
Spoilage Potential of Contaminating Yeast Species Kluyveromyces marxianus, Pichia kudriavzevii and Torulaspora delbrueckii during Cold Storage of Skyr

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Reagents
2.2. Sequencing of 26S rRNA Gene and 5.8S-ITS Region
2.3. Pulsed-Field Gel Electrophoresis
2.4. Yeast Propagation in Skyr
2.5. Measurements of Viable Counts, pH and Carbon Dioxide
2.6. GC-MS Analysis of the Volatile Compounds and Data Processing
2.7. High Performance Liquid Chromatography
2.8. Phenotypic Tests
2.9. Statistical Analysis
2.10. Nucleotide Sequence Accession Number
3. Results
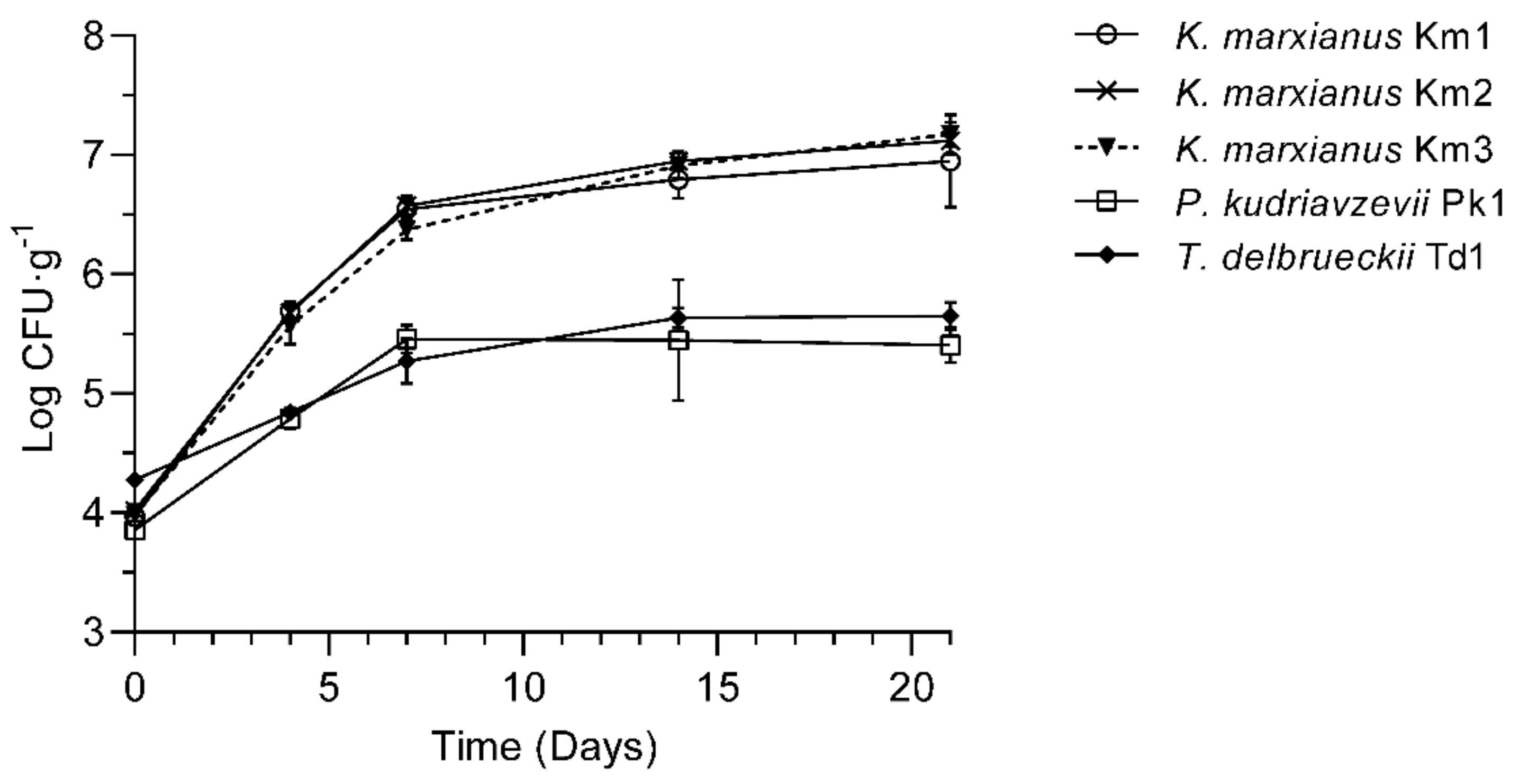
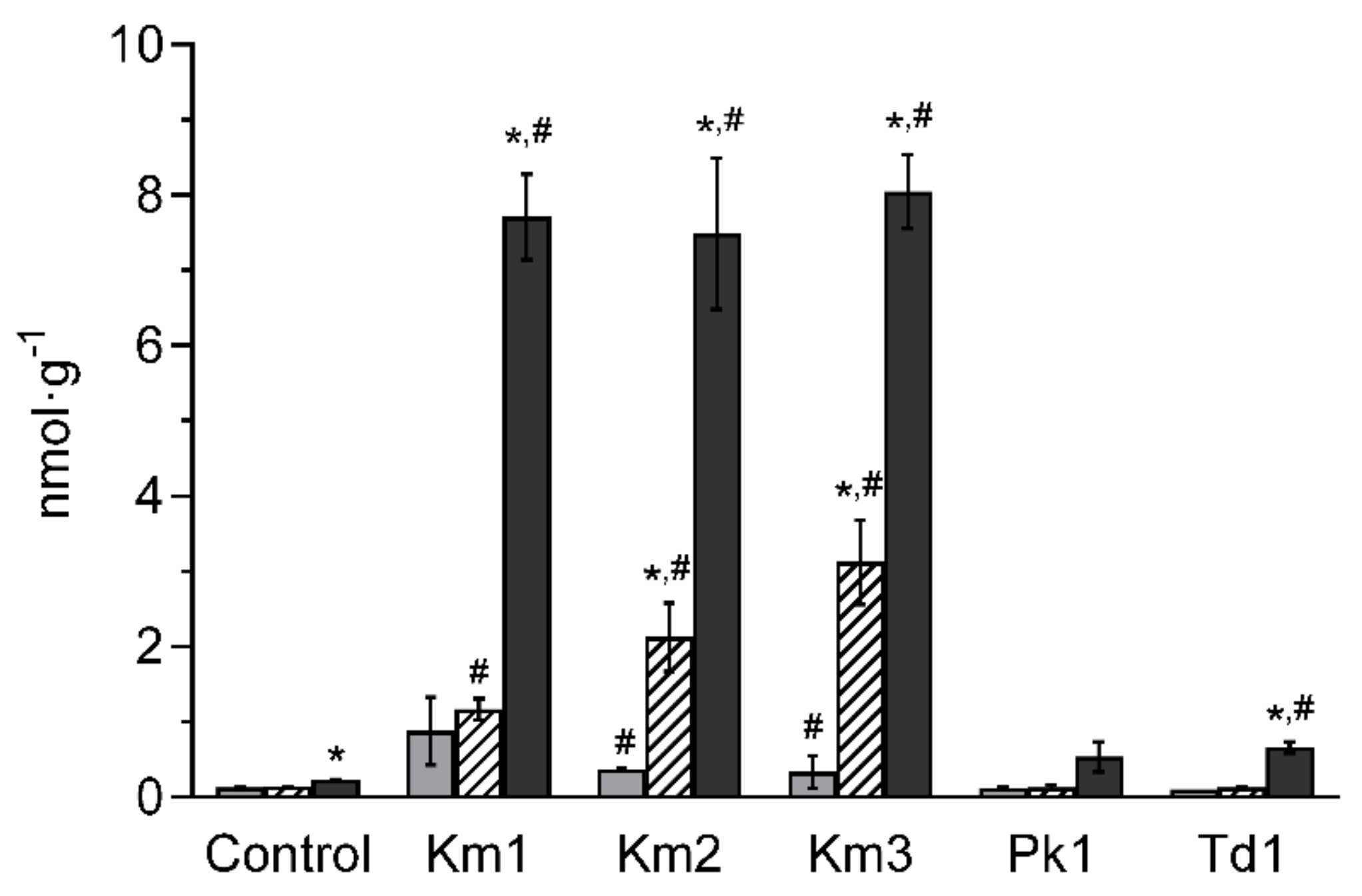
3.1. Yeasts Identification and Growth
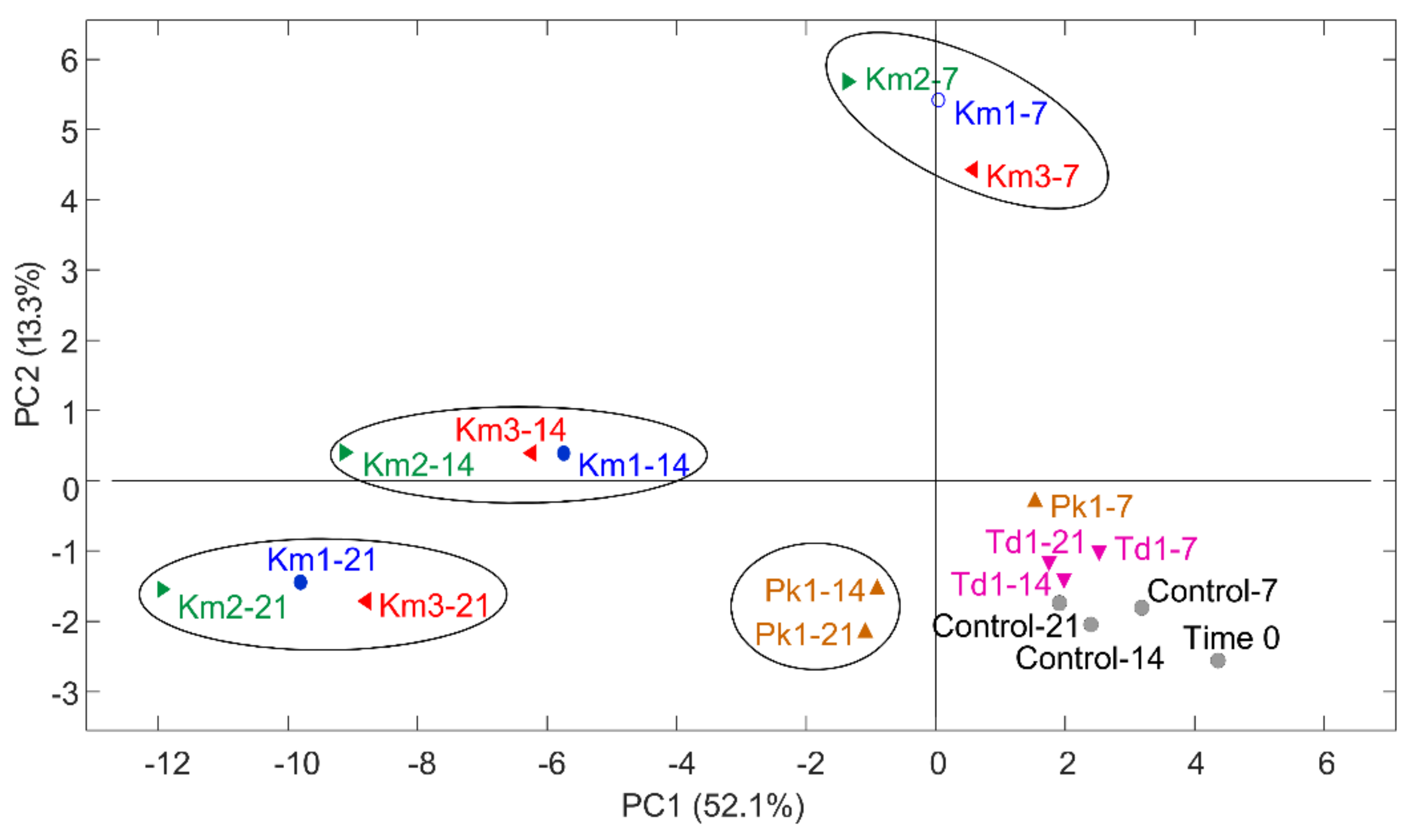
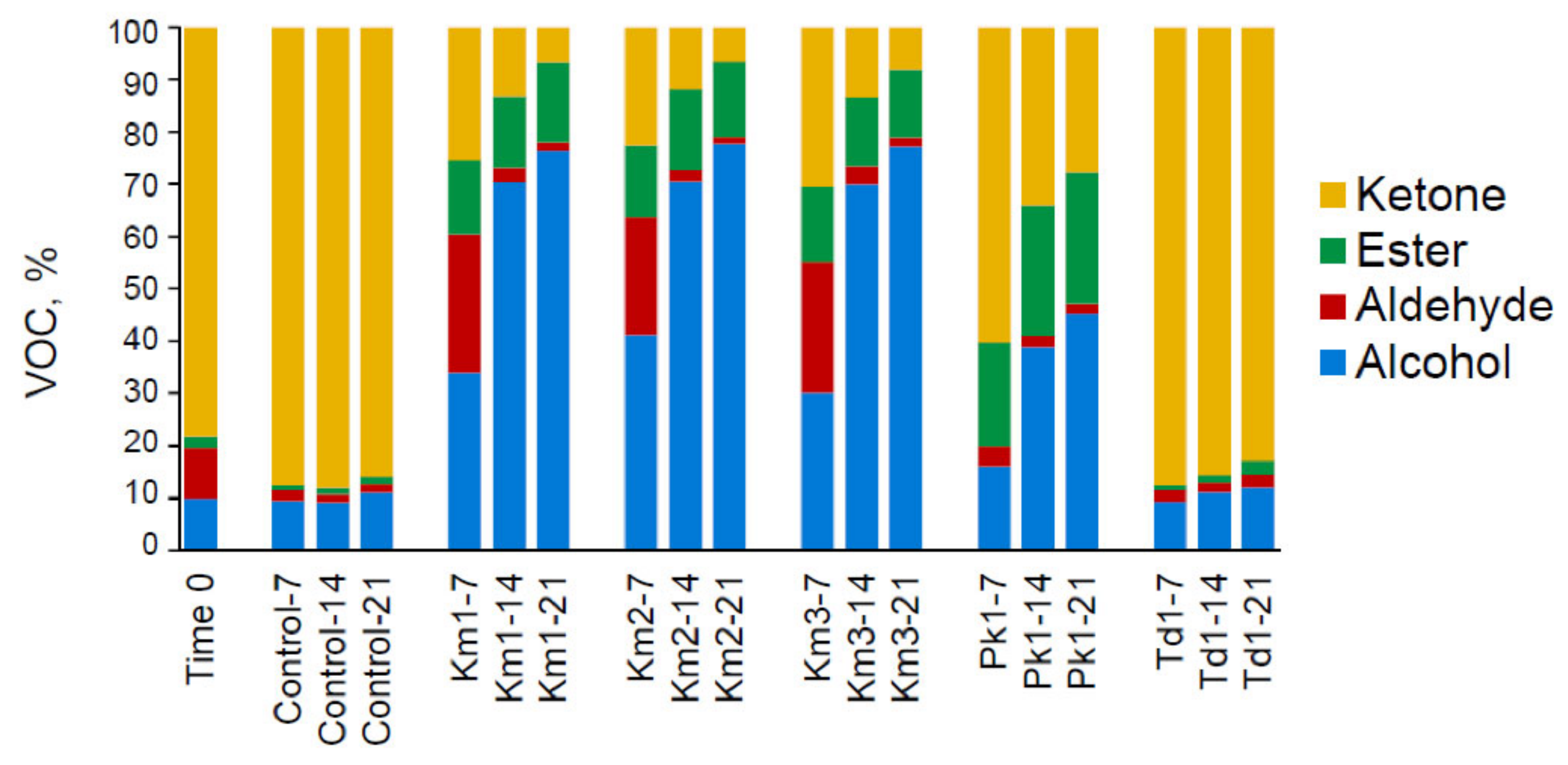
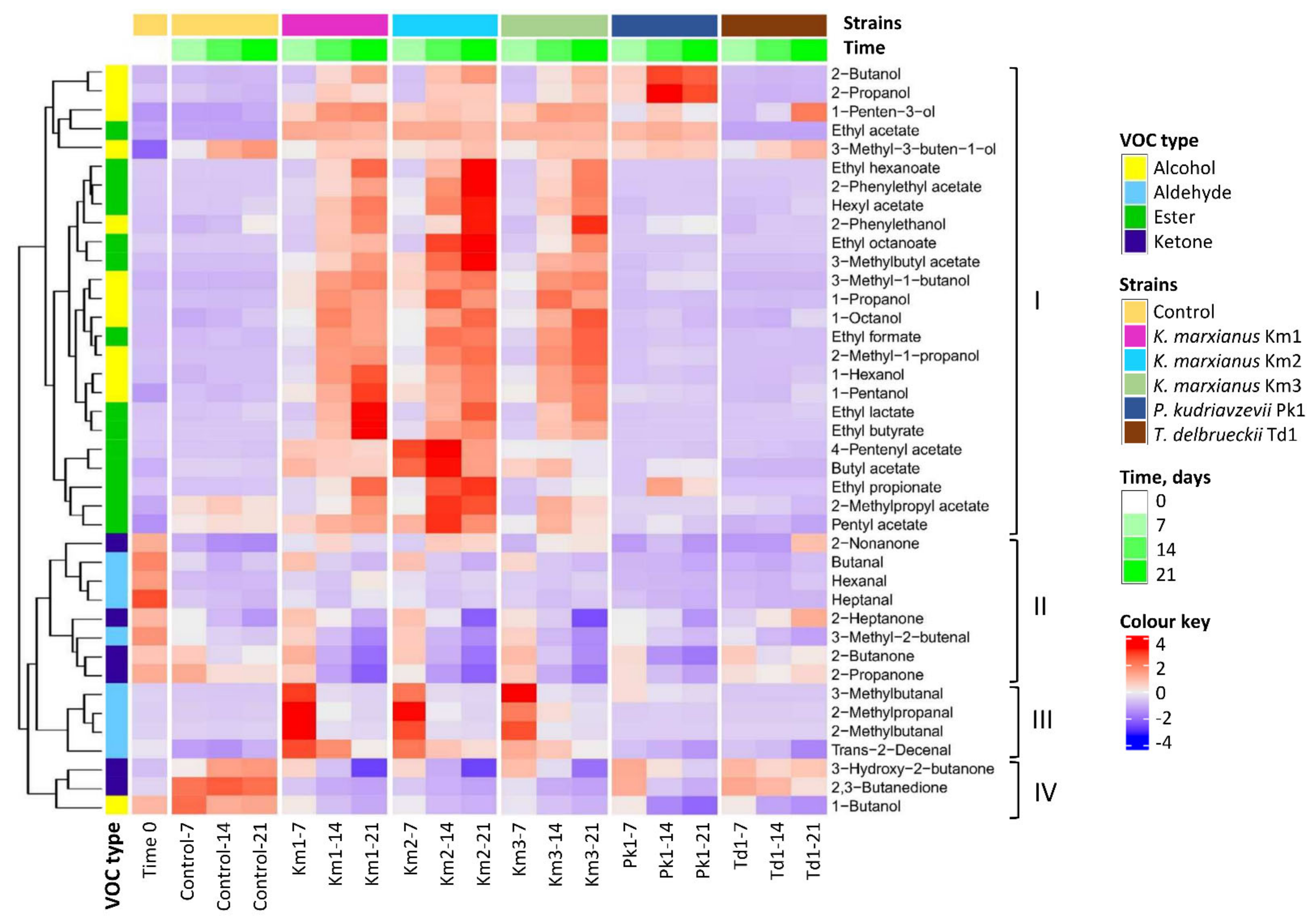
3.2. Production of Volatile Compounds
3.3. Utilization of Lactose and Galactose
3.4. Production of Organic Acids, Ethanol and Carbon Dioxide
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arla Foods Ingredients. Skyr is predicted a promising future in Europe. Int. Dairy Mag. 2020, 15, 1–36. [Google Scholar]
- Gudmundsson, G.; Kristbergsson, K. Modernization of skyr processing: Icelandic acid-curd soft cheese. In Modernization of Traditional Food Processes and Products, 1st ed.; McElhatton, A., el Idrissi, M.M., Eds.; Springer: New York, NY, USA, 2016; pp. 45–53. [Google Scholar] [CrossRef]
- Valsdóttir, Þ.; Sveinsson, Þ.E. Uniqueness of Traditional Skyr; Matís: Reykjavík, Iceland, 2011. [Google Scholar]
- Chen, C.; Zhao, S.; Hao, G.; Yu, H.; Tian, H.; Zhao, G. Role of lactic acid bacteria on the yogurt flavour: A review. Int. J. Food Prop. 2017, 20, S316–S330. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H. Volatile flavor compounds in yogurt: A review. Crit. Rev. Food Sci. Nutr. 2010, 50, 938–950. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Yu, B.; Yu, H.; Chen, C. Evaluation of the synergistic olfactory effects of diacetyl, acetaldehyde, and acetoin in a yogurt matrix using odor threshold, aroma intensity, and electronic nose analyses. J. Dairy Sci. 2020, 103, 7957–7967. [Google Scholar] [CrossRef]
- Miyaji, K.; Kuwano, Y.; Murakami, Y.; Hirata, S.; Imayoshi, Y.; Maruyama, H.; Koizumi, R.; Inoue, H.; Azuma, N. Off-flavors generated during long-term ambient storage of pasteurized drinking yogurt from skim milk. Biosci. Biotechnol. Biochem. 2020, 85, 391–400. [Google Scholar] [CrossRef]
- Garnier, L.; Valence, F.; Pawtowski, A.; Auhustsinava-Galerne, L.; Frotté, N.; Baroncelli, R.; Deniel, F.; Coton, E.; Mounier, J. Diversity of spoilage fungi associated with various French dairy products. Int. J. Food Microbiol. 2017, 241, 191–197. [Google Scholar] [CrossRef]
- Awasti, N.; Anand, S. The role of yeast and molds in dairy industry: An update. In Dairy Processing: Advanced Research to Applications, 1st ed.; Minj, J., Sudhakaran, A.V., Kumari, A., Eds.; Springer: Singapore, 2020; pp. 243–262. [Google Scholar] [CrossRef]
- Garnier, L.; Valence, F.; Mounier, J. Diversity and Control of Spoilage Fungi in Dairy Products: An Update. Microorganisms 2017, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Geronikou, A.; Srimahaeak, T.; Rantsiou, K.; Triantafillidis, G.; Larsen, N.; Jespersen, L. Occurrence of Yeasts in White-Brined Cheeses: Methodologies for Identification, Spoilage Potential and Good Manufacturing Practices. Front. Microbiol. 2020, 11, 582778. [Google Scholar] [CrossRef]
- Karim, A.; Gerliani, N.; Aïder, M. Kluyveromyces marxianus: An emerging yeast cell factory for applications in food and biotechnology. Int. J. Food Microbiol. 2020, 333, 108818. [Google Scholar] [CrossRef]
