
A Targeted Metabolomics Approach to Study Secondary Metabolites and Antioxidant Activity in ‘Kinnow Mandarin’ during Advanced Fruit Maturity

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Fruit Material and Sampling
2.3. Flavonoids and Phenolics
2.4. Limonoid Aglycones and Glycosides
2.5. MS/MS Conditions
2.6. Limit of Detection (LOD), Limit of Quantification (LOQ), Correlation Coefficient (r) and Calibration Range
2.7. Antioxidant Activity
2.8. Statistical and Multivariate Analysis
3. Results and Discussion
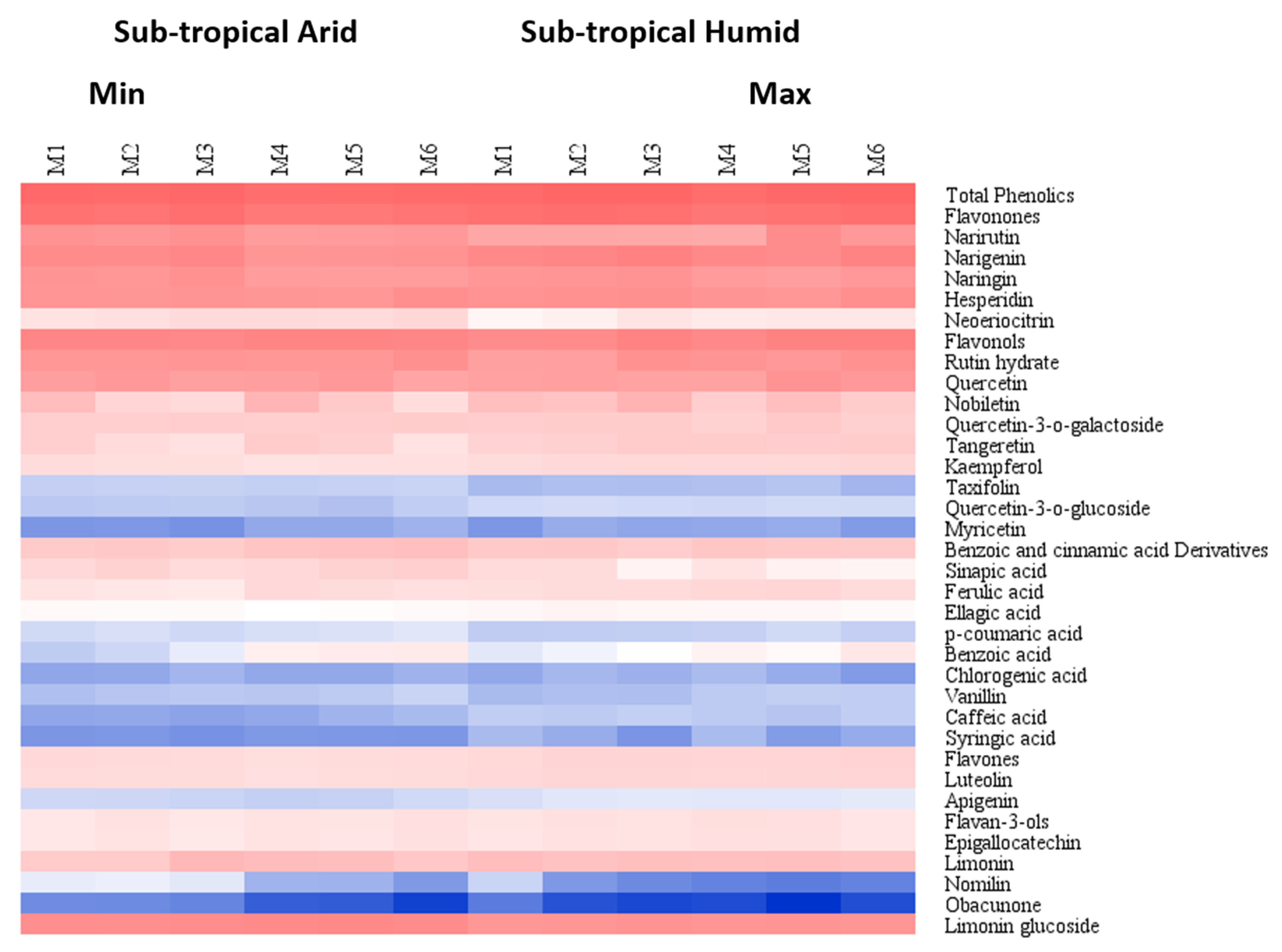
3.1. Flavonoids and Phenolics
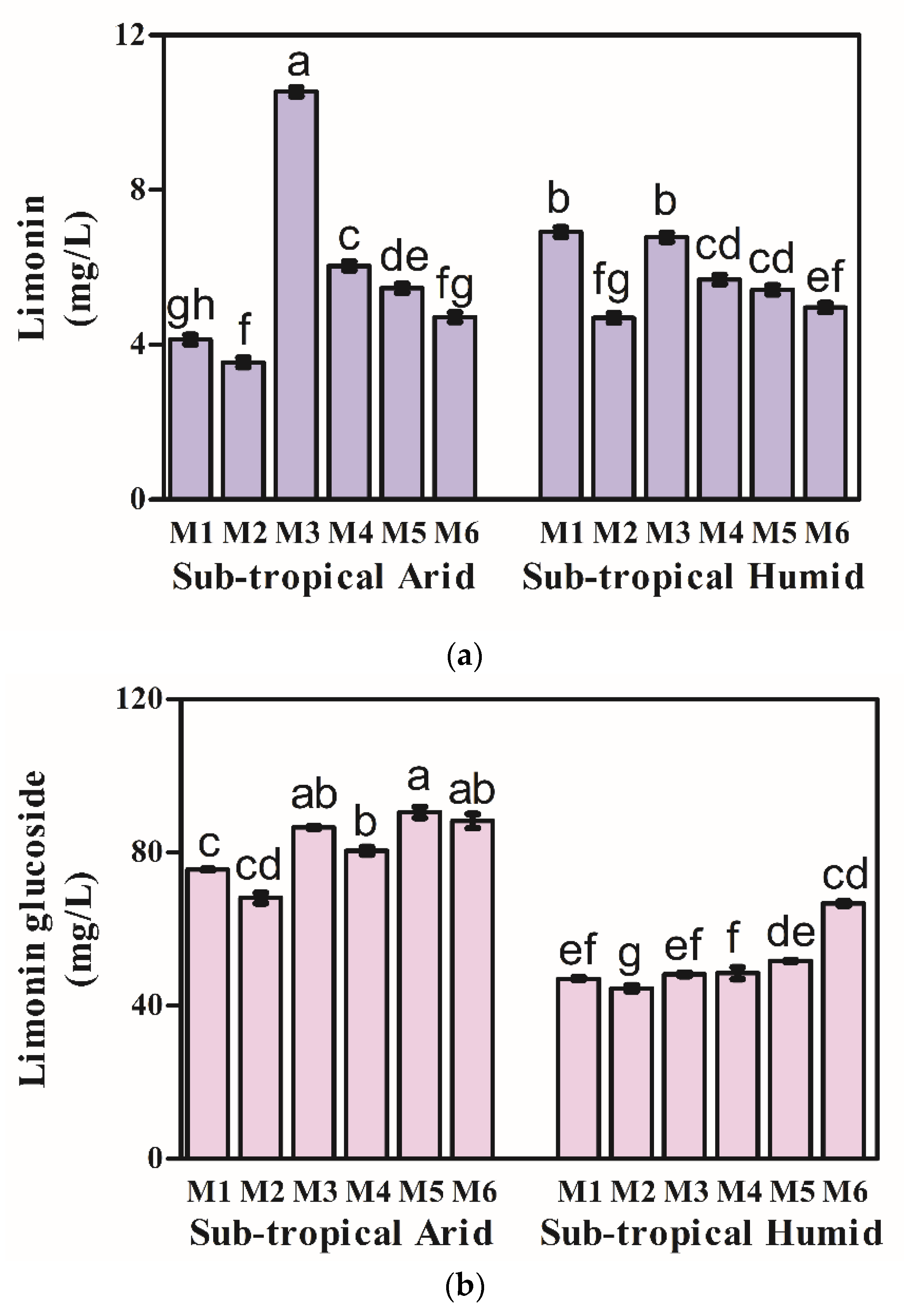
3.2. Limonoids
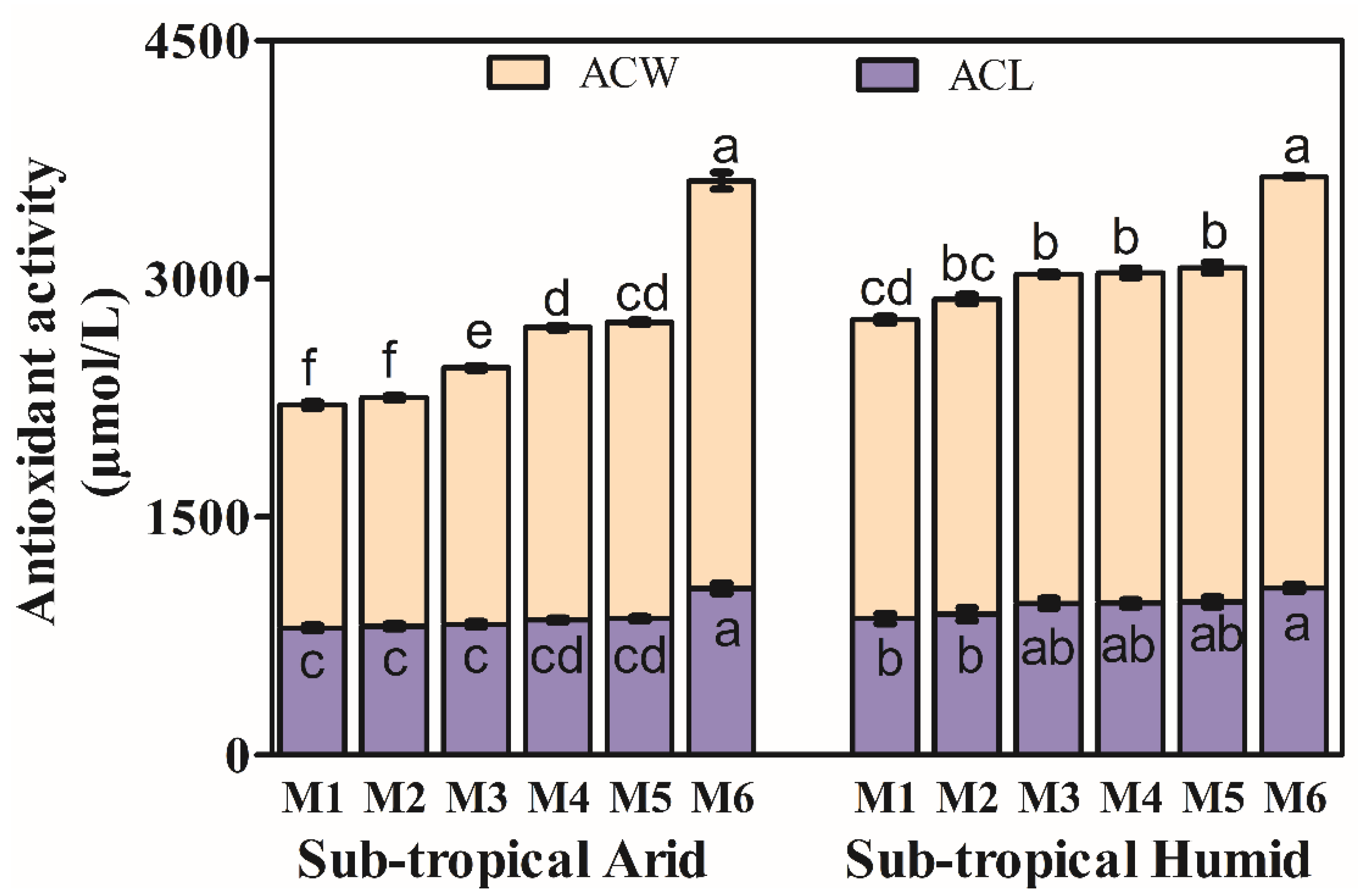
3.3. Antioxidant Activity
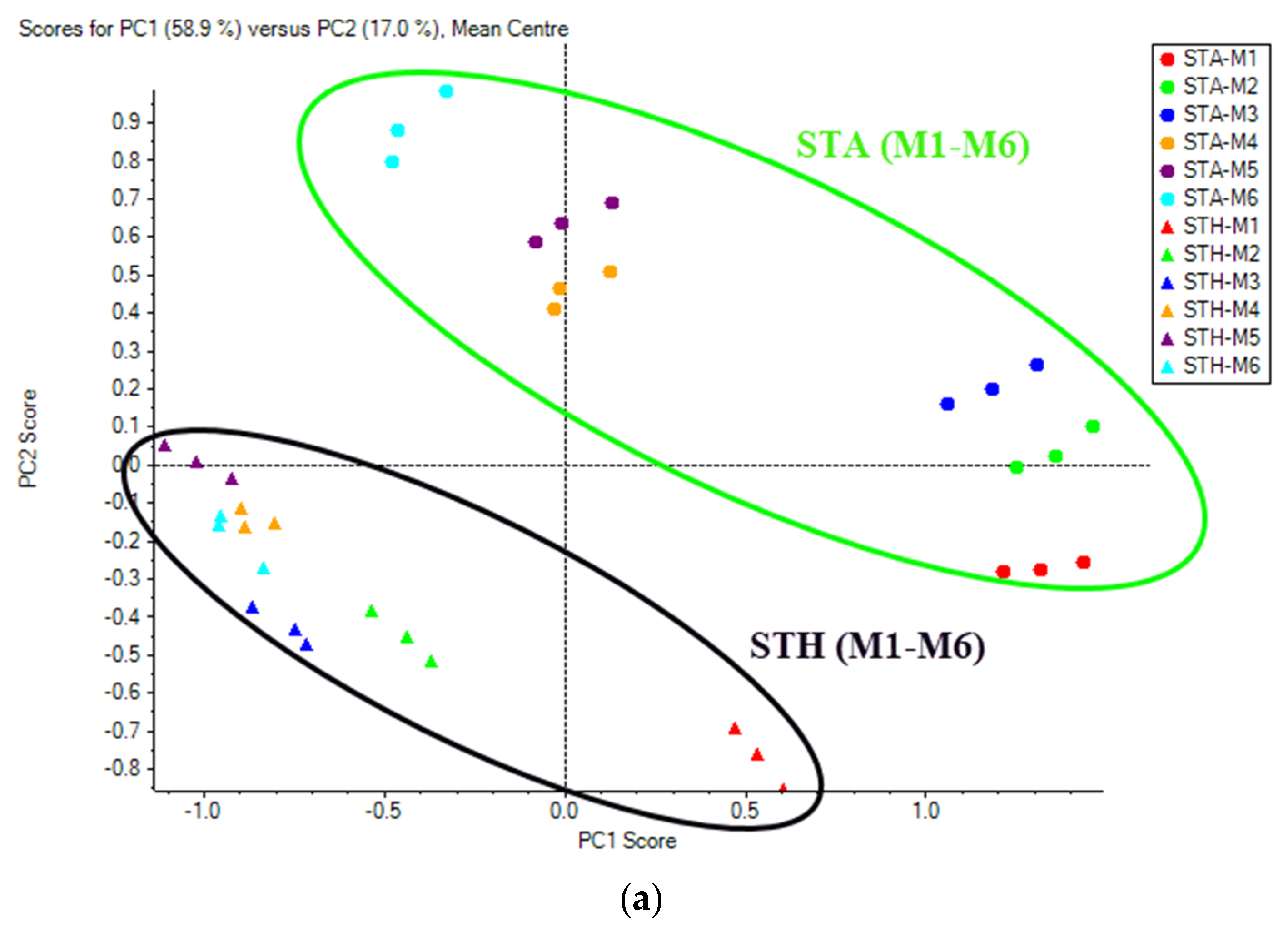
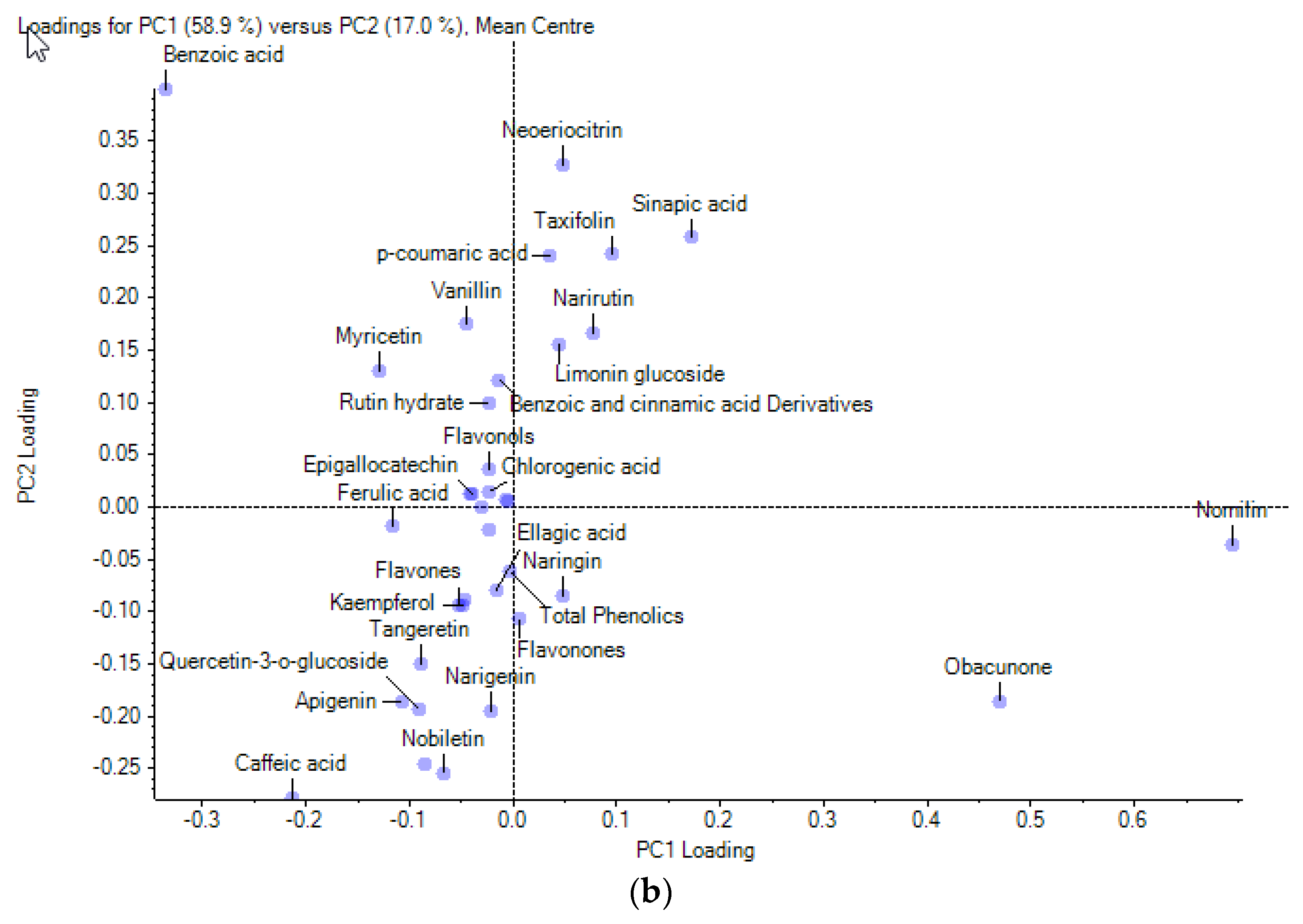
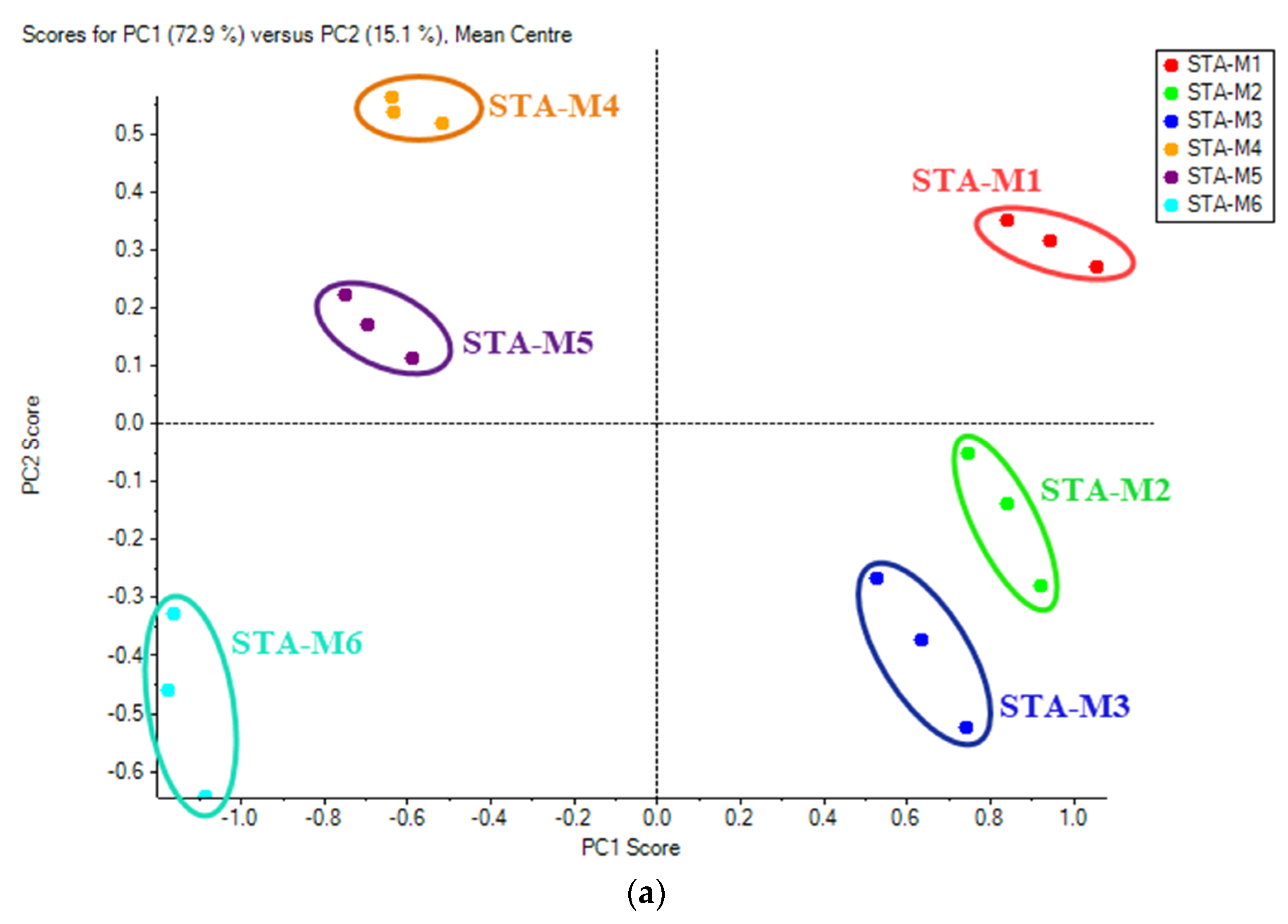
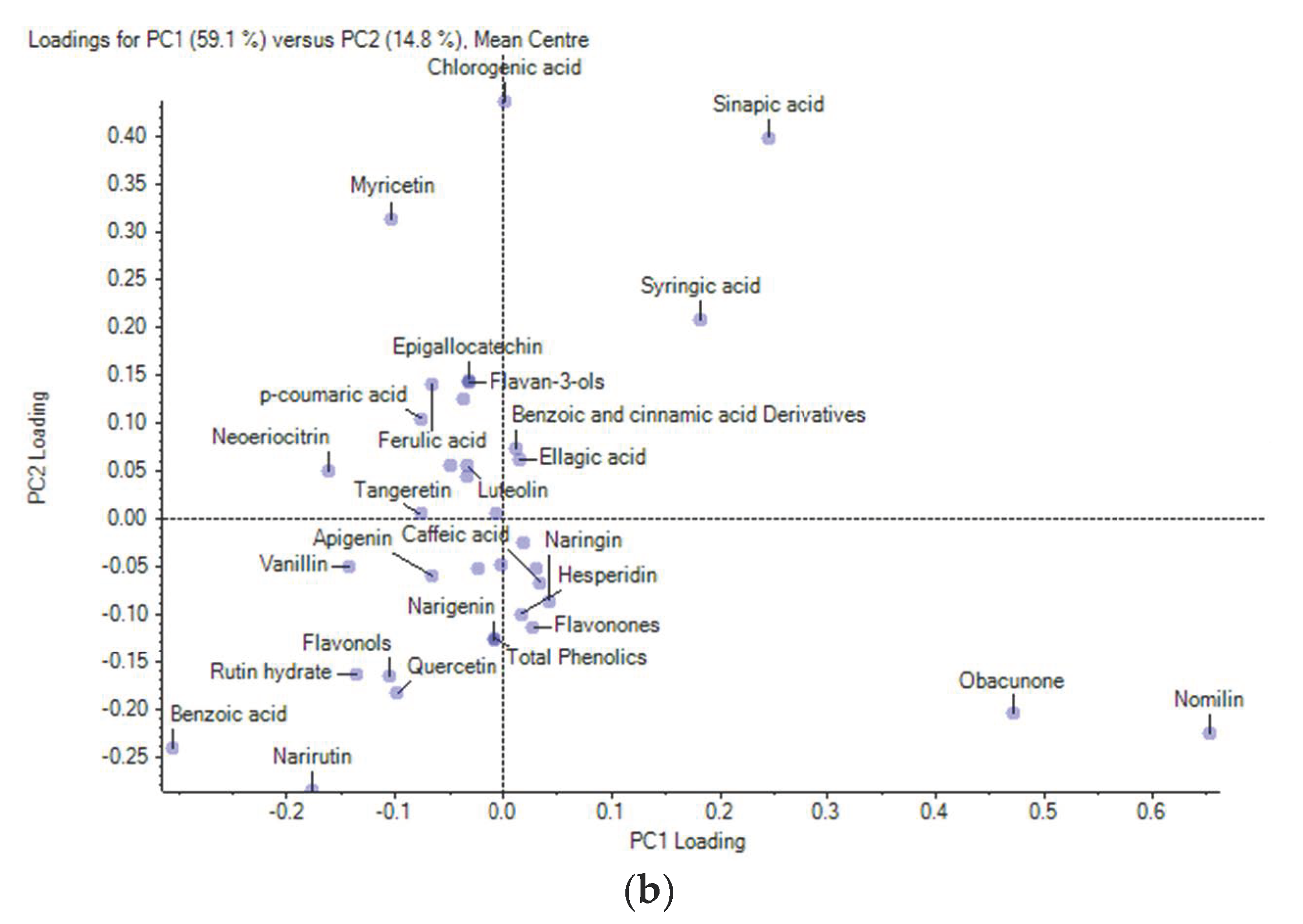
3.4. Multivariate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Qian, J.; Cao, J.; Wang, D.; Liu, C.; Yang, R.; Li, X.; Sun, C. Antioxidant Capacity, Anticancer Ability and Flavonoids Composition of 35 Citrus (Citrus Reticulata Blanco) Varieties. Molecules 2017, 22, 1114. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Ma, S.; Ren, D. Phytochemistry and Bioactivity of Citrus Flavonoids: A Focus on Antioxidant, Anti-Inflammatory, Anticancer and Cardiovascular Protection Activities. Phytochem. Rev. 2017, 16, 479–511. [Google Scholar] [CrossRef]
- Kashyap, D.; Mittal, S.; Sak, K.; Singhal, P.; Tuli, H.S. Molecular Mechanisms of Action of Quercetin in Cancer: Recent Advances. Tumor Biol. 2016, 37, 12927–12939. [Google Scholar] [CrossRef] [PubMed]
