
Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses?



Abstract
:1. Introduction
2. Materials and Methods
2.1. MICs Determination
2.2. Challenge Tests
2.3. Statistical Analysis
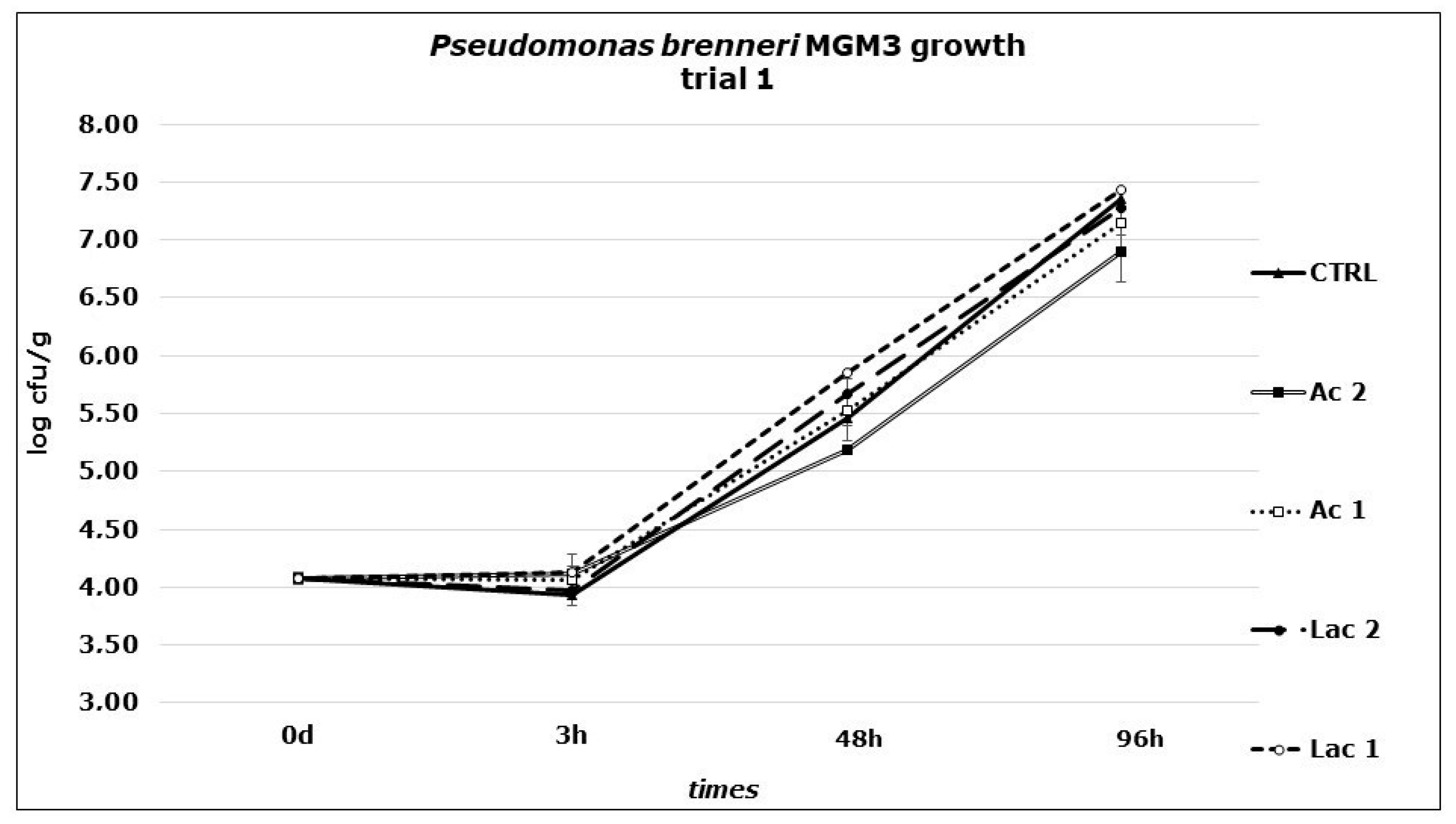
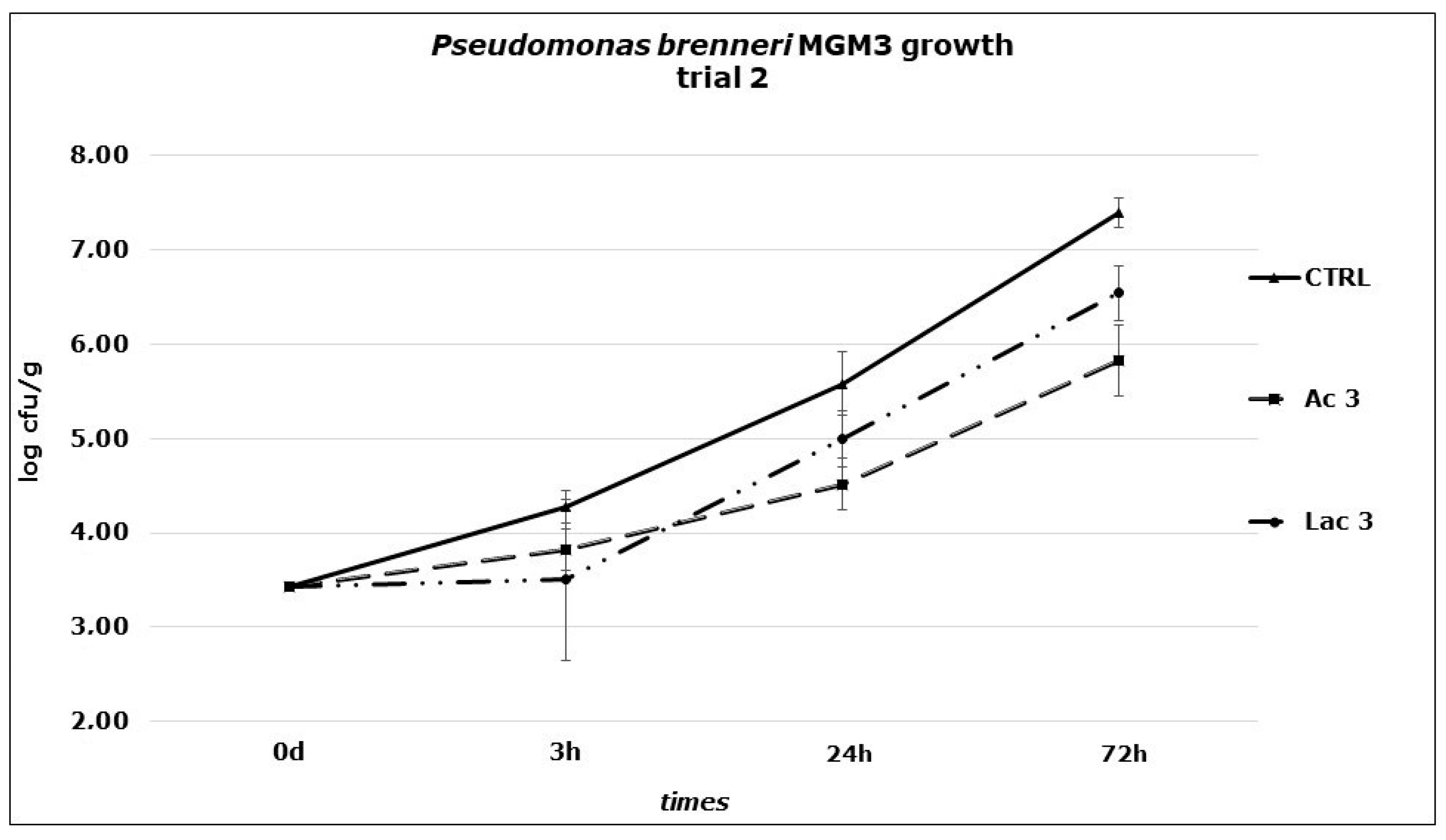
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gustavsson, J.; Cederberg, C.; Sonesson, U.; Van Otterdijk, R.; Meybeck, A. Global Food Losses and Food Waste; Food and Agriculture Organization of the United Nations: Rome, Italy, 2011. [Google Scholar]
- Sperber, W.H.; Doyle, M.P. Compendium of the Microbiological Spoilage of Foods and Beverages; Springer: New York, NY, USA, 2009. [Google Scholar]
- Petruzzi, L.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Chapter 1—Microbial spoilage of foods: Fundamentals. In The Micro-Biological Quality of Food—Foodborne Spoilers; Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Woodhead Publishing: Sawston, Cambridge, UK, 2017; pp. 1–21. [Google Scholar]
- Kraft, A.A. Psychrotrophic Bacteria in Foods: Disease and Spoilage; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Wiedmann, M.; Weilmeier, D.; Dineen, S.S.; Ralyea, R.; Boor, K.J. Molecular and Phenotypic Characterization ofPseudomonas spp. Isolated from Milk. Appl. Environ. Microbiol. 2000, 66, 2085–2095. [Google Scholar] [CrossRef] [Green Version]
- Spanu, C.; Piras, F.; Mocci, A.; Nieddu, G.; De Santis, E.; Scarano, C. Use of Carnobacterium spp protective culture in MAP packed Ricotta fresca cheese to control Pseudomonas spp. Food Microbiol. 2018, 74, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Cantoni, C.; Stella, S.; Ripamonti, B.; Marchese, R. Pseudomonas fluorescens and anomalous colourations of mozzarella cheese. Ind. Aliment. 2001, 40, 33–35. [Google Scholar]
- Cantoni, C.; Stella, S.; Cozzi, M.; Iacumin, L.; Comi, G. Blue colouring in mozzarella cheese. Ind. Aliment. 2003, 42, 840–843. [Google Scholar]