- Caballero, R.; Olguín, P.; Cruz-Guerrero, A.; Gallardo, F.; García-Garibay, M.; Gómez-Ruiz, L. Evaluation of Kluyveromyces marxianus as baker’s yeast. Food Res. Int. 1995, 28, 37–41. [Google Scholar] [CrossRef]
- Struyf, N.; Laurent, J.; Verspreet, J.; Verstrepen, K.J.; Courtin, C.M. Saccharomyces cerevisiae and Kluyveromyces marxianus Cocultures Allow Reduction of Fermentable Oligo-, Di-, and Monosaccharides and Polyols Levels in Whole Wheat Bread. J. Agric. Food Chem. 2017, 65, 8704–8713. [Google Scholar] [CrossRef] [Green Version]
- Sá, A.G.A.; Meneses, A.; de Araújo, P.H.H.; Oliveira, D. A review on enzymatic synthesis of aromatic esters used as flavor ingredients for food, cosmetics and pharmaceuticals industries. Trends Food Sci. Technol. 2017, 69, 95–105. [Google Scholar] [CrossRef]
- Mayoral, M.B.; Martín, R.; Sanz, A.; Hernández, P.E.; González, I.; García, T. Detection of Kluyveromyces marxianus and other spoilage yeasts in yoghurt using a PCR-culture technique. Int. J. Food Microbiol. 2005, 105, 27–34. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast Spoilage of Foods and Beverages. In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2011; pp. 53–63. [Google Scholar] [CrossRef]
- Fonseca, G.G.; Heinzle, E.; Wittmann, C.; Gombert, A.K. The yeast Kluyveromyces marxianus and its biotechnological potential. Appl. Microbiol. Biotechnol. 2008, 79, 339–354. [Google Scholar] [CrossRef]
- Šalomskienė, J.; Mačionienė, I. The influence of contamination yoghurt, quark and semi-hard cheese by yeasts on their sensory properties. Vet. Med. Zoot 2009, 48, 72–76. [Google Scholar]
- Zhang, D.-D.; Liu, J.-L.; Jiang, T.-M.; Li, L.; Fang, G.-Z.; Liu, Y.-P.; Chen, L.-J. Influence of Kluyveromyces marxianus on proteins, peptides, and amino acids in Lactobacillus-fermented milk. Food Sci. Biotechnol. 2017, 26, 739–748. [Google Scholar] [CrossRef]
- Maïworé, J.; Ngoune, L.T.; Piro-Metayer, I.; Montet, D. Identification of yeasts present in artisanal yoghurt and traditionally fermented milks consumed in the northern part of Cameroon. Sci. Afr. 2019, 6, e00159. [Google Scholar] [CrossRef]
- Lourens-Hattingh, A.; Viljoen, B.C. Survival of dairy-associated yeasts in yoghurt and yoghurt-related products. Food Microbiol. 2002, 19, 597–604. [Google Scholar] [CrossRef]
- Johansen, P.G.; Owusu-Kwarteng, J.; Parkouda, C.; Padonou, S.W.; Jespersen, L. Occurrence and Importance of Yeasts in Indigenous Fermented Food and Beverages Produced in Sub-Saharan Africa. Front. Microbiol. 2019, 10, 1789. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Li, K.; Shi, X.; Ni, Y.; Li, B.; Zhuge, B. Potential characterization of yeasts isolated from Kazak artisanal cheese to produce flavoring compounds. MicrobiologyOpen 2017, 7, e00533. [Google Scholar] [CrossRef] [Green Version]
- Gori, K.; Ryssel, M.; Arneborg, N.; Jespersen, L. Isolation and Identification of the Microbiota of Danish Farmhouse and Industrially Produced Surface-Ripened Cheeses. Microb. Ecol. 2012, 65, 602–615. [Google Scholar] [CrossRef] [Green Version]
- Kühle, A.V.D.A.; Jespersen, L. The Taxonomic Position of Saccharomyces boulardii as Evaluated by Sequence Analysis of the D1/D2 Domain of 26S rDNA, the ITS1-5.8S rDNA-ITS2 Region and the Mitochondrial Cytochrome-c Oxidase II Gene. Syst. Appl. Microbiol. 2003, 26, 564–571. [Google Scholar] [CrossRef]
- Jespersen, L.; Nielsen, D.S.; Hønholt, S.; Jakobsen, M. Occurrence and diversity of yeasts involved in fermentation of West African cocoa beans. FEMS Yeast Res. 2005, 5, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Renault, P.E.; Albertin, W.; Bely, M. An innovative tool reveals interaction mechanisms among yeast populations under oenological conditions. Appl. Microbiol. Biotechnol. 2013, 97, 4105–4119. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, C.; Johansen, P.G.; Petersen, M.A.; Poojary, M.M.; Lund, M.N.; Jespersen, L.; Arneborg, N. The utilisation of amino acids by Debaryomyces hansenii and Yamadazyma triangularis associated with cheese. Int. Dairy J. 2021, 121, 105135. [Google Scholar] [CrossRef]
