
Non-Specific Binding and Cross-Reaction of ELISA: A Case Study of Porcine Hemoglobin Detection



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. Indirect sELISA
2.4. Western Blot
2.5. Statistical Analysis
3. Results and Discussion
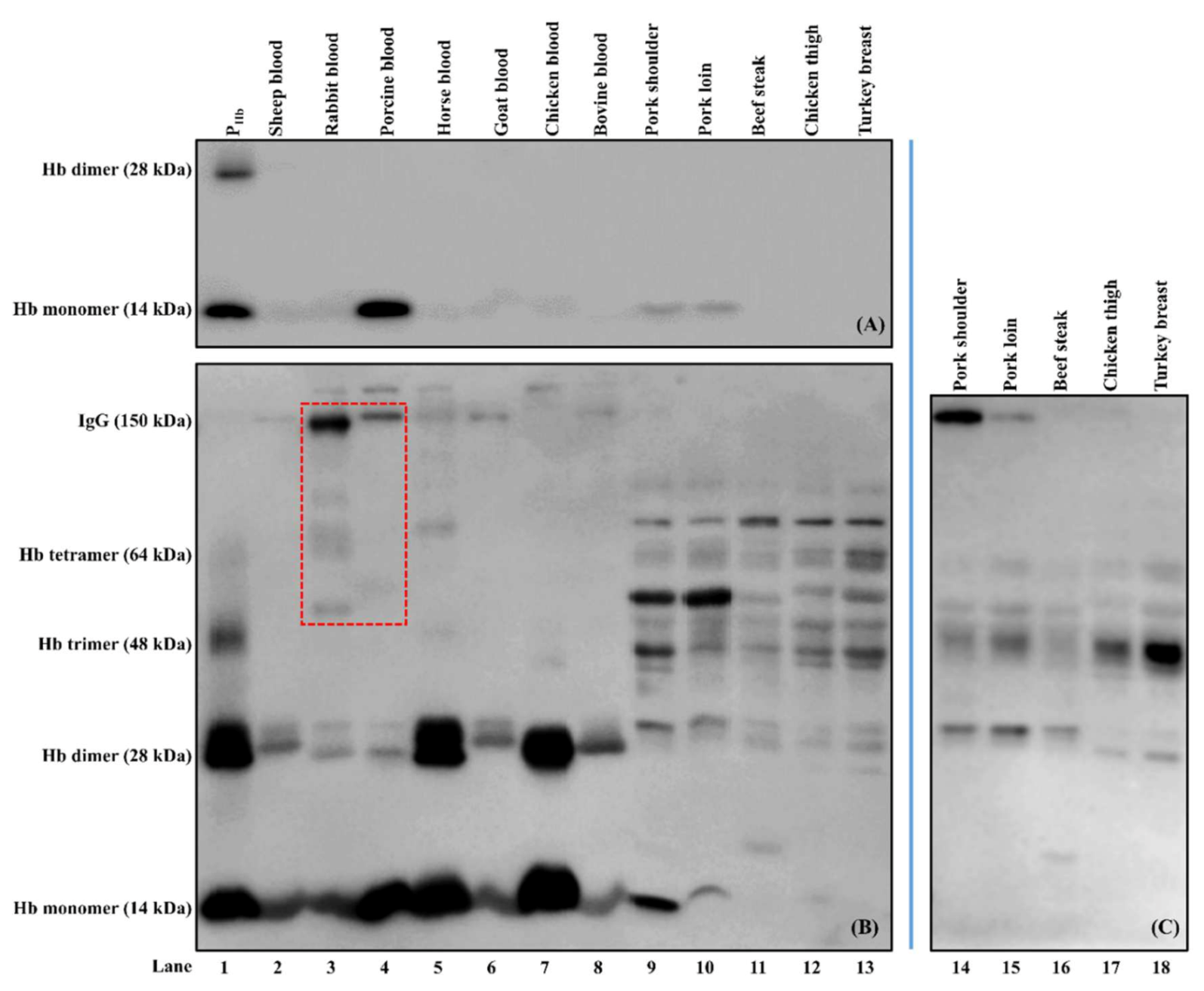
3.1. Antibody Characterization
3.2. Non-Specific Binding (NSB)
3.2.1. Effect of Tween-20
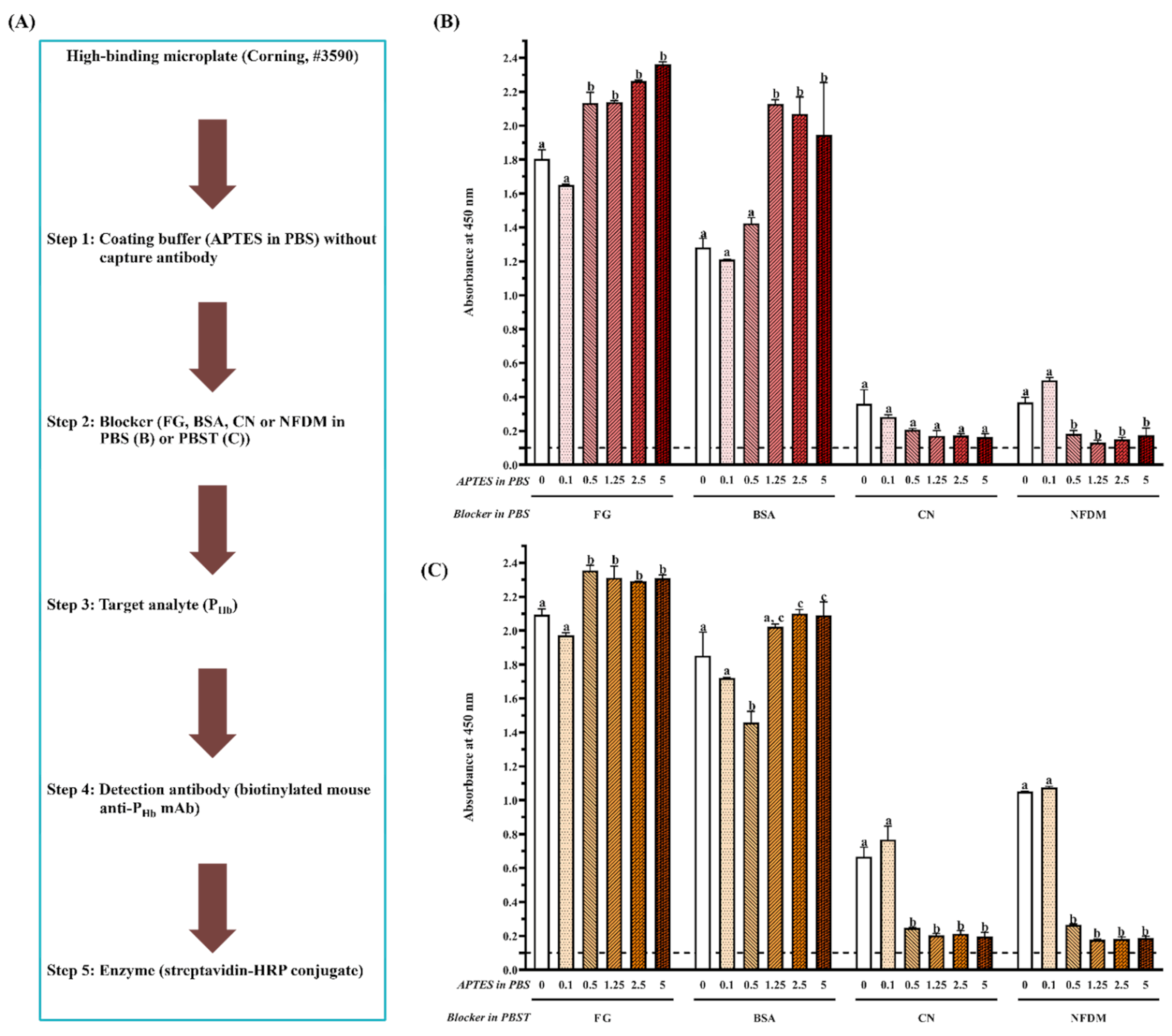
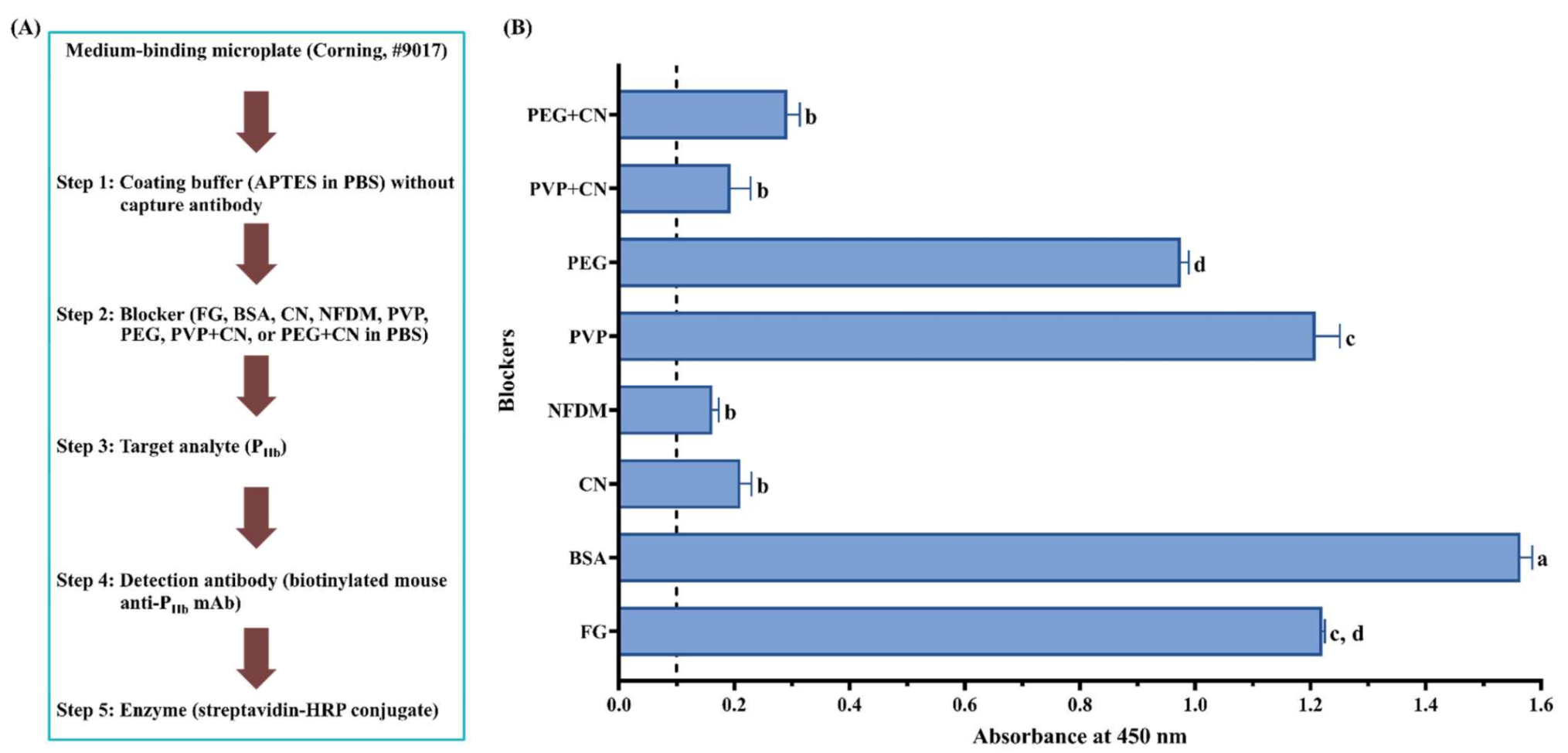