- Cirmi, S.; Maugeri, A.; Ferlazzo, N.; Gangemi, S.; Calapai, G.; Schumacher, U.; Navarra, M. Anticancer Potential of Citrus Juices and Their Extracts: A Systematic Review of Both Preclinical and Clinical Studies. Front. Pharmacol. 2017, 8, 420. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Ren, C.; Cao, Y.; Wang, Y.; Duan, W.; Xie, L.; Sun, C.; Li, X. Characterization and Purification of Bergamottin from Citrus Grandis (L.) Osbeck Cv. Yongjiazaoxiangyou and Its Antiproliferative Activity and Effect on Glucose Consumption in HepG2 Cells. Molecules 2017, 22, 1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shankar, E.; Goel, A.; Gupta, K.; Gupta, S. Plant Flavone Apigenin: An Emerging Anticancer Agent. Curr. Pharmacol. Rep. 2017, 3, 423–446. [Google Scholar] [CrossRef]
- Yang, J.; Fa, J.; Li, B. Apigenin Exerts Anticancer Effects on Human Cervical Cancer Cells via Induction of Apoptosis and Regulation of Raf/MEK/ERK Signalling Pathway. Trop. J. Pharm. Res. 2018, 17, 1615. [Google Scholar] [CrossRef]
- Mallick, N.; Khan, R. Antihyperlipidemic Effects of Citrus Sinensis, Citrus Paradisi, and Their Combinations. J. Pharm. Bioallied Sci. 2016, 8, 112. [Google Scholar] [CrossRef]
- Zeka, K.; Ruparelia, K.; Arroo, R.; Budriesi, R.; Micucci, M. Flavonoids and Their Metabolites: Prevention in Cardiovascular Diseases and Diabetes. Diseases 2017, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Xie, X.; Lin, Q.; Chen, J.; Grierson, D.; Yin, X.; Sun, C.; Chen, K. Differential Expression of Organic Acid Degradation-Related Genes During Fruit Development of Navel Oranges (Citrus Sinensis) in Two Habitats. Plant. Mol. Biol. Rep. 2013, 31, 1131–1140. [Google Scholar] [CrossRef]