- Johnsen, L.G.; Skou, P.B.; Khakimov, B.; Bro, R. Gas chromatography—Mass spectrometry data processing made easy. J. Chromatogr. A 2017, 1503, 57–64. [Google Scholar] [CrossRef]
- Xiao, J.; Chen, Y.; Li, J.; Shi, X.; Deng, L.; Wang, B. Evaluation of the Effect of Auxiliary Starter Yeasts With Enzyme Activities on Kazak Cheese Quality and Flavor. Front. Microbiol. 2020, 11, 3154. [Google Scholar] [CrossRef]
- Leite, A.; Leite, D.C.D.A.; Del Aguila, E.; Alvares, T.; Peixoto, R.; Miguel, M.; Silva, J.; Paschoalin, V. Microbiological and chemical characteristics of Brazilian kefir during fermentation and storage processes. J. Dairy Sci. 2013, 96, 4149–4159. [Google Scholar] [CrossRef]
- Zeppa, G.; Conterno, L.; Gerbi, V. Determination of Organic Acids, Sugars, Diacetyl, and Acetoin in Cheese by High-Performance Liquid Chromatography. J. Agric. Food Chem. 2001, 49, 2722–2726. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Methods for isolation, phenotypic characterization and maintenance of yeasts. In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 87–110. [Google Scholar] [CrossRef]
- Canganella, F.; Ovidi, M.; Paganini, S.; Vettraino, A.; Bevilacqua, L.; Trovatelli, L. Survival of undesirable micro-organisms in fruit yoghurts during storage at different temperatures. Food Microbiol. 1998, 15, 71–77. [Google Scholar] [CrossRef]
- Moineau-Jean, A.; Guévremont, E.; Champagne, C.P.; Roy, D.; Raymond, Y.; LaPointe, G. Fate of Escherichia coli and Kluyveromyces marxianus contaminants during storage of Greek-style yogurt produced by centrifugation or ultrafiltration. Int. Dairy J. 2017, 72, 36–43. [Google Scholar] [CrossRef]
- Belloch, C.; Querol, A.; Barrio, E. Yeast and Moulds|Kluyveromyces spp. In Encyclopedia Dairy Science, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 754–764. [Google Scholar] [CrossRef]
- Fasoli, G.; Tofalo, R.; Lanciotti, R.; Schirone, M.; Patrignani, F.; Perpetuini, G.; Grazia, L.; Corsetti, A.; Suzzi, G. Chromosome arrangement, differentiation of growth kinetics and volatile molecule profiles in Kluyveromyces marxianus strains from Italian cheeses. Int. J. Food Microbiol. 2015, 214, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Bosset, J.; Gauch, R. Comparison of the volatile flavour compounds of six european ‘AOC’ cheeses by using a new dynamic headspace GC-MS method. Int. Dairy J. 1993, 3, 359–377. [Google Scholar] [CrossRef]
- Mounchili, A.; Wichtel, J.; Bosset, J.; Dohoo, I.; Imhof, M.; Altieri, D.; Mallia, S.; Stryhn, H. HS-SPME gas chromatographic characterization of volatile compounds in milk tainted with off-flavour. Int. Dairy J. 2005, 15, 1203–1215. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Daran, J.-M.; van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich Pathway for Fusel Alcohol Production: A Century of Research on Saccharomyces cerevisiae Metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Wittmann, C.; Hans, M.; Bluemke, W. Metabolic physiology of aroma-producingKluyveromyces marxianus. Yeast 2002, 19, 1351–1363. [Google Scholar] [CrossRef]
- Cadwallader, K.R.; Singh, T.K. Flavours and off-flavours in milk and dairy products. In Advanced Dairy Chemistry, 3rd ed; McSweeney, P., Fox, P.F., Eds.; Springer: New York, NY, USA, 2009; pp. 631–690. [Google Scholar] [CrossRef]
- Güler, Z. Changes in salted yoghurt during storage. Int. J. Food Sci. Technol. 2007, 42, 235–245. [Google Scholar] [CrossRef]