3.2.2. Effect of Protein and Non-Protein Blockers
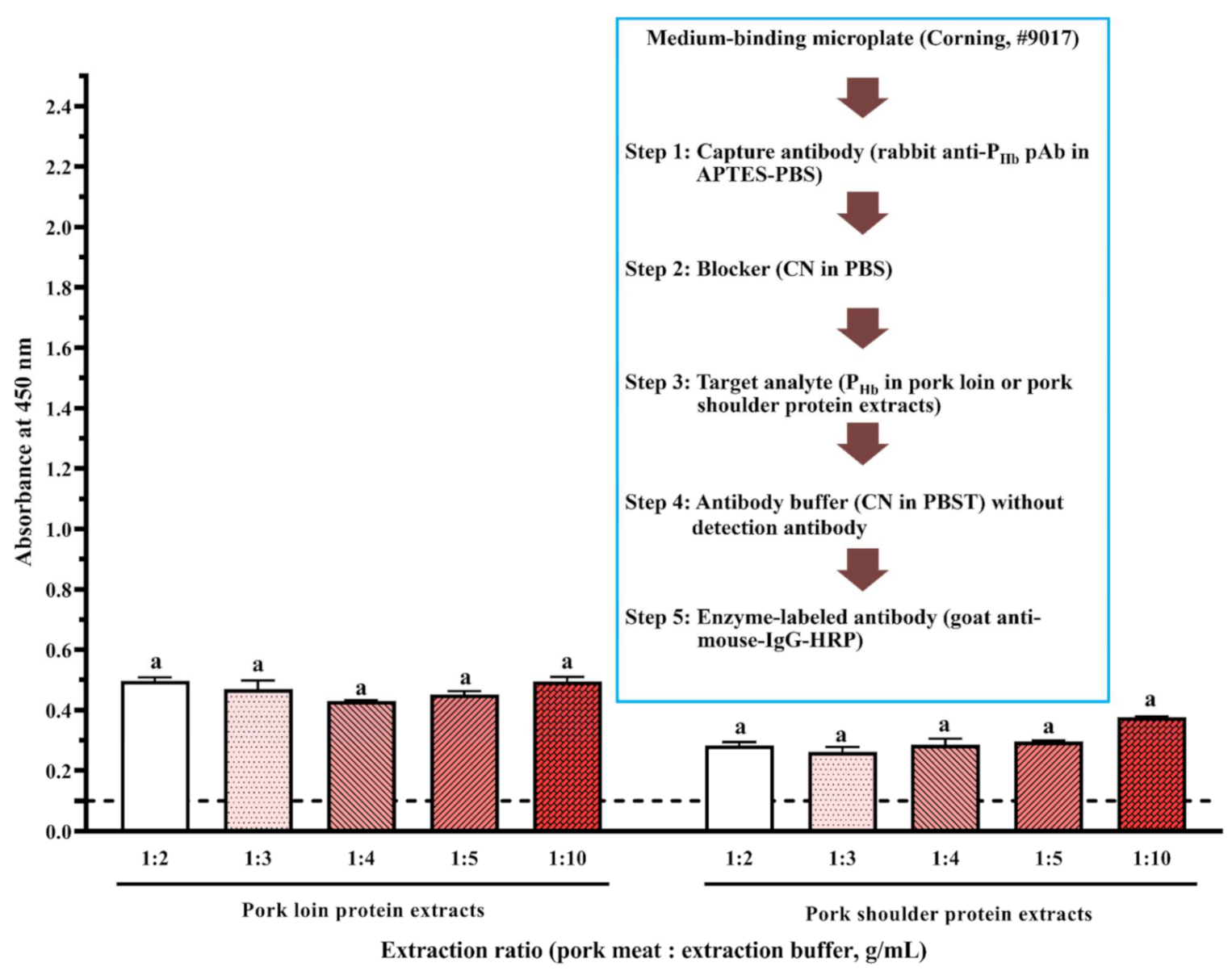
3.2.3. Effect of Microplate Type
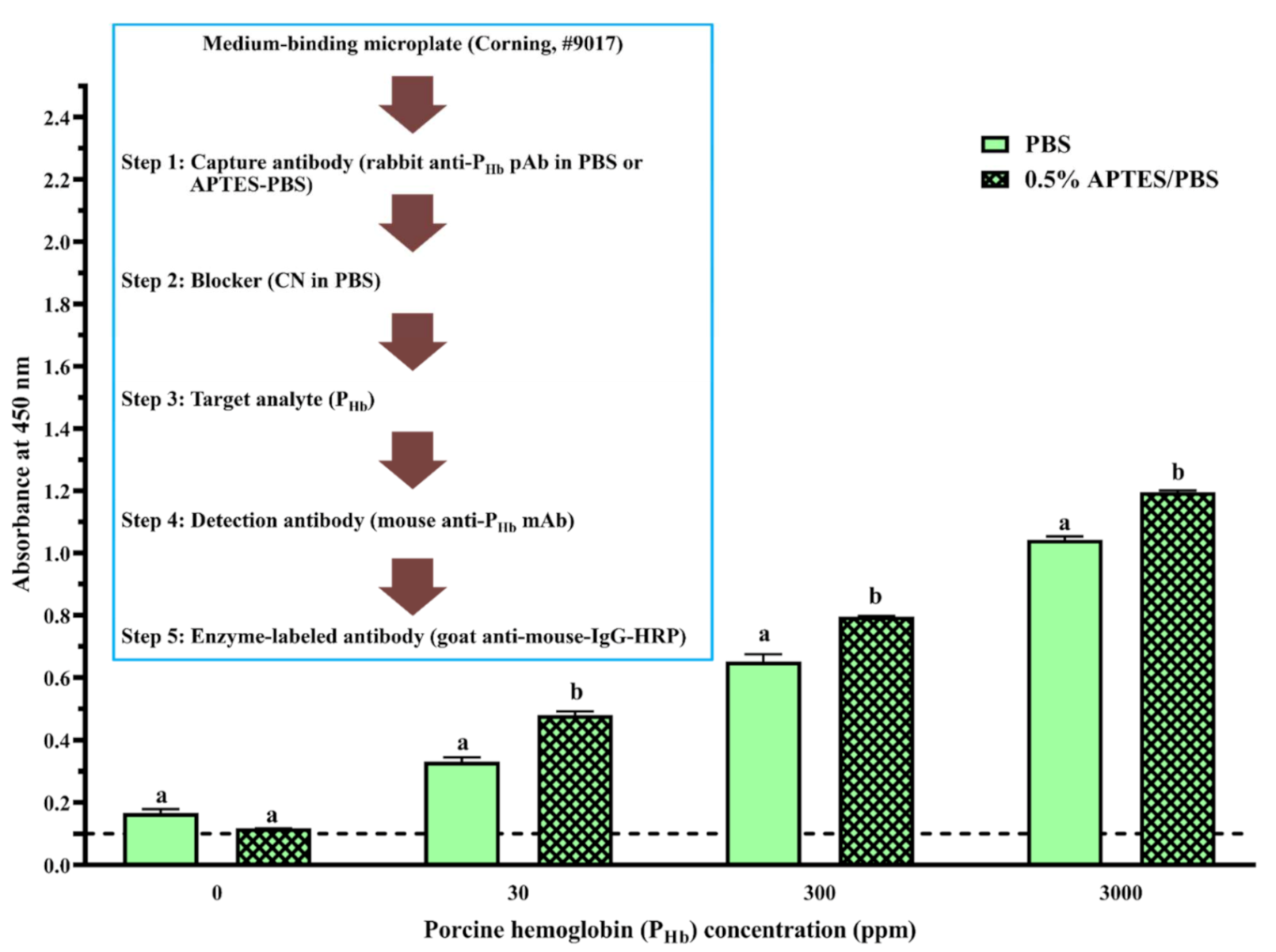
3.2.4. Effect of APTES during Coating
3.3. Cross-Reaction
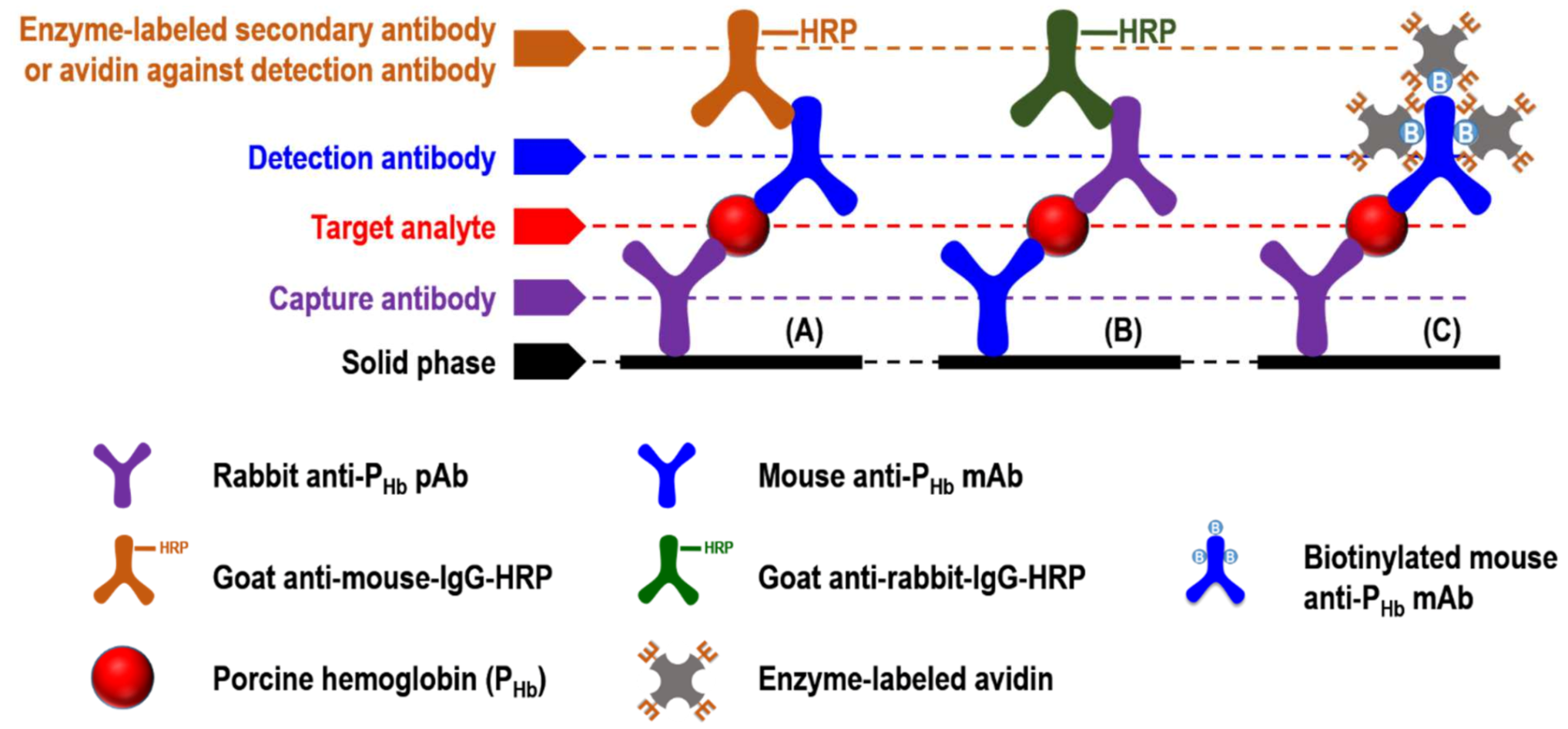
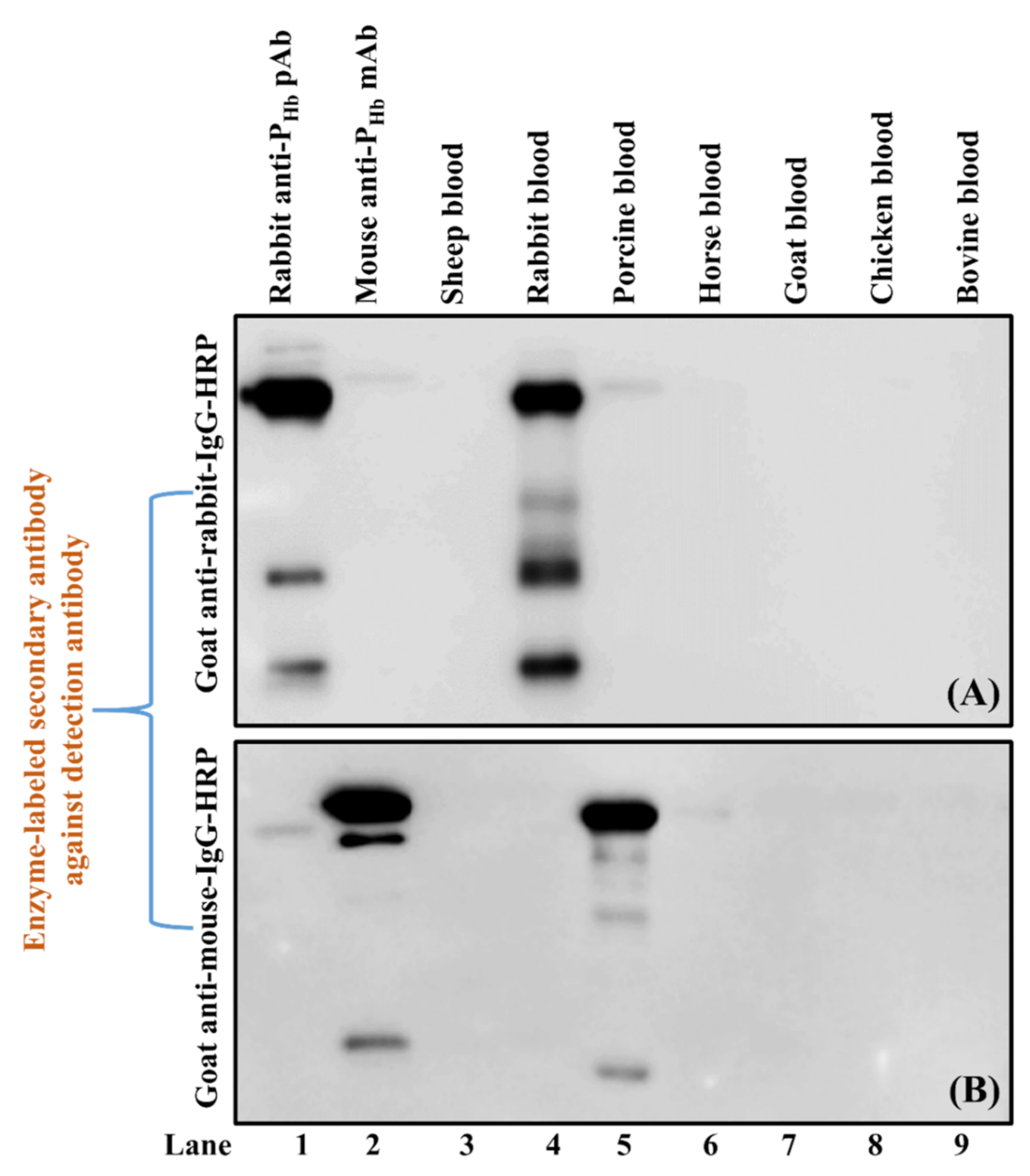
3.3.1. Cross-Reaction of the Enzyme-Labeled Secondary Antibody and Unintended IgGs
3.3.2. Cross-Reaction among Capture Antibody, Non-PHb Proteins, and Enzyme-Labeled Secondary Antibody
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Spink, J.; Moyer, D.C. Defining the public health threat of food fraud. J. Food Sci. 2011, 76, R157–R163. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.C.; Spink, J.; Lipp, M. Development and application of a database of food ingredient fraud and economically motivated adulteration from 1980 to 2010. J. Food Sci. 2012, 77, R118–R126. [Google Scholar] [CrossRef]
- Everstine, K.; Spink, J.; Kennedy, S. Economically motivated adulteration (EMA) of food: Common characteristics of EMA incidents. J. Food Protect. 2013, 76, 723–735. [Google Scholar] [CrossRef]
- Johnson, R. Food Fraud and “Economically Motivated Adulteration” of Food and Food Ingredients. Available online: https://fas.org/sgp/crs/misc/R43358.pdf (accessed on 12 July 2021).