- Carminati, D.; Bonvini, B.; Rossetti, L.; Zago, M.; Tidona, F.; Giraffa, G. Investigation on the presence of blue pigment-producing Pseudomonas strains along a production line of fresh mozzarella cheese. Food Control 2019, 100, 321–328. [Google Scholar] [CrossRef]
- Dogan, B.; Boor, K.J. Genetic Diversity and Spoilage Potentials among Pseudomonas spp. Isolated from Fluid Milk Products and Dairy Processing Plants. Appl. Environ. Microbiol. 2003, 69, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Carrascosa, C.; Millán, R.; Jaber, J.R.; Lupiola, P.; del Rosario-Quintana, C.; Mauricio, C.; Sanjuán, E. Blue pigment in fresh cheese produced by Pseudomonas fluorescens. Food Control 2015, 54, 95–102. [Google Scholar] [CrossRef]
- Scatamburlo, T.; Yamazi, A.; Cavicchioli, V.; Pieri, F.; Nero, L.A.; Moreira, T. Spoilage potential of Pseudomonas species isolated from goat milk. J. Dairy Sci. 2015, 98, 759–764. [Google Scholar] [CrossRef]
- Rossi, C.; Serio, A.; Chaves-López, C.; Anniballi, F.; Auricchio, B.; Goffredo, E.; Cenci-Goga, B.T.; Lista, F.; Fillo, S.; Paparella, A. Biofilm formation, pigment production and motility in Pseudomonas spp. isolated from the dairy industry. Food Control 2018, 86, 241–248. [Google Scholar] [CrossRef]
- Arslan, S.; Eyi, A.; Özdemir, F. Spoilage potentials and antimicrobial resistance of Pseudomonas spp. isolated from cheeses. J. Dairy Sci. 2011, 94, 5851–5856. [Google Scholar] [CrossRef] [Green Version]
- Van Tassell, J.; Martin, N.; Murphy, S.; Wiedmann, M.; Boor, K.; Ivy, R. Evaluation of various selective media for the detection of Pseudomonas species in pasteurized milk. J. Dairy Sci. 2012, 95, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, P.J. Coupling of Phosphorylation to Electron and Hydrogen Transfer by a Chemi-Osmotic type of Mechanism. Nat. Cell Biol. 1961, 191, 144–148. [Google Scholar] [CrossRef]
- European Union. Regulation (EC) No 1333/2008. In Proceedings of the European Parliament and of the Council of 16 December 2008 on Food Additives, Bruxelles, Belgium, 31 December 2008; Volume L 354, pp. 16–33. [Google Scholar]
