
Glucosinolates as Markers of the Origin and Harvesting Period for Discrimination of Bee Pollen by UPLC-MS/MS


 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Materials
2.2. Standards
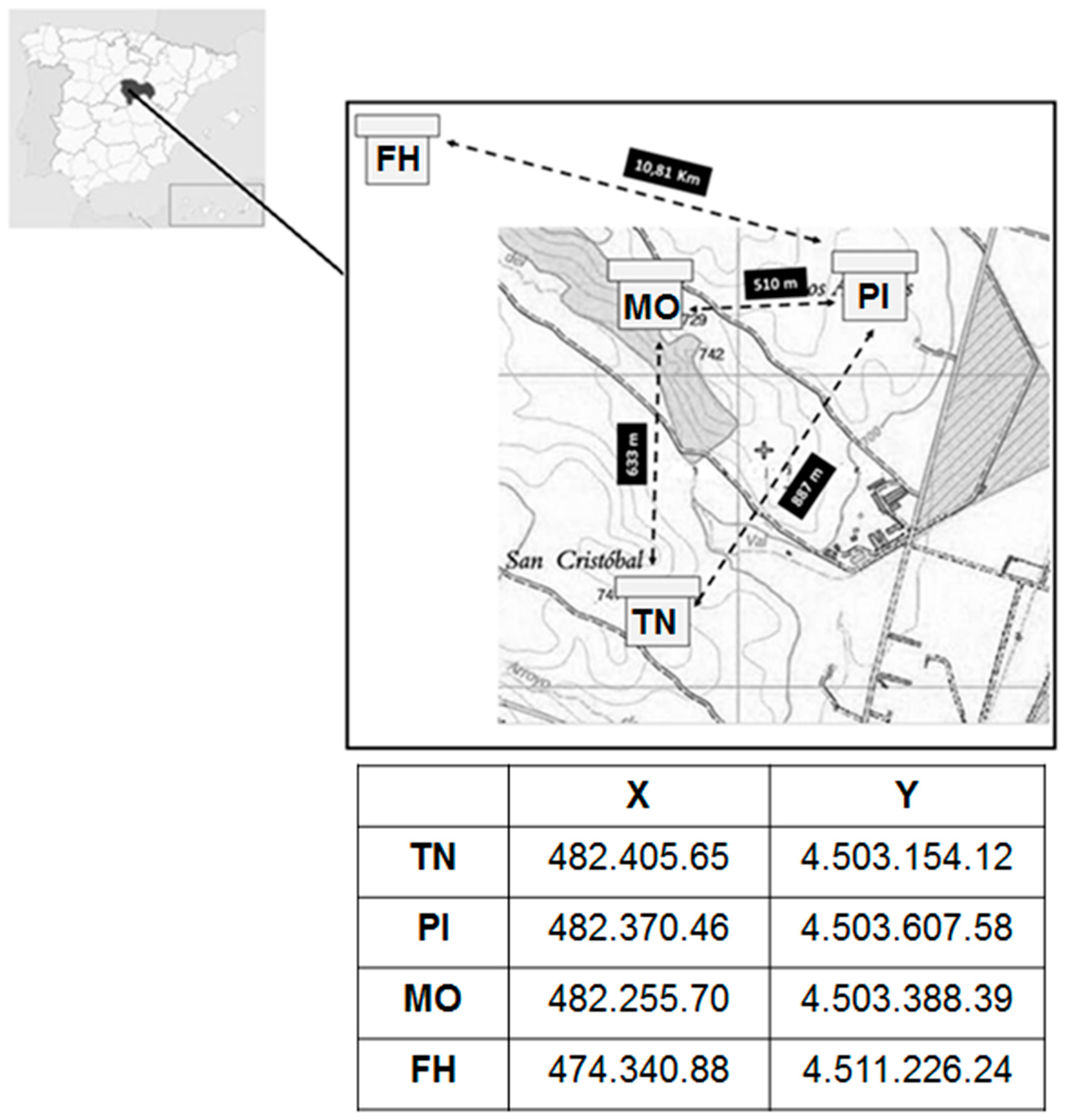
2.3. Sample Procurement and Treatment
2.3.1. Samples
2.3.2. Palynological Analysis
2.3.3. Sample Treatment
2.4. UPLC-MS/MS System
2.5. Canonical Discriminant Analysis
3. Results and Discussion
3.1. Glucosinolate Content
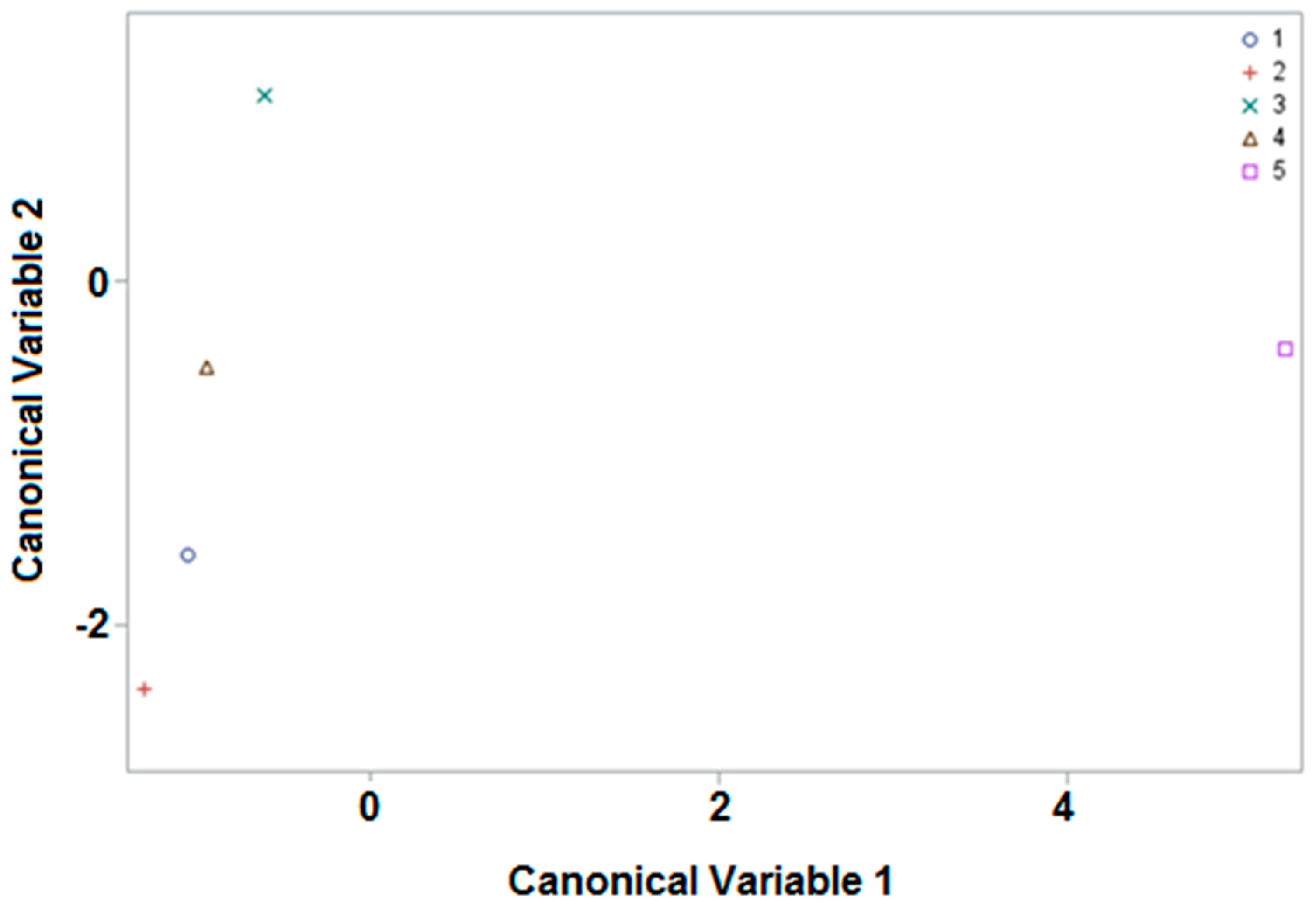
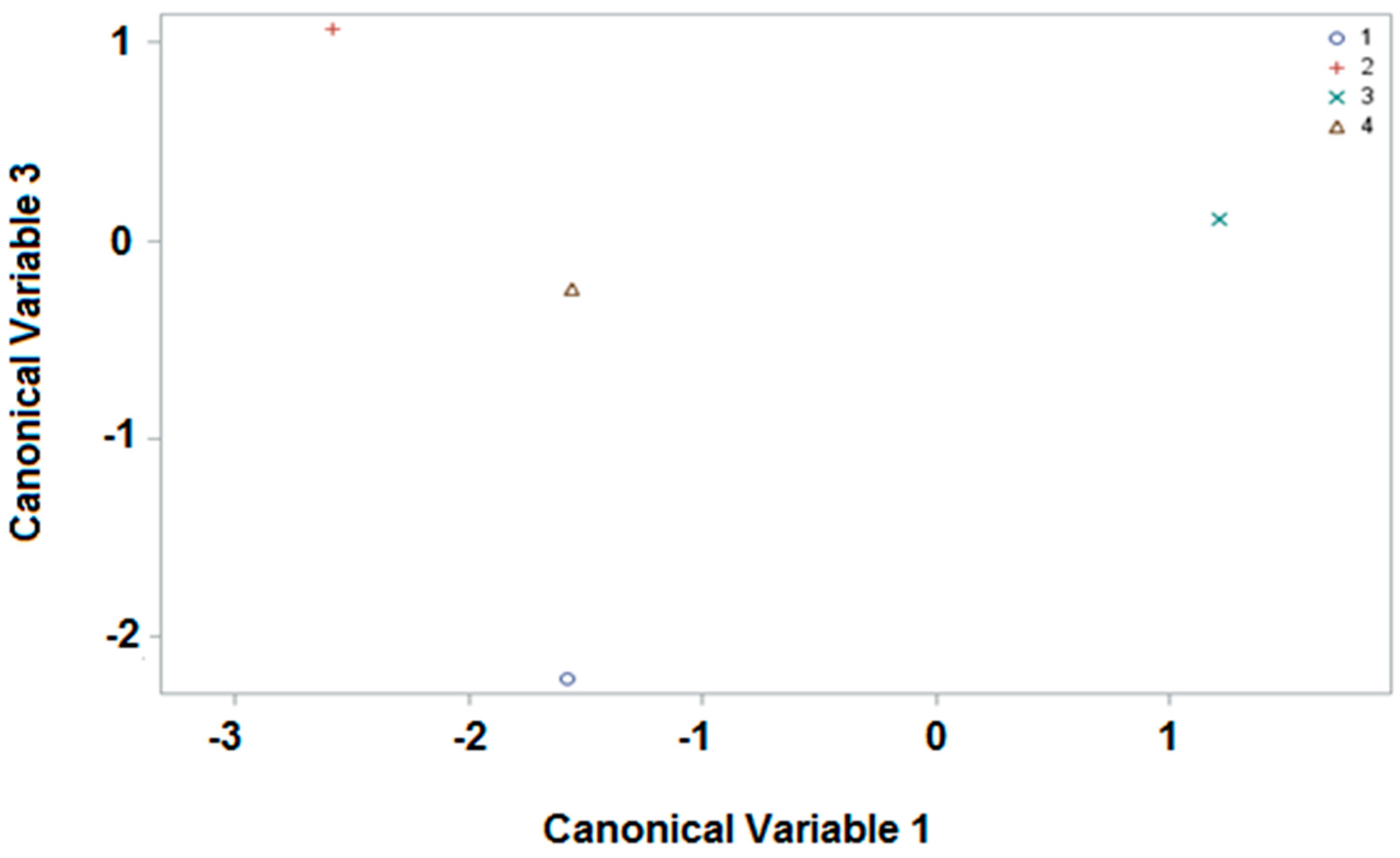
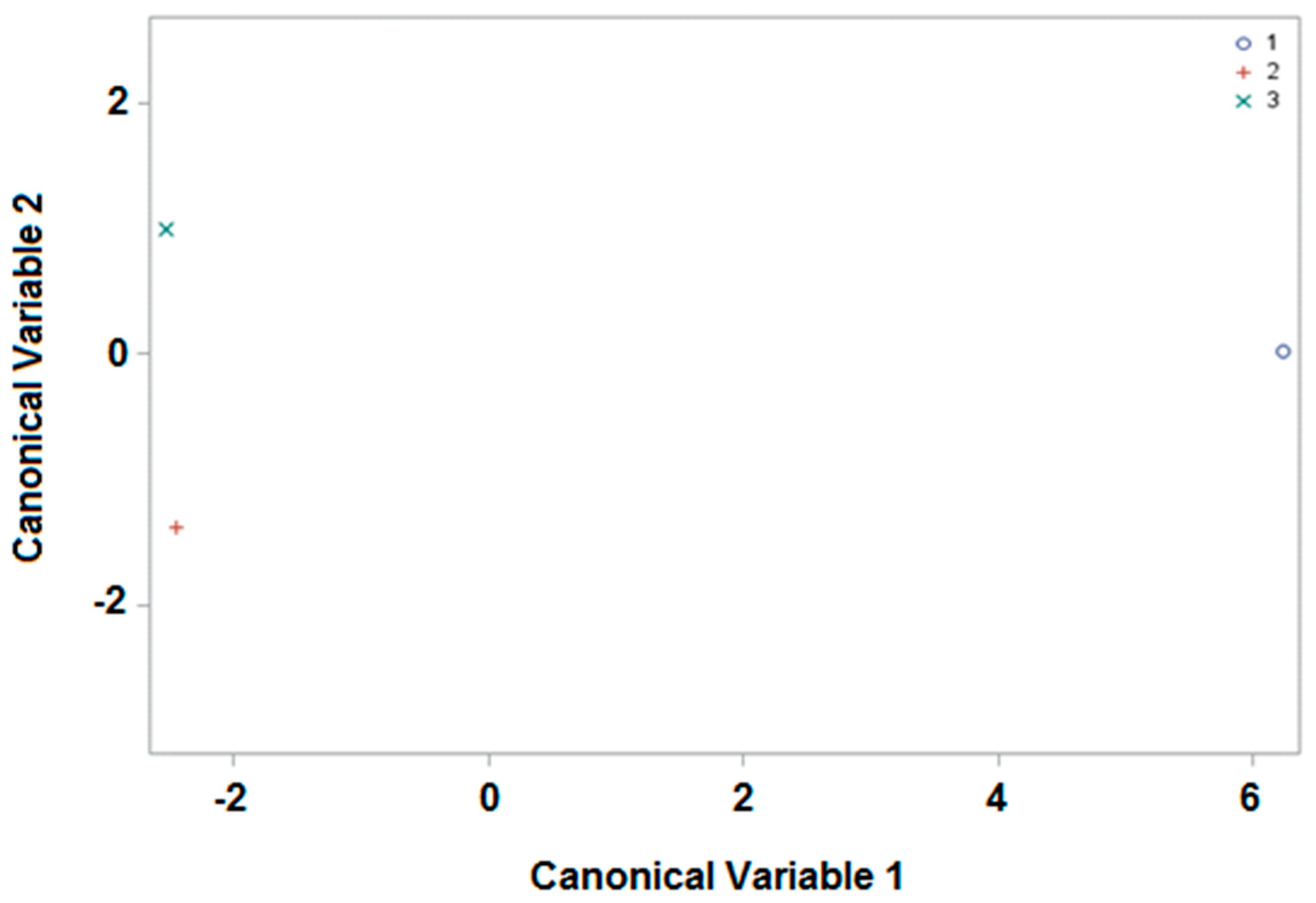
3.2. Canonical Discrimant Analysis
3.2.1. Origin
All Samples
Apiary Samples
3.2.2. Harvesting Period
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Z.; Ren, P.; Wu, Y.; He, Q. Recent advances in analytical techniques for the detection of adulteration and authenticity of bee products—A review. Food Addit. Contam. Part A 2021, 38, 533–549. [Google Scholar] [CrossRef] [PubMed]
- Trifković, J.; Andrić, F.; Ristivojević, P.; Guzelmeric, E.; Yesilada, E. Analytical methods in tracing honey authenticity. J. AOAC Int. 2017, 100, 827–839. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.Y.; Zhong, M.C.; Hao, N.; Wang, T.; Wang, H.T. Botanical origin authenticity control of pine pollen food products using multiplex species-specific PCR method. Food Anal. Methods 2022, 15, 421–427. [Google Scholar] [CrossRef]
- Ares, A.M.; Valverde, S.; Bernal, J.L.; Nozal, M.J.; Bernal, J. Extraction and determination of bioactive compounds from bee pollen. J. Pharm. Biomed. Anal. 2018, 147, 110–124. [Google Scholar] [CrossRef]