- Points, J.; Mannings, L. Facing up to food fraud in a pandemic. Food Sci. Technol. 2020, 34, 16–20. [Google Scholar] [CrossRef]
- Knorr, D.; Khoo, C.S.H. COVID-19 and food: Challenges and research needs. Front. Nutr. 2020, 7, 598913. [Google Scholar] [CrossRef] [PubMed]
- Van Ruth, S.M. Impact of the COVID-19 Pandemic on Food Fraud Vulnerability in Food Supply Networks. Available online: https://research.wur.nl/en/publications/impact-of-the-covid-19-pandemic-on-food-fraud-vulnerability-in-fo (accessed on 12 July 2021).
- Asensio, L.; Gonzalez, I.; Garcia, T.; Martin, R. Determination of food authenticity by enzyme-linked immunosorbent assay (ELISA). Food Control 2008, 19, 1–8. [Google Scholar] [CrossRef]
- BCC Research. Global Markets and Technologies for Food Safety Testing. Available online: https://academic.bccresearch.com/market-research/food-and-beverage/food-safety-testing-technologies-markets-report.html (accessed on 12 July 2021).
- Grand View Research. Immunoassay Market Size, Share & Trends Analysis Report by Product (Reagent & Kits, Analyzers/Instruments, Software & Services), by Technology (RIA, ELISA, Rapid Test), by Application, by End Use, by Region, and Segment Forecasts, 2021–2028. Available online: https://www.grandviewresearch.com/industry-analysis/immunoassay-market (accessed on 12 July 2021).
- BCC Research. Immunoassays: Technologies and Global Markets—Focus on Enzyme Immunoassays. Available online: https://www.bccresearch.com/market-research/biotechnology/enzyme-immunoassays-bio120a.html (accessed on 12 July 2021).
- Engvall, E. The ELISA, enzyme-linked immunosorbent assay. Clin. Chem. 2010, 56, 319–320. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.; Maghsoudlou, P. Enzyme-linked immunosorbent assay (ELISA): The basics. Brit. J. Hosp. Med. 2016, 77, C98–C101. [Google Scholar] [CrossRef]
- Ciaurriz, P.; Fernandez, F.; Tellechea, E.; Moran, J.F.; Asensio, A.C. Comparison of four functionalization methods of gold nanoparticles for enhancing the enzyme-linked immunosorbent assay (ELISA). Beilstein J. Nanotech. 2017, 8, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Aaby, P.; Aftahi, N.; Aldrich, C.; Alzari, P.M.; Bevan, M.M. Chapter 4 Specificity and Cross-Reactivity. In Immunology and Evolution of Infectious Disease; Frank, S.A., Ed.; Princeton University Press: Princeton, NJ, USA, 2002; pp. 34–56. [Google Scholar]
- USFDA. Guidance for Industry Bioanalytical Method Validation. Available online: https://www.fda.gov/downloads/drugs/guidances/ucm070107.pdf (accessed on 12 July 2021).
- Crowther, J.R. Chapter 3 Stages in ELISA. In The ELISA Guidebook; Walker, J.M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2009; pp. 43–78. [Google Scholar]
- Jørgensen, C.S.; Hansen, K.B.; Jacobsen, S.; Halberg, P.; Ullman, S.; Hansen, D.; Mikkelsen, T.L.; Weile, B.; Madsen, M.H.; Wiik, A.; et al. Absence of high-affinity calreticulin autoantibodies in patients with systemic rheumatic diseases and coeliac disease. Scand. J. Clin. Lab. Inv. 2005, 65, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Péterfi, Z.; Kocsis, B. Comparison of blocking agents for an ELISA for LPS. J. Immunoass. 2000, 21, 341–354. [Google Scholar] [CrossRef]
- Güven, E.; Duus, K.; Lydolph, M.C.; Jørgensen, C.S.; Laursen, I.; Houen, G. Non-specific binding in solid phase immunoassays for autoantibodies correlates with inflammation markers. J. Immunol. Methods 2014, 403, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Konishi, E.; Kitai, Y.; Nishimura, K.; Harada, S. Antibodies to bovine serum albumin in human sera: Problems and solutions with casein-based ELISA in the detection of natural Japanese encephalitis virus infections. Jpn. J. Infect. Dis. 2010, 63, 296–298. [Google Scholar]
- Xiao, Y.; Isaacs, S.N. Enzyme-linked immunosorbent assay (ELISA) and blocking with bovine serum albumin (BSA)—Not all BSAs are alike. J. Immunol. Methods 2012, 384, 148–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chart, H.; Evans, J.; Chalmers, R.M.; Salmon, R.L. Escherichia coli O157 serology: False-positive ELISA results caused by human antibodies binding to bovine serum albumin. Lett. Appl. Microbiol. 1998, 27, 76–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, X.Y.; Wu, M.; Dong, W.Y.; Rao, Q.C.; Huo, H.L.; Han, Q.G. Monoclonal antibody-based sandwich enzyme-linked immunosorbent assay for porcine hemoglobin quantification. Food Chem. 2020, 324, 126880. [Google Scholar] [CrossRef]
- Corning Inc. Corning and Falcon Microplates Selection Guide: For Assays and Drug Discovery. Available online: https://www.corning.com/media/worldwide/cls/documents/CLS-C-DL-MP-014REV9.pdf (accessed on 12 July 2021).
