting profile. J. Sci. Food Agric. 2019, 99, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Fiore, E.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of enterococci. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Pillar, C.M.; Gilmore, M.S. Enterococcal virulence–pathogenicity island of E. faecalis. Front. BioSci. 2004, 9, 2335–2346. [Google Scholar] [CrossRef] [Green Version]
- Ramos, S.; Silva, V.; Dapkevicius, M.L.N.E.; Igrejas, G.; Poeta, P. Enterococci, from harmless bacteria to a pathogen. Microorganisms 2020, 8, 1118. [Google Scholar] [CrossRef]
- Silvetti, T.; Capra, E.; Morandi, S.; Cremonesi, P.; Decimo, M.; Gavazzi, F.; Giannico, R.; De Noni, I.; Brasca, M. Microbial population profile during ripening of Protected Designation of Origin (PDO) Silter cheese, produced with and without autochthonous starter culture. LWT Food Sci. Technol. 2017, 84, 821–831. [Google Scholar] [CrossRef]
- Barlow, S.; Chesson, A.; Collins, J.D.; Dybing, E.; Flynn, A.; Fruijtier-Pölloth, C.; Hardy, A.; Knapp, A.; Kuiper, H.; Le Neindre, P.; et al. Opinion of the Scientific Committee on a request from EFSA on the introduction of a Qualified Presumption of Safety (QPS) approach for assessment of selected microorganisms referred to EFSA. EFSA J. 2007, 587, 1–16. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef]
- Palmer, K.L.; Kos, V.N.; Gilmore, M.S. Horizontal gene transfer and the genomics of enterococcal antibiotic resistance. Curr. Opin. Microbiol. 2010, 13, 632–639. [Google Scholar] [CrossRef] [Green Version]
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in enterococcus. Virulence 2012, 3, 421–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Jeannot, K.; Xiao, Y.; Knapp, C.W. Horizontal gene transfer mediated bacterial antibiotic resistance. Front. Microbiol. 2019, 27, 933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeder, M.; Brooks, B.D.; Brooks, A.E. The complex relationship between virulence and antibiotic resistance. Genes 2017, 8, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clewell, D.; Weaver, K.; Dunny, G.; Coque, T.M.; Francia, M.V.; Hayes, F. Extrachromosomal and Mobile Elements in Enterococci: Transmission, Maintenance, and Epidemiology. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection [Internet]; Gilmore, M., Clewell, D.D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014; pp. 309–420. [Google Scholar]
- Palmer, K.L.; Gilmore, M.S. Multidrug-resistant enterococci lack CRISPR-cas. mBio 2010, 12, e00227-10. [Google Scholar] [CrossRef] [Green Version]