- Chen, H.; Fink, G.R. Feedback control of morphogenesis in fungi by aromatic alcohols. Genes Dev. 2006, 20, 1150–1161. [Google Scholar] [CrossRef] [Green Version]
- Gori, K.; Knudsen, P.B.; Nielsen, K.F.; Arneborg, N.; Jespersen, L. Alcohol-based quorum sensing plays a role in adhesion and sliding motility of the yeast Debaryomyces hansenii. FEMS Yeast Res. 2011, 11, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Balbino, T.R.; da Silveira, F.A.; Ventorim, R.Z.; Nascimento, A.G.D.; de Oliveira, L.L.; da Silveira, W.B. Adaptive responses of Kluyveromyces marxianus CCT 7735 to 2-phenylethanol stress: Alterations in membrane fatty-acid composition, ergosterol content, exopolysaccharide production and reduction in reactive oxygen species. Fungal Genet. Biol. 2021, 151, 103561. [Google Scholar] [CrossRef]
- Kern, K.; Nunn, C.D.; Pichová, A.; Dickinson, J.R. Isoamyl alcohol-induced morphological change in Saccharomyces cerevisiae involves increases in mitochondria and cell wall chitin content. FEMS Yeast Res. 2004, 5, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zhou, W.; Yu, H.; Yuan, J.; Tian, H. Evaluation of the Perceptual Interactions among Aldehydes in a Cheddar Cheese Matrix According to Odor Threshold and Aroma Intensity. Molecules 2020, 25, 4308. [Google Scholar] [CrossRef]
- Hickey, C.; O’Sullivan, M.; Davis, J.; Scholz, D.; Kilcawley, K.; Wilkinson, M.; Sheehan, J. The effect of buttermilk or buttermilk powder addition on functionality, textural, sensory and volatile characteristics of Cheddar-style cheese. Food Res. Int. 2018, 103, 468–477. [Google Scholar] [CrossRef] [Green Version]
- McSweeney, P.L.H.; Sousa, M.J. Biochemical pathways for the production of flavour compounds in cheeses during ripening: A review. Lait 2000, 80, 293–324. [Google Scholar] [CrossRef]
- Carrillo-Carrión, C.; Cárdenas, S.; Valcárcel, M. Vanguard/rearguard strategy for the evaluation of the degradation of yoghurt samples based on the direct analysis of the volatiles profile through headspace-gas chromatography–mass spectrometry. J. Chromatogr. A 2007, 1141, 98–105. [Google Scholar] [CrossRef]
- Sfakianakis, P.; Tzia, C. Flavour profiling by gas chromatography–mass spectrometry and sensory analysis of yoghurt derived from ultrasonicated and homogenised milk. Int. Dairy J. 2017, 75, 120–128. [Google Scholar] [CrossRef]
- Hanková, M.; Čížková, H. Sensory active substances causing off-odour in liquid whey during storage. Potravinarstvo Slovak J. Food Sci. 2020, 14, 729–734. [Google Scholar] [CrossRef]
- Morrissey, J.P.; Etschmann, M.M.W.; Schrader, J.; de Billerbeck, G.M. Cell factory applications of the yeast Kluyveromyces marxianus for the biotechnological production of natural flavour and fragrance molecules. Yeast 2014, 32, 3–16. [Google Scholar] [CrossRef]
- Hoffmann, A.; Kupsch, C.; Walther, T.; Löser, C. Synthesis of ethyl acetate from glucose by Kluyveromyces marxianus, Cyberlindnera jadinii and Wickerhamomyces anomalus depending on the induction mode. Eng. Life Sci. 2020, 21, 154–168. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2009, 3, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Moineau-Jean, A.; Raymond, Y.; Sabik, H.; Graveline, N.; Champagne, C.P.; Roy, D.; LaPointe, G. Effect of manufacturing processes and storage on aroma compounds and sensory properties of yoghurt. Int. Dairy J. 2020, 105, 104662. [Google Scholar] [CrossRef]