- Ofori, J.A.; Hsieh, Y.-H.P. Monoclonal antibodies as probes for the detection of porcine blood derived food ingredients. J. Agr. Food Chem. 2016, 64, 3705–3711. [Google Scholar] [CrossRef]
- Jiang, X.Y.; Fuller, D.; Hsieh, Y.-H.P.; Rao, Q.C. Monoclonal antibody-based ELISA for the quantification of porcine hemoglobin in meat products. Food Chem. 2018, 250, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Prusaksochaczewski, E.; Luong, J.H.T. An improved ELISA method for the detection of Salmonella typhimurium. J. Appl. Bacteriol. 1989, 66, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Steinitz, M. Quantitation of the blocking effect of Tween 20 and bovine serum albumin in ELISA microwells. Anal. Biochem. 2000, 282, 232–238. [Google Scholar] [CrossRef]
- Hakami, A.R.; Ball, J.K.; Tarr, A.W. Non-ionic detergents facilitate non-specific binding of M13 bacteriophage to polystyrene surfaces. J. Virol. Methods 2015, 221, 1–8. [Google Scholar] [CrossRef]
- Julian, E.; Cama, M.; Martinez, P.; Luquin, M. An ELISA for five glycolipids from the cell wall of Mycobacterium tuberculosis: Tween 20 interference in the assay. J. Immunol. Methods 2001, 251, 21–30. [Google Scholar] [CrossRef]
- Bird, C.R.; Gearing, A.J.H.; Thorpe, R. The use of Tween-20 alone as a blocking-agent for immunoblotting can cause artefactual results. J. Immunol. Methods 1988, 106, 175–179. [Google Scholar] [CrossRef]
- Waritani, T.; Chang, J.; McKinney, B.; Terato, K. An ELISA protocol to improve the accuracy and reliability of serological antibody assays. MethodsX 2017, 4, 153–165. [Google Scholar] [CrossRef]
- Mohammad, K.; Esen, A. A blocking agent and a blocking step are not needed in ELISA, immunostaining dot-blots and Western blots. J. Immunol. Methods 1989, 117, 141–145. [Google Scholar] [CrossRef]
- Galva, C.; Gatto, C.; Milanick, M. Soymilk: An effective and inexpensive blocking agent for immunoblotting. Anal. Biochem. 2012, 426, 22–23. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, J. Effective Blocking Procedures: ELISA Technical Bulletin—No. 3. Available online: http://www.labcluster.com/news4_3/Corning_elisa3.pdf (accessed on 12 July 2021).
- Vogt, R.F.; Phillips, D.L.; Henderson, L.O.; Whitfield, W.; Spierto, F.W. Quantitative differences among various proteins as blocking agents for ELISA microtiter plates. J. Immunol. Methods 1987, 101, 43–50. [Google Scholar] [CrossRef]
- Gravel, P. Protein blotting by the semidry method. In The Protein Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 621–629. [Google Scholar]
- Yu, F.Y.; Vdovenko, M.M.; Wang, J.J.; Sakharov, I.Y. Comparison of enzyme-linked immunosorbent assays with chemiluminescent and colorimetric detection for the determination of ochratoxin A in food. J. Agr. Food Chem. 2011, 59, 809–813. [Google Scholar] [CrossRef] [PubMed]
- He, S.F.; Li, X.; Gao, J.Y.; Tong, P.; Chen, H.B. Development of sandwich ELISA for testing bovine beta-lactoglobulin allergenic residues by specific polyclonal antibody against human IgE binding epitopes. Food Chem. 2017, 227, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Zhang, Q.; Chen, D.F.; Liu, Y.; Li, C.Y.; Hu, B.S.; Du, D.; Liu, F.Q. Development of a specific enzyme-linked immunosorbent assay (ELISA) for the analysis of the organophosphorous pesticide fenthion in real samples based on monoclonal antibody. Anal. Lett. 2011, 44, 1591–1601. [Google Scholar] [CrossRef]
- Rajasekariah, G.H.R.; Ryan, J.R.; Hillier, S.R.; Yi, L.P.; Stiteler, J.M.; Cui, L.W.; Smithyman, A.M.; Martin, S.K. Optimisation of an ELISA for the serodiagnosis of visceral leishmaniasis using in vitro derived promastigote antigens. J. Immunol. Methods 2001, 252, 105–119. [Google Scholar] [CrossRef]
- Farajollahi, M.M.; Cook, D.B.; Hamzehlou, S.; Self, C.H. Reduction of non-specific binding in immunoassays requiring long incubations. Scand. J. Clin. Lab. Inv. 2012, 72, 531–539. [Google Scholar] [CrossRef]
- Ahirwar, R.; Bariar, S.; Balakrishnan, A.; Nahar, P. BSA blocking in enzyme-linked immunosorbent assays is a non-mandatory step: A perspective study on mechanism of BSA blocking in common ELISA protocols. RSC Adv. 2015, 5, 100077–100083. [Google Scholar] [CrossRef]
- Farrell, H.M.; Jimenez-Flores, R.; Bleck, G.T.; Brown, E.M.; Butler, J.E.; Creamer, L.K.; Hicks, C.L.; Hollar, C.M.; Ng-Kwai-Hang, K.F.; Swaisgood, H.E. Nomenclature of the proteins of cows’ milk—sixth revision. J. Dairy Sci. 2004, 87, 1641–1674. [Google Scholar] [CrossRef] [Green Version]
- Pratt, R.P.; Roser, B. Comparison of Blocking Agents for ELISA. Available online: https://assets.thermofisher.com/TFS-Assets/LSG/Application-Notes/D19561.pdf (accessed on 12 July 2021).