- Bonacina, J.; Suárez, N.; Hormigo, R.; Fadda, S.; Lechner, M.; Saavedra, L. A genomic view of food-related and probiotic Enterococcus strains. DNA Res. 2017, 24, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.O.; Baptiste, K.E. Vancomycin-resistant enterococci: A review of antimicrobial resistance mechanisms and perspectives of human and animal health. Microb. Drug Resist. 2018, 24, 590–606. [Google Scholar] [CrossRef] [Green Version]
- Agudelo Higuita, N.I.; Huycke, M.M. Enterococcal Disease, Epidemiology, and Implications for Treatment. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014; pp. 65–100. [Google Scholar]
- Maietti, L.; Bonvini, B.; Huys, G.; Giraffa, G. Incidence of antibiotic resistance and virulence determinants among Enterococcus italicus isolates from dairy products. Syst. Appl. Microbiol. 2007, 30, 509–517. [Google Scholar] [CrossRef]
- Mannu, L.; Paba, A.; Daga, E.; Comunian, R.; Zanetti, S.; Duprè, I.; Sechi, L.A. Comparison of the incidence of virulence determinants and antibiotic resistance between Enterococcus faecium strains of dairy, animal and clinical origin. Int. J. Food Microbiol. 2003, 88, 291–304. [Google Scholar] [CrossRef]
- WHO. World Health Organization Model List of Essential Medicines, 21st ed.; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Castaño-Arriba, A.; González-Machado, C.; Igrejas, G.; Poeta, P.; Alonso-Calleja, C.; Capita, R. Antibiotic resistance and biofilm-forming ability in enterococcal isolates from red meat and poultry preparations. Pathogens 2020, 9, 1021. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
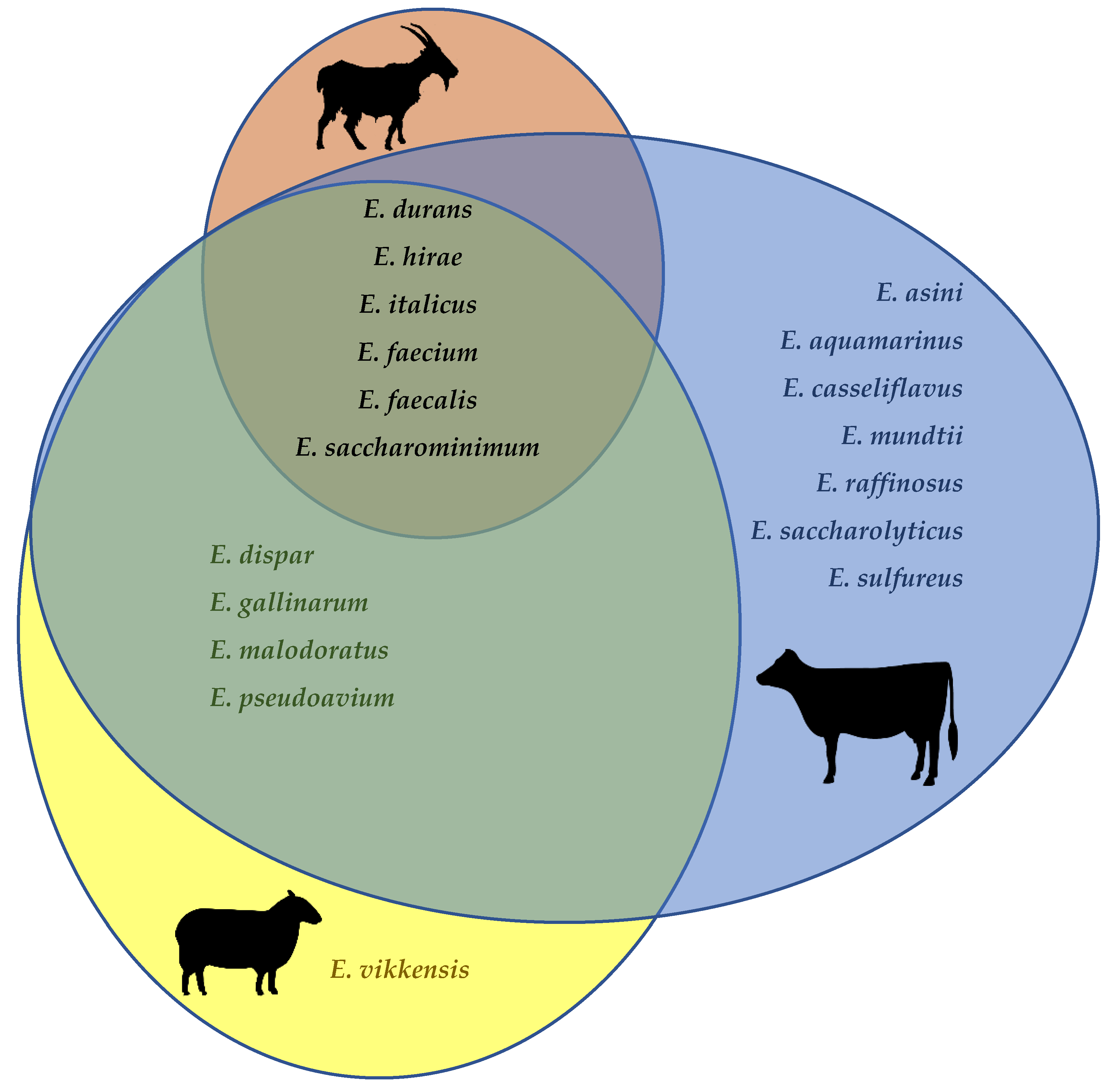
| Cheese (Type *) | Country | Milk Species/Treatment | Maturation Time | Enterococcal Species | Ref. |
|---|---|---|---|---|---|
| Alberquilla (H) | Spain | Ewe/goat | - | E. devriesei, E. faecium | [117] |
| Arzúa-Ulloa (S) | Spain | Cow (raw, pasteurized) | 6 mo | E. durans | [122] |
| Bitto (H) | Italy | Cow (raw) | >70 d | E. durans, E. faecalis, E. faecium, E. lactis | [73,123] |
| Blue-veined cheese (MR) | UK | Cow (raw) | 3 mo | E. durans, E. faecalis | [120] |
| Bryndza (S) | Slovakia | Ewe (raw) | ≤14 d | E. casseliflavus, E. durans, E. faecalis, E. faecium, E. mundtii, E. pallens | [124,125,126] |
| Cabrales (MR) | Spain | Cow/ewe/goat (raw) | 2–5 mo | E. durans, E. faecalis | [122] |
| Caciocavallo Palermit. (PF) | Italy | Cow (raw) | 4 mo | E. casseliflavus, E durans, E. faecalis, E. gallinarum | [75,127] |
| Calenzana (S) | Italy | Ewe/goat | 4–8 mo | E. faecium, E. durans, E. hirae | [76] |