- Tirloni, E.; Nauta, M.; Vasconi, M.; Di Pietro, V.; Bernardi, C.; Stella, S. Growth of Listeria monocytogenes in ready-to-eat “shrimp cocktail”: Risk assessment and possible preventive interventions. Int. J. Food Microbiol. 2020, 334, 108800. [Google Scholar] [CrossRef] [PubMed]
- Baïda, N.; Yazourh, A.; Singer, E.; Izard, D. Pseudomonas brenneri sp. nov., a new species isolated from natural mineral waters. Res. Microbiol. 2001, 152, 493–502. [Google Scholar] [CrossRef]
- Andreani, N.; Martino, M.; Fasolato, L.; Carraro, L.; Montemurro, F.; Mioni, R.; Bordin, P.; Cardazzo, B. Reprint of ‘Tracking the blue: A MLST approach to characterise the Pseudomonas fluorescens group’. Food Microbiol. 2015, 45, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.R.; Moss, M.O. Food Microbiology, 2nd ed.; Royal Society of Chemistry: London, UK, 2000. [Google Scholar]
- Gunasekera, T.; Dorsch, M.; Slade, M.; Veal, D. Specific detection of Pseudomonas spp. in milk by fluorescence in situ hybridi-zation using ribosomal RNA directed probes. J. Appl. Microbiol. 2003, 94, 936–945. [Google Scholar] [CrossRef] [Green Version]
- Baruzzi, F.; Lagonigro, R.; Quintieri, L.; Morea, M.; Caputo, L. Occurrence of non-lactic acid bacteria populations involved in protein hydrolysisof cold-stored high moisture Mozzarella cheese. Food Microbiol. 2012, 30, 37–44. [Google Scholar] [CrossRef]
- Nogarol, C.; Acutis, P.L.; Bianchi, D.M.; Maurella, C.; Peletto, S.; Gallina, S.; Adriano, D.; Zuccon, F.; Borrello, S.; Caramelli, M.; et al. Molecular Characterization of Pseudomonas fluorescens Isolates Involved in the Italian “Blue Mozzarella” Event. J. Food Prot. 2013, 76, 500–504. [Google Scholar] [CrossRef]
- Del Olmo, A.; Calzada, J.; Nuñez, M. The blue discoloration of fresh cheeses: A worldwide defect associated to specific contamination by Pseudomonas fluorescens. Food Control 2018, 86, 359–366. [Google Scholar] [CrossRef]