- Gonçalves, A.C.; Lahlou, R.A.; Alves, G.; Garcia-Viguera, C.; Moreno, D.A.; Silva, L.R. Potential activity of Abrantes pollen extract: Biochemical and cellular model studies. Foods 2021, 10, 2894. [Google Scholar] [CrossRef] [PubMed]
- Prđun, S.; Svečnjak, L.; Valentić, M.; Marijanović, Z.; Jerković, I. Characterization of Bee Pollen: Physico-Chemical Properties, Headspace Composition and FTIR Spectral Profiles. Foods 2021, 10, 2103. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.J.S.; Estevinho, L.M.; Pereira, A.P.; Sousa, J.M.C.; Anjos, O. Computational intelligence applied to discriminate bee pollen quality and botanical origin. Food Chem. 2018, 267, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sattler, J.A.G.; de Melo, I.L.P.; Granato, D.; Araújo, E.; de Freitas, A.S.; Barth, O.M.; Sattler, A.; Almeida-Muradian, L.B. Impact of origin on bioactive compounds and nutritional composition of bee pollen from southern Brazil: A screening study. Food Res. Int. 2015, 77, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Taha, E.K.A. Chemical composition and amounts of mineral elements in honeybee-collected pollen in relation to botanical origin. J. Apic. Sci. 2015, 59, 75–81. [Google Scholar] [CrossRef]
- Kaškonienė, V.; Ruočkuvienė, G.; Kaškonas, P.; Akuneca, I.; Maruška, A. Chemometric analysis of bee pollen based on volatile and phenolic compound compositions and antioxidant properties. Food Anal. Methods 2015, 8, 1150–1163. [Google Scholar] [CrossRef]