| Canestrato Pugliese (H) | Italy | Ewe (raw) | 4 mo | E. faecalis, E. faecium | [128] |
| Casera Valtellina (H) | Italy | Cow (raw) | 70 d | E. faecalis, E. faecium, E. gilvus | [129,130] |
| Casín (SH/H) | Spain | Cow (raw) | 38 d | E. faecium | [91] |
| Cazisolu (PF) | Italy | Cow (raw) | 1 mo | E. durans, E. faecium, E. italicus, E. lactis | [131] |
| Cueva de la Magahá (H) | Spain | Goat (raw) | 8 mo | E. devriesei, E. faecalis, E. faecium, E. hirae, E. malodoratus | [92] |
| Danbo (SS) | Denmark | Cow | 18 wk | E. faecalis, E. faecium | [132] |
| Feta (S; B) | Greece | Ewe/goat | 2 mo | E. canis, E. faecalis, E. faecium | [133] |
| Fiore Sardo (H) | Italy | Ewe (raw) | 9 mo | E. durans, E. faecalis, E. faecium | [77] |
| Fontina (SS) | Italy | Cow (raw) | ≤9 mo | E. durans, E. faecalis, E. faecium, E. hirae | [48,134,135] |
| Fossa (H) | Italy | Cow/ewe | 1–3 mo | E. durans, E. faecium, E. faecalis | [136] |
| Fresh cheese (F) | Croatia | Cow (pasteurized) | none | E. durans, E. faecium, E. hirae, E. ratti, E. villorum | [137] |
| Galotyri (F) | Greece | Ewe/goat | none | E. durans, E. faecalis, E. faecium | [98] |
| Genestoso (S) | Spain | Cow (raw) | 20–30 d | E. faecalis | [138] |
| Gorgonzolla (MR) | Italy | Cow | 3–4 mo | E. faecalis, E. faecium | [129] |
| Gouda-type (SH/H) | Belgium | Cow (raw) | 2–4 mo | E. casseliflavus/malodoratus group, E. faecalis, E faecium/durans group | [121,139] |
| Graviera (H) | Greece | Ewe/goat (thermized) | 3 mo | E. durans, E. faecalis, E. faecium, E. hirae | [99,100] |
| Hervé (S) | Belgium | Cow (raw) | 63 d | E. casseliflavus, E. faecalis, E. faecium | [140] |
| Idiazábal-type (S; Smk) | Spain | Ewe (raw) | 70–72 d | E. durans, E. faecalis, E. faecium, E. casseliflavus, E. hirae, E. gallinarum | [37] |
| Istrian (H) | Croatia | Ewe (raw) | 3 mo | E. faecalis, E. faecium, E. italicus, E. saccharominimus, E. sulfureus | [141] |
| Kalathaki Lemnou (S) | Greece | Ewe/goat (raw) | 3 mo | E. durans, E. faecalis, E. faecium, E. gilvus | [101] |
| Livanjski (SH) | Bos.-Herz. | Cow/ewe (raw) | 1–2 mo | E. durans, E. faecalis, E. faecium, E. gilvus, E. hirae, E. lactis, E. malodoratus | [95] |
| Manchego (SH/H) | Spain | Ewe (raw, pasteurized) | 2–24 mo | E. avium, E. faecalis, E. faecium, E. hirae | [142] |
| Maroilles (S) | France | Cow (raw) | ≤4 mo | E. devriesei | [143] |
| Melichloro (H) | Greece | Ewe/goat (raw) | 4–6 d | E. avium, E. durans, E. faecium, E. pseudoavium | [101] |
| Montasio (SH) | Italy | Cow (raw) | 2–5 mo | E. durans, E. faecalis, E. faecium, E. gallinarum | [78,79] |