- Hassan, Z.H.; Lesmayati, S.; Qomariah, R.; Hasbianto, A. Effects of Wax Coating Applications and Storage Temperatures on the Quality of Tangerine Citrus (Citrus Reticulata) var. Siam Banjar. Int. Food Res. J. 2014, 21, 641. [Google Scholar]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and Anti-Inflammatory Properties of the Citrus Flavonoids Hesperidin and Hesperetin: An Updated Review of Their Molecular Mechanisms and Experimental Models: Hesperidin and Hesperetin as Antioxidant and Anti-Inflammatory Agents. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Gualdani, R.; Cavalluzzi, M.; Lentini, G.; Habtemariam, S. The Chemistry and Pharmacology of Citrus Limonoids. Molecules 2016, 21, 1530. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Ren, Y.; Lin, D.; Peng, S.; Zhong, B.; Ma, Z. The Anti-Inflammatory Properties of Citrus Wilsonii Tanaka Extract in LPS-Induced RAW 264.7 and Primary Mouse Bone Marrow-Derived Dendritic Cells. Molecules 2017, 22, 1213. [Google Scholar] [CrossRef]
- da Silva, S.A.V.; Clemente, A.; Rocha, J.; Direito, R.; Marques, H.C.; Sepodes, B.; Figueira, M.-E.; Ribeiro, M.H. Anti-Inflammatory Effect of Limonin from Cyclodextrin (Un)Processed Orange Juices in In Vivo Acute Inflammation and Chronic Rheumatoid Arthritis Models. J. Funct. Foods 2018, 49, 146–153. [Google Scholar] [CrossRef]
- Hagenlocher, Y.; Gommeringer, S.; Held, A.; Feilhauer, K.; Köninger, J.; Bischoff, S.C.; Lorentz, A. Nobiletin Acts Anti-Inflammatory on Murine IL-10-/- Colitis and Human Intestinal Fibroblasts. Eur. J. Nutr. 2019, 58, 1391–1401. [Google Scholar] [CrossRef]