- Zhou, J.; Qi, Y.; Ritho, J.; Zhang, Y.; Zheng, X.; Wu, L.; Li, Y.; Sun, L. Flavonoid glycosides as floral origin markers to discriminate of unifloral bee pollen by LC-MS/MS. Food Control 2015, 57, 54–61. [Google Scholar] [CrossRef]
- Hwang, I.M.; Park, B.; Dang, Y.M.; Kim, S.Y.; Seo, H.Y. Simultaneous direct determination of 15 glucosinolates in eight Brassica species by UHPLC-Q-Orbitrap-MS. Food Chem. 2019, 282, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Kim, J.S. Anti-carcinogenic glucosinolates in cruciferous vegetables and their antagonistic effects on prevention of cancers. Molecules 2018, 23, 2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ares, A.M.; Nozal, M.J.; Bernal, J. Extraction, chemical characterization and biological activity determination of broccoli health promoting compounds. J. Chromatogr. A 2013, 1313, 78–95. [Google Scholar] [CrossRef]
- Dong, M.; Tian, Z.; Ma, Y.; Yang, Z.; Ma, Z.; Wang, X.; Li, Y.; Jiang, H. Rapid screening and characterization of glucosinolates in 25 Brassicaceae tissues by UHPLC-Q-exactive orbitrap-MS. Food Chem. 2021, 365, 130493. [Google Scholar] [CrossRef] [PubMed]
- Truchado, P.; Tourn, E.; Gallez, L.M.; Moreno, D.A.; Ferreres, F.; Tomás-Barberán, F.A. Identification of botanical biomarkers in Argentinean Diplotaxis honeys: Flavonoids and glucosinolates. J. Agric. Food Chem. 2010, 58, 12678–12685. [Google Scholar] [CrossRef]