| Mozzarella (F; PF) | Italy | Cow (raw) | fresh | E. faecalis, E. sulfureus | [144] |
| Nostrano (S) | Italy | Cow (raw) | 8 mo | E. faecalis | [80] |
| Nostrano di Primiero (S) | Italy | Cow (raw) | 2 mo | E. faecalis, E. faecium | [81] |
| Oscypek (SH; Smk) | Poland | Ewe (raw) | 5–18 d | E. durans, E. italicus | [145] |
| Pecorino (central It.) (H) | Italy | Ewe (raw) | 1–12 mo | E. durans, E. faecalis, E. faecium | [146] |
| Pecorino Abruzzese (H) | Italy | Ewe (raw) | 3 mo | E. durans, E. faecalis, E. faecium | [102] |
| Pecorino Crotonese (H) | Italy | Ewe (r., therm., past.) | 4 mo | E. faecalis | [147] |
| Pecorino di Tramonti (H) | Italy | Ewe (raw) | 1–3 mo | E. durans, E. faecalis, E. faecium | [105] |
| Pecorino Siciliano (H) | Italy | Ewe (raw) | 0.5–6 mo | E. durans, E. faecalis, E. faecium, E. hirae | [82,106,148] |
| Picante (SH) | Portugal | Ewe/goat (raw) | 4–6 mo | E. durans, E. faecalis, E. faecium, | [149,150,151] |
| Pico (SS) | Portugal | Cow (raw) | 21 d | E. faecalis, E. italicus, E. pseudoavium | [152,153,154] |
| Provolone del Monaco (SH) | Italy | Cow (raw) | 6–12 mo | E. durans, E. faecalis, E. faecium | [83] |
| Quesailla Arochena (NS) | Spain | Goat (raw) | 4 mo | E. devriesei, E. faecalis, E. malodoratus | [118] |
| Ragusano (H; PF) | Italy | Ewe (raw) | 6 mo | E. durans, E. faecalis, E. faecium, E. hirae | [148] |
| Raschera (H) | Italy | Cow (raw) | 1–6 mo | E. casseliflavus, E. faecalis, E. faecium | [84] |
| Raw goat milk (SH) | Spain | Goat (raw) | 2 mo | E. casseliflavus, E. durans, E. faecalis, E. faecium, E. gallinarum, E. hirae, E. italicus, E. lactis | [93] |
| S. Jorge (SH/H) | Portugal | Cow (raw) | 6–12 mo | E. faecalis, E. faecium | [155] |
| Saint-Nectaire (S) | France | Cow (raw) | 28 d | E. faecalis | [156] |
| Salers (SH) | France | Cow (raw) | 5 mo | E. faecalis, E. faecium | [157,158,159] |
| San Simón da Costa (SH;Smk) | Spain | Cow (raw) | 45–60 d | E. faecalis, E. sulfureus | [160] |
| Scimudin (S) | Italy | Cow (raw) | >10 d | E. faecalis, E. faecium | [129] |
| Semicotto caprine (H) | Italy | Goat (raw) | 2 mo | E. durans, E. faecalis, E. faecium, E. gallinarum, E. hirae | [85] |
| Serbian artisanal (NS) | Serbia | Cow (raw) | - | E. durans, E. faecalis | [161] |
| Serpa (SS) | Portugal | Ewe (raw) | 30 d | E. faecalis, E. faecium, E. hirae | [111] |
| Serra (S) | Portugal | Ewe (raw) | 2–6 mo | E. faecium | [162,163] |
| Sokobanja (NS) | Serbia | Cow (raw) | 3d | E. faecalis, E. faecium | [116] |
| Stilton (MR) | UK | Cow (pasteurized) | 9–12 wk | E. faecalis | [164] |