- Tejada, S.; Pinya, S.; Martorell, M.; Capó, X.; Tur, J.A.; Pons, A.; Sureda, A. Potential Anti-Inflammatory Effects of Hesperidin from the Genus Citrus. Curr. Med. Chem. 2019, 25, 4929–4945. [Google Scholar] [CrossRef]
- Battinelli, L.; Mengoni, F.; Lichtner, M.; Mazzanti, G.; Saija, A.; Mastroianni, C.M.; Vullo, V. Effect of Limonin and Nomilin on HIV-1 Replication on Infected Human Mononuclear Cells. Planta Med. 2003, 69, 910–913. [Google Scholar]
- Farooq, T.; Hameed, A.; Rehman, K.; Ibrahim, M.; Qadir, M.I.; Akash, M.S.H. Antiretroviral Agents: Looking for the Best Possible Chemotherapeutic Options to Conquer HIV. Crit. Rev. Eukaryot. Gene Expr. 2016, 26, 363–381. [Google Scholar] [CrossRef]
- Lee, G.-E.; Kim, J.; Shin, C.-G. Antiviral Activities of Hydroxylated Flavones on Feline Foamy Viral Proliferation. Appl. Biol. Chem. 2017, 60, 419–425. [Google Scholar] [CrossRef]
- Lado, J.; Gambetta, G.; Zacarias, L. Key Determinants of Citrus Fruit Quality: Metabolites and Main Changes during Maturation. Sci. Hortic. 2018, 233, 238–248. [Google Scholar] [CrossRef] [Green Version]
- Iglesias, D.J.; Cercós, M.; Colmenero-Flores, J.M.; Naranjo, M.A.; Ríos, G.; Carrera, E.; Ruiz-Rivero, O.; Lliso, I.; Morillon, R.; Tadeo, F.R.; et al. Physiology of Citrus Fruiting. Braz. J. Plant Physiol. 2007, 19, 333–362. [Google Scholar] [CrossRef]
- Noomhorm, A.; Kasemsuksakul, N. Effect of Maturity and Processing on Bitter Compounds in Thai Tangerine Juice. Int. J. Food Sci. Technol. 2007, 27, 65–72. [Google Scholar] [CrossRef]