- Sinigaglia, M.; Bevilacqua, A.; Corbo, M.R.; Pati, S.; Del Nobile, M.A. Use of active compounds for prolonging the shelf life of mozzarella cheese. Int. Dairy J. 2008, 18, 624–630. [Google Scholar] [CrossRef]
- Cruz-Romero, M.; Murphy, T.; Morris, M.; Cummins, E.; Kerry, J. Antimicrobial activity of chitosan, organic acids and nano-sized solubilisates for potential use in smart antimicrobially-active packaging for potential food applications. Food Control 2013, 34, 393–397. [Google Scholar] [CrossRef]
- Moreira, T.S.M.; de Oliveira, A.; Barlati, T.; da Silva, V.; Rodrigues Dos Santos, A.; Gonçalves, O.H.; da Silva Gonzalez, O.; Droval, A.A.; Leimann, F.V. Hydrogels based on gelatin: Effect of lactic and acetic acids on microstructural modifications, water absorption mechanisms and antibacterial activity. LWT 2019, 103, 69–77. [Google Scholar] [CrossRef]
- Nakai, S.A.; Siebert, K.J. Organic acid inhibition models for Listeria innocua, Listeria ivanovii, Pseudomonas aeruginosa and Oenococcus oeni. Food Microbiol. 2004, 21, 67–72. [Google Scholar] [CrossRef]
- Bushell, F.M.L.; Tonner, P.D.; Jabbari, S.; Schmid, A.K.; Lund, P.A. Synergistic Impacts of Organic Acids and pH on Growth of Pseudomonas aeruginosa: A Comparison of Parametric and Bayesian Non-parametric Methods to Model Growth. Front. Microbiol. 2019, 9, 3196. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.; McCoy, C.P.; Irwin, N.J. Synergistic activity of weak organic acids against uropathogens. J. Hosp. Infect. 2021. [Google Scholar] [CrossRef]
- Leriche, F.; Bordessoules, A.; Fayolle, K.; Karoui, R.; Laval, K.; Leblanc, L.; Dufour, E. Alteration of raw-milk cheese by Pseudomonas spp.: Monitoring the sources of contamination using fluorescence spectroscopy and metabolic profiling. J. Microbiol. Methods 2004, 59, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Lucera, A.; Mastromatteo, M.; Conte, A.; Zambrini, A.V.; Faccia, M.; Del Nobile, M.A. Effect of active coating on microbiological and sensory properties of fresh mozzarella cheese. Food Packag. Shelf 2014, 1, 25–29. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Acetic Acid | t0 | t24 | t48 | t72 |