| Taleggio (SS) | Italy | Cow (raw, past.) | 6–10 wk | E. faecalis | [129] |
| Terrincho (SS) | Portugal | Ewe (raw) | 1 mo | E. casseliflavus, E. durans, E. faecalis, E. faecium, E. gallinarum | [165] |
| Tetilla (S) | Spain | Cow (raw) | 1 mo | E. durans, E. faecalis | [122] |
| Tolminc (H) | Slovenia | Cow (raw) | 2 mo | E. faecalis | [166] |
| Toma Piemontese (SS) | Italy | Cow (pasteurized) | 20–45 d | E. durans, E. faecalis, E. faecium | [167] |
| Torta Arochena (NS) | Spain | Goat (raw) | 4 mo | E. avium, E. devriesei, E. faecalis, E. malodoratus | [118] |
| Valsesia (SH) | Italy | Goat (raw) | 1–2 mo | E. casseliflavus, E. durans, E. faecalis, E. faecium, E. gallinarum, E. gilvus | [86] |
| Vlasina (S; B) | Serbia | Goat (raw) | 2 mo | E. durans, E. faecalis, E. faecium | [168] |
| White pickled (S; B) | Serbia, Croatia | Cow (raw) | 1–10 d | E. durans, E. faecalis, E. faecium | [169] |
| Zlatar (SH; B) | Serbia | Cow (raw) | 2 mo | E. faecalis, E. faecium | [170] |
| Enterococcal Species | Genetic Determinants of Resistance | Phenotypic Resistance to 1 | Reference(s) |
|---|---|---|---|
| E. durans | Not tested | ERY; CIP; PEN; TET; CHL | [102] |
| E. faecalis | tetK,L,M,S; aadE; aph3′; cat; erm; vanA | GENT; STR; KAN; NEO; CFT; CFO; PEN; OXA; TET; MYN; ERY; VAN; CHL; Q-D; SxT | [44,102,152,166,179,273] |
| E. faecium | tetM | ERY; CIP; LEV; NOR; NIT | [179,273,298] |
| E. gilvus | None detected | None detected | [179] |
| E. italicus | tetK,S | RIF; TET; TRI | [193,297] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dapkevicius, M.d.L.E.; Sgardioli, B.; Câmara, S.P.A.; Poeta, P.; Malcata, F.X. Current Trends of Enterococci in Dairy Products: A Comprehensive Review of Their Multiple Roles. Foods 2021, 10, 821. https://doi.org/10.3390/foods10040821
Dapkevicius MdLE, Sgardioli B, Câmara SPA, Poeta P, Malcata FX. Current Trends of Enterococci in Dairy Products: A Comprehensive Review of Their Multiple Roles. Foods. 2021; 10(4):821. https://doi.org/10.3390/foods10040821
Chicago/Turabian StyleDapkevicius, Maria de Lurdes Enes, Bruna Sgardioli, Sandra P. A. Câmara, Patrícia Poeta, and Francisco Xavier Malcata. 2021. "Current Trends of Enterococci in Dairy Products: A Comprehensive Review of Their Multiple Roles" Foods 10, no. 4: 821. https://doi.org/10.3390/foods10040821
APA StyleDapkevicius, M. d. L. E., Sgardioli, B., Câmara, S. P. A., Poeta, P., & Malcata, F. X. (2021). Current Trends of Enterococci in Dairy Products: A Comprehensive Review of Their Multiple Roles. Foods, 10(4), 821. https://doi.org/10.3390/foods10040821