- Yoo, K.M.; Lee, K.W.; Park, J.B.; Lee, H.J.; Hwang, I.K. Variation in Major Antioxidants and Total Antioxidant Activity of Yuzu (Citrus Junos Sieb Ex Tanaka) during Maturation and between Cultivars. J. Agric. Food Chem. 2004, 52, 5907–5913. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Ye, X.; Liu, D.; Ma, Y.; Chen, J. Composition and Distribution of Phenolic Acids in Ponkan (Citrus Poonensis Hort. Ex Tanaka) and Huyou (Citrus Paradisi Macf. Changshanhuyou) during Maturity. J. Food Compos. Anal. 2008, 21, 382–389. [Google Scholar] [CrossRef]
- Fong, C.H.; Hasegawa, S.; Coggins, C.W.; Atkin, D.R.; Miyake, M. Contents of Limonoids and Limonin 17-.Beta.-D-Glucopyranoside in Fruit Tissue of Valencia Orange during Fruit Growth and Maturation. J. Agric. Food Chem. 1992, 40, 1178–1181. [Google Scholar] [CrossRef]
- Barreca, D.; Bellocco, E.; Caristi, C.; Leuzzi, U.; Gattuso, G. Flavonoid Composition and Antioxidant Activity of Juices from Chinotto (Citrus × Myrtifolia Raf.) Fruits at Different Ripening Stages. J. Agric. Food Chem. 2010, 58, 3031–3036. [Google Scholar] [CrossRef]
- Kimball, D.A. Factors Affecting the Rate of Maturation of Citrus Fruits. In Proceedings of the Florida State Horticultural Society. 1984, Volume 97, pp. 40–44. Available online: https://journals.flvc.org/fshs/article/download/95254/91271 (accessed on 3 March 2022).
- Reuther, W. Climatic Effects and Quality of Citrus in the Tropics. In Proceedings of the Tropical Region, American Society for Horticultural Science. 1980, Volume 24, pp. 15–18. Available online: https://www.cabdirect.org/cabdirect/abstract/19850331515 (accessed on 3 March 2022).