- Ares, A.M.; Redondo, M.; Tapia, J.; González-Porto, A.V.; Higes, M.; Martín-Hernández, R.; Bernal, J. Differentiation of bee pollen samples according to their intact-glucosinolate content using canonical discriminant analysis. LWT-Food Sci. Technol. 2020, 129, 109559. [Google Scholar] [CrossRef]
- Bernal, J.; González, D.; Valverde, S.; Toribio, L.; Ares, A.M. Improved separation of intact glucosinolates in bee pollen by using ultra-high-performance liquid chromatography coupled to quadrupole time-of-flight mass spectrometry. Food Anal. Methods 2019, 12, 1170–1178. [Google Scholar] [CrossRef]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; Wiley: New York, NY, USA, 1989. [Google Scholar]
- Cepero, A.; Ravoet, J.; Gómez-Moracho, T.; Bernal, J.L.; Del Nozal, M.J.; Bartolomé, C.; Maside, X.; Meana, A.; González-Porto, A.V.; de Graaf, D.; et al. Holistic screening of collapsing honey bee colonies in Spain: A case study. BMC Res. Notes 2014, 7, 649. [Google Scholar] [CrossRef] [Green Version]
- Ferrauto, G.; Pavone, P. Palynological, physico-chemical and organoleptic characteristics of carob tree (Ceratonia siliqua L.) honey from Sicily. Int. J. Food Sci. 2013, 48, 1596–1602. [Google Scholar] [CrossRef]
- Escuredo, O.; Silva, L.R.; Valentão, P.; Seijo, M.C.; Andrade, P.B. Assessing Rubus honey value: Pollen and phenolic compounds content and antibacterial capacity. Food Chem. 2012, 130, 671–678. [Google Scholar] [CrossRef]