- Padilla, B.; Manzanares, P.; Belloch, C. Yeast species and genetic heterogeneity within Debaryomyces hansenii along the ripening process of traditional ewes’ and goats’ cheeses. Food Microbiol. 2014, 38, 160–166. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Huang, L.; Xing, G.; Xu, X.; Tu, C.; Dong, M. Effect of Co-Fermentation with Lactic Acid Bacteria and K. marxianus on Physicochemical and Sensory Properties of Goat Milk. Foods 2020, 9, 299. [Google Scholar] [CrossRef] [Green Version]
- Leclercq-Perlat, M.-N.; Corrieu, G.; Spinnler, H.-E. Comparison of Volatile Compounds Produced in Model Cheese Medium Deacidified by Debaryomyces hansenii or Kluyveromyces marxianus. J. Dairy Sci. 2004, 87, 1545–1550. [Google Scholar] [CrossRef] [Green Version]
- Benozzi, E.; Romano, A.; Capozzi, V.; Makhoul, S.; Cappellin, L.; Khomenko, I.; Aprea, E.; Scampicchio, M.; Spano, G.; Märk, T.D.; et al. Monitoring of lactic fermentation driven by different starter cultures via direct injection mass spectrometric analysis of flavour-related volatile compounds. Food Res. Int. 2015, 76, 682–688. [Google Scholar] [CrossRef]
- Zhao, L.; Feng, R.; Ren, F.; Mao, X. Addition of buttermilk improves the flavor and volatile compound profiles of low-fat yogurt. LWT 2018, 98, 9–17. [Google Scholar] [CrossRef]
- Cárcoba, R.; Delgado, T.; Rodríguez, A. Comparative performance of a mixed strain starter in cow’s milk, ewe’s milk and mixtures of these milks. Eur. Food Res. Technol. 2000, 211, 141–146. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, ecology and industrial applications of aroma formation in yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef] [Green Version]
- Tamime, A.Y.; Robinson, R.K. Biochemistry of fermentation. In Tamime and Robinson’s Yoghurt, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 535–607. [Google Scholar] [CrossRef]
- Aunsbjerg, S.D.; Honoré, A.; Marcussen, J.; Ebrahimi, P.; Vogensen, F.; Benfeldt, C.; Skov, T.; Knøchel, S. Contribution of volatiles to the antifungal effect of Lactobacillus paracasei in defined medium and yogurt. Int. J. Food Microbiol. 2015, 194, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Mende, S.; Rohm, H.; Jaros, D. Influence of exopolysaccharides on the structure, texture, stability and sensory properties of yoghurt and related products. Int. Dairy J. 2016, 52, 57–71. [Google Scholar] [CrossRef]
- Tian, H.; Xu, X.; Sun, X.; Chen, C.; Yu, H. Evaluation of the perceptual interaction among key aroma compounds in milk fan by gas chromatography−olfactometry, odor threshold, and sensory analyses. J. Dairy Sci. 2020, 103, 5863–5873. [Google Scholar] [CrossRef] [PubMed]
- Serra, M.; Trujillo, A.J.; Guamis, B.; Ferragut, V. Flavour profiles and survival of starter cultures of yoghurt produced from high-pressure homogenized milk. Int. Dairy J. 2009, 19, 100–106. [Google Scholar] [CrossRef]
- Adhikari, K.; Grün, I.; Mustapha, A.; Fernando, L. Changes in the Profile of Organic Acids in Plain Set and Stirred Yogurts during Manufacture and Refrigerated Storage. J. Food Qual. 2002, 25, 435–451. [Google Scholar] [CrossRef]
- Nuryana, I.; Andriani, A.; Lisdiyanti, P. Yopi Analysis of organic acids produced by lactic acid bacteria. IOP Conf. Series: Earth Environ. Sci. 2019, 251, 012054. [Google Scholar] [CrossRef]
- Gänzle, M. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Lachance, M.A. Kluyveromyces van der Walt (1971). In The Yeasts, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2011; pp. 471–481. [Google Scholar] [CrossRef]
- Tofalo, R.; Fasoli, G.; Schirone, M.; Perpetuini, G.; Pepe, A.; Corsetti, A.; Suzzi, G. The predominance, biodiversity and biotechnological properties of Kluyveromyces marxianus in the production of Pecorino di Farindola cheese. Int. J. Food Microbiol. 2014, 187, 41–49. [Google Scholar] [CrossRef]
- Fernández-García, E.; McGregor, J.U. Determination of Organic Acids During the Fermentation and Cold Storage of Yogurt. J. Dairy Sci. 1994, 77, 2934–2939. [Google Scholar] [CrossRef]