- Grogan, C.; Raiteri, R.; O’Connor, G.M.; Glynn, T.J.; Cunningham, V.; Kane, M.; Charlton, M.; Leech, D. Characterisation of an antibody coated microcantilever as a potential immuno-based biosensor. Biosens. Bioelectron. 2002, 17, 201–207. [Google Scholar] [CrossRef]
- Kenna, J.G.; Major, G.N.; Williams, R.S. Methods for reducing non-specific antibody binding in enzyme-linked immunosorbent assays. J. Immunol. Methods 1985, 85, 409–419. [Google Scholar] [CrossRef]
- Sentandreu, M.A.; Aubry, L.; Toldra, F.; Ouali, A. Blocking agents for ELISA quantification of compounds coming from bovine muscle crude extracts. Eur. Food Res. Technol. 2007, 224, 623–628. [Google Scholar] [CrossRef]
- Douglas, J.T.; Wu, Q.X.; Agustin, G.P.; Madarang, M.G. Evaluation of inexpensive blocking agents for ELISA in the detection of antibody in leprosy. Leprosy Rev. 1988, 59, 37–43. [Google Scholar] [CrossRef]
- Akerstedt, J. An indirect ELISA for detection of Encephalitozoon cuniculi infection in farmed blue foxes (Alopex lagopus). Acta Vet. Scand. 2002, 43, 211–220. [Google Scholar] [CrossRef]
- Huber, D.; Rudolf, J.; Ansari, P.; Galler, B.; Fuhrer, M.; Hasenhindl, C.; Baumgartner, S. Effectiveness of natural and synthetic blocking reagents and their application for detecting food allergens in enzyme-linked immunosorbent assays. Anal. Bioanal. Chem. 2009, 394, 539–548. [Google Scholar] [CrossRef]
- Hoffman, W.L.; Jump, A.A. Inhibition of the streptavidin-biotin interaction by milk. Anal. Biochem. 1989, 181, 318–320. [Google Scholar] [CrossRef]
- Leite, M.C.; Galland, F.; Brolese, G.; Guerra, M.C.; Bortolotto, J.W.; Freitas, R.; de Almeida, L.M.V.; Gottfried, C.; Goncalves, C.A. A simple, sensitive and widely applicable ELISA for S100B: Methodological features of the measurement of this glial protein. J. Neurosci. Methods 2008, 169, 93–99. [Google Scholar] [CrossRef]
- Tang, R.H.; Yang, H.; Choi, J.R.; Gong, Y.; Hu, J.; Feng, S.S.; Pingguan-Murphy, B.; Mei, Q.B.; Xu, F. Improved sensitivity of lateral flow assay using paper-based sample concentration technique. Talanta 2016, 152, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.X.; He, R.; Ju, X.R.; Zhang, J.; Wang, M.J.; Xing, C.R.; Yuan, J. Development and optimization of an immunoassay for the detection of Hg(II) in lake water. Food Sci. Nutr. 2019, 7, 1615–1622. [Google Scholar] [CrossRef] [Green Version]
- Thermo Fisher Scientific. Immunoassay Plate Guide. Available online: http://assets.thermofisher.com/TFS-Assets/LCD/Scientific-Resources/Immunoassay_Plate_Guide.pdf (accessed on 12 July 2021).
- Jones, E.; Michael, S.; Sittampalam, G. Basics of assay equipment and instrumentation for high throughput screening. In Assay Guidance Manual; Sittampalam, G.S., Coussens, N.P., Brimacombe, K., Grossman, A., Arkin, M., Auld, D., Austin, C., Baell, J., Bejcek, B., Caaveiro, J.M.M., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2012. [Google Scholar]
- Bergeron, A.B.; Bortz, C.J.; Rossi, A. Corning Medium and High Binding ELISA Microplates for Select Target Size Binding Assays Application Note. Available online: https://www.corning.com/media/worldwide/cls/documents/applications/CLS-AN-497%20DL.pdf (accessed on 12 July 2021).
- Gibbs, J.; Vessels, M.; Rothenberg, M. Immobilization Principles—Selecting the Surface. Available online: http://www.labcluster.com/news4_2/Corning_elisa1_2.pdf (accessed on 12 July 2021).
- The UniProt Consortium. UniProt: The universal protein knowledgebase. Nucleic Acids Res. 2017, 45, D158–D169. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Seidler, P.; Wan, L.S.; Fill, C. Formation, structure, and reactivity of amino-terminated organic films on silicon substrates. J. Colloid Interface Sci. 2009, 329, 114–119. [Google Scholar] [CrossRef]
- Dixit, C.K.; Vashist, S.K.; MacCraith, B.D.; O’Kennedy, R. Multisubstrate-compatible ELISA procedures for rapid and high-sensitivity immunoassays. Nat. Protoc. 2011, 6, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.H.; Jin, G. Silicon surface modification with a mixed silanes layer to immobilize proteins for biosensor with imaging ellipsometry. Colloid Surf. B 2004, 34, 173–177. [Google Scholar] [CrossRef]
- Cunliffe, D.; Smart, C.A.; Alexander, C.; Vulfson, E.N. Bacterial adhesion at synthetic surfaces. Appl. Environ. Microb. 1999, 65, 4995–5002. [Google Scholar] [CrossRef] [Green Version]
- Vashist, S.K.; Marion Schneider, E.; Lam, E.; Hrapovic, S.; Luong, J.H.T. One-step antibody immobilization-based rapid and highly-sensitive sandwich ELISA procedure for potential in vitro diagnostics. Sci. Rep. 2014, 4, 4407. [Google Scholar] [CrossRef]
- Dixit, C.K.; Vashist, S.K.; O’Neill, F.T.; O’Reilly, B.; MacCraith, B.D.; O’Kennedy, R. Development of a high sensitivity rapid sandwich ELISA procedure and its comparison with the conventional approach. Anal. Chem. 2010, 82, 7049–7052. [Google Scholar] [CrossRef]
- David, C.; Stålberg, J.; Larsson, A. Chicken antibodies: A clinical chemistry perspective. Ups. J. Med. Sci. 1999, 104, 179–189. [Google Scholar] [CrossRef]
- Boscato, L.M.; Stuart, M.C. Incidence and specificity of interference in two-site immunoassays. Clin. Chem. 1986, 32, 1491–1495. [Google Scholar] [CrossRef]
- Tate, J.; Ward, G. Interferences in immunoassay. Clin. Biochem. Rev. 2004, 25, 105–120. [Google Scholar] [PubMed]
- R&D Systems. Goat Anti-Mouse IgG Biotinylated Antibody. Available online: https://www.rndsystems.com/products/goat-anti-mouse-igg-biotinylated-antibody_baf007 (accessed on 12 July 2021).
- Thermo Fisher Scientific. Secondary Antibody Cross-Adsorption and Cross-Reactivity. Available online: https://www.thermofisher.com/us/en/home/life-science/antibodies/antibodies-learning-center/antibodies-resource-library/antibody-methods/secondary-antibody-cross-adsorption-cross-reactivity.html#:~:text=Cross%2Dadsorbed%20secondary%20antibodies%20are,cross%2Dreactivity%20and%20increases%20specificity (accessed on 12 July 2021).
- Abcam. Pre-Adsorbed Secondary Antibodies. Available online: https://www.abcam.com/secondary-antibodies/pre-adsorbed-secondary-antibodies (accessed on 12 July 2021).
- Sigma-Aldrich. Anti-Mouse IgG (H + L), Highly Cross Adsorbed Antibody Produced in Goat. Available online: https://www.sigmaaldrich.com/catalog/product/sigma/sab3701071?lang=en®ion=US (accessed on 12 July 2021).