- Ares, A.M.; Nozal, M.J.; Bernal, J. Development and validation of a liquid chromatography-tandem mass spectrometry method to determine intact glucosinolates in bee pollen. J. Chromatogr. B 2015, 1000, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Glauser, G.; Schweizer, F.; Turlings, T.C.J.; Reymond, P. Rapid profiling of intact glucosinolates in Arabidopsis leaves by UHPLC- QTOFMS using a charged surface hybrid column. Phytochem. Anal. 2012, 23, 520–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobson, J.D. Applied Multivariate Data Analysis. Regression and Experimental Design, 1st ed.; Springer: New York, NY, USA, 1991. [Google Scholar]
- Dungey, S.G.; Sang, J.P.; Rothnie, N.E.; Palmer, M.V.; Burke, D.G.; Knox, R.B.; Williams, E.; Hilliard, E.; Salisbury, P. Glucosinolates in the pollen of rapeseed and Indian mustard. Phytochemistry 1988, 27, 815–817. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name (Abbreviation) | Molecular Weight | Ions (m/z) | LOD (µg/kg) | LOQ (µg/kg) |
|---|---|---|---|---|
| Glucoiberin (GIB) | 423 | 422.0044 A,QC, 96.9540 B,C | 18 | 60 |
| Progoitrin (PRO) | 389 | 388.0203 A,QC, 96.9543B,C | 18 | 60 |
| Sinigrin (SIN) | 359 | 358.0120 A,QC, 96.9544 B,C | 16 | 52 |
| Epiprogoitrin (EPI) | 389 | 388.0283 A,QC, 96.9539 B,C | 12 | 38 |
| Glucoraphanin (GRA) | 437 | 436.0291 A,QC, 96.9537 B,C | 14 | 44 |
| Gluconapin (GNA) | 373 | 372.0269 A,QC, 96.9542 B,C | 7 | 23 |
| Glucoalyssin (ALY) | 451 | 450.0500 A,QC, 96.9541 B,C | 16 | 55 |
| 4-hydroxyglucobrassicin (4-OH) | 464 | 463.0337 A,QC, 96.9540 B,C | 25 | 80 |
| Glucobrassicanapin (GBN) | 387 | 386.0379 A,QC, 96.9544 B,C | 8 | 26 |
| Glucotropaeolin (GTL) | 409 | 408.0224 A,QC, 96.9542 B,C | 7 | 21 |