- Vénica, C.I.; Perotti, M.C.; Bergamini, C.V. Organic acids profiles in lactose-hydrolyzed yogurt with different matrix composition. Dairy Sci. Technol. 2014, 94, 561–580. [Google Scholar] [CrossRef] [Green Version]
- La Torre, L.; Tamime, A.Y.; Muir, D.D. Rheology and sensory profiling of set-type fermented milks made with different commercial probiotic and yoghurt starter cultures. Int. J. Dairy Technol. 2003, 56, 163–170. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain ID | Identity, % | GeneBank Accession nr. | Origin |
|---|---|---|---|---|
| Kluyveromyces marxianus | Km1 | 100 | OM854769 | This study |
| Kluyveromyces marxianus | Km2 | 100 | OM854770 | This study |
| Kluyveromyces marxianus | Km3 | 99.8 | OM854771 | This study |
| Pichia kudriavzevii | Pk1 | 100 | OM854772 | Arla Foods, Denmark |
| Torulaspora delbrueckii | Td1 | 100 | OM854773 | Arla Foods, Denmark |
| Compound | RI Calculated | RI 1 Reference | VOCt0, µg·kg−1 3 | VOCt21, µg·kg−1 2 | Odour Threshold µg·kg−1 3 | Odour Description 4 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | K. marxianus Km1 | K. marxianus Km2 | K. marxianus Km3 | P. kudriavzevii Pk1 | T. delbrueckii Td1 | ||||||
| Alcohols | |||||||||||
| 1-Butanol | 1166 | 1166 | 37 ± 6.9 | 39 ± 1.4 A | 22 ± 1.9 B | 23 ± 1.9 B | 23 ± 5.1 B | 14 ± 4.3 B | 19 ± 2.7 B | 500-2500 | rancid, sweet |
| 2-Butanol | 1031 | 988–1053 | 2.1 ± 0.5 | 1.6 ± 0.05 B | 19 ± 3.2 A | 21 ± 0.62 A | 17 ± 3.4 A | 28 ± 7.9 A | 1.6 ± 0.17 B | 3300 | vinous, fruity |
| 1-Hexanol | 1355 | 1372 | 3.8 ± 1.0 | 4.5 ± 1.4 C | 56 ± 3.1 A | 46 ± 2.4 B | 48 ± 2.1 B | 4.8 ± 0.53 C | 5.9 ± 1.3 C | 2500 | sweet alcohol |
| 3-Methyl-1-butanol | 1227 | 1174–1255 | 21 ± 4.9 | 34 ± 13 C | 1780 ± 48 A | 1918 ± 100 A | 1781 ± 163 A | 412 ± 63 B | 30 ± 5 C | 3060 | whiskey, malt, burnt |
| 3-Methyl-3-buten-1-ol | 1262 | 1226–1277 | 10.1 ± 2.4 | 32 ± 1.8 | 26 ± 1.2 | 27 ± 1.6 | 26 ± 3.4 | 25 ± 2.4 | 28 ± 2.6 | 250 | sweet fruity |
| 2-Methyl-1-propanol | 1110 | 1110 | 0.61 ± 0.25 | 1.4 ± 0.62 B | 446 ± 58 A | 562 ± 17 A | 591 ± 102 A | 38 ± 8.9 AB | 0.90 ± 0.15 B | 40,000 | sweet musty, fusel |
| 1-Pentanol | 1267 | 1273 | 6.5 ± 1.6 | 8.1 ± 0.68 C | 30 ± 0.76 A | 24 ± 2.1 B | 23 ± 2.4 B | 9.2 ± 0.85 C | 9.9 ± 1.7 C | 120 | fusel |
| 1-Penten-3-ol | 1179 | 1177 | 0.28 ± 0.12 | 0.40 ± 0.15 B | 2.2 ± 0.15 A | 1.4 ± 0.19 AB | 1.9 ± 0.36 A | 0.99 ± 0.25 AB | 2.3 ± 0.83 A | 500 | pungent, fruity |
| 2-Phenylethanol | 1923 | 1930 | 0.36 ± 0.16 | 1.1 ± 1.04 BC | 3.2 ± 0.20 AB | 4.7 ± 0.41 A | 4.5 ± 0.80 A | 1.04 ± 0.71 BC | 0.36 ± 0.41 C | 750–1100 | mild rose |
| 1-Propanol | 1043 | 1041 | 0.35 ± 0.08 | 0.35 ± 0.05 B | 14 ± 3.4 A | 15 ± 2.3 A | 14 ± 2.02 A | 0.69 ± 0.01 B | 0.34 ± 0.06 B | 9000 | alcoholic |
| 2-Propanol | 936 | 884–963 | 1.1 ± 0.20 | 0.65 ± 0.41 B | 2.7 ± 0.81 AB | 3.3 ± 1.3 AB | 3.5 ± 0.53 AB | 6.9 ± 4.01 A | 0.68 ± 0.06 B | 40,000-78,000 | rubbing alcohol |
| 1-Octanol | 1559 | 1570 | 5.4 ± 1.7 | 5.8 ± 2.4 C | 15 ± 0.80 B | 20 ± 1.7 AB | 21 ± 1.4 A | 4.8 ± 0.44 C | 7.0 ± 1.9 C | 110–130 | aromatic, orange |
| Aldehydes | |||||||||||
| Butanal | 875 | 830–911 | 2.9 ± 0.92 | 0.82 ± 0.17 | 0.59 ± 0.13 | 0.45 ± 0.02 | 0.60 ± 0.15 | 0.33 ± 0.15 | 0.70 ± 0.17 | 7–37 | pungent |
| Heptanal | 1191 | 1189 | 12 ± 3.3 | 1.1 ± 0.14 B | 3.9 ± 1.1 A | 2.5 ± 0.22 AB | 2.3 ± 0.63 AB | 0.76 ± 0.10 B | 1.1 ± 0.34 B | 3–60 | fatty, pungent, fruity |
| Hexanal | 1079 | 1082 | 21 ± 5.7 | 1.3 ± 1.1 B | 8.6 ± 0.55 A | 4.2 ± 0.22 AB | 3.8 ± 0.79 B | 0.64 ± 0.07 B | 2.7 ± 1.9 B | 5–9 | sharp, fruity, grass |
| 2-Methylbutanal | 912 | 912 | 4.9 ± 1.1 | 3.3 ± 0.07 B | 10.5 ± 0.72 A | 11 ± 1.7 A | 14 ± 3.5 A | 0.35 ± 0.33 B | 3.6 ± 0.34 B | 1–6 | apple, malt |