- Jackson ImmunoResearch Laboratories Inc. Cross-Adsorbed (Min X) Secondary Antibodies and Cross-Reactivity. Available online: https://www.jacksonimmuno.com/technical/products/cross-adsorbed-secondary-antibodies (accessed on 12 July 2021).
- Lipman, N.S.; Jackson, L.R.; Trudel, L.J.; Weis-Garcia, F. Monoclonal versus polyclonal antibodies: Distinguishing characteristics, applications, and information resources. ILAR J. 2005, 46, 258–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Wang, Y.; Li, Z.; Pramod, S.N.; Zhang, L.; Lin, H. Development of ELISA method for detecting crustacean major allergen tropomyosin in processed food samples. Food Anal. Methods 2019, 12, 2719–2729. [Google Scholar] [CrossRef]
- Bordeaux, J.; Welsh, A.W.; Agarwal, S.; Killiam, E.; Baquero, M.T.; Hanna, J.A.; Anagnostou, V.K.; Rimm, D.L. Antibody validation. BioTechniques 2010, 48, 197–209. [Google Scholar] [CrossRef] [Green Version]
- Weller, M.G. Ten basic rules of antibody validation. Anal. Chem. Insights 2018, 13, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlen, M.; Bandrowski, A.; Carr, S.; Edwards, A.; Ellenberg, J.; Lundberg, E.; Rimm, D.L.; Rodriguez, H.; Hiltke, T.; Snyder, M.; et al. A proposal for validation of antibodies. Nat. Methods 2016, 13, 823–828. [Google Scholar] [CrossRef]
- Staggs, C.G.; Sealey, W.M.; McCabe, B.J.; Teague, A.M.; Mock, D.M. Determination of the biotin content of select foods using accurate and sensitive HPLC/avidin binding. J. Food Compost. Anal. 2004, 17, 767–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USFDA. Biotin Interference with Troponin Lab Tests—Assays Subject to Biotin Interference. Available online: https://www.fda.gov/medical-devices/in-vitro-diagnostics/biotin-interference-troponin-lab-tests-assays-subject-biotin-interference (accessed on 12 July 2021).
- Holmes, E.W.; Samarasinghe, S.; Emanuele, M.A.; Meah, F. Biotin interference in clinical immunoassays: A cause for concern. Arch. Pathol. Lab. Med. 2017, 141, 1459–1460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colon, P.J.; Greene, D.N. Biotin interference in clinical immunoassays. J. Appl. Lab. Med. 2018, 2, 941–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luong, J.H.T.; Vashist, S.K. Chemistry of biotin–streptavidin and the growing concern of an emerging biotin interference in clinical immunoassays. ACS Omega 2020, 5, 10–18. [Google Scholar] [CrossRef] [Green Version]
- Gifford, J.L.; de Koning, L.; Sadrzadeh, S.M.H. Strategies for mitigating risk posed by biotin interference on clinical immunoassays. Clin. Biochem. 2019, 65, 61–63. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Preparation Steps | Sample Model 1 | Sample Model 2 | Sample Model 3 |
|---|---|---|---|
| Meat Protein Extracts | Whole Blood Protein Extracts | PHb (H4131, Sigma-Aldrich) | |
| 1. Extraction/dissolving (12.5 mM NaHCO3 and 25 mM NaCl, pH 8.3) | 1:2, 1:3, 1:4, 1:5, 1:10 (g/mL) * | 1:100 (mL/mL) | 3 mg/mL |
| 2. Homogenization (11,000 rpm, 2 min) | Yes | No | No |
| 3. Sonication (130 W, 15 min) | Yes | Yes | No |
| 4. End-over-end rotation (1 h, 4 °C) | Yes | Yes | No |
| 5. Centrifugation (20,000× g, 15 min, 4 °C) | Yes | Yes | No |
| 6. Protein concentration determination (BCA assay) | Yes | Yes | No |
| 7. Relevant figures | 6 and 7 | 7 and 8 | 2, 3, 4 and 5 |
| Blockers | Component I | Concentration (%, g/mL) | Component II | Concentration (%, g/mL) |
|---|---|---|---|---|
| Protein-based | ||||
| BSA | Bovine serum albumin | 5 | ||
| CN | Casein | 1 | ||
| NFDM | Non-fat dry milk | 5 | ||
| FG | Fish gelatin | 1 | ||
| Non-protein-based | ||||
| PEG + CN | Polyethylene glycol | 5 | Casein | 1 |
| PVP + CN | Polyvinylpyrrolidone | 5 | Casein | 1 |
| Experiment No. | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Schematics | Figure 1B | Figure 1B | Figure 1C | Figure 1A | Figure 1B |
| Objectives | Non-specific binding | Cross-reaction | |||
| Step 1: Capture antibody in PBS | None | Rabbit anti-PHb pAb | Mouse anti-PHb mAb | ||
| Step 2: Blocker | CN in PBS | ||||
| Step 3: Target analyte (PHb) | None | ||||
| Step 4: Detection antibody in antibody buffer (CN in PBST) | None | Mouse anti-PHb mAb | Biotinylated mouse anti-PHb mAb | None | None |
| Step 5: Enzyme-labeled antibody against detection antibody in antibody buffer | Goat anti-mouse-IgG-HRP (RRID: AB_258008) | None | Goat anti-mouse-IgG-HRP (RRID: AB_258008) | Goat anti-rabbit-IgG-HRP (RRID: AB_257896) | |
| Streptavidin-HRP conjugate in PBST | |||||
| A450 (mean ± SEM) | 0.044 ± 0 | 0.050 ± 0.003 | 0.050 ± 0.003 | 0.176 ± 0.007 | 0.644 ± 0.013 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.; Wu, M.; Albo, J.; Rao, Q. Non-Specific Binding and Cross-Reaction of ELISA: A Case Study of Porcine Hemoglobin Detection. Foods 2021, 10, 1708. https://doi.org/10.3390/foods10081708
Jiang X, Wu M, Albo J, Rao Q. Non-Specific Binding and Cross-Reaction of ELISA: A Case Study of Porcine Hemoglobin Detection. Foods. 2021; 10(8):1708. https://doi.org/10.3390/foods10081708
Chicago/Turabian StyleJiang, Xingyi, Meng Wu, Jonathan Albo, and Qinchun Rao. 2021. "Non-Specific Binding and Cross-Reaction of ELISA: A Case Study of Porcine Hemoglobin Detection" Foods 10, no. 8: 1708. https://doi.org/10.3390/foods10081708
APA StyleJiang, X., Wu, M., Albo, J., & Rao, Q. (2021). Non-Specific Binding and Cross-Reaction of ELISA: A Case Study of Porcine Hemoglobin Detection. Foods, 10(8), 1708. https://doi.org/10.3390/foods10081708