| Glucoerucin (GER) | 421 | 420.0400 A,QC, 96.9536 B,C | 10 | 32 |
| Glucobrassicin (GBC) | 448 | 447.0348 A,QC, 96.9542 B,C | 9 | 34 |
| Gluconasturtiin (NAS) | 423 | 422.0401 A,QC, 96.9543 B,C | 6 | 19 |
| 4-Metoxyglucobrassicin (4-ME) | 478 | 477.0403 A,QC, 96.9538 B,C | 28 | 88 |
| Neoglucobrassicin (NEO) | 478 | 477.0440 A,QC, 96.9540 B,C | 8 | 25 |
| Sample | Major Taxon | Harvesting Period |
|---|---|---|
| MO-1 | Brassica t. + MF | April–May |
| MO-2 | Brassica t. | April–May |
| MO-3 | Quercus | April–May |
| MO-4 | Brassica t.+ MF | April–May |
| MO-5 | Papaver + Rubus | June |
| MO-6 | MF | June |
| MO-7 | Papaver + MF | June |
| MO-8 | MF | June |
| MO-9 | Papaver + Retama t. | June |
| MO-10 | MF | July–August |
| MO-11 | Rosa t. | July–August |
| MO-12 | MF | July–August |
| PI-1 | Brassica t. | April–May |
| PI-2 | Brassica t. | April–May |
| PI-3 | Brassica t. | April–May |
| PI-4 | Brassica t. | April–May |
| PI-5 | Brassica t. | April–May |
| PI-6 | Brassica t. | April–May |
| PI-7 | Brassica t. | April–May |
| PI-8 | Brassica t. | April–May |
| PI-9 | Brassica t. | April–May |
| PI-10 | Quercus ilex t. | April–May |
| PI-11 | Brassica t. | April–May |
| PI-12 | Brassica t. | April–May |
| PI-13 | Brassica t. | April–May |
| PI-14 | MF | June |
| PI-15 | MF | June |
| PI-16 | Papaver + Rosa t. | June |
| PI-17 | Teucrium + Rosa t. | June |
| PI-18 | MF | June |
| PI-19 | MF | June |
| PI-20 | Rubus | June |
| PI-21 | Papaver + Rosaceae | June |
| PI-22 | Reseda + Retama t. | June |