| 3-Methylbutanal | 916 | 916 | 7.6 ± 1.9 | 2.9 ± 1.0 B | 9.3 ± 0.45 A | 10.2 ± 0.75 A | 9.8 ± 1.8 A | 12 ± 2.3 A | 4.03 ± 0.24 B | 1–3 | apple, malt |
| 3-Methyl-2-butenal | 1199 | 1189–1236 | 6.8 ± 1.5 | 2.5 ± 0.45 A | 0.46 ± 0.01 C | 0.66 ± 0.07 C | 0.67 ± 0.15 C | 2.1 ± 0.30 AB | 1.3 ± 0.42 BC | nr | metallic, herbaceous |
| 2-Methylpropanal | 811 | 812 | 7.1 ± 1.1 | 5.3 ± 0.53 D | 10.2 ± 1.6 BC | 12 ± 0.32 B | 19 ± 2.2 A | 5.4 ± 0.90 D | 6.2 ± 0.70 CD | 1.5 | sharp, pungent |
| Trans-2-Decenal | 1660 | 1662 | 4.9 ± 1.7 | 2.9 ± 2.3 AB | 5.3 ± 0.33 AB | 6.9 ± 0.55 A | 6.8 ± 1.3 A | 1.9 ± 0.28 B | 3.2 ± 1.3 AB | 0.4 | orange, tallow |
| Esters | |||||||||||
| Butyl acetate | 1070 | 1082 | 0.019 ± 0.036 | 0.51 ± 0.10 AB | 1.8 ± 0.71 AB | 2.8 ± 1.6 A | 0.92 ± 0.15 AB | 1.01 ± 0.60 AB | 0.094 ± 0.023 B | 10–100 | sweet, fruity |
| Ethyl acetate | 891 | 891 | nd | nd B | 289 ± 4.7 A | 283 ± 15 A | 291 ± 0.8 A | 278 ± 15 A | ndB | 5–5000 | pineapple, solvent |
| Ethyl butyrate | 1033 | 1036 | 0.51 ± 0.15 | 0.87 ± 0.23 D | 81 ± 4.2 A | 44 ± 2.2 B | 34 ± 2.3 C | 1.63 ± 0.48 D | 0.57 ± 0.06 D | 1 | pineapple |
| Ethyl hexanoate | 1244 | 1251 | 0.093 ± 0.039 | 0.072 ± 0.077 C | 23 ± 2.1 B | 32 ± 1.5 A | 19 ± 1.2 B | nd C | 0.18 ± 0.06 C | 1 | fruity, aniseed, apple |
| Ethyl octanoate | 1419 | 1445 | 0.55 ± 0.24 | 0.26 ± 0.37 C | 10.1 ± 1.2 B | 23 ± 5.7 A | 14 ± 2.1 B | nd C | 0.11 ± 0.15 C | 15 | fruity, sour apple |
| Ethyl formate | 823 | 823 | 0.27 ± 0.17 | 0.038 ± 0.054 B | 5.7 ± 2.9 A | 7.5 ± 0.36 A | 8.5 ± 1.5 A | 0.39 ± 0.08 B | 0.27 ± 0.04 B | 150,000 | fruity, pungent |
| Ethyl lactate | 1344 | 1356 | 0.20 ± 0.07 | 1.3 ± 0.89 C | 21 ± 1.3 A | 17 ± 1.7 AB | 14 ± 2.4 B | 0.58 ± 0.26 C | 0.82 ± 0.68 C | 14,000 | buttery, fruity |
| Ethyl propionate | 956 | 956 | nd | 0.076 ± 0.069 B | 6.2 ± 2.6 A | 7.6 ± 4.9 AB | 1.3 ± 0.15 B | 2.1 ± 1.1 AB | nd B | 10 | fruity, rum, pineapple |
| Hexyl acetate | 1281 | 1291 | 0.13 ± 0.04 | 0.26 ± 0.04 B | 1.3 ± 0.23 A | 1.7 ± 0.30 A | 1.2 ± 0.12 A | 0.21 ± 0.01 B | 0.24 ± 0.04 B | 2 | sweet, fruity, pear |
| 2-Methylpropyl acetate | 1013 | 1018 | 0.011 ± 0.007 | 1.78 ± 0.33 | 3.6 ± 1.3 | 5.1 ± 3.5 | 2.1 ± 0.12 | 0.87 ± 1.01 | 0.33 ± 0.14 | 66 | fruity, floral |
| 3-Methylbutyl acetate | 1136 | 1142 | 0.17 ± 0.091 | 0.30 ± 0.31 C | 33 ± 8.1 AB | 57 ± 21 A | 29 ± 0.49 AB | 3.8 ± 2.02 BC | 0.31 ± 0.31 C | 5 | sweet, fruity, banana |
| 4-Pentenyl acetate | 1199 | 1204 | 0.078 ± 0.036 | 0.08 ± 0.02 B | 2.08 ± 1.04 AB | 3.6 ± 2.4 A | 0.92 ± 0.36 AB | 0.13 ± 0.11 B | 0.01 ± 0.01 B | nr | green, plastic, weedy |
| Pentyl acetate | 1183 | 1185 | 0.22 ± 0.11 | 0.58 ± 0.18 AB | 0.85 ± 0.24 AB | 0.94 ± 0.31 A | 0.59 ± 0.01 AB | 0.36 ± 0.12 AB | 0.23 ± 0.04 B | 80 | banana |
| 2-Phenylethyl acetate | 1821 | 1835 | 0.074 ± 0.041 | 0.10 ± 0.13 C | 2.3 ± 0.44 B | 5.03 ± 0.28 A | 2.9 ± 0.47 B | 0.04 ± 0.05 C | 0.10 ± 0.07 C | 650 | sweet, floral, fruity |
| Ketones | |||||||||||
| 2,3-Butanedione | 984 | 985 | 178 ± 36 | 526 ± 11 A | 48 ± 4.3 C | 49 ± 10 C | 48 ± 5.4 C | 72 ± 9.3 C | 251 ± 47 B | 1.1–6.5 | chlorine, buttery |
| 2-Butanone | 905 | 906 | 191 ± 25 | 136 ± 5.6 A | 40 ± 7.9 B | 44 ± 2.9 B | 62 ± 11 B | 47 ± 11 B | 142 ± 10.5 A | 8400 | ethereal, fruity |
| 2-Heptanone | 1189 | 1189 | 60 ± 9 | 46 ± 4.2 B | 47 ± 1.4 B | 41 ± 1.9 B | 40.2 ± 2.7 B | 46 ± 0.22 B | 60.1 ± 2.6 A | 140–3000 | fruity, banana, spicy |
| 3-Hydroxy-2-butanone | 1290 | 1302 | 152 ± 28 | 201 ± 10 A | 55 ± 0.92 B | 56 ± 15 B | 79 ± 15 B | 128 ± 30.3 AB | 173 ± 48 A | 800 | buttery, woody, yogurt |
| 2-Nonanone | 1380 | 1398 | 31 ± 10 | 0.41 ± 0.58 B | 12 ± 0.80 AB | 20.5 ± 1.5 A | 18 ± 2.5 A | 2.5 ± 0.33 B | 24 ± 10.6 A | 5-200 | hot milk, soap, green |