- Peng, Y.; Liu, F.; Ye, J. Quantitative and Qualitative Analysis of Flavonoid Markers in Frucus Aurantii of Different Geographical Origin by Capillary Electrophoresis with Electrochemical Detection. J. Chromatogr. B 2006, 830, 224–230. [Google Scholar] [CrossRef]
- Goulas, V.; Manganaris, G.A. Exploring the Phytochemical Content and the Antioxidant Potential of Citrus Fruits Grown in Cyprus. Food Chem. 2012, 131, 39–47. [Google Scholar] [CrossRef]
- Zhang, M.; Nan, H.; Wang, Y.; Jiang, X.; Li, Z. Comparison of Flavonoid Compounds in the Flavedo and Juice of Two Pummelo Cultivars (Citrus grandis L. Osbeck) from Different Cultivation Regions in China. Molecules 2014, 19, 17314–17328. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.D.; Ko, W.J.; Koh, K.S.; Jeon, Y.J.; Kim, S.H. Composition of Flavonoids and Antioxidative Activity from Juice of Jeju Native Citrus Fruits during Maturation. Korean J. Nutr. 2009, 42, 278. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Heying, E.; Tanumihardjo, S.A. History, Global Distribution, and Nutritional Importance of Citrus Fruits. Compr. Rev. Food Sci. Food Saf. 2012, 11, 530–545. [Google Scholar] [CrossRef]
- Lado, J.; Cuellar, F.; Rodrigo, M.J.; Zacarías, L. Nutritional Composition of Mandarins. In Nutritional Composition of Fruit Cultivars; Elsevier, 2016; pp. 419–443. ISBN 978-0-12-408117-8. Available online: https://doi.org/10.1016/B978-0-12-408117-8.00018-0 (accessed on 3 March 2022).
- Dettmer, K.; Aronov, P.A.; Hammock, B.D. Mass Spectrometry-Based Metabolomics. Mass Spectrom. Rev. 2007, 26, 51–78. [Google Scholar] [CrossRef] [PubMed]
- Gika, H.G.; Theodoridis, G.A.; Plumb, R.S.; Wilson, I.D. Current Practice of Liquid Chromatography–Mass Spectrometry in Metabolomics and Metabonomics. J. Pharm. Biomed. Anal. 2014, 87, 12–25. [Google Scholar] [CrossRef]
- Ara, T.; Waki, N.; Sakurai, N.; Aizawa, K.; Suganuma, H.; Matsumura, Y.; Kawada, T.; Shibata, D.; Takahashi, S. Comparative Metabolomic Analysis of Two Tomato Cultivars during Fruit Maturation. In Proceedings of the XIV International Symposium on Processing Tomato 1159. 2016, pp. 101–106. Available online: https://doi.org/10.17660/ActaHortic.2017.1159.16 (accessed on 3 March 2022).
- Monti, L.L.; Bustamante, C.A.; Osorio, S.; Gabilondo, J.; Borsani, J.; Lauxmann, M.A.; Maulión, E.; Valentini, G.; Budde, C.O.; Fernie, A.R.; et al. Metabolic Profiling of a Range of Peach Fruit Varieties Reveals High Metabolic Diversity and Commonalities and Differences during Ripening. Food Chem. 2016, 190, 879–888. [Google Scholar] [CrossRef]
- Duan, S.; Wu, Y.; Fu, R.; Wang, L.; Chen, Y.; Xu, W.; Zhang, C.; Ma, C.; Shi, J.; Wang, S. Comparative Metabolic Profiling of Grape Skin Tissue along Grapevine Berry Developmental Stages Reveals Systematic Influences of Root Restriction on Skin Metabolome. Int. J. Mol. Sci. 2019, 20, 534. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Sun, L.; Xie, L.; He, Y.; Luo, T.; Sheng, L.; Luo, Y.; Zeng, Y.; Xu, J.; Deng, X.; et al. Regulation of Cuticle Formation during Fruit Development and Ripening in ‘Newhall’ Navel Orange (Citrus Sinensis Osbeck) Revealed by Transcriptomic and Metabolomic Profiling. Plant. Sci. 2016, 243, 131–144. [Google Scholar] [CrossRef]
- Ledesma-Escobar, C.A.; Priego-Capote, F.; Robles-Olvera, V.J.; Luque de Castro, M.D. Changes in the Composition of the Polar Fraction of Persian Lime (Citrus Latifolia) during Fruit Growth by LC–QTOF MS/MS Analysis. Food Chem. 2017, 234, 262–268. [Google Scholar] [CrossRef]
- Centonze, V.; Lippolis, V.; Cervellieri, S.; Damascelli, A.; Casiello, G.; Pascale, M.; Logrieco, A.F.; Longobardi, F. Discrimination of Geographical Origin of Oranges (Citrus Sinensis L. Osbeck) by Mass Spectrometry-Based Electronic Nose and Characterization of Volatile Compounds. Food Chem. 2019, 277, 25–30. [Google Scholar] [CrossRef]
- Stephan, N.; Halama, A.; Mathew, S.; Hayat, S.; Bhagwat, A.; Mathew, L.S.; Diboun, I.; Malek, J.; Suhre, K. A Comprehensive Metabolomic Data Set of Date Palm Fruit. Data Brief. 2018, 18, 1313–1321. [Google Scholar] [CrossRef]
- Punjab Economic Survey 2020–2021. Available online: https://esopb.gov.in/Static/PDF/EconomicSurvey-2020-21.pdf (accessed on 3 March 2022).
- Saini, M.K.; Capalash, N.; Kaur, C.; Singh, S.P. Targeted Metabolic Profiling Indicates Differences in Primary and Secondary Metabolites in Kinnow Mandarin (C. Nobilis × C. Deliciosa) from Different Climatic Conditions. J. Food Compos. Anal. 2019, 83, 103278. [Google Scholar] [CrossRef]
- Saini, M.K.; Capalash, N.; Kaur, C.; Singh, S.P. Comprehensive Metabolic Profiling to Decipher the Influence of Rootstocks on Fruit Juice Metabolome of Kinnow (C. Nobilis × C. Deliciosa). Sci. Hortic. 2019, 257, 108673. [Google Scholar] [CrossRef]
- Saini, M.K.; Capalash, N.; Varghese, E.; Kaur, C.; Singh, S.P. Quantitative Metabolomics Approach Reveals Dynamics of Primary Metabolites in ‘Kinnow’ Mandarin (C. Nobilis × C. Deliciosa) during Advanced Stages of Fruit Maturation under Contrasting Growing Climates. J. Hortic. Sci. Biotechnol. 2020, 95, 106–112. [Google Scholar] [CrossRef]
- Vrhovsek, U.; Masuero, D.; Gasperotti, M.; Franceschi, P.; Caputi, L.; Viola, R.; Mattivi, F. A Versatile Targeted Metabolomics Method for the Rapid Quantification of Multiple Classes of Phenolics in Fruits and Beverages. J. Agric. Food Chem. 2012, 60, 8831–8840. [Google Scholar] [CrossRef]