| PI-23 | Reseda | June |
| PI-24 | Papaver + Retama t. | June |
| PI-25 | Cytisus t. | June |
| PI-26 | MF | June |
| PI-27 | MF | June |
| PI-28 | MF | June |
| PI-29 | Papaver + Rosa t. | June |
| PI-30 | MF | June |
| PI-31 | MF | June |
| PI-32 | MF | July–August |
| PI-33 | MF | July–August |
| PI-34 | MF | July–August |
| PI-35 | Rosa t. | July–August |
| PI-36 | MF | July–August |
| PI-37 | MF | July–August |
| PI-38 | Rosa t. | July–August |
| PI-39 | MF | July–August |
| PI-40 | Retama t. | July–August |
| PI-41 | MF | July–August |
| PI-42 | MF | July–August |
| PI-43 | MF | July–August |
| PI-44 | Rubus | July–August |
| PI-45 | MF | July–August |
| TN-1 | MF | June |
| TN-2 | MF | June |
| TN-3 | MF | June |
| TN-4 | MF | June |
| TN-5 | MF | June |
| TN-6 | MF | July–August |
| TN-7 | MF | July–August |
| TN-8 | Rosa t | July–August |
| FH-1 | Brassica t. | April–May |
| FH-2 | MF | April–May |
| FH-3 | Brassica t. | April–May |
| FH-4 | Vicia t. | June |
| FH-5 | Rosaceae | June |
| FH-6 | Rosa t. | July–August |
| FH-7 | Rosa t. | July–August |
| GSL | Frequency A (%) | Concentration Range * (µg/kg; Dry Weight) |
|---|---|---|
| GIB | 0 | <LOD |
| PRO | 33 | 130–6690 |
| SIN | 33 | 84–2721 |
| EPI | 18 | 41–55 |
| GRA | 26 | 47–1172 |
| GNA | 56 | 27–1354 |
| ALY | 29 | 143–7916 |
| 4-OH | 32 | 105–3370 |
| GBN | 69 | 31–8469 |
| GTL | 11 | 22–76 |
| GER | 26 | 34–152 |
| GBC | 26 | 39–1467 |
| NAS | 47 | 19–9936 |
| 4-ME | 26 | 109–1664 |
| NEO | 61 | 29–5562 |