|---|---|---|---|---|
| Average ± Std.Dev | Average ± Std.Dev | Average ± Std.Dev | Average ± Std.Dev | |
| 49.96 mM | −0.02 ± 0.03 | 0.01 ± 0.04 | 0.01 ± 0.04 | 0.01 ± 0.04 |
| 24.98 mM | −0.05 ± 0.01 | −0.03 ± 0.42 | 0.28 ± 0.42 | 0.57 ± 0.16 |
| 12.49 mM | −0.04 ± 0.01 | 0.12 ± 0.01 | 1.54 ± 0.01 | 1.76 ± 0.13 |
| 5.00 mM | −0.12 ± 0.01 | 0.20 ± 0.18 | 1.55 ± 0.18 | 1.55 ± 0.14 |
| 2.50 mM | −0.09 ± 0.01 | 0.28 ± 0.02 | 1.47 ± 0.02 | 1.61 ± 0.03 |
| CTRL | 0.01 ± 0.09 | 0.35 ± 0.01 | 1.65 ± 0.01 | 1.72 ± 0.06 |
| Lactic Acid | t0 | t24 | t48 | t72 |
| Average ± std.dev | Average ± std.dev | Average ± std.dev | Average ± std.dev | |
| 44.40 mM | −0.06 ± 0.03 | −0.04 ± 0.02 | −0.05 ± 0.02 | 0.01 ± 0.01 |
| 22.20 mM) | −0.08 ± 0.01 | −0.03 ± 0.04 | 1.42 ± 0.01 | 1.79 ± 0.06 |
| 11.10 mM | −0.08 ± 0.05 | 0.24 ± 0.03 | 1.60 ± 0.04 | 1.65 ± 0.04 |
| 5.55 mM | −0.22 ± 0.26 | 0.35 ± 0.01 | 1.58 ± 0.15 | 1.56 ± 0.12 |
| 2.78 mM | 0.24 ± 0.39 | 0.35 ± 0.07 | 1.57 ± 0.04 | 1.80 ± 0.09 |
| CTRL | 0.01 ± 0.09 | 0.35 ± 0.05 | 1.65 ± 0.01 | 1.72 ± 0.06 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tirloni, E.; Bernardi, C.; Stella, S. Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses? Foods 2021, 10, 891. https://doi.org/10.3390/foods10040891
Tirloni E, Bernardi C, Stella S. Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses? Foods. 2021; 10(4):891. https://doi.org/10.3390/foods10040891
Chicago/Turabian StyleTirloni, Erica, Cristian Bernardi, and Simone Stella. 2021. "Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses?" Foods 10, no. 4: 891. https://doi.org/10.3390/foods10040891
APA StyleTirloni, E., Bernardi, C., & Stella, S. (2021). Pseudomonas spp.: Are Food Grade Organic Acids Efficient against These Spoilage Microorganisms in Fresh Cheeses? Foods, 10(4), 891. https://doi.org/10.3390/foods10040891