- Manners, G.D.; Breksa, A.P.; Schoch, T.K.; Hidalgo, M.B. Analysis of Bitter Limonoids in Citrus Juices by Atmospheric Pressure Chemical Ionization and Electrospray Ionization Liquid Chromatography-Mass Spectrometry. J. Agric. Food Chem. 2003, 51, 3709–3714. [Google Scholar] [CrossRef]
- Jayaprakasha, G.K.; Dandekar, D.V.; Tichy, S.E.; Patil, B.S. Simultaneous Separation and Identification of Limonoids from Citrus Using Liquid Chromatography-Collision-Induced Dissociation Mass Spectra. J. Sep. Sci. 2011, 34, 2–10. [Google Scholar] [CrossRef]
- Tripoli, E.; Guardia, M.L.; Giammanco, S.; Majo, D.D.; Giammanco, M. Citrus Flavonoids: Molecular Structure, Biological Activity and Nutritional Properties: A Review. Food Chem. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Omidbaigi, R.; Nasiri, M.F. Quantitative Distribution of Hesperidin in Citrus Species, during Fruit Maturation and Optimal Harvest Time. Nat. Prod. Radiance 2004, 3, 12–15. [Google Scholar]
- Benavente-García, O.; Castillo, J.; Marin, F.R.; Ortuño, A.; Del Río, J.A. Uses and Properties of Citrus Flavonoids. J. Agric. Food Chem. 1997, 45, 4505–4515. [Google Scholar] [CrossRef]
- Kawaii, S.; Tomono, Y.; Katase, E.; Ogawa, K.; Yano, M. Quantitation of Flavonoid Constituents in Citrus Fruits. J. Agric. Food Chem. 1999, 47, 3565–3571. [Google Scholar] [CrossRef]
- Khan, M.K.; Zill-E-Huma; Dangles, O. A Comprehensive Review on Flavanones, the Major Citrus Polyphenols. J. Food Compos. Anal. 2014, 33, 85–104. [Google Scholar] [CrossRef]
- Peterson, J.J.; Dwyer, J.T.; Beecher, G.R.; Bhagwat, S.A.; Gebhardt, S.E.; Haytowitz, D.B.; Holden, J.M. Flavanones in Oranges, Tangerines (Mandarins), Tangors, and Tangelos: A Compilation and Review of the Data from the Analytical Literature. J. Food Compos. Anal. 2006, 19, S66–S73. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Mahasneh, M.A.; Ereifej, K.; Al-Karaki, G.; Al-Duais, M.; Andrade, J.E.; Tranchant, C.C.; Kubow, S.; et al. Profiles of Free and Bound Phenolics Extracted from Citrus Fruits and Their Roles in Biological Systems: Content, and Antioxidant, Anti-Diabetic and Anti-Hypertensive Properties. Food Funct. 2017, 8, 3187–3197. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Liu, D.; Chen, J.; Ye, X.; Ma, Y.; Shi, J. Juice Components and Antioxidant Capacity of Citrus Varieties Cultivated in China. Food Chem. 2008, 106, 545–551. [Google Scholar] [CrossRef]
- Tounsi, M.S.; Wannes, W.A.; Ouerghemmi, I.; Jegham, S.; Njima, Y.B.; Hamdaoui, G.; Zemni, H.; Marzouk, B. Juice Components and Antioxidant Capacity of Four Tunisian Citrus Varieties. J. Sci. Food Agric. 2011, 91, 142–151. [Google Scholar] [CrossRef]
- Chaudhary, P.R.; Bang, H.; Jayaprakasha, G.K.; Patil, B.S. Variation in Key Flavonoid Biosynthetic Enzymes and Phytochemicals in ‘Rio Red’ Grapefruit (Citrus Paradisi Macf.) during Fruit Development. J. Agric. Food Chem. 2016, 64, 9022–9032. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, H.; Pang, Y.; Cheng, Y.; Deng, X.; Xu, J. Comparative Study of Flavonoid Production in Lycopene-Accumulated and Blonde-Flesh Sweet Oranges (Citrus Sinensis) during Fruit Development. Food Chem. 2015, 184, 238–246. [Google Scholar] [CrossRef]
- Del Río, J.A.; Fuster, M.D.; Gómez, P.; Porras, I.; García-Lidón, A.; Ortuño, A. Citrus Limon: A Source of Flavonoids of Pharmaceutical Interest. Food Chem. 2004, 84, 457–461. [Google Scholar] [CrossRef]
- Ortuño, A.; Reynaldo, I.; Fuster, M.D.; Botía, J.; Puig, D.G.; Sabater, F.; Lidón, A.G.; Porras, I.; Del Río, J. Citrus Cultivars with High Flavonoid Contents in the Fruits. Sci. Hortic. 1997, 68, 231–236. [Google Scholar] [CrossRef]
- Castillo, J.; Benavente, O.; del Rio, J.A. Naringin and Neohesperidin Levels during Development of Leaves, Flower Buds, and Fruits of Citrus Aurantium. Plant Physiol. 1992, 99, 67–73. [Google Scholar] [CrossRef]
- Moriguchi, T.; Kita, M.; Ogawa, K.; Tomono, Y.; Endo, T.; Omura, M. Flavonol Synthase Gene Expression during Citrus Fruit Development. Physiol. Plant. 2002, 114, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Agati, G.; Azzarello, E.; Pollastri, S.; Tattini, M. Flavonoids as Antioxidants in Plants: Location and Functional Significance. Plant. Sci. 2012, 196, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Crifò, T.; Puglisi, I.; Petrone, G.; Recupero, G.R.; Lo Piero, A.R. Expression Analysis in Response to Low Temperature Stress in Blood Oranges: Implication of the Flavonoid Biosynthetic Pathway. Gene 2011, 476, 1–9. [Google Scholar] [CrossRef]
- Wang, F.; Yu, X.; Liu, X.; Shen, W.; Zhu, S.; Zhao, X. Temporal and Spatial Variations on Accumulation of Nomilin and Limonin in the Pummelos. Plant. Physiol. Biochem. 2016, 106, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Baldwin, E.; McCollum, G.; Plotto, A.; Manthey, J.; Widmer, W.; Luzio, G.; Cameron, R. Changes in Volatile and Non-Volatile Flavor Chemicals of “Valencia” Orange Juice over the Harvest Seasons. Foods 2016, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, S.; Miyake, M. Biochemistry and Biological Functions of Citrus Limonoids. Food Rev. Int. 1996, 12, 413–435. [Google Scholar] [CrossRef]
- Yang, D.; Du, X.; Yang, Z.; Liang, Z.; Guo, Z.; Liu, Y. Transcriptomics, Proteomics, and Metabolomics to Reveal Mechanisms Underlying Plant Secondary Metabolism. Eng. Life Sci. 2014, 14, 456–466. [Google Scholar] [CrossRef]