| GSL | Frequency A (%) | Concentration Range * (µg/kg; Dry Weight) |
|---|---|---|
| GIB | 0 | <LOD |
| PRO | 0 | <LOD |
| SIN | 9 | 92 |
| EPI | 36 | 57–557 |
| GRA | 36 | 69–249 |
| GNA | 91 | 49–1216 |
| ALY | 9 | <LOQ |
| 4-OH | 100 | 639–4193 |
| GBN | 27 | <LOQ |
| GTL | 64 | 23–1593 |
| GER | 0 | <LOD |
| GBC | 27 | 73–419 |
| NAS | 45 | 23–623 |
| 4-ME | 27 | <LOQ |
| NEO | 18 | <LOQ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ares, A.M.; Tapia, J.A.; González-Porto, A.V.; Higes, M.; Martín-Hernández, R.; Bernal, J. Glucosinolates as Markers of the Origin and Harvesting Period for Discrimination of Bee Pollen by UPLC-MS/MS. Foods 2022, 11, 1446. https://doi.org/10.3390/foods11101446
Ares AM, Tapia JA, González-Porto AV, Higes M, Martín-Hernández R, Bernal J. Glucosinolates as Markers of the Origin and Harvesting Period for Discrimination of Bee Pollen by UPLC-MS/MS. Foods. 2022; 11(10):1446. https://doi.org/10.3390/foods11101446
Chicago/Turabian StyleAres, Ana M., Jesús A. Tapia, Amelia V. González-Porto, Mariano Higes, Raquel Martín-Hernández, and José Bernal. 2022. "Glucosinolates as Markers of the Origin and Harvesting Period for Discrimination of Bee Pollen by UPLC-MS/MS" Foods 11, no. 10: 1446. https://doi.org/10.3390/foods11101446
APA StyleAres, A. M., Tapia, J. A., González-Porto, A. V., Higes, M., Martín-Hernández, R., & Bernal, J. (2022). Glucosinolates as Markers of the Origin and Harvesting Period for Discrimination of Bee Pollen by UPLC-MS/MS. Foods, 11(10), 1446. https://doi.org/10.3390/foods11101446