| 2-Propanone | 815 | 815 | 109 ± 12 | 83 ± 8.2 A | 8.3 ± 2.9 B | 10.4 ± 1.4 B | 19 ± 1.5 B | 38 ± 28 B | 86 ± 8.7 A | 20,000 | fruity |
| Organic Acids | Initial Concentration, mg·100 g−1 | Yeast Species | Concentration, mg·100 g−1 1 | ||
|---|---|---|---|---|---|
| Day 7 | Day 14 | Day 21 | |||
| Acetic acid | 54 ± 1.4 | Control | 57 ± 2.9 | 57 ± 2.2 | 67 ± 4.2 A * |
| K. marxianus Km1 | 54 ± 3.2 | 52 ± 1.1 | 48 ± 2.4 C * | ||
| K. marxianus Km2 | 61 ± 4.7 | 56 ± 0.67 * | 50.4 ± 2.5 BC | ||
| K. marxianus Km3 | 56 ± 4.8 | 54 ± 1.09 | 58 ± 7.2 ABC | ||
| P. kudriavzevii Pk1 | 58 ± 2.4 | 54 ± 3.4 | 60.2 ± 2.9 AB | ||
| T. delbrueckii Td1 | 55 ± 6.3 | 55 ± 0.94 | 58 ± 4.1 ABC | ||
| Citric acid | 120 ± 9.1 | Control | 94 ± 3.4 A * | 98 ± 13 A | 96 ± 5.7 A * |
| K. marxianus Km1 | 86 ± 1.8 AB * | 74 ± 1.1 B * | 48 ± 8.1 B * | ||
| K. marxianus Km2 | 81 ± 6.9 AB * | 74 ± 1.6 B * | 80 ± 8.4 A * | ||
| K. marxianus Km3 | 82 ± 7.7 AB * | 74 ± 2.7 B * | 83 ± 7.9 A * | ||
| P. kudriavzevii Pk1 | 81 ± 7.3 AB * | 81 ± 7.3 AB * | 75 ± 14 A * | ||
| T. delbrueckii Td1 | 73 ± 3.9 B * | 71 ± 3.1 B * | 79 ± 0.64 A * | ||
| Lactic acid | 224 ± 8.5 | Control | 244 ± 13 A | 267 ± 1.4 A * | 287 ± 31 * |
| K. marxianus Km1 | 223 ± 4.8 AB | 233 ± 3.1 B | 257 ± 5.3 * | ||
| K. marxianus Km2 | 218 ± 5.2 B | 245 ± 18 AB | 261 ± 6.4 * | ||
| K. marxianus Km3 | 219 ± 7.9 B | 236 ± 0.57 B | 285 ± 19 * | ||
| P. kudriavzevii Pk1 | 223 ± 6.1 AB | 241 ± 9.7 B | 283 ± 12 * | ||
| T. delbrueckii Td1 | 221 ± 9.2 B | 231 ± 1.8 B | 271 ± 16 * | ||
| Orotic acid | 5.1 ± 0.16 | Control | 4.7 ± 0.38 | 4.6 ± 0.09 A * | 4.8 ± 0.57 A |
| K. marxianus Km1 | 4.3 ± 0.09 * | 4.1 ± 0.03 B * | 3.3 ± 0.14 B * | ||
| K. marxianus Km2 | 4.2 ± 0.17 * | 5.1 ± 0.29 A | 4.4 ± 0.17 A * | ||
| K. marxianus Km3 | 4.1 ± 0.19 * | 4.1 ± 0.08 B * | 4.8 ± 0.41 A | ||
| P. kudriavzevii Pk1 | 4.2 ± 0.17 * | 4.2 ± 0.23 B * | 5.01 ± 0.29 A | ||
| T. delbrueckii Td1 | 4.02 ± 0.39 * | 4.1 ± 0.06 B * | 4.6 ± 0.34 A | ||
| Uric acid | 3.7 ± 0.36 | Control | 3.1 ± 0.91 | 3.5 ± 0.20 A | 3.7 ± 0.73 |
| K. marxianus Km1 | 2.7 ± 0.38 | 2.5 ± 0.20 B * | 2.6 ± 0.19 * | ||
| K. marxianus Km2 | 3.1 ± 0.24 | 3.4 ± 0.20 A | 2.8 ± 0.58 | ||
| K. marxianus Km3 | 3.03 ± 0.29 | 3.2 ± 0.40 A | 3.2 ± 0.74 | ||
| P. kudriavzevii Pk1 | 3.02 ± 0.40 | 3.2 ± 0.18 A * | 3.2 ± 0.60 | ||
| T. delbrueckii Td1 | 2.9 ± 0.45 * | 3.01 ± 0.21 AB | 3.5 ± 0.40 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srimahaeak, T.; Petersen, M.A.; Lillevang, S.K.; Jespersen, L.; Larsen, N. Spoilage Potential of Contaminating Yeast Species Kluyveromyces marxianus, Pichia kudriavzevii and Torulaspora delbrueckii during Cold Storage of Skyr. Foods 2022, 11, 1776. https://doi.org/10.3390/foods11121776
Srimahaeak T, Petersen MA, Lillevang SK, Jespersen L, Larsen N. Spoilage Potential of Contaminating Yeast Species Kluyveromyces marxianus, Pichia kudriavzevii and Torulaspora delbrueckii during Cold Storage of Skyr. Foods. 2022; 11(12):1776. https://doi.org/10.3390/foods11121776
Chicago/Turabian StyleSrimahaeak, Thanyaporn, Mikael Agerlin Petersen, Søren K. Lillevang, Lene Jespersen, and Nadja Larsen. 2022. "Spoilage Potential of Contaminating Yeast Species Kluyveromyces marxianus, Pichia kudriavzevii and Torulaspora delbrueckii during Cold Storage of Skyr" Foods 11, no. 12: 1776. https://doi.org/10.3390/foods11121776
APA StyleSrimahaeak, T., Petersen, M. A., Lillevang, S. K., Jespersen, L., & Larsen, N. (2022). Spoilage Potential of Contaminating Yeast Species Kluyveromyces marxianus, Pichia kudriavzevii and Torulaspora delbrueckii during Cold Storage of Skyr. Foods, 11(12), 1776. https://doi.org/10.3390/foods11121776