- Besco, E.; Braccioli, E.; Vertuani, S.; Ziosi, P.; Brazzo, F.; Bruni, R.; Sacchetti, G.; Manfredini, S. The Use of Photochemiluminescence for the Measurement of the Integral Antioxidant Capacity of Baobab Products. Food Chem. 2007, 102, 1352–1356. [Google Scholar] [CrossRef]
- Zou, Z.; Xi, W.; Hu, Y.; Nie, C.; Zhou, Z. Antioxidant Activity of Citrus Fruits. Food Chem. 2016, 196, 885–896. [Google Scholar] [CrossRef]
- Yoo, K.-M.; Moon, B. Comparative Carotenoid Compositions during Maturation and Their Antioxidative Capacities of Three Citrus Varieties. Food Chem. 2016, 196, 544–549. [Google Scholar] [CrossRef]
- Almeida, I.M.C.; Barreira, J.C.M.; Oliveira, M.B.P.P.; Ferreira, I.C.F.R. Dietary Antioxidant Supplements: Benefits of Their Combined Use. Food Chem. Toxicol. 2011, 49, 3232–3237. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Climate (C) | Subtropical Arid (STA) | Subtropical Humid (STH) | S.E.M | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Harvest Maturity (M) | M1 | M2 | M3 | M4 | M5 | M6 | M1 | M2 | M3 | M4 | M5 | M6 | |
| Total Phenolics | 309.1 d | 274.1 fg | 375.5 b | 266.9 g | 264.7 g | 226.0 h | 310.1 d | 288.8 ef | 414.1 a | 327.4 c | 294.7 de | 267.2 g | 3.34 |
| Flavonones | 213.4 c | 180.0 fg | 259.5 b | 170.0 g | 177.9 fg | 152.3 h | 197.6 de | 181.1 fg | 275.6 a | 206.4 cd | 190.8 ef | 172.1 g | 2.56 |
| Hesperidin | 70.6 c | 62.3 d | 94.2 b | 48.9 fg | 43.9 gh | 39.8 h | 54.2 ef | 50.1 efg | 110.1 a | 72.5 c | 56.7 de | 54.3 ef | 1.53 |
| Naringin | 47.3 bc | 40.0 d | 60.11 a | 32.1 e | 30.1 e | 29.6 e | 52.2 b | 45.7 c | 50.7 bc | 31.5 e | 34.9 de | 33.9 e | 1.07 |
| Narirutin | 49.9 bc | 41.0 de | 62.2 a | 49.9 bc | 35.1 ef | 30.3 f | 46.1 cd | 46.5 cd | 56.1 ab | 56.5 ab | 54.4 b | 42.6 cde | 1.51 |
| Naringenin | 44.3 b | 41.2 b | 56.6 a | 53.1 a | 40.3 b | 39.9 b | 44.7 b | 38.3 b | 57.5 a | 45.1 b | 44.0 b | 40.4 b | 1.51 |
| Neoeriocitrin | 1.5 c | 1.33 c | 2.3 a | 2.16 a | 1.80 b | 1.8 b | 0.4 e | 0.5 e | 1.2 c | 0.8 d | 0.8 d | 0.8 d | 0.06 |
| Flavonols | 88.3 f | 86.7 f | 108.7 bc | 88.5 f | 78.3 g | 64.8 h | 105.1 cd | 100.7 de | 131.5 a | 113.9 b | 96.4 e | 87.0 f | 1.36 |
| Quercetin | 30.9 de | 35.6 d | 36.0 cd | 38.6 cd | 30.9 de | 30.5 de | 45.3 b | 43.0 b | 55.3 a | 53.0 a | 42.5 b | 43.9 b | 1.30 |
| Rutin hydrate | 41.9 cd | 39.9 d | 53.5 a | 51.2 ab | 39.4 d | 37.8 de | 46.2 bc | 46.4 bc | 54.5 a | 46.7 bc | 43.3 cd | 33.1 e | 1.20 |
| Nobiletin | 7.3 c | 3.9 g | 9.8 b | 2.2 i | 2.0 i | 1.4 j | 6.3 d | 4.6 f | 11.8 a | 5.7 e | 3.1 h | 3.0 h | 0.09 |
| Tangeretin | 3.2 b | 2.7 cd | 3.4 b | 1.5 e | 1.4 e | 1.0 f | 3.3 b | 2.8 c | 3.7 a | 3.3 b | 2.5 d | 2.5 d | 0.05 |
| Quercetin-3-O-galactoside | 3.3 bcd | 3.0 cde | 4.0 a | 3.5 b | 3.1 bcde | 2.9 de | 2.4 fg | 2.3 g | 4.0 a | 3.3 bc | 3.1 cde | 2.8 ef | 0.08 |
| Kaempferol | 1.5 c | 1.3 de | 1.7 b | 1.3 e | 1.2 f | 1.0 g | 1.5 c | 1.4 cd | 2.1 a | 1.7 b | 1.7 b | 1.5 c | 0.02 |
| Flavan-3-ols | 1.2 ab | 1.1 ab | 1.4 ab | 1.2 ab | 1.1 ab | 1.0 ab | 1.0 ab | 1.1 ab | 1.4 a | 1.3 ab | 1.2 ab | 0.9 b | 0.09 |
| Epigallocatechin | 1.2 ab | 1.1 ab | 1.4 ab | 1.2 ab | 1.1 ab | 1.0 ab | 1.0 ab | 1.1 ab | 1.4 a | 1.3 ab | 1.2 ab | 0.9 b | 0.09 |
| Flavones | 1.8 cd | 1.6 de | 1.9 bc | 1.6 de | 1.5 ef | 1.3 f | 1.9 bc | 1.8 c | 2.4 a | 2.1 b | 2.1 b | 1.9 c | 0.04 |
| Luteolin | 1.7 c | 1.5 d | 1.8 bc | 1.5 d | 1.4 e | 1.2 f | 1.8 c | 1.7 c | 2.2 a | 1.9 b | 1.9 b | 1.7 c | 0.02 |
| Genistein | 0.1 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a | 0.1 a | 0.2 a | 0.2 a | 0.2 a | 0.2 a | 0.02 |
| Benzoic and Cinnamic Acid Derivatives | 4.0 de | 4.3 cd | 3.7 de | 5.3 bc | 5.8 ab | 6.4 a | 4.1 de | 3.6 e | 3.6 e | 4.3 cd | 4.9 b | 5.9 ab | 0.16 |
| Sinapic acid | 2.1 d | 2.7 b | 1.9 d | 2.2 cd | 2.7 bc | 3.3 a | 1.8 de | 1.4 e | 1.4 e | 1.4 e | 1.5 e | 2.2 cd | 0.10 |
| Ferulic acid | 1.1 def | 0.8 f | 0.8 ef | 1.3 bcdef | 1.5 abcd | 1.7 abc | 1.4 abcde | 1.3 bcdef | 1.2 cdef | 1.7 abc | 1.9 a | 1.8 ab | 0.11 |
| Ellagic acid | 0.5 abcd | 0.5 cde | 0.6 ab | 0.5 cde | 0.4 ef | 0.4 f | 0.5 abcd | 0.6 abc | 0.4 ef | 0.5 de | 0.5 bcd | 0.6 a | 0.02 |
| Benzoic acid | 0.1 e | 0.1 e | 0.3 d | 1.1 a | 1.0 a | 0.8 b | 0.2 de | 0.3 d | 0.5 c | 0.5 c | 0.7 b | 1.1 a | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saini, M.K.; Capalash, N.; Varghese, E.; Kaur, C.; Singh, S.P. A Targeted Metabolomics Approach to Study Secondary Metabolites and Antioxidant Activity in ‘Kinnow Mandarin’ during Advanced Fruit Maturity. Foods 2022, 11, 1410. https://doi.org/10.3390/foods11101410
Saini MK, Capalash N, Varghese E, Kaur C, Singh SP. A Targeted Metabolomics Approach to Study Secondary Metabolites and Antioxidant Activity in ‘Kinnow Mandarin’ during Advanced Fruit Maturity. Foods. 2022; 11(10):1410. https://doi.org/10.3390/foods11101410
Chicago/Turabian StyleSaini, Manpreet Kaur, Neena Capalash, Eldho Varghese, Charanjit Kaur, and Sukhvinder Pal Singh. 2022. "A Targeted Metabolomics Approach to Study Secondary Metabolites and Antioxidant Activity in ‘Kinnow Mandarin’ during Advanced Fruit Maturity" Foods 11, no. 10: 1410. https://doi.org/10.3390/foods11101410
APA StyleSaini, M. K., Capalash, N., Varghese, E., Kaur, C., & Singh, S. P. (2022). A Targeted Metabolomics Approach to Study Secondary Metabolites and Antioxidant Activity in ‘Kinnow Mandarin’ during Advanced Fruit Maturity. Foods, 11(10), 1410. https://doi.org/10.3390/